
Virus-Induced Gene Silencing (VIGS) in Chinese Jujube
Abstract
:1. Introduction
2. Results
2.1. TRV Can Infect Jujube Seedlings
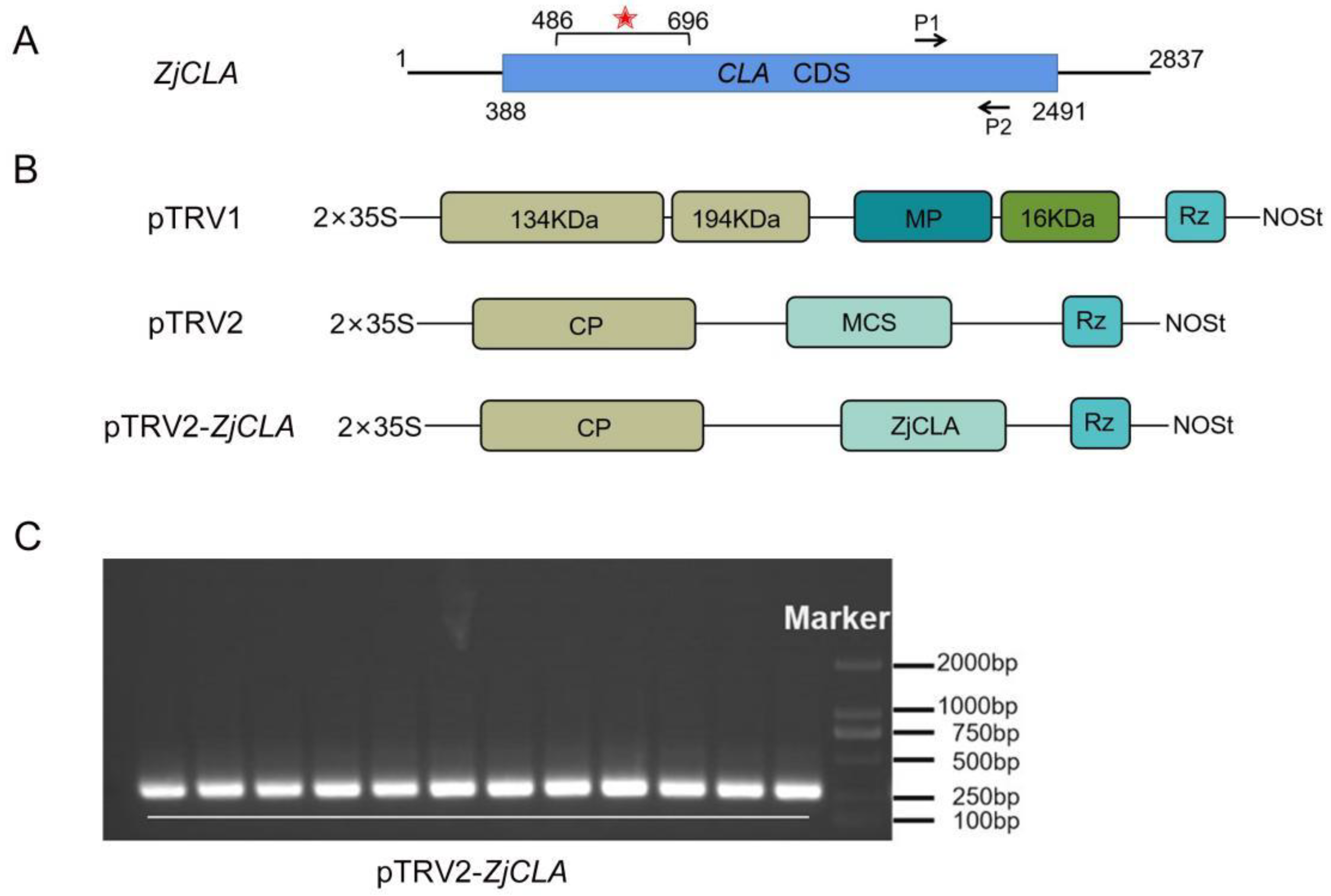
2.2. The Construction of pTRV2-ZjCLA Recombination
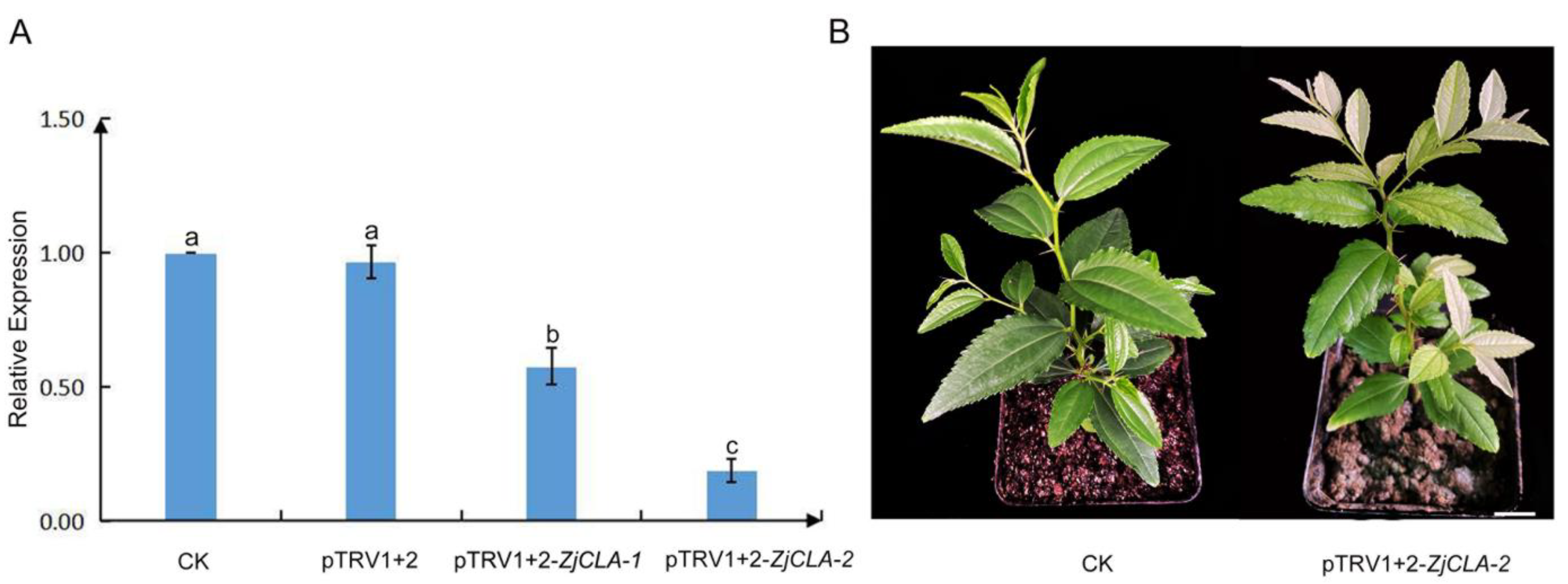
2.3. Silencing of ZjCLA in Chinese jujube
2.4. Optimization of the TRV-VIGS System in Jujube
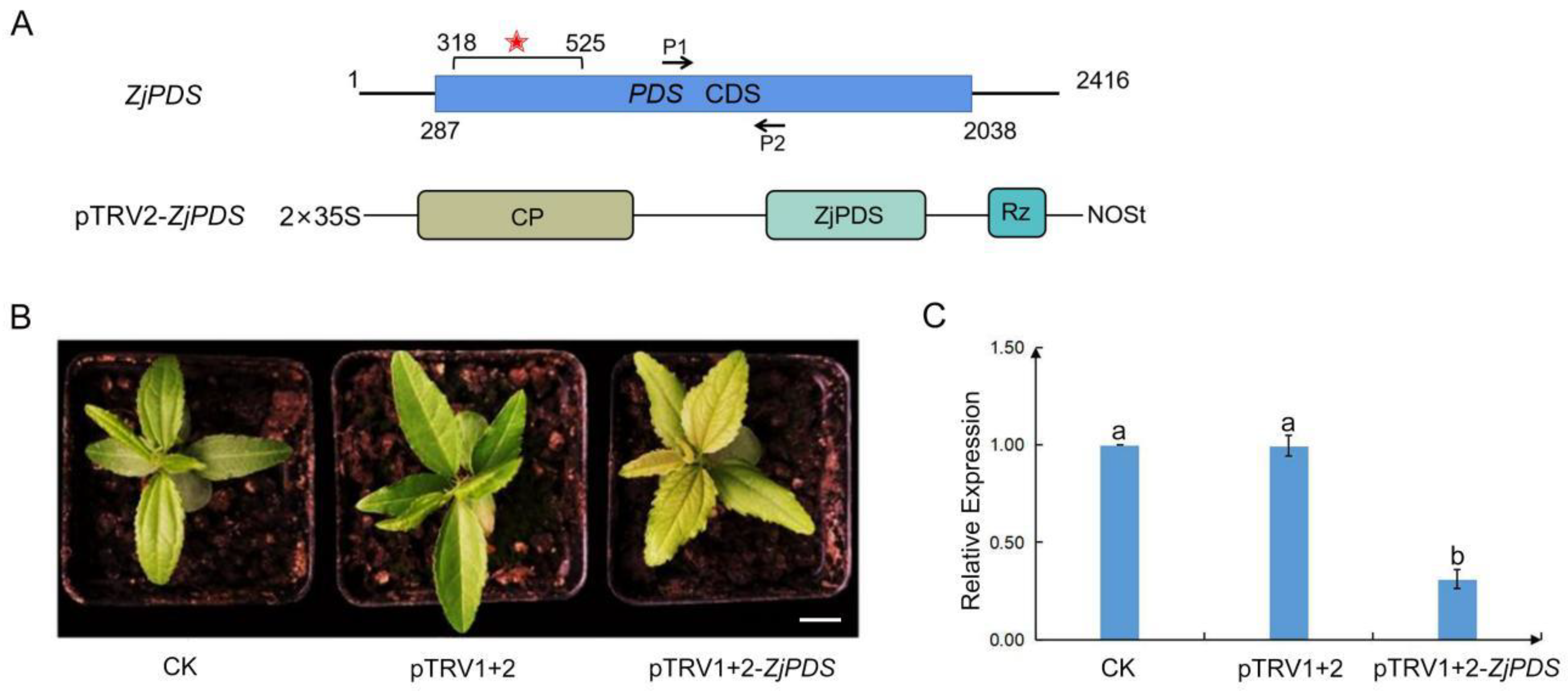
2.5. ZjPDS Silencing by Using the TRV-VIGS System in Jujube
3. Discussion
4. Materials and Methods
4.1. Cultivation of Jujube Seedlings
4.2. Identification and Cloning of ZjCLA and ZjPDS in Jujube
4.3. pTRV2-ZjCLA and pTRV2-ZjPDS Vector Construction
4.4. Preparation and Infection of Bacterial Solution
4.5. Infection of Jujube Seedlings
4.6. RNA Isolation and Gene Expression Analysis
4.7. Determinations of Chlorophyll Content
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rössner, C.; Lotz, D.; Becker, A. VIGS goes viral: How VIGS transforms our understanding of plant science. Annu. Rev. Plant Biol. 2022, 73, 703–728. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.Y.; Martinho, C.; Baulcombe, D.C. Post-transcriptional gene silencing using virus-induced gene silencing to study plant gametogenesis in tomato. Methods Mol. Biol. 2022, 2484, 201–212. [Google Scholar] [PubMed]
- Kammen, A.V. Virus-induced gene silencing in infected and transgenic plants. Trends Plant Sci. 1997, 2, 409–411. [Google Scholar] [CrossRef]
- Scheid, O.M.; Paszkowski, J. Transcriptional gene silencing mutants. Plant Mol. Biol. 2000, 43, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.A.; Dinesh Kumar, V.; Soma Sekhar Reddy, P.; Anil Kumar, C.; Sai Kumar, K.; Narasimha Rao, N.; Tarakeswari, M.; Sujatha, M. Post-transcriptional gene silencing: Basic concepts and applications. J. Biosci. 2020, 45, 128. [Google Scholar] [CrossRef]
- Suzuki, K.; Kelleher, A.D. Transcriptional regulation by promoter targeted RNAs. Curr. Top. Med. Chem. 2009, 9, 1079–1087. [Google Scholar] [CrossRef]
- Akbar, S.; Wei, Y.; Zhang, M.Q. RNA interference: Promising approach to combat plant viruses. Int. J. Mol. Sci. 2022, 23, 5312. [Google Scholar] [CrossRef]
- Felippes, F.F.; Wang, J.W.; Weigel, D. MIGS: miRNA-induced gene silencing. Plant J. 2012, 70, 541–547. [Google Scholar] [CrossRef]
- Tretiakova, P.; Voegele, R.T.; Soloviev, A.; Link, T.I. Successful silencing of the mycotoxin synthesis gene TRI5 in Fusarium culmorum and observation of reduced virulence in VIGS and SIGS experiments. Genes 2022, 13, 395. [Google Scholar] [CrossRef]
- Tiedge, K.; Destremps, J.; Solano-Sanchez, J.; Arce-Rodriguez, M.L.; Zerbe, P. Foxtail mosaic virus-induced gene silencing (VIGS) in switchgrass (Panicum virgatum L.). Plant Methods 2022, 18, 71. [Google Scholar] [CrossRef]
- Huang, C.; Qian, Y.; Li, Z.; Zhou, X. Virus-induced gene silencing and its application in plant functional genomics. Sci. China Life Sci. 2012, 55, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Creager, A.N.H. Tobacco mosaic virus and the history of molecular biology. Annu. Rev. Virol. 2022, 29, 39–55. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.Q.; Ding, N.Z. Evolution of Potato virus X. Mol. Phylogenetics Evol. 2022, 167, 107336. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, J.R.; Haigler, C.H.; Robertson, D. Method: Low-cost delivery of the cotton leaf crumple virus-induced gene silencing system. Plant Methods 2012, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Meziadi, C.; Blanchet, S.; Geffroy, V.; Pflieger, S. Virus-Induced gene silencing (VIGS) and foreign gene expression in Pisum sativum L. using the "one-step" bean pod mottle virus (BPMV) viral vector. Methods Mol. Biol. 2017, 1654, 311–319. [Google Scholar] [PubMed]
- Holzberg, S.; Brosio, P.; Gross, C.; Pogue, G.P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J. 2002, 30, 315–327. [Google Scholar] [CrossRef]
- Tan, Y.; Bukys, A.; Molnár, A.; Hudson, A. Rapid, high efficiency virus-mediated mutant complementation and gene silencing in Antirrhinum. Plant Methods 2020, 16, 145. [Google Scholar] [CrossRef]
- Mustafa, R.; Shafiq, M.; Mansoor, S.; Briddon, R.W.; Scheffler, B.E.; Scheffler, J.; Amin, I. Virus- induced gene silencing in cultivated cotton (Gossypium spp.) using tobacco rattle virus. Mol. Biotechnol. 2016, 58, 65–72. [Google Scholar] [CrossRef]
- Zaidi, S.S.; Mansoor, S. Viral vectors for plant genome engineering. Front. Plant Sci. 2017, 8, 539. [Google Scholar] [CrossRef]
- Jia, H.F.; Chai, Y.M.; Li, C.L.; Lu, D.; Luo, J.J.; Qin, L.; Shen, Y.Y. Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant Physiol. 2011, 157, 188–199. [Google Scholar] [CrossRef]
- Meng, L.H.; Wang, R.H.; Zhu, B.Z.; Zhu, H.L.; Luo, Y.B.; Fu, D.Q. Efficient virus-induced gene silencing in Solanum rostratum. PLoS ONE 2016, 11, e0156228. [Google Scholar] [CrossRef]
- Xu, Y.; Ji, X.; Xu, Z.; Yuan, Y.; Chen, X.; Kong, D.; Zhang, Y.; Sun, D. Transcriptome profiling reveals a petunia transcription factor, PhCOL4, contributing to antiviral RNA silencing. Front. Plant Sci. 2022, 13, 876428. [Google Scholar] [CrossRef]
- Liu, H.; Fu, D.; Zhu, B.; Yan, H.; Shen, X.; Zuo, J.; Zhu, Y.; Luo, Y. Virus-induced gene silencing in eggplant (Solanum melongena). J. Integr. Plant Biol. 2012, 54, 422–429. [Google Scholar] [CrossRef]
- Tian, J.; Pei, H.; Zhang, S.; Chen, J.; Chen, W.; Yang, R.; Meng, Y.; You, J.; Gao, J.; Ma, N. TRV-GFP: A modified Tobacco rattle virus vector for efficient and visualizable analysis of gene function. J. Exp. Bot. 2014, 65, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Liu, C.; Song, L.; Dong, Y.; Chen, L.; Li, M. A sweet cherry glutathione s-transferase gene, PavGST1, plays a central role in fruit skin coloration. Cells 2022, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-J.; Zhao, J.; Cai, Q.-L.; Liu, G.-C.; Wang, J.-R.; Zhao, Z.-H.; Liu, P.; Dai, L.; Yan, G.; Wang, W.-J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 5315. [Google Scholar] [CrossRef]
- Shen, L.-Y.; Luo, H.; Wang, X.-L.; Wang, X.-M.; Qiu, X.-J.; Liu, H.; Zhou, S.-S.; Jia, K.-H.; Nie, S.; Bao, Y.-T.; et al. Chromosome-scale genome assembly for Chinese sour jujube and insights into its genome evolution and domestication signature. Front. Plant Sci. 2021, 12, 773090. [Google Scholar] [CrossRef]
- Gu, X.F.; Meng, H.; Qi, G.; Zhang, J. Agrobacterium-mediated transformation of the winter jujube (Zizyphus jujuba Mill.). Plant Cell Tissue Organ Cult. 2008, 94, 23–32. [Google Scholar] [CrossRef]
- Rahman, J.; Baldwin, I.T.; Gase, K. California TRV-based VIGS vectors mediate gene silencing at elevated temperatures but with greater growth stunting. BMC Plant Biol. 2021, 21, 553. [Google Scholar] [CrossRef] [PubMed]
- Purkayastha, A.; Dasgupta, I. Virus-induced gene silencing: A versatile tool for discovery of gene functions in plants. Plant Physiol. Biochem. 2009, 47, 967–976. [Google Scholar] [CrossRef]
- Ryu, C.M.; Anand, A.; Kang, L.; Mysore, K.S. Agrodrench: A novel and effective agroinoculation method for virus-induced gene silencing in roots and diverse Solanaceous species. Plant J. 2004, 40, 322–331. [Google Scholar] [CrossRef]
- Schachtsiek, J.; Hussain, T.; Azzouhri, K.; Kayser, O.; Stehle, F. Virus-induced gene silencing (VIGS) in Cannabis sativa L. Plant Methods 2019, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Peng, J.Y.; Zeng, M.J.; Wu, L.X.; Fan, Y.X.; Zeng, L.H. Virus-induced gene silencing (VIGS) in Chinese narcissus and is use in functional analysis of NtMYB3. Hortic. Plant J. 2021, 7, 565–572. [Google Scholar] [CrossRef]
- Ma, Q.; Li, S.; Bi, C.; Hao, Z.; Sun, C.; Ye, N. Complete chloroplast genome sequence of a major economic species, Ziziphus jujuba (Rhamnaceae). Curr. Genet. 2017, 63, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhang, Z.; Li, S.; Lian, Q.; Fu, P.; He, Y.; Qiao, J.; Xu, K.; Liu, L.; Wu, M.; et al. Genomic analyses of diverse wild and cultivated accessions provide insights into the evolutionary history of jujube. Plant Biotechnol. J. 2021, 19, 517–531. [Google Scholar] [CrossRef]
- Fu, P.C.; Zhang, Y.Z.; Ya, H.Y.; Gao, Q.B. Characterization of SSR genomic abundance and identification of SSR markers for population genetics in Chinese jujube (Ziziphus jujuba Mill.). PeerJ 2016, 4, 1735. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Yellina, A.L.; Orashakova, S.; Becker, A. Virus-induced gene silencing (VIGS) in plants: An overview of target species and the virus-derived vector systems. Methods Mol. Biol. 2013, 975, 1–14. [Google Scholar]
- Jiang, C.Z.; Chen, J.C.; Reid, M. Virus-induced gene silencing in ornamental plants. Methods Mol. Biol. 2011, 744, 81–96. [Google Scholar]
- Zhang, J.; Yu, D.; Zhang, Y.; Liu, K.; Xu, K.; Zhang, F.; Wang, J.; Tan, G.; Nie, X.; Ji, Q.; et al. Vacuum and co-cultivation Agroinfiltration of (Germinated) seeds results in Tobacco Rattle Virus (TRV) mediated whole-plant virus-induced gene silencing (VIGS) in wheat and maize. Front. Plant Sci. 2017, 8, 393. [Google Scholar] [CrossRef]
- Jiao, Y.; Ma, R.J.; Shen, Z.J.; Yan, J.; Yu, M.L. Gene regulation of anthocyanin biosynthesis in two blood-flesh peach (Prunus persica (L.) Batsch) cultivars during fruit development. J. Zhejiang Univ. Sci. B 2014, 15, 809–819. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, L.X.; Xu, C. The effect of silencing chalcone synthase on anthocyanin metabolism in peach. Acta Hortic. Sin. 2015, 42, 31–37. [Google Scholar]
- Li, Z.X.; Lan, J.B.; Liu, Y.Q.; Qi, L.W.; Tang, J.M. Investigation of the role of AcTPR2 in kiwifruit and its response to Botrytis cinerea infection. BMC Plant Biol. 2020, 20, 557. [Google Scholar] [CrossRef]
- Becker, A.; Lange, M. VIGS-genomics goes functional. Trends Plant Sci. 2010, 15, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pozo, N.; Rosli, H.G.; Martin, G.B.; Mueller, L.A. The SGN VIGS tool: User-friendly software to design virus-induced gene silencing (VIGS) constructs for functional genomics. Mol. Plant. 2015, 8, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Zhao, J.; Liu, M. Expression stabilities of candidate reference genes for RT-qPCR in Chinese jujube (Ziziphus jujuba Mill.) under a variety of conditions. PLoS ONE 2016, 11, e0154212. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Usage | Primer Sequence |
|---|---|
| Partial gene cloning of ZjCLA | F: GAGCAGATCTATTGGGCTTAGC; R: ATGACATCAGACCTTAGTTCATCC |
| Vector construction of pTRV2-ZjCLA | F: TGTGAGTAAGGTTACCGAATTCGAGCAGATCTATTGGGCTTAGC R: GGGACATGCCCGGGCCTCGAGATGACATCAGACCTTAGTTCATCC |
| qRT-PCR of ZjCLA | F: ATCAATTGGAGGCTTTGGAT; R: ACTGTTGCTGCAATATGAGA |
| Partial gene cloning of ZjPDS | F: ACTTGAGCTGGCAAACTA; R: CACACAAACTACCTTCAAA |
| Vector construction of pTRV2-ZjPDS | F: AGAAGGCCTCCATGGGGATCCACTTGAGCTGGCAAACTA R: GGGACATGCCCGGGCCTCGAGCACACAAACTACCTTCAAA |
| qRT-PCR of ZjPDS | F: AGGAGAGTTCAGTCGATTTG; R: TGACATGGCAATAAACACCT |
| TRV virus testing of pTRV1 | F: GCTCTTGGGAACTACATGGTG; R: CGCCTCAATCGTCTTCATCTCC |
| TRV virus testing of pTRV2 | F: GGTACGTAGTAGAGTCCCAC; R: ACAAAAGACTTACCGATCAATC |
| qRT-PCR of ZjACT | F: TTGCTTCTCACCCTTGATGC; R: AGCCTTCCTGCCAACGAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Niu, N.; Li, S.; Liu, Y.; Xue, C.; Wang, H.; Liu, M.; Zhao, J. Virus-Induced Gene Silencing (VIGS) in Chinese Jujube. Plants 2023, 12, 2115. https://doi.org/10.3390/plants12112115
Zhang Y, Niu N, Li S, Liu Y, Xue C, Wang H, Liu M, Zhao J. Virus-Induced Gene Silencing (VIGS) in Chinese Jujube. Plants. 2023; 12(11):2115. https://doi.org/10.3390/plants12112115
Chicago/Turabian StyleZhang, Yao, Nazi Niu, Shijia Li, Yin Liu, Chaoling Xue, Huibin Wang, Mengjun Liu, and Jin Zhao. 2023. "Virus-Induced Gene Silencing (VIGS) in Chinese Jujube" Plants 12, no. 11: 2115. https://doi.org/10.3390/plants12112115