
Molecular Analysis of Genetic Diversity and Structure of the Lablab (Lablab purpureus (L.) Sweet) Gene Pool Reveals Two Independent Routes of Domestication


Abstract
:1. Introduction
2. Results
2.1. Morphological Variations in Lablab
2.2. Nuclear SSR Variation and Genetic Diversity of Lablab
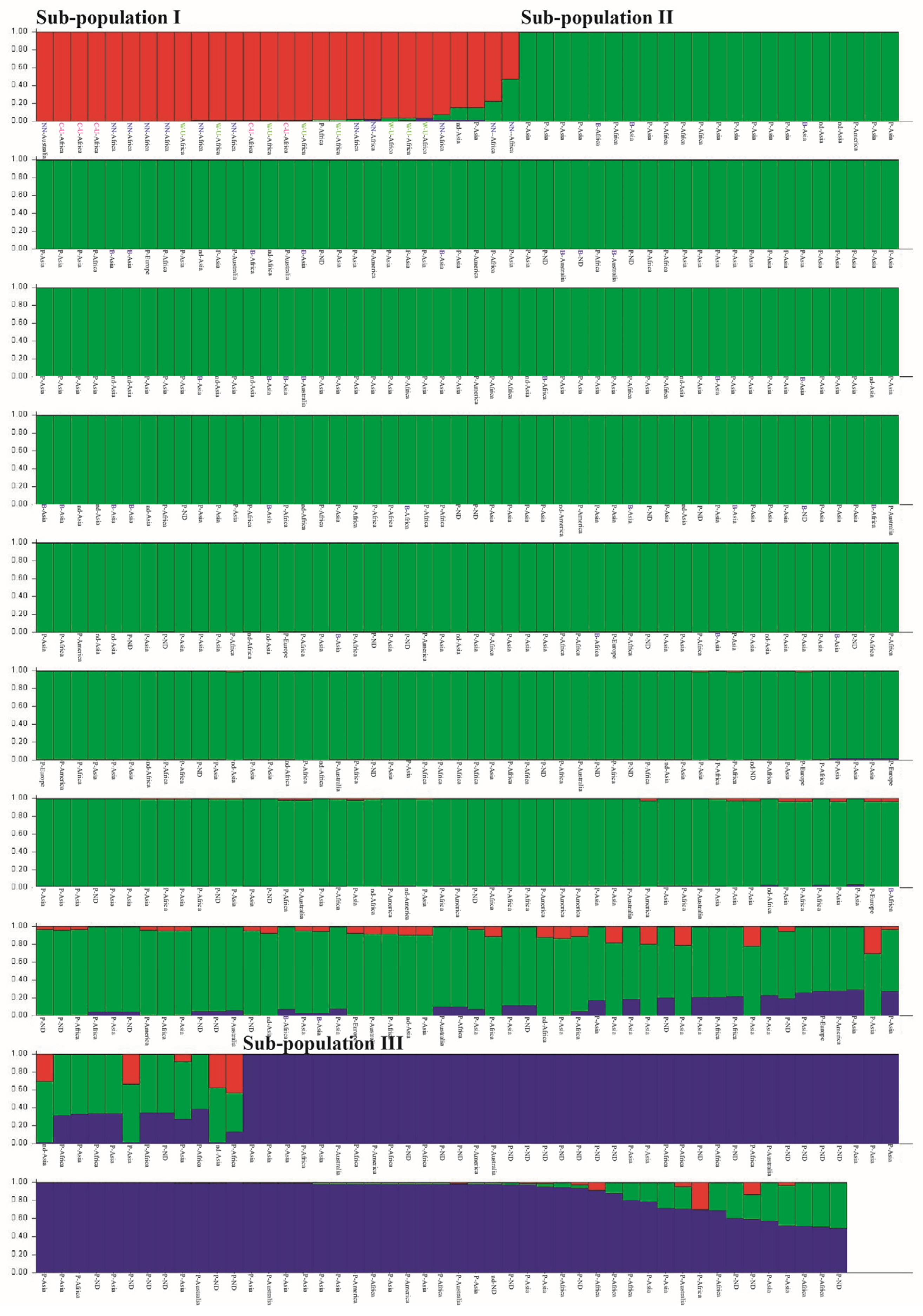
2.3. Population Structure Analysis
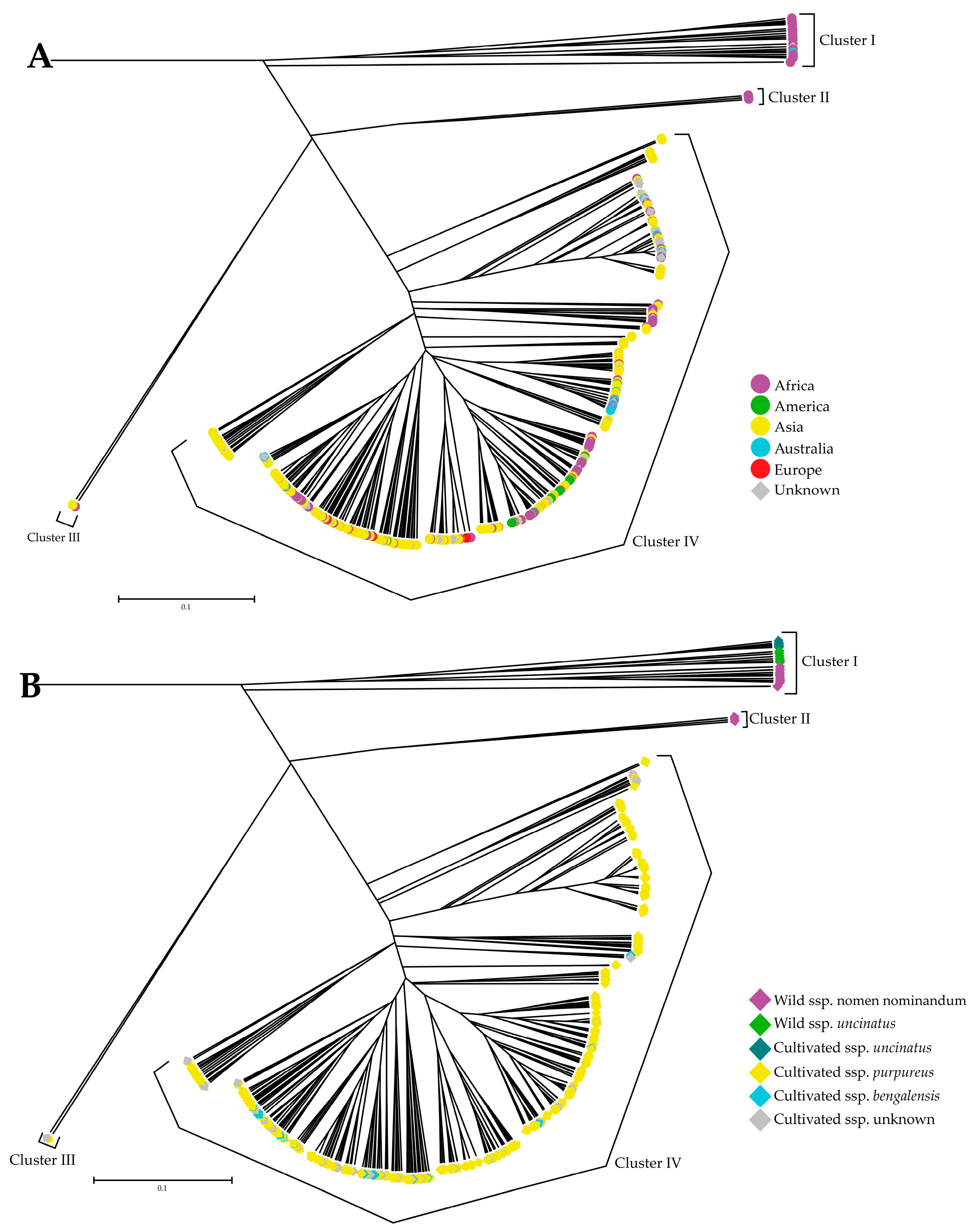
2.4. UPGMA Analysis and Neighbor-Joining Analysis
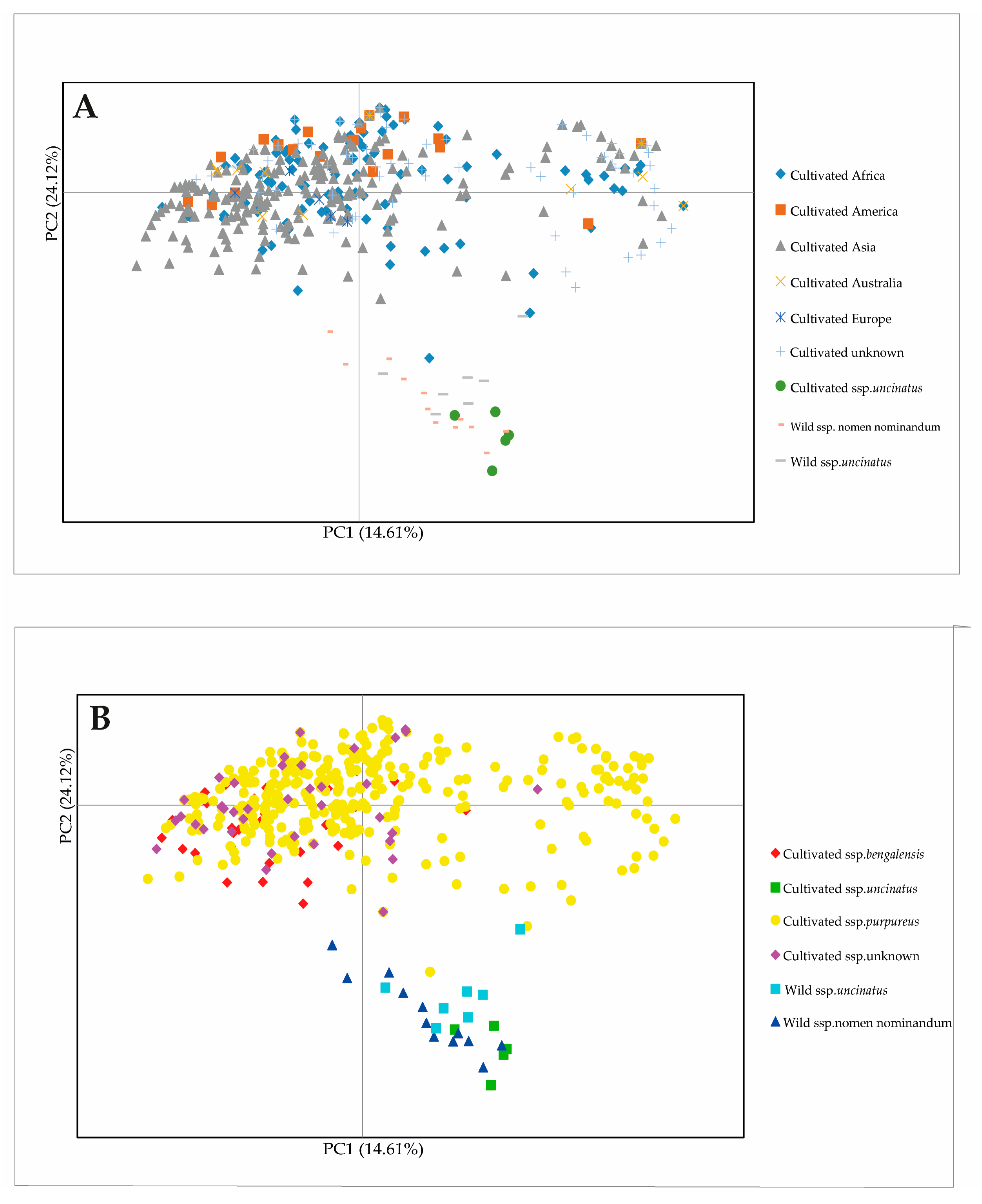
2.5. Principal Coordinate Analysis
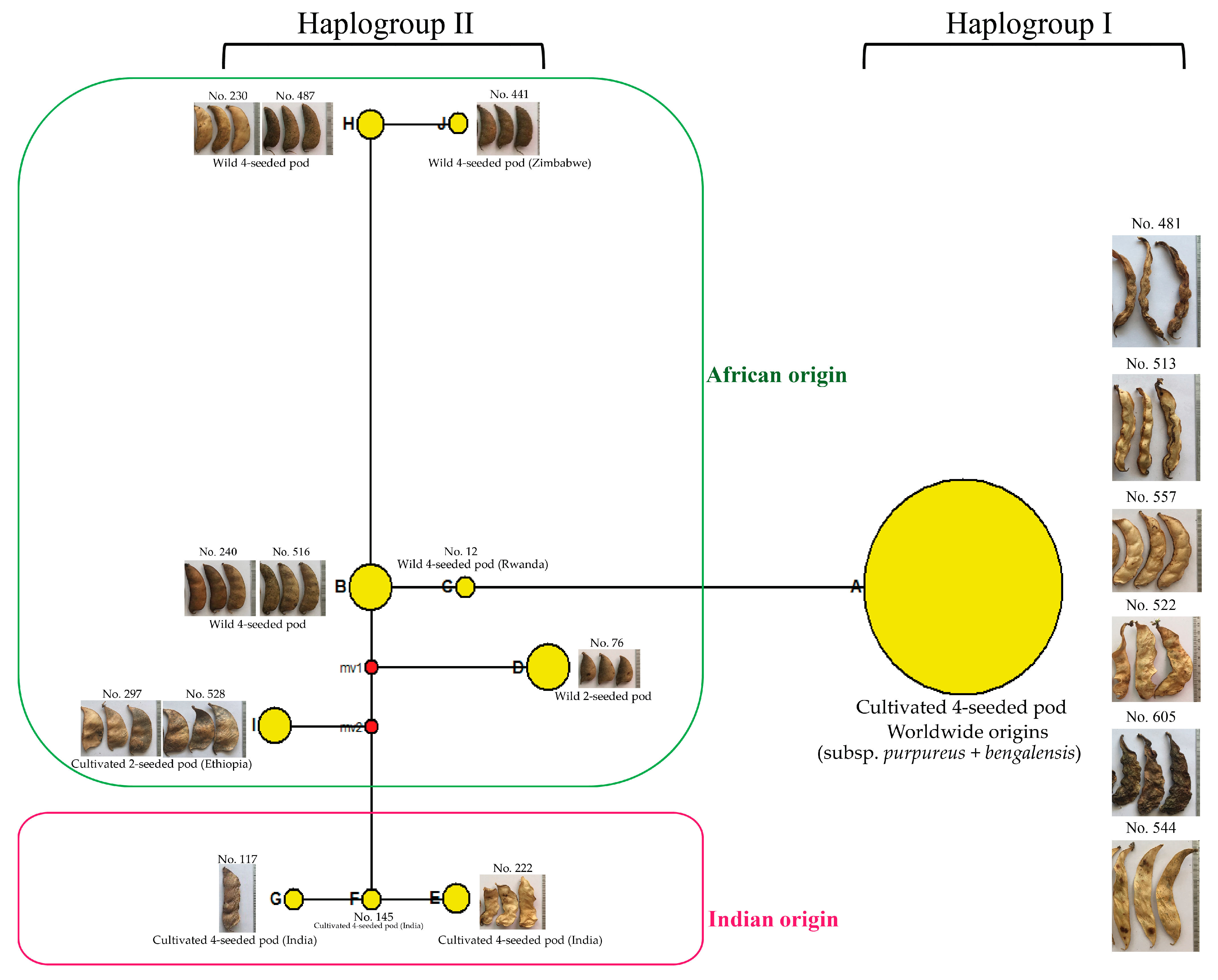
2.6. Chloroplast SSR Variation and Haplotype Diversity of Lablab
2.7. Core Collection Development of Lablab
3. Discussion
Center of Origins, Diversity and Domestication of Lablab
4. Materials and Methods
4.1. Lablab Germplasm and DNA Extraction
4.2. Characterization of Phenotypic Traits
4.3. Nuclear and Chloroplast SSR Markers Analysis
4.4. Genetic Data Analysis
4.5. Development of Lablab Core Collection
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimani, E.N.; Wachira, F.N.; Kinyua, M.G. Molecular diversity of kenyan lablab bean (Lablab purpureus (L.) Sweet) accessions using amplified fragment length polymorphism markers. Am. J. Plant Sci. 2012, 3, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Maass, B.L.; Knox, M.R.; Venkatesha, S.C.; Angessa, T.T.; Ramme, S.; Pengelly, B.C. Lablab purpureus—A crop lost for Africa? Trop. Plant. Biol. 2010, 3, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, S.; Ahmed, R.; Bhowmick, S.; Mamun, A.A.; Hashimoto, M. Proximate composition and fatty acid analysis of Lablab purpureus (L.) legume seed: Implicates to both protein and essential fatty acid supplementation. Springer Plus 2016, 5, 1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deka, R.K.; Sarkar, C.R. Nutrient composition and antinutritional factors of Dolichos lablab L. seeds. Food Chem. 1990, 38, 239–246. [Google Scholar] [CrossRef]
- Kala, B.K.; Soris, P.T.; Mohan, V.R.; Vadivel, V. Nutrient and chemical evaluation of raw seeds of five varieties of Lablab purpureus (L.) sweet. Adv. Bio Res. 2010, 1, 44–53. [Google Scholar]
- El Hardallo, S.B.; el Tiny, A.H.; Nour, M. Chemical characteristics of some legumes grown in Sudan. Sudan J. Food Sci. Technol. 1980, 12, 35–42. [Google Scholar]
- Shaahu, D.T.; Kaankuka, F.G.; Okpanachi, U. Proximate, amino acid, anti-nutritional factor and mineral composition of different varieties of raw lablab purpureus seeds. Intl. J. Sci. Technol. Res. 2015, 4, 157–161. [Google Scholar]
- Murphy, A.M.; Colucci, P.E. A tropical forage solution to poor quality ruminant diets: A review of Lablab purpureus. Livest. Res. Rural. Dev. 1999, 11, 1–17. [Google Scholar]
- Pengelly, B.C.; Maass, B.L. Lablab purpureus (L.) Sweet- diversity, potential use and determination of a core collection of this multi-purpose tropical legume. Genet. Res. Crop Evol. 2001, 48, 261–272. [Google Scholar] [CrossRef]
- Maass, B.L.; Jamnadass, R.H.; Hanson, J.; Pengelly, B.C. Determining sources of diversity in cultivated and wild Lablab purpureus related to provenance of germplasm by using amplified fragment length polymorphism. Genet. Res. Crop Evol. 2005, 52, 683–695. [Google Scholar] [CrossRef]
- Liu, C.J. Genetic diversity and relationships among Lablab purpureus genotypes evaluated using RAPD as markers. Euphytica 1996, 90, 115–119. [Google Scholar] [CrossRef]
- Robotham, O.; Chapman, M. Population genetic analysis of hyacinth bean (Lablab purpureus (L.) Sweet, Leguminosae) indicates an East African origin and variation in drought tolerance. Genet. Resour. Crop Evol. 2017, 64, 139–148. [Google Scholar] [CrossRef]
- D’Souza, M.R.; Devaraj, V.R. Biochemical responses of Hyacinth bean (Lablab purpureus) to salinity stress. Acta Physiol. Plant. 2010, 32, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Cook, B.G.; Pengelly, B.C.; Brown, S.D.; Donnelly, J.L.; Eagles, D.A.; Franco, M.A.; Hanson, J.; Mullen, B.F.; Partridge, I.J.; Peters, M.; et al. Tropical forages: An interactive selection tool. Lablab purpureus. CSIRO, DPI&F(Qld), CIAT, and ILRI, Brisbane. Australia. 2005. Available online: http://www.tropicalforages.info/key/Forages/Media/Html/Lablab_purpureus.htm (accessed on 24 July 2012).
- Amkul, K.; Sookbang, J.M.; Somta, P. Genetic diversity and structure of landrace of lablab (Lablab purpureus (L.) Sweet) cultivars in Thailand revealed by SSR markers. Breed Sci. 2021, 71, 176–183. [Google Scholar] [CrossRef]
- Mihailović, V.; Mikić, A.; Ćeran, M.; Ćupina, B.; Đorđević, V.; Marjanović-Jeromela, A.; Mikić, S.; Perić, V.; Savić, A.; Srebrić, M.; et al. Some aspects of biodiversity, applied genetics and agronomy in hyacinth bean (Lablab purpureus) research. Legume Perspect. 2016, 13, 9–15. [Google Scholar]
- Verdcourt, B. Studies in the Leguminosae-Papilionoideae for the Flora of Tropical East Africa III. Kew Bull. 1970, 24, 379–447. [Google Scholar] [CrossRef] [Green Version]
- Westphal, E. Pulses in Ethiopia, Their Taxonomy and Ecological Significance. Centre for Agricultural Publishing and Documentation, Wageningen. 1974, p. 279. Available online: https://edepot.wur.nl/197905 (accessed on 10 June 2022).
- Fuller, D.Q. African crops in prehistoric South Asia: A critical review. In Food, Fuel and Fields: Progress in African Archaeobotany; Neumann, K., Butler, E.A., Kahlheber, S., Eds.; Africa Praehistorica 15; Heinrich-Barth-Institut: Köln, Germany, 2003; pp. 239–271. [Google Scholar]
- Venkatesha, S.C.; Gowda, M.B.; Mahadevu, P.; Rao, A.M.; Kim, D.-J.; Ellis, T.H.N.; Knox, M.R. Genetic diversity within Lablab purpureus and the application of gene-specific markers from a range of legume species. Plant Genet. Resour. Characterisation Util. 2007, 5, 154–171. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.L.; Morris, J.B.; Barkley, N.A.; Dean, R.E.; Jenkins, T.M.; Pederson, G.A. Evaluation of genetic diversity of the USDA Lablab purpureus germplasm collection using simple sequence repeat markers. J. Hortic. Sci. Biotechnol. 2007, 82, 571–578. [Google Scholar] [CrossRef]
- Rai, N.; Singh, P.K.; Rai, A.C.; Rai, V.P.; Singh, M. Genetic diversity in Indian bean (Lablab purpureus) germplasm based on morphological traits and RAPD markers. Indian J. Agric. Sci. 2011, 81, 801–806. [Google Scholar]
- Shivachi, A.; Kiplagat, K.O.; Kinyua, G.M. Microsatellite analysis of selected Lablab purpureus genotypes in Kenya. Rwanda J. 2012, 28, 39–52. [Google Scholar]
- Zhang, G.; Xu, S.; Mao, W.; Gong, Y.; Hu, Q. Development of EST-SSR markers to study genetic diversity in hyacinth bean (Lablab purpureus L.). POJ 2013, 6, 295–301. [Google Scholar]
- Maass, B.L.; Usongo, M. Changes in seed characteristics during the domestication of the lablab bean (Lablab purpureus (L.) Sweet: Papilionoideae). Crop Pasture Sci. 2007, 58, 9–19. [Google Scholar] [CrossRef]
- Sultana, N.; Ozaki, Y.; Okubo, H. The use of RAPD markers in lablab bean (Lablab purpureus (L.) Sweet) phylogeny. Bull. Inst. Trop Agric. Kyushu Univ. 2000, 23, 45–51. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Maass, B.L. Origin, domestication and global dispersal of Lablab purpureus (L.) Sweet (Fabaceae): Current understanding. Legume Perspect. 2016, 13, 5–8. [Google Scholar]
- Maass, B.L.; Robotham, O.; Chapman, M.A. Evidence for two domestication events of hyacinth bean (Lablab purpureus (L.) Sweet): A comparative analysis of population genetic data. Genet. Resour. Crop Evol. 2017, 64, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Rivals, F. Le dolique d’Egypte ou lablab. 2. Sous-espèce, bases de classement des variétés, variabilité des conditions de floraison, intérêt agricole des variétés de jours courts. Rev. Int. Bot. Appl. Agric. Trop. 1953, 33, 518–537. [Google Scholar]
- Frankel, O.H.; Brown, A.H.D. Plant genetic resources today: A critical appraisal. In Crop Genetic Resources Conservation and Evaluation; Holden, J.H.W., Williams, J.T., Eds.; George Allan and Unwin: London, UK, 1984; pp. 249–257. [Google Scholar]
- Lodhi, M.A.; Ye, G.N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol. Biol. Rep. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Keerthi, C.M.; Ramesh, S.; Byregowda, M.; Vaijayanthi, P.V. Simple sequence repeat (SSR) marker assay-based genetic diversity among dolichos bean (Lablab purpureus L. Sweet) advanced breeding lines differing for productivity per se traits. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 3736–3744. [Google Scholar] [CrossRef]
- Wang, X.W.; Kaga, A.; Tomooka, N.; Vaughan, D.A. The development of SSR markers by a new method in plants and their application to gene flow studies in azuki bean [Vigna angularis (Willd.) Ohwi & Ohashi]. Theor. Appl. Genet. 2004, 109, 352–360. [Google Scholar]
- Chankaew, S.; Isemura, T.; Isobe, S.; Kaga, A.; Tomooka, N.; Somta, P.; Shirasawa, H.H.K.; Vaughan, D.A.; Srinives, P. Detection of genome donor species of neglected tetraploid crop Vigna reflexo-pilosa (Cre’ole Bean), and genetic structure of diploid species based on newly developed EST-SSR markers from azuki Bean (Vigna angularis). PLoS ONE 2014, 9, e104990. [Google Scholar] [CrossRef]
- Somta, P.; Seehalak, W.; Srinives, P. Development, characterization and cross-species amplification of mungbean (Vigna radiata) genic microsatellite markers. Conserv. Genet. 2009, 10, 1939–1943. [Google Scholar] [CrossRef]
- Pan, L.; Li, Y.; Guo, R.; Wu, H.; Hu, Z.; Chen, C. Development of 12 chloroplast microsatellite markers in Vigna unguiculata (Fabaceae) and amplification in Phaseolus vulgaris. Appl. Plant Sci. 2014, 2, 1300075. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for STRUCTURE Software: Version 2.2. 2007. Available online: http://pritch.bsd.uchicago.edu/software/structure22/readme.pdf (accessed on 28 April 2020).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research–an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Kim, K.W.; Chung, H.K.; Cho, G.T.; Ma, K.H.; Chandrabalan, D.; Gwag, J.G.; Kim, T.S.; Cho, E.G.; Park, Y.J. PowerCore: A program applying the advanced M strategy with a heuristic search for establishing core sets. Bioinformatics 2007, 23, 2155–2162. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Cultivated | Wild | t-Test (Cultivated vs. Wild) | ||
|---|---|---|---|---|---|
| uncinatus | purpureus + bengalensis | uncinatus | nomen nominandum | ||
| Stem | |||||
| Stem color | Purple, Green | Purple, Green | Purple, Green | Purple, Green | - |
| Leave | |||||
| Leave color | Green | Purple, Green | Green | Green | - |
| Flower | |||||
| Flower color | Purple | Purple, White | Purple | Purple | - |
| Day to 1st flower (days) | 60–82, average 73.20 | 17–154, average 93.97 | 58–98, average 68.57 | 62–149, average 115.40 | ns |
| Pod | |||||
| Fresh pod length (cm) | 3.20–4.90, average 4.11 | 3.30–12.50, average 5.98 | 2.74–3.10, average 2.95 | 3.00–5.20, average 3.74 | ** |
| Fresh pod width (cm) | 2.20–2.52, average 2.37 | 0.63–3.30, average 1.99 | 1.32–1.78, average 1.56 | 0.30–1.74, average 1.21 | ** |
| Dry pod length (cm) | 4.12–5.68, average 4.67 | 3.10–15.04, average 5.95 | 3.08–3.46, average 3.25 | 3.18–4.35, average 3.70 | ** |
| Dry pod width (cm) | 2.14–2.54, average 2.36 | 0.86–6.40, average 1.90 | 1.30–1.88, average 1.65 | 1.20–1.50, average 1.32 | ** |
| Fresh pod color | Green | Purple, Green | Green | Green | |
| Dry pod color | Brown | Brown | Brown | Brown | |
| Seed | |||||
| Seed length (mm) | 13.62–14.16, average 13.89 | 6.08–14.29, average 10.91 | 7.74–8.92, average 8.31 | 5.40–7.39, average 6.52 | ** |
| Seed width (mm) | 10.13–10.30, average 10.21 | 4.42–10.23, average 7.74 | 5.60–6.66, average 6.17 | 4.26–6.13, average 5.00 | ** |
| Seed thickness (mm) | 6.44–6.72, average 6.58 | 1.92–8.19, average 4.90 | 2.06–3.54, average 2.97 | 2.45–6.65, average 3.45 | ** |
| Number of seeds per pod (count) | 1.50–2.20, average 1.90 | 2.20–6.00, average 3.75 | 2.00–2.20, average 2.09 | 2.60–4.60, average 3.71 | ** |
| Marker Name | NA | Allele Size Range (Base Pairs) | Major Allele Frequency | Gene Diversity (HE) | Observed Heterozygosity (HO) | PIC |
|---|---|---|---|---|---|---|
| c13319_g1_i1 | 7 | 184-218 | 0.9440 | 0.1078 | 0.0474 | 0.1058 |
| c13353_g1_i1 | 10 | 252-270 | 0.7550 | 0.4055 | 0.0177 | 0.3775 |
| c17963_g1_i1 | 17 | 200-232 | 0.5195 | 0.6844 | 0.0453 | 0.6587 |
| c21512_g1_i1 | 9 | 225-281 | 0.7616 | 0.3903 | 0.0486 | 0.3558 |
| c22788_g1_i1 | 11 | 333-389 | 0.8443 | 0.2809 | 0.0773 | 0.2721 |
| c23309_g1_i1 | 8 | 273-301 | 0.7031 | 0.4765 | 0.0271 | 0.4473 |
| Hbp_006 | 4 | 170-200 | 0.9400 | 0.1137 | 0.0105 | 0.1089 |
| Hbp_009 | 10 | 374-390 | 0.7458 | 0.4175 | 0.0935 | 0.3882 |
| Hbp_010 | 6 | 240-296 | 0.9633 | 0.0715 | 0.0000 | 0.0706 |
| Hbp_012 | 2 | 256-260 | 0.9958 | 0.0084 | 0.0000 | 0.0083 |
| KTD184 | 5 | 176-187 | 0.8650 | 0.2451 | 0.0082 | 0.2352 |
| KTD225 | 10 | 133-162 | 0.5514 | 0.5375 | 0.0535 | 0.4410 |
| KTD241 | 8 | 144-158 | 0.6301 | 0.4995 | 0.0369 | 0.4172 |
| KTD245 | 19 | 220-310 | 0.6667 | 0.5275 | 0.0380 | 0.5021 |
| KTD249 | 5 | 248-260 | 0.7589 | 0.3951 | 0.0418 | 0.3615 |
| Overall | 131 | |||||
| Mean | 8.73 | 0.7763 | 0.3441 | 0.0364 | 0.3167 |
| Type/Region | Subregion | Sample Size | NA | MAF | HE | HO |
|---|---|---|---|---|---|---|
| Cultivated | 474 | 112 | 0.80 | 0.3139 | 0.0325 | |
| Africa | 120 | 79 | 0.78 | 0.3393 | 0.0421 | |
| Central | 23 | 23 | 0.90 | 0.1320 | 0.0381 | |
| East | 61 | 61 | 0.77 | 0.3565 | 0.0391 | |
| North | 19 | 19 | 0.88 | 0.1250 | 0.0333 | |
| South | 52 | 52 | 0.78 | 0.3158 | 0.0519 | |
| West | 40 | 40 | 0.80 | 0.2917 | 0.0373 | |
| America | 22 | 33 | 0.88 | 0.1869 | 0.0219 | |
| North | 20 | 20 | 0.93 | 0.1063 | 0.0167 | |
| South | 32 | 32 | 0.87 | 0.2006 | 0.0229 | |
| Asia | 78 | 78 | 0.79 | 0.3018 | 0.0273 | |
| East | 31 | 31 | 0.84 | 0.2467 | 0.0417 | |
| South | 73 | 73 | 0.79 | 0.3175 | 0.0306 | |
| Southeast | 45 | 45 | 0.83 | 0.2370 | 0.0166 | |
| West | 19 | 19 | 0.89 | 0.1259 | 0.0000 | |
| Australia | 15 | 29 | 0.81 | 0.2426 | 0.0249 | |
| Europe | 5 | 25 | 0.85 | 0.2197 | 0.0433 | |
| unknown | 75 | 58 | 0.82 | 0.2568 | 0.0378 | |
| Wild | 19 | 79 | 0.52 | 0.6059 | 0.1417 | |
| Africa | 17 | 73 | 0.52 | 0.6024 | 0.1491 | |
| Central | 20 | 1.33 | 0.82 | - | - | |
| East | 38 | 2.53 | 0.63 | 0.4570 | 0.1044 | |
| South | 57 | 3.80 | 0.58 | 0.5489 | 0.1807 | |
| Australia | 1 | 14 | 0.83 | - | - | |
| unknown | 1 | 13 | 0.77 | - | - | |
| Type/Origin | Sample Size | Major Allele Frequency | Average Alleles per Locus | Gene Diversity (HE) | Observed Heterozygosity (HO) |
|---|---|---|---|---|---|
| Cultivated | 474 | 0.80 | 7.47 | 0.3139 | 0.0325 |
| ssp. purpureus | 397 | 0.81 | 6.60 | 0.2971 | 0.0332 |
| ssp. bengalensis | 33 | 0.81 | 2.53 | 0.2584 | 0.0306 |
| ssp. uncinatus | 5 | 0.91 | 1.47 | 0.1222 | 0.0689 |
| Unknown | 39 | 0.79 | 3.67 | 0.3066 | 0.0251 |
| Wild | 19 | 0.52 | 5.27 | 0.6059 | 0.1417 |
| ssp. uncinatus | 7 | 0.80 | 2.00 | 0.2775 | 0.1616 |
| ssp. nomen nominandum | 12 | 0.57 | 4.60 | 0.5643 | 0.1338 |
| VgcpSSR04 (Base Pair) | VgcpSSR05 (Base Pair) | VgcpSSR10 (Base Pair) | VgcpSSR11 (Base Pair) | VgcpSSR12 (Base Pair) | VgcpSSR14 (Base Pair) | ||
|---|---|---|---|---|---|---|---|
| Haplogroup I | Frequency | ||||||
| A | 216 | 206 | 183 | 204 | 236 | 229 | 420 |
| Haplogroup II | |||||||
| B | 222 | 211 | 187 | 206 | 244 | 229 | 5 |
| C | 222 | 211 | 187 | 206 | 240 | 229 | 1 |
| D | 222 | 211 | 186 | 212 | 244 | 229 | 5 |
| E | 222 | 212 | 187 | 202 | 244 | 225 | 1 |
| F | 222 | 215 | 186 | 202 | 244 | 225 | 1 |
| G | 222 | 216 | 186 | 202 | 244 | 225 | 1 |
| H | 222 | 211 | 187 | 202 | 249 | 238 | 2 |
| I | 222 | 215 | 185 | 206 | 244 | 229 | 3 |
| J | 224 | 211 | 187 | 202 | 249 | 238 | 1 |
| Mean | |||||||
| No. alleles per locus | 3 | 5 | 4 | 5 | 4 | 4 | 4.1667 |
| Gene diversity | 0.1067 | 0.1105 | 0.1089 | 0.1028 | 0.1027 | 0.0371 | 0.0948 |
| PIC | 0.1015 | 0.1073 | 0.1068 | 0.1011 | 0.0995 | 0.0368 | 0.0922 |
| Marker | No. of Alleles per Locus | Allelic Richness | Observed Heterozygosity (HO) | Gene Diversity (HE) |
|---|---|---|---|---|
| c13319 | 7 | 8 | 0.1364 | 0.4556 |
| c22788 | 11 | 13 | 0.1538 | 0.5562 |
| KTD225 | 10 | 12 | 0.0833 | 0.727 |
| c17963 | 17 | 20 | 0.0889 | 0.8054 |
| Hbp006 | 4 | 5 | 0.0455 | 0.4708 |
| Hbp009 | 10 | 13 | 0.2286 | 0.68 |
| KTD184 | 5 | 7 | 0.0417 | 0.5684 |
| KTD249 | 5 | 7 | 0.1277 | 0.5593 |
| KTD241 | 8 | 9 | 0.0208 | 0.7029 |
| KTD245 | 19 | 21 | 0.1087 | 0.8003 |
| c23309 | 8 | 8 | 0.0000 | 0.6686 |
| Hbp012 | 2 | 2 | 0.0000 | 0.0416 |
| Hbp010 | 6 | 6 | 0.0000 | 0.3894 |
| c13353 | 10 | 13 | 0.0667 | 0.5899 |
| c21512 | 9 | 11 | 0.1163 | 0.6003 |
| Overall | 131 | 10.33 | 0.0812 | 0.5744 |
| Organ | Traits | Evaluation |
|---|---|---|
| Stem | Stem color | Green or Purple |
| Leave | Leave color | Green or Purple |
| Flower | Flower color | White or Purple |
| Day to 1st flower | Number of days from planting to 1st flowering | |
| Pod | Fresh pod length (cm) | Length of straight pod (use 5 pods) |
| Fresh pod width (cm) | Maximum width (use 5 pods) | |
| Dry pod length (cm) | Length of straight pod (use 5 pods) | |
| Dry pod width (cm) | Maximum width (use 5 pods) | |
| Fresh pod color | Green or Purple | |
| Dry pod color | Black or Brown | |
| Seed | Seed length (mm) | Maximum distance from top to bottom of the seed (use 5 seeds) |
| Seed width (mm) | Maximum distance from hilum to its opposite side (use 5 seeds) | |
| Seed thickness (mm) | Maximum distance between both sides of hilum (use 5 seeds) | |
| Number of seeds per pod (count) | Number of seed per pod |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kongjaimun, A.; Takahashi, Y.; Yoshioka, Y.; Tomooka, N.; Mongkol, R.; Somta, P. Molecular Analysis of Genetic Diversity and Structure of the Lablab (Lablab purpureus (L.) Sweet) Gene Pool Reveals Two Independent Routes of Domestication. Plants 2023, 12, 57. https://doi.org/10.3390/plants12010057
Kongjaimun A, Takahashi Y, Yoshioka Y, Tomooka N, Mongkol R, Somta P. Molecular Analysis of Genetic Diversity and Structure of the Lablab (Lablab purpureus (L.) Sweet) Gene Pool Reveals Two Independent Routes of Domestication. Plants. 2023; 12(1):57. https://doi.org/10.3390/plants12010057
Chicago/Turabian StyleKongjaimun, Alisa, Yu Takahashi, Yosuke Yoshioka, Norihiko Tomooka, Rachsawan Mongkol, and Prakit Somta. 2023. "Molecular Analysis of Genetic Diversity and Structure of the Lablab (Lablab purpureus (L.) Sweet) Gene Pool Reveals Two Independent Routes of Domestication" Plants 12, no. 1: 57. https://doi.org/10.3390/plants12010057