
The Key Metabolic Network and Genes Regulating the Fresh Fruit Texture of Jujube (Ziziphus jujuba Mill.) Revealed via Metabolomic and Transcriptomic Analysis
 , ,
, ,
Abstract
:1. Introduction
2. Results
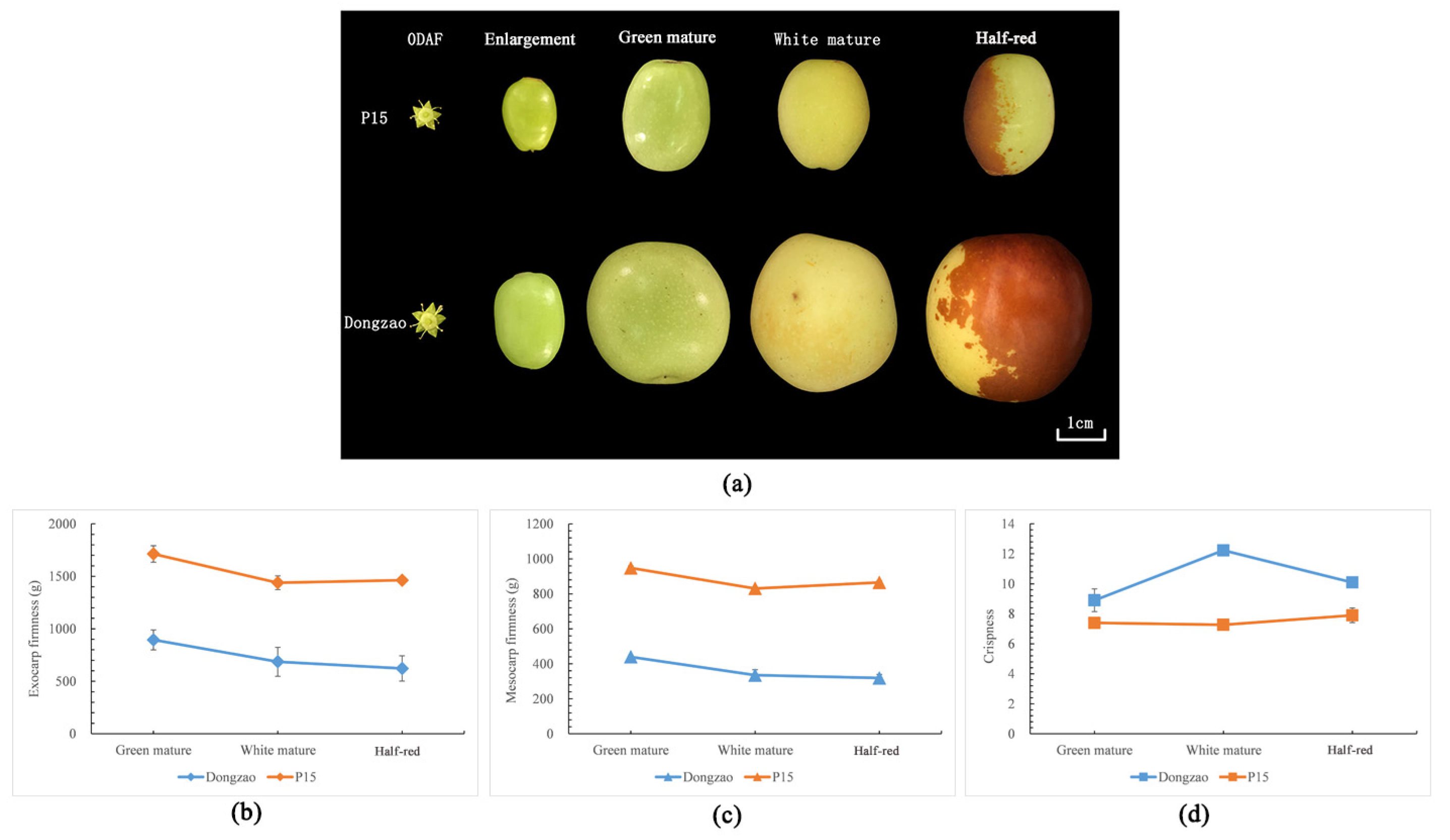
2.1. Texture Difference between ‘Dongzao’ and ‘P15’
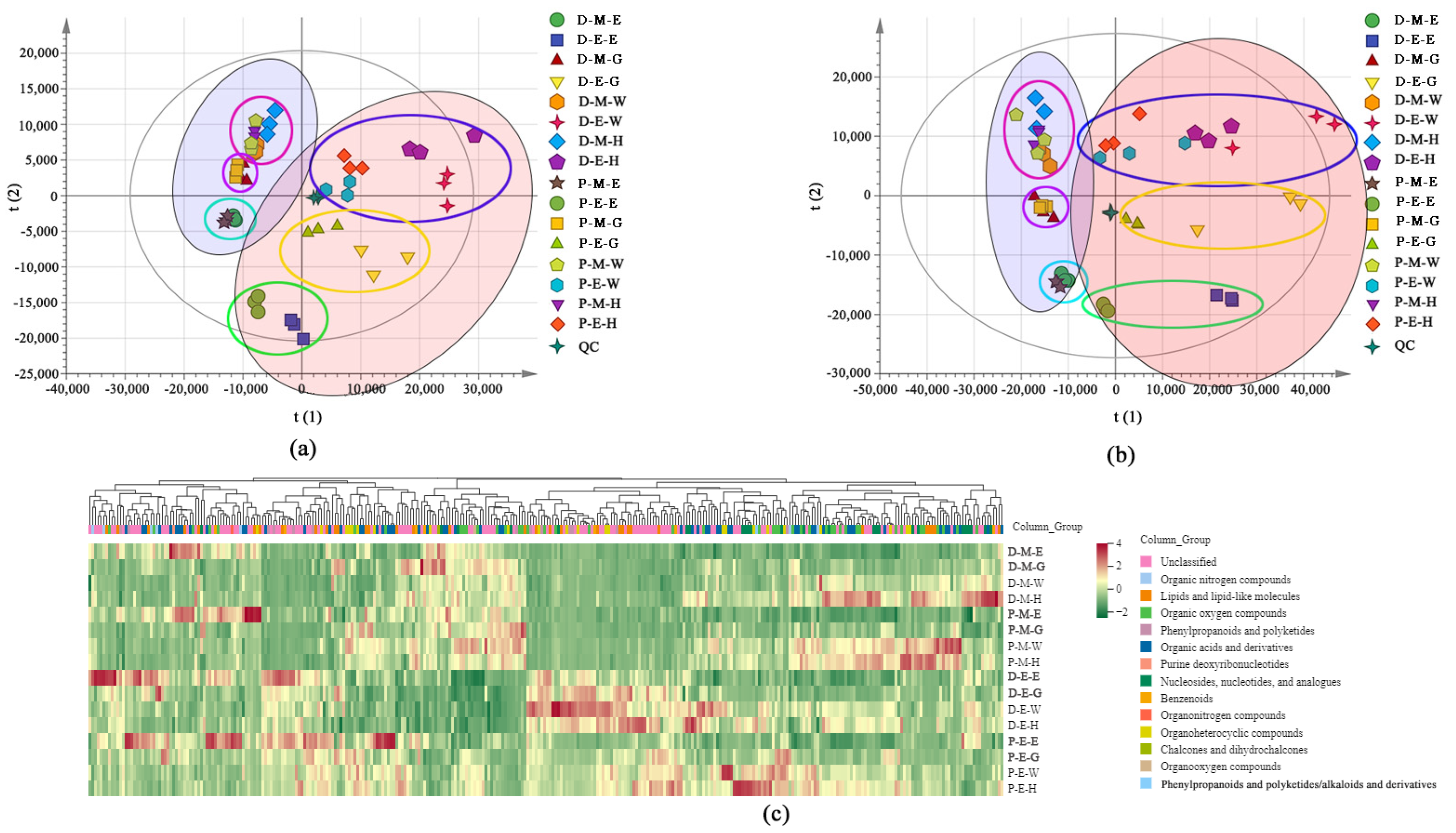
2.2. Metabolomic Profiles
2.3. DAMs Profiling in Exocarp and Mesocarp of Two Varieties at Different Development Stages
2.4. Transcriptomic Analysis in Exocarp and Mesocarp of Two Varieties at Different Development Stages
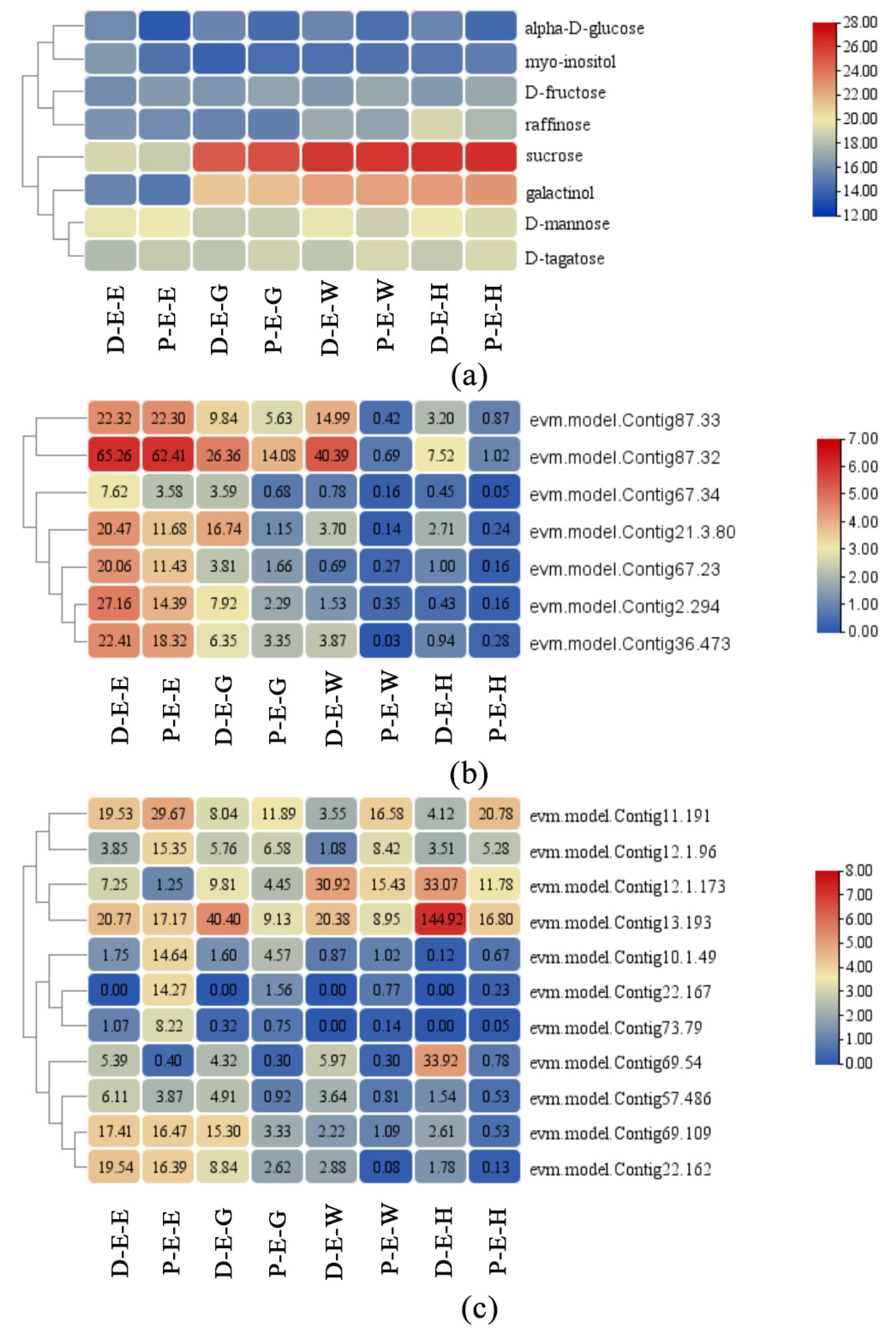
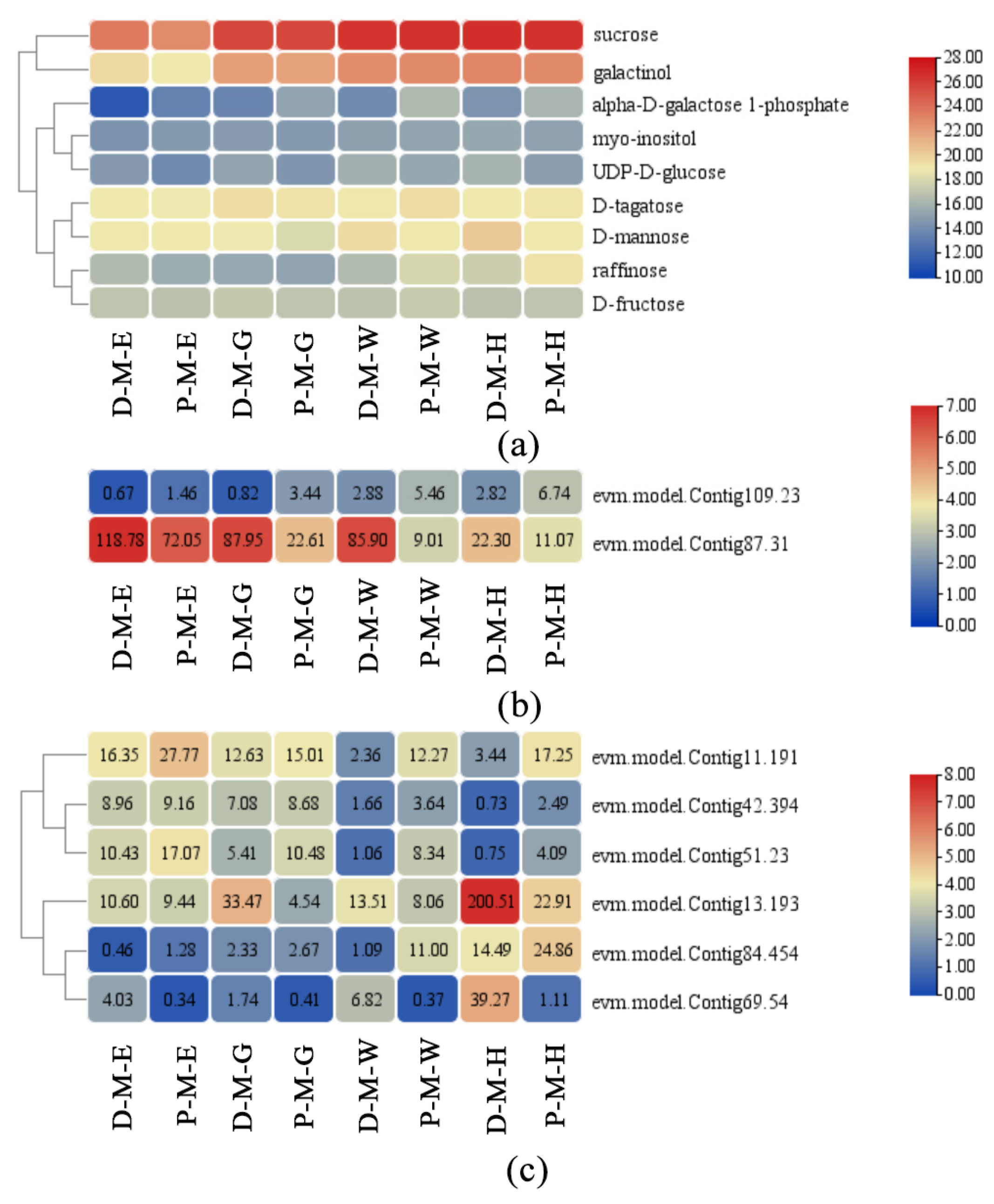
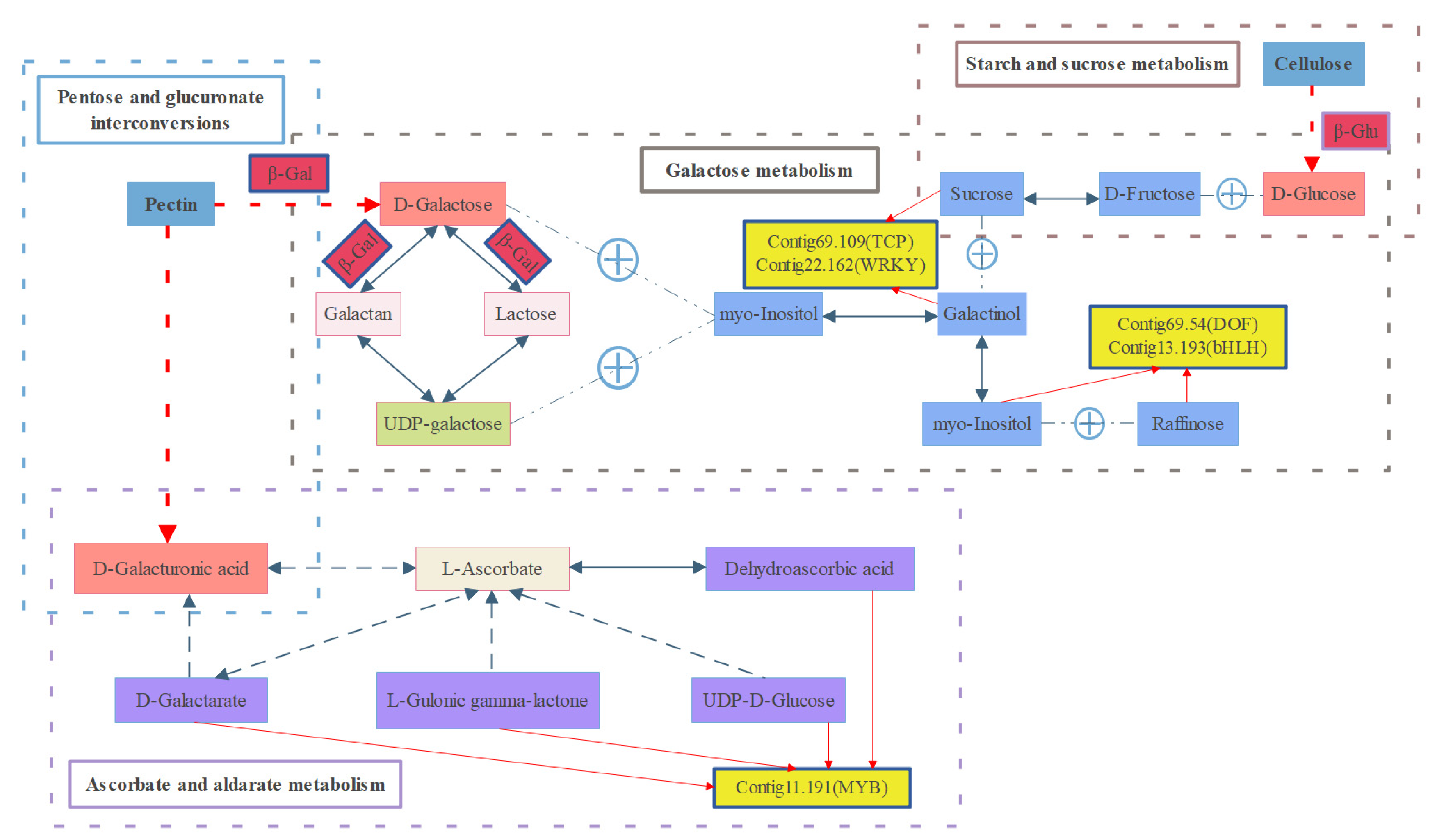
2.5. ‘Galactose Metabolism’ Is the Most Closely Related Pathway between the Two-Omics Data
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Fruit Texture Detection
4.3. Metabolite Profiling Using LC-MS/MS and Data Analysis
4.4. Transcriptome Sequencing and Data Analysis
4.5. Combined Metabolomic and Transcriptomic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Ma, H.; Jung, S.; Main, D.; Guo, L. Developmental Mechanisms of Fleshy Fruit Diversity in Rosaceae. Annu. Rev. Plant Biol. 2020, 71, 547–573. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.H.; Wu, C.S.; Wang, M. The jujube (Ziziphus jujuba Mill.) fruit: A review of current knowledge of fruit composition and health benefits. J. Agric. Food Chem. 2013, 61, 3351–3363. [Google Scholar] [CrossRef]
- Péneau, S.; Hoehn, E.; Roth, H.; Escher, F.; Nuessli, J. Importance and consumer perception of freshness of apples. Food Qual. Prefer. 2006, 17, 19. [Google Scholar] [CrossRef]
- Bianchi, T.; Guerrero, L.; Gratacós-Cubarsí, M.; Claret, A.; Argyris, J.; Garcia-Mas, J.; Hortós, M. Textural properties of different melon (Cucumis melo L.) fruit types: Sensory and physical-chemical evaluation. Sci. Hortic. 2016, 201, 46–56. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, G.; Liang, L. Establishment of the Detecting Method on the Fruit Texture of Dongzao by Puncture Test. Sci. Agric. Sin. 2011, 44, 1210–1217. [Google Scholar]
- Zhao, A.; Xu, X.; Wang, Y.; Ren, H.; Gong, G.; Jiao, J.; Sui, C.; Li, D. Measuring texture quality of fresh jujube fruit using texture analyser. J. Fruit Sci. 2018, 35, 631–641. [Google Scholar]
- Kong, X.; Xu, M.; Wan, H.; Han, L.; Liu, X.; Li, Q.; Hao, B.; Zhang, S.; Li, X.; Liu, Y.; et al. Development of a texture evaluation system for winter jujube (Ziziphus jujuba ‘Dongzao’). J. Integr. Agric. 2022, 21, 3658–3668. [Google Scholar] [CrossRef]
- Matas, A.J.; Gapper, N.E.; Chung, M.Y.; Giovannoni, J.J.; Rose, J.K. Biology and genetic engineering of fruit maturation for enhanced quality and shelf-life. Curr. Opin. Biotechnol. 2009, 20, 197–203. [Google Scholar] [CrossRef]
- Albenne, C.; Canut, H.; Jamet, E. Plant cell wall proteomics: The leadership of Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 111. [Google Scholar] [CrossRef]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jorgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic cross-links: Building and de-constructing the plant cell wall. Nat. Prod. Rep. 2020, 37, 919–961. [Google Scholar] [CrossRef]
- Goulao, L.F.; Oliveira, C.M. Cell wall modifications during fruit ripening: When a fruit is not the fruit. Trends Food Sci. Technol. 2008, 19, 25. [Google Scholar] [CrossRef]
- Cybulska, J.; Zdunek, A.; Psonka-Antonczyk, K.M.; Stokke, B.R.T. The relation of apple texture with cell wall nanostructure studied using an atomic force microscope. Carbohydr. Polym. 2013, 92, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Candelas, P.; Rosario, B.P.; Marta, B.; García-Gago, J.A.; Waldron, K.W.; Quesada, M.A.; Juan, M.; Mercado, J.A. Antisense down-regulation of the strawberry β-galactosidase gene FaβGal4 increases cell wall galactose levels and reduces fruit softening. J. Exp. Bot. 2015, 67, 619–631. [Google Scholar] [CrossRef]
- Xu, Z.; Dai, J.; Kang, T.; Shah, K.; Li, Q.; Liu, K.; Xing, L.; Ma, J.; Zhang, D.; Zhao, C. PpePL1 and PpePL15 Are the Core Members of the Pectate Lyase Gene Family Involved in Peach Fruit Ripening and Softening. Front. Plant Sci. 2022, 13, 844055. [Google Scholar] [CrossRef]
- Wen, W.; Li, D.; Li, X.; Gao, Y.; Li, W.; Li, H.; Liu, J.; Liu, H.; Chen, W.; Luo, J.; et al. Metabolome-based genome-wide association study of maize kernel leads to novel biochemical insights. Nat. Commun. 2014, 5, 3438. [Google Scholar] [CrossRef]
- Noor, E.; Cherkaoui, S.; Sauer, U. Biological insights through omics data integration. Curr. Opin. Syst. Biol. 2019, 15, 39–47. [Google Scholar] [CrossRef]
- Shen, B.; Zhang, Z.; Shi, Q.; Du, J.; Xue, Q.; Li, X. Active compound analysis of Ziziphus jujuba cv. Jinsixiaozao in different developmental stages using metabolomic and transcriptomic approaches. Plant Physiol. Biochem. 2022, 189, 14–23. [Google Scholar] [CrossRef]
- Shi, Q.; Du, J.; Zhu, D.; Li, X.; Li, X. Metabolomic and Transcriptomic Analyses of Anthocyanin Biosynthesis Mechanisms in the Color Mutant Ziziphus jujuba cv. Tailihong. J. Agric. Food Chem. 2020, 68, 15186–15198. [Google Scholar] [CrossRef]
- Wu, C.S.; Gao, Q.H.; Guo, X.D.; Yu, J.G.; Min, W. Effect of ripening stage on physicochemical properties and antioxidant profiles of a promising table fruit ‘pear-jujube’ (Zizyphus jujuba Mill.). Sci. Hortic. 2012, 148, 177–184. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.; Maiwulanjiang, M.; Zhang, W.L.; Zhan, J.Y.; Lam, C.T.; Zhu, K.Y.; Yao, P.; Choi, R.C.; Lau, D.T.; et al. Chemical and biological assessment of Ziziphus jujuba fruits from China: Different geographical sources and developmental stages. J. Agric. Food Chem. 2013, 61, 7315–7324. [Google Scholar] [CrossRef]
- Seymour, G.B.; Manning, K.; Eriksson, E.M.; Popovich, A.H.; King, G.J. Genetic identification and genomic organization of factors affecting fruit texture. J. Exp. Bot. 2002, 53, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Mercado, J.A.; Matas, A.J.; Posé, S. Fruit and Vegetable Texture: Role of Their Cell Walls. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–7. [Google Scholar]
- Yang, L.; Cong, P.; He, J.; Bu, H.; Qin, S.; Lyu, D. Differential pulp cell wall structures lead to diverse fruit textures in apple (Malus domestica). Protoplasma 2022, 259, 1205–1217. [Google Scholar] [CrossRef]
- Balic, I.; Olmedo, P.; Zepeda, B.; Rojas, B.; Ejsmentewicz, T.; Barros, M.; Aguayo, D.; Moreno, A.A.; Pedreschi, R.; Meneses, C.; et al. Metabolomic and biochemical analysis of mesocarp tissues from table grape berries with contrasting firmness reveals cell wall modifications associated to harvest and cold storage. Food Chem. 2022, 389, 133052. [Google Scholar] [CrossRef] [PubMed]
- Belge, B.; Comabella, E.; Graell, J.; Lara, I. Post-storage cell wall metabolism in two sweet cherry (Prunus avium L.) cultivars displaying different postharvest performance. Food Sci. Technol. Int. 2015, 21, 416–427. [Google Scholar] [CrossRef]
- Smith, D.L.; Gross, K.C. A Family of at Least Seven β-Galactosidase Genes Is Expressed during Tomato Fruit Development. Plant Physiol. 2000, 123, 1173–1183. [Google Scholar] [CrossRef]
- Yang, H.; Liu, J.; Dang, M.; Zhang, B.; Li, H.; Meng, R.; Qu, D.; Yang, Y.; Zhao, Z. Analysis of beta-Galactosidase During Fruit Development and Ripening in Two Different Texture Types of Apple Cultivars. Front. Plant Sci. 2018, 9, 539. [Google Scholar] [CrossRef]
- Celia, M.R.M.; John, O.; Seymour, G.B. Pectate lyases, cell wall degradation and fruit softening. J. Exp. Bot. 2002, 377, 2115–2119. [Google Scholar] [CrossRef]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.; Sheldon, J.; Stiegelmeyer, S.; Perez, L.; et al. Genetic improvement of tomato by targeted control of fruit softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef]
- Wang, D.; Samsulrizal, N.; Yan, C.; Allcock, N.S.; Seymour, G.B. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant. Physiol. 2018, 179, 1187–2018. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Pandey, A.; Ganansounou, E. Genetic modification: A tool for enhancing beta-glucosidase production for biofuel application. Bioresour. Technol. 2017, 245, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Daras, G.; Templalexis, D.; Avgeri, F.; Tsitsekian, D.; Karamanou, K.; Rigas, S. Updating Insights into the Catalytic Domain Properties of Plant Cellulose synthase (CesA) and Cellulose synthase-like (Csl) Proteins. Molecules 2021, 26, 4335. [Google Scholar] [CrossRef] [PubMed]
- Song, C.B.; Shan, W.; Yang, Y.Y.; Tan, X.L.; Fan, Z.Q.; Chen, J.Y.; Lu, W.J.; Kuang, J.F. Heterodimerization of MaTCP proteins modulates the transcription of MaXTH10/11 genes during banana fruit ripening. Biochim. Biophys. Acta-Gene Regul. Mech. 2018, 1861, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Negi, J.; Moriwaki, K.; Konishi, M.; Yokoyama, R.; Nakano, T.; Kusumi, K.; Hashimoto-Sugimoto, M.; Schroeder, J.I.; Nishitani, K.; Yanagisawa, S.; et al. A Dof transcription factor, SCAP1, is essential for the development of functional stomata in Arabidopsis. Curr. Biol. 2013, 23, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.H.; Han, Y.C.; Xiao, Y.Y.; Kuang, J.F.; Fan, Z.Q.; Chen, J.Y.; Lu, W.J. The banana fruit Dof transcription factor MaDof23 acts as a repressor and interacts with MaERF9 in regulating ripening-related genes. J. Exp. Bot. 2016, 67, 2263–2275. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Qiao, H.; Ma, M.; Wang, X.; Tian, Y.; Bai, S.; Hasi, A. Genome-Wide Identification and Characterization of Melon bHLH Transcription Factors in Regulation of Fruit Development. Plants 2021, 10, 2721. [Google Scholar] [CrossRef]
- Huang, J.; Chen, X.; He, A.; Ma, Z.; Gong, T.; Xu, K.; Chen, R. Integrative Morphological, Physiological, Proteomics Analyses of Jujube Fruit Development Provide Insights Into Fruit Quality Domestication from Wild Jujube to Cultivated Jujube. Front. Plant Sci. 2021, 12, 773825. [Google Scholar] [CrossRef]
- Webb, K.; Bristow, T.; Sargent, M.; Stein, B. Methodology for Accurate Mass Measurement of Small Molecules; LGC Limited: Teddington, UK, 2004; ISBN 0-948926-22-8. [Google Scholar]
- Gao, Y.; Li, J.T.; Li, X.; Li, X.; Yang, S.W.; Chen, N.H.; Li, L.; Zhang, L. Tetrahydroxy stilbene glycoside attenuates acetaminophen-induced hepatotoxicity by UHPLC-Q-TOF/MS-based metabolomics and multivariate data analysis. J. Cell. Physiol. 2020, 236, 3832–3862. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, C.; Zhao, X.; Fei, Z.; Wan, K.; Zhang, Z.; Pang, X.; Yin, X.; Bai, Y.; Sun, X.; et al. The Jujube Genome Provides Insights into Genome Evolution and the Domestication of Sweetness/Acidity Taste in Fruit Trees. PLoS Genet. 2016, 12, e1006433. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages | ID | Names |
|---|---|---|
| Enlargement stage | map00073 | Cutin, suberine and wax biosynthesis |
| map00410 | beta-alanine metabolism | |
| Green mature stage | map00052 | Galactose metabolism |
| map00630 | Glyoxylate and dicarboxylate metabolism | |
| map02010 | ABC transporters | |
| map00350 | Tyrosine metabolism | |
| White mature stage | map00053 | Ascorbate and aldarate metabolism |
| map00052 | Galactose metabolism | |
| map00051 | Fructose and mannose metabolism | |
| map00908 | Zeatin biosynthesis | |
| Half-red stage | map00052 | Galactose metabolism |
| map02010 | ABC transporters | |
| map00073 | Cutin, suberine and wax biosynthesis | |
| map00561 | Glycerolipid metabolism |
| DEG | Annotation | DAM | Pathway | Pearson’s r |
|---|---|---|---|---|
| evm.model.Contig21.3.80 | β-Gal | D-Fructose | Fructose and mannose metabolism | −0.80 |
| Galactose metabolism | ||||
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.81 | ||
| evm.model.Contig67.34 | β-Gal | Sucrose | Galactose metabolism | −0.80 |
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.84 | ||
| evm.model.Contig67.23 | β-Gal | Galactinol | Galactose metabolism | −0.84 |
| evm.model.Contig87.33 | β-Glu | Galactinol | Galactose metabolism | −0.81 |
| D-Fructose | Fructose and mannose metabolism | −0.82 | ||
| Galactose metabolism | ||||
| Starch and sucrose metabolism | ||||
| evm.model.Contig87.32 | β-Glu | Galactinol | Galactose metabolism | −0.81 |
| D-Fructose | Fructose and mannose metabolism | −0.81 | ||
| Galactose metabolism | ||||
| Starch and sucrose metabolism | ||||
| evm.model.Contig36.473 | β-Glu | Sucrose | Galactose metabolism | −0.81 |
| Starch and sucrose metabolism | ||||
| evm.model.Contig2.294 | PL | Sucrose | Galactose metabolism | −0.85 |
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.85 | ||
| evm.model.Contig10.1.49 | TCP | D-galacturonic acid | Pentose and glucuronate interconversions | 0.95 |
| Ascorbate and aldarate metabolism | ||||
| evm.model.Contig69.109 | TCP | Sucrose | Galactose metabolism | −0.88 |
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.91 | ||
| evm.model.Contig22.162 | WRKY | Sucrose | Galactose metabolism | −0.87 |
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.92 | ||
| evm.model.Contig22.167 | WRKY | D-galacturonic acid | Pentose and glucuronate interconversions | 0.99 |
| Ascorbate and aldarate metabolism | ||||
| evm.model.Contig11.191 | MYB | L-Gulonic gamma-lactone | Ascorbate and aldarate metabolism | 0.91 |
| D-Galactarate | Ascorbate and aldarate metabolism | 0.94 | ||
| Dehydroascorbic acid | Ascorbate and aldarate metabolism | 0.94 | ||
| evm.model.Contig12.1.173 | MYB | D-Mannose 1-phosphate | Fructose and mannose metabolism | 0.81 |
| Raffinose | Galactose metabolism | 0.82 | ||
| evm.model.Contig57.486 | AP2/ERF | Alpha-D-Glucose | Fructose and mannose metabolism | −0.86 |
| Galactose metabolism | ||||
| evm.model.Contig73.79 | AP2/B3 | D-galacturonic acid | Pentose and glucuronate interconversions | 0.99 |
| Ascorbate and aldarate metabolism | ||||
| evm.model.Contig12.1.96 | GATA | D-galacturonic acid | Pentose and glucuronate interconversions | 0.85 |
| Ascorbate and aldarate metabolism | ||||
| D-Galactarate | Ascorbate and aldarate metabolism | 0.84 | ||
| Dehydroascorbic acid | Ascorbate and aldarate metabolism | 0.81 | ||
| L-Gulonic gamma-lactone | Ascorbate and aldarate metabolism | 0.83 | ||
| evm.model.Contig69.54 | DOF | Raffinose | Galactose metabolism | 0.82 |
| evm.model.Contig13.193 | bHLH | Raffinose | Galactose metabolism | 0.80 |
| Stages | ID | Names |
|---|---|---|
| Enlargement stage | map00410 | beta-Alanine metabolism |
| map01040 | Biosynthesis of unsaturated fatty acids | |
| Green mature stage | map00630 | Glyoxylate and dicarboxylate metabolism |
| White mature stage | map02010 | ABC transporters |
| map00052 | Galactose metabolism | |
| Half-red stage | map00052 | Galactose metabolism |
| map00053 | Ascorbate and aldarate metabolism | |
| map02010 | ABC transporters | |
| map00330 | Arginine and proline metabolism | |
| map00908 | Zeatin biosynthesis | |
| map00020 | Citrate cycle (TCA cycle) |
| DEG | Annotation | DAM | Pathway | Pearson’s r |
|---|---|---|---|---|
| evm.model.Contig109.23 | CesA | α-D-Galactose 1-phosphate | Galactose metabolism | 0.93 |
| evm.model.Contig87.31 | β-Glu | α-D-Galactose 1-phosphate | Galactose metabolism | −0.82 |
| evm.model.Contig84.454 | ERF | L-(-)Sorbose | Fructose and mannose metabolism | 0.82 |
| Raffinose | Galactose metabolism | 0.93 | ||
| Isomaltose | Starch and sucrose metabolism | 0.81 | ||
| evm.model.Contig42.394 | MADS-box | UDP-D-Glucose | Pentose and glucuronate interconversions | −0.85 |
| Galactose metabolism | ||||
| Ascorbate and aldarate metabolism | ||||
| Starch and sucrose metabolism | ||||
| L-(-)Sorbose | Fructose and mannose metabolism | −0.92 | ||
| Galactinol | Galactose metabolism | −0.93 | ||
| Sucrose | Galactose metabolism | −0.92 | ||
| Starch and sucrose metabolism | ||||
| Isomaltose | Starch and sucrose metabolism | −0.93 | ||
| evm.model.Contig51.23 | AP2/B3 | UDP-D-Glucose | Pentose and glucuronate interconversions | −0.87 |
| Galactose metabolism | ||||
| Ascorbate and aldarate metabolism | ||||
| Starch and sucrose metabolism | ||||
| Galactinol | Galactose metabolism | −0.83 | ||
| Sucrose | Galactose metabolism | −0.81 | ||
| Starch and sucrose metabolism | ||||
| L-Gulonic gamma-lactone | Ascorbate and aldarate metabolism | 0.88 | ||
| evm.model.Contig11.191 | MYB | UDP-D-Glucose | Pentose and glucuronate interconversions | −0.93 |
| Galactose metabolism | ||||
| Ascorbate and aldarate metabolism | ||||
| Starch and sucrose metabolism | ||||
| L-Gulonic gamma-lactone | Ascorbate and aldarate metabolism | 0.82 | ||
| evm.model.Contig69.54 | DOF | D-Mannose | Fructose and mannose metabolism | 0.94 |
| Galactose metabolism | ||||
| myo-Inositol | Galactose metabolism | 0.94 | ||
| Ascorbate and aldarate metabolism | ||||
| D-Tagatose | Galactose metabolism | 0.91 | ||
| evm.model.Contig13.193 | bHLH | D-Mannose | Fructose and mannose metabolism | 0.90 |
| Galactose metabolism | ||||
| myo-Inositol | Galactose metabolism | 0.88 | ||
| Ascorbate and aldarate metabolism | ||||
| D-Tagatose | Galactose metabolism | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Jin, J.; Li, M.; Kong, D.; Cao, M.; Wang, X.; Li, Y.; Chen, X.; Zhang, X.; Pang, X.; et al. The Key Metabolic Network and Genes Regulating the Fresh Fruit Texture of Jujube (Ziziphus jujuba Mill.) Revealed via Metabolomic and Transcriptomic Analysis. Plants 2023, 12, 2087. https://doi.org/10.3390/plants12112087
Song S, Jin J, Li M, Kong D, Cao M, Wang X, Li Y, Chen X, Zhang X, Pang X, et al. The Key Metabolic Network and Genes Regulating the Fresh Fruit Texture of Jujube (Ziziphus jujuba Mill.) Revealed via Metabolomic and Transcriptomic Analysis. Plants. 2023; 12(11):2087. https://doi.org/10.3390/plants12112087
Chicago/Turabian StyleSong, Shuang, Juan Jin, Meiyu Li, Decang Kong, Ming Cao, Xue Wang, Yingyue Li, Xuexun Chen, Xiuli Zhang, Xiaoming Pang, and et al. 2023. "The Key Metabolic Network and Genes Regulating the Fresh Fruit Texture of Jujube (Ziziphus jujuba Mill.) Revealed via Metabolomic and Transcriptomic Analysis" Plants 12, no. 11: 2087. https://doi.org/10.3390/plants12112087