
Effect of Recurrent Salt and Drought Stress Treatments on the Endangered Halophyte Limonium angustebracteatum Erben

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
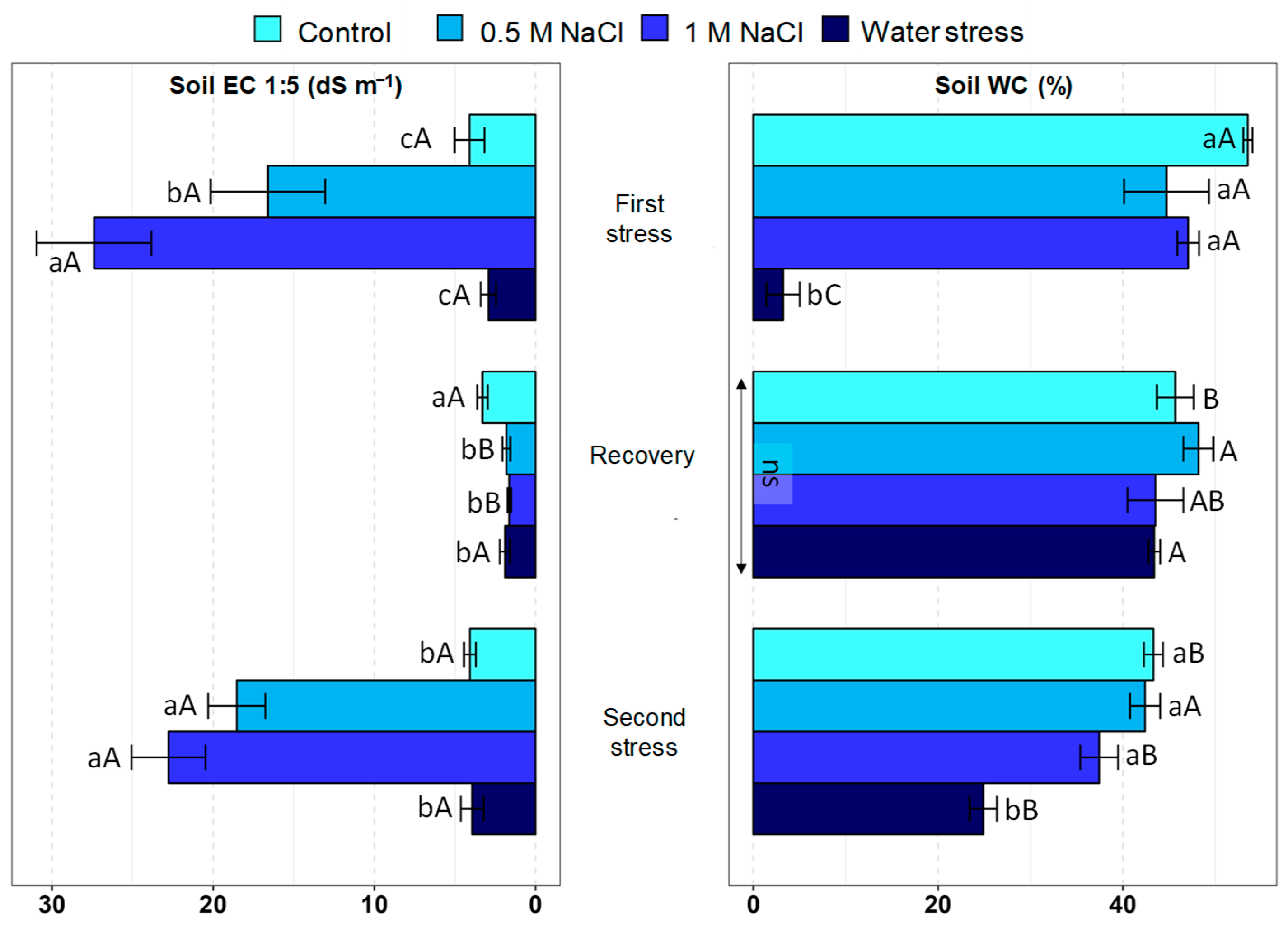
2.1. Substrate Salinity and Water Content
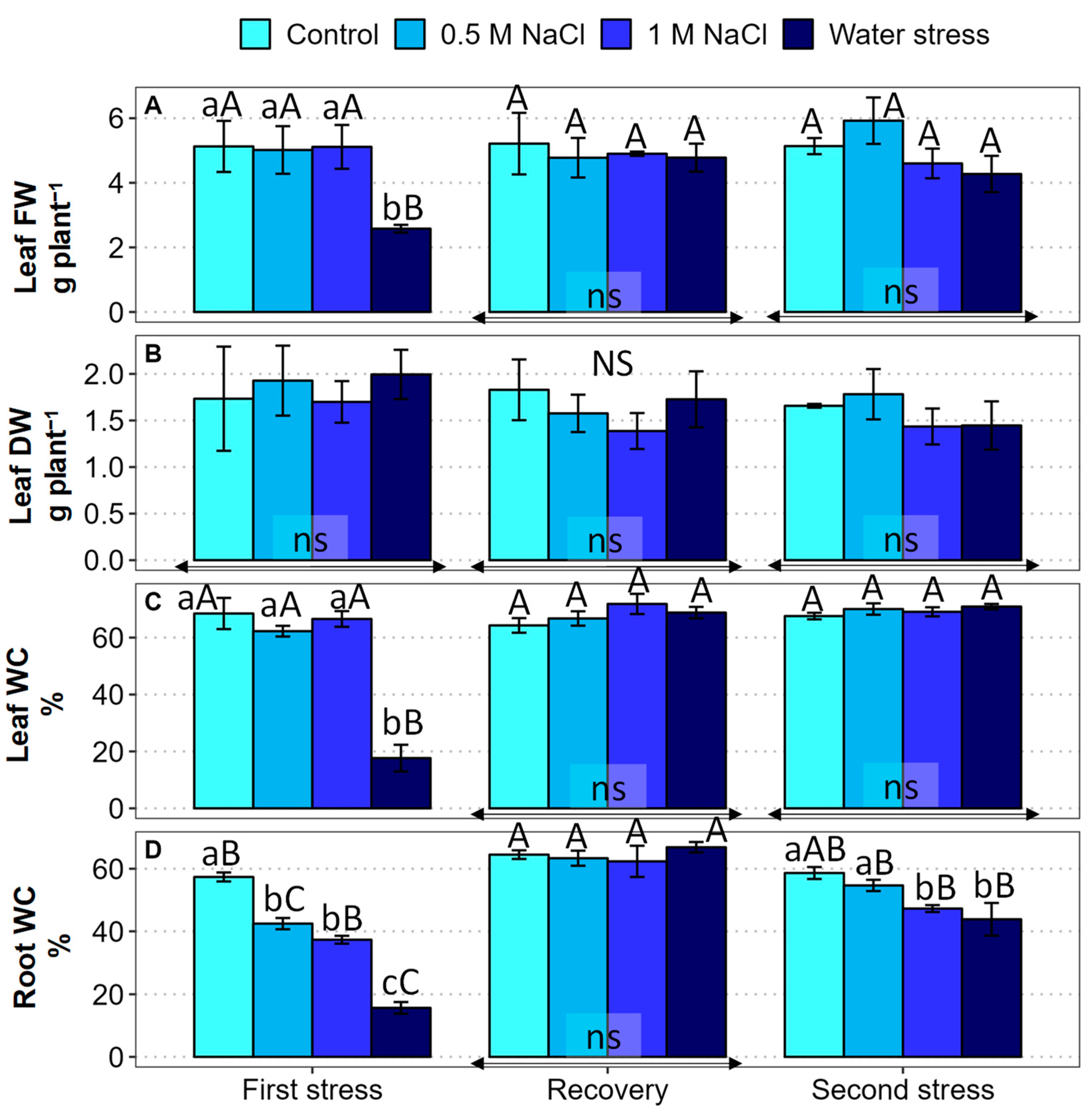
2.2. Growth Parameters
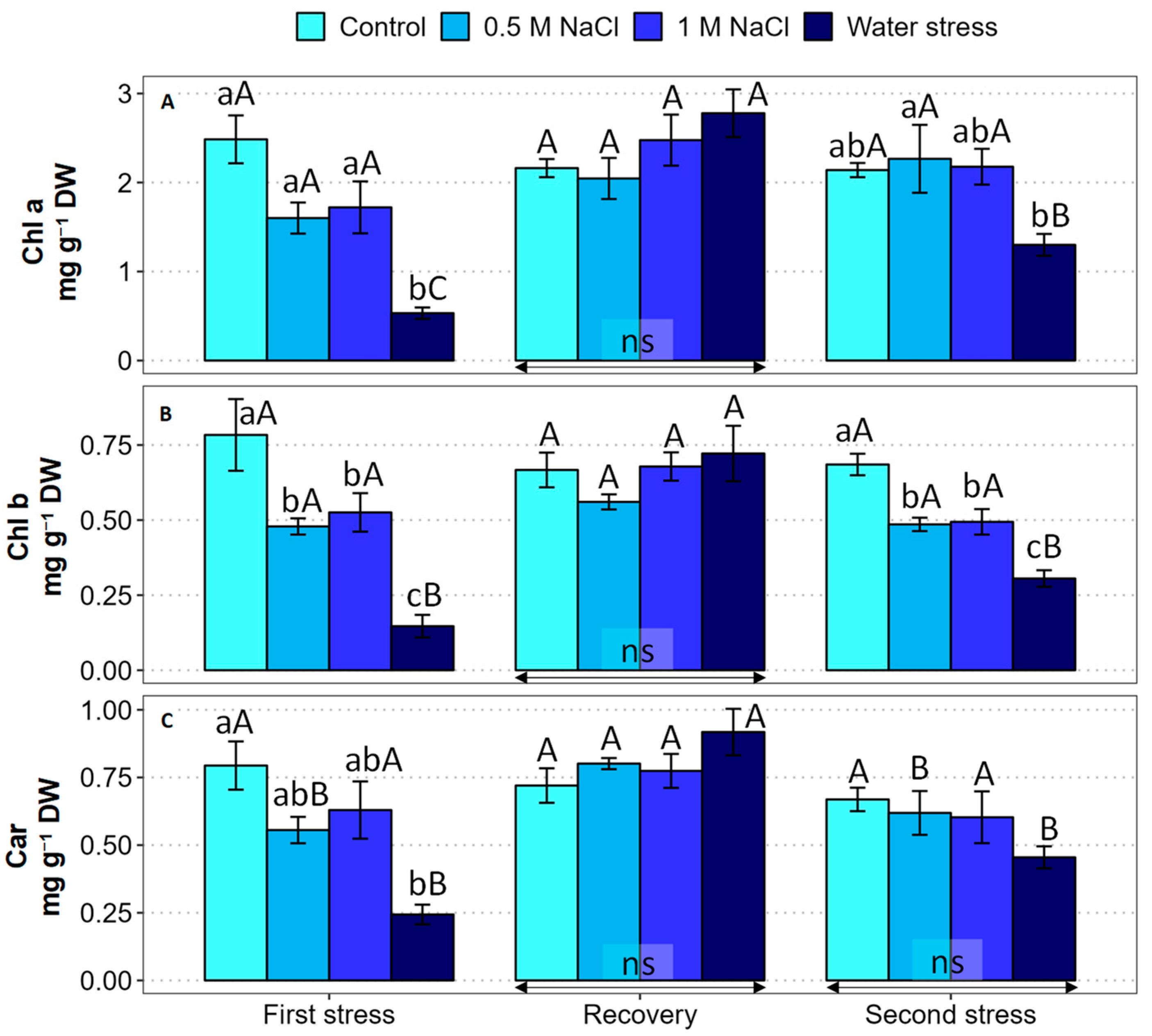
2.3. Photosynthetically Active Pigments
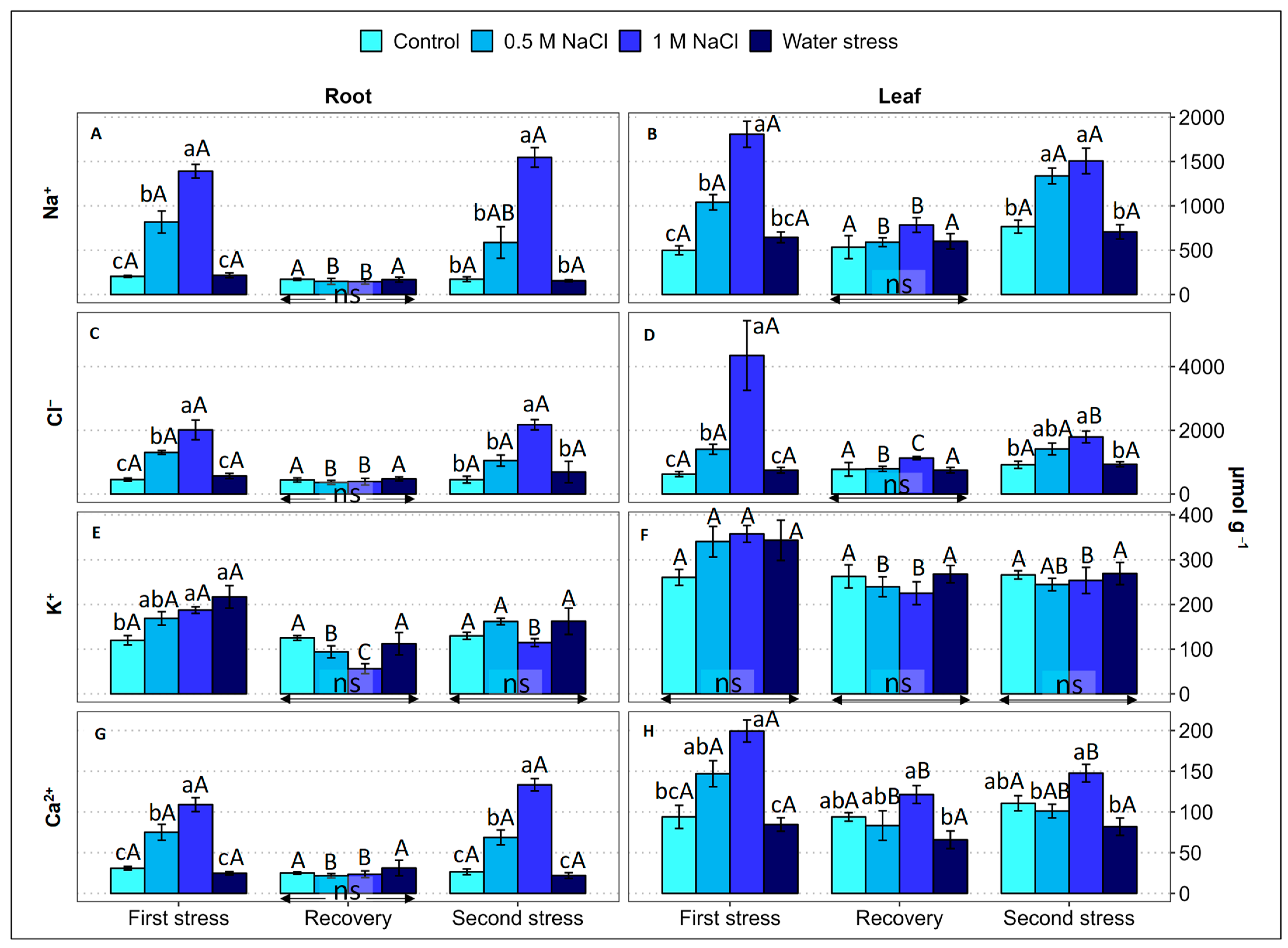
2.4. Ion Accumulation
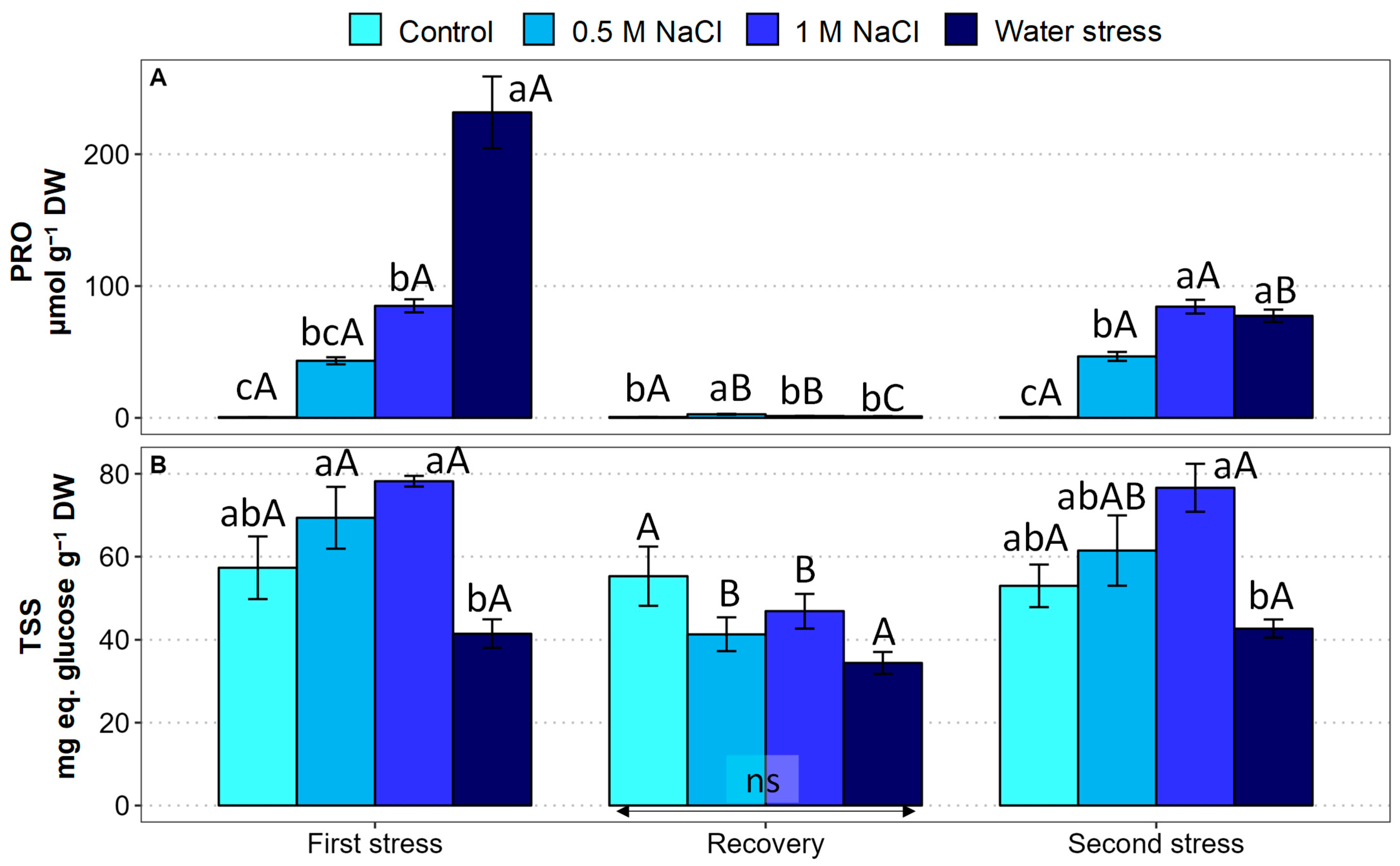
2.5. Osmolyte Contents
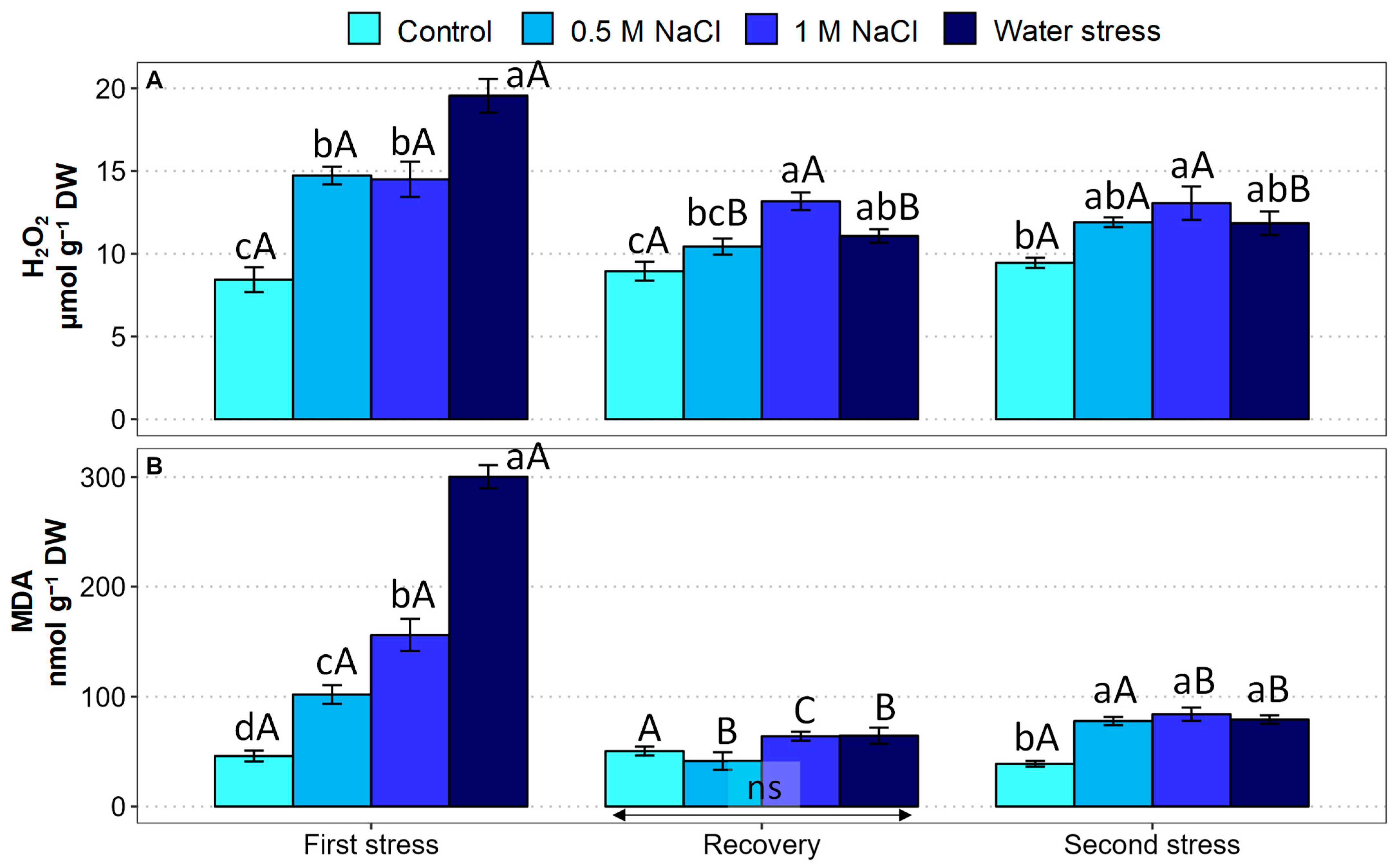
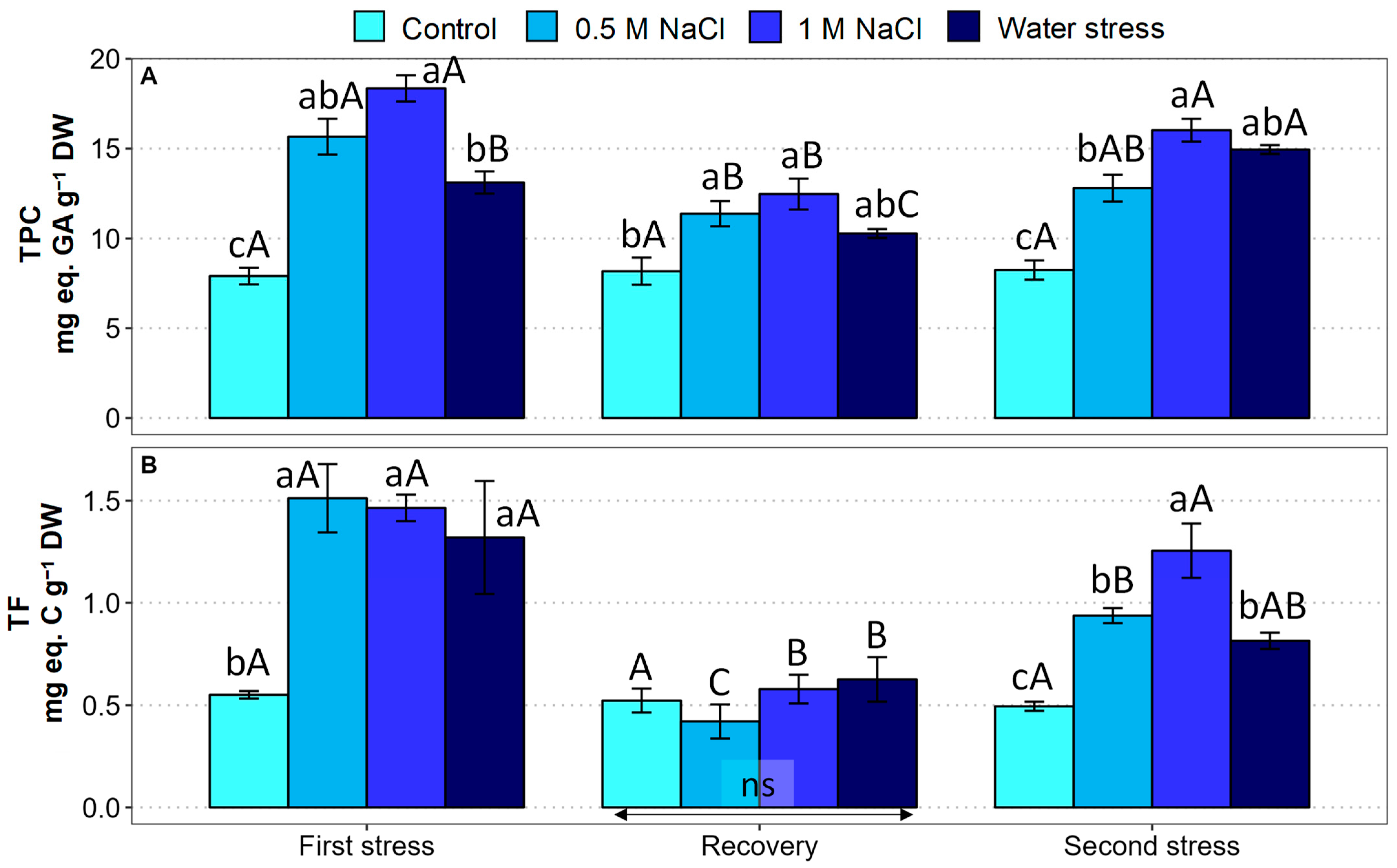
2.6. Oxidative Stress Markers and Antioxidant Compounds
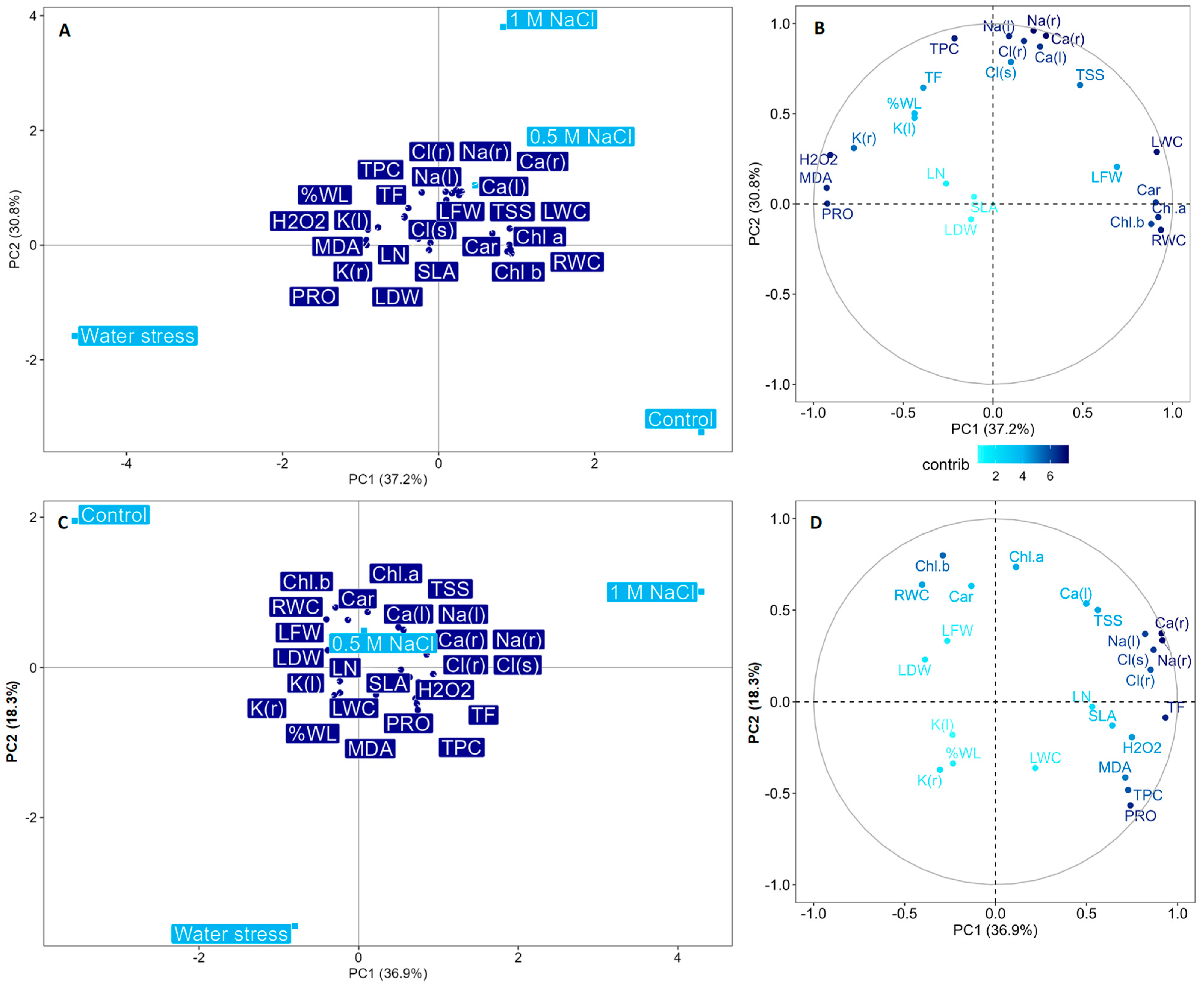
2.7. General Pattern of Trait Responses
3. Discussion
4. Materials and Methods
4.1. Plant Material
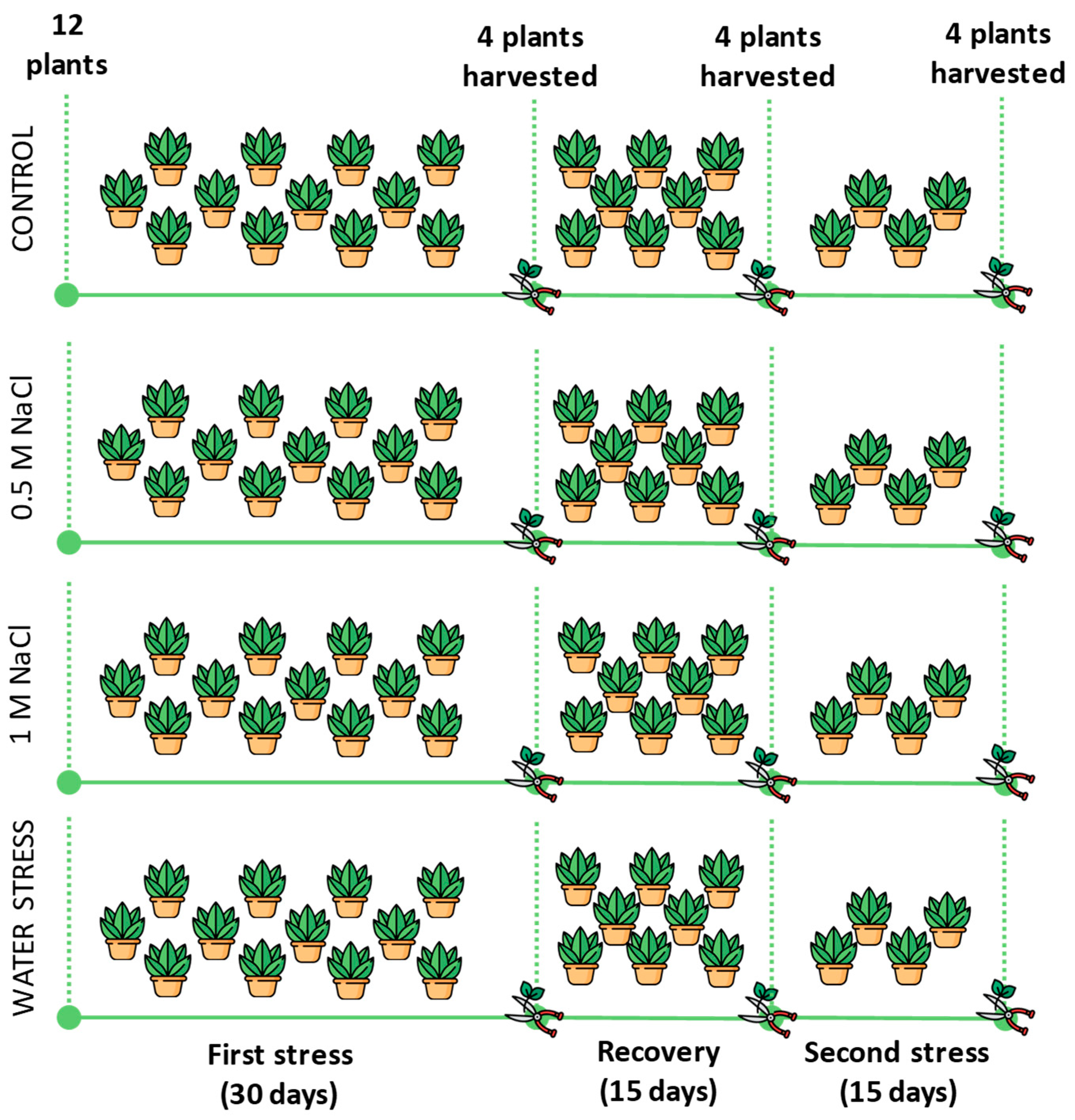
4.2. Experimental Design
4.3. Morphological Parameters
4.4. Photosynthetic Pigments
4.5. Quantification of Ions
4.6. Quantification of Osmolytes
4.7. Determination of Oxidative Stress Markers and Antioxidant Compounds
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zedler, J.B.; Bonin, C.L.; Larkin, D.J.; Varty, A. Salt Marshes. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; pp. 614–622. ISBN 978-0-444-64130-4. [Google Scholar]
- Gedan, K.B.; Kirwan, M.L.; Wolanski, E.; Barbier, E.B.; Silliman, B.R. The Present and Future Role of Coastal Wetland Vegetation in Protecting Shorelines: Answering Recent Challenges to the Paradigm. Clim. Chang. 2011, 106, 7–29. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Duarte, C.M. Dimensions of Blue Carbon and Emerging Perspectives. Biol. Lett. 2019, 15, 20180781. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ge, Z.M.; Fei, B.L.; Zhang, C.; Liu, Q.X.; Zhang, L.Q. Soil Carbon and Nitrogen Storage in Recently Restored and Mature Native Scirpus Marshes in the Yangtze Estuary, China: Implications for Restoration. Ecol. Eng. 2017, 104, 150–157. [Google Scholar] [CrossRef]
- Taylor, B.W.; Paterson, D.M.; Baxter, J.M. Sediment Dynamics of Natural and Restored Bolboschoenus maritimus Saltmarsh. Front. Ecol. Evol. 2019, 7, 237. [Google Scholar] [CrossRef] [Green Version]
- Dee, L.E.; Keyes, A.A. Analyzing Ecosystem Services as Part of Ecological Networks in Three Salt Marsh Ecosystems. Ecology 2022, 103, e3609. [Google Scholar] [CrossRef] [PubMed]
- Wedding, L.M.; Reiter, S.; Moritsch, M.; Hartge, E.; Reiblich, J.; Gourlie, D.; Guerry, A. Embedding the Value of Coastal Ecosystem Services into Climate Change Adaptation Planning. PeerJ 2022, 10, e13463. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A Blueprint for Blue Carbon: Toward an Improved Understanding of the Role of Vegetated Coastal Habitats in Sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global Carbon Sequestration in Tidal, Saline Wetland Soils. Global Biogeochem. Cycles 2003, 17, 1111. [Google Scholar] [CrossRef]
- Lombardi, T.; Bertacchi, A.; Pistelli, L.; Pardossi, A.; Pecchia, S.; Toffanin, A.; Sanmartin, C. Biological and Agronomic Traits of the Main Halophytes Widespread in the Mediterranean Region as Potential New Vegetable Crops. Horticulturae 2022, 8, 195. [Google Scholar] [CrossRef]
- Matinzadeh, Z.; López-Angulo, J.; Escudero, A.; Palacio, S.; Abedi, M.; Akhani, H. Functional Structure of Plant Communities along Salinity Gradients in Iranian Salt Marshes. Plant-Environ. Interact. 2022, 3, 16–27. [Google Scholar] [CrossRef]
- Álvarez-Rogel, J.; Alcaraz, F.; Silla, R. Soil Salinity and Moisture Gradients and Plant Zonation in Mediterranean Salt Marshes of Southeast Spain. Wetlands 2000, 20, 357–372. [Google Scholar] [CrossRef]
- Gil, R.; Bautista, I.; Boscaiu, M.; Lidón, A.; Wankhade, S.; Sánchez, H.; Llinares, J.; Vicente, O. Responses of Five Mediterranean Halophytes to Seasonal Changes in Environmental Conditions. AoB Plants 2014, 6, plu049. [Google Scholar] [CrossRef] [PubMed]
- Koutroumpa, K.; Theodoridis, S.; Warren, B.H.; Jiménez, A.; Celep, F.; Doğan, M.; Romeiras, M.M.; Santos-Guerra, A.; Fernández-Palacios, J.M.; Caujapé-Castells, J. An Expanded Molecular Phylogeny of Plumbaginaceae, with Emphasis on Limonium (Sea Lavenders): Taxonomic Implications and Biogeographic Considerations. Ecol. Evol. 2018, 8, 12397–12424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brullo, S.; Erben, M. The Genus Limonium (Plumbaginaceae) in Greece. Phytotaxa 2016, 240, 1–212. [Google Scholar] [CrossRef]
- Baker, H.G. The Evolution, Functioning and Breakdown of Heteromorphic Incompatibility Systems. I. The Plumbaginaceae. Evolution 1966, 20, 349–368. [Google Scholar] [PubMed]
- Moreno, J.; Terrones, A.; Juan, A. Germination Patterns along a Salinity Gradient of Closely-Related Halophytes in Sympatry. Estuar. Coast. Shelf Sci. 2022, 264, 107690. [Google Scholar] [CrossRef]
- Grigore, M.-N.; Toma, C. Anatomical Adaptations of Halophytes: A Review of Classic Literature and Recent Findings, 1st ed.; Springer Nature, Ed.; Springer: Cham, Switzerland, 2017; ISBN 978-3-319-66480-4. [Google Scholar]
- Mi, P.; Yuan, F.; Guo, J.; Han, G.; Wang, B. Salt Glands Play a Pivotal Role in the Salt Resistance of Four Recretohalophyte Limonium Mill. Species. Plant Biol. 2021, 23, 1063–1073. [Google Scholar] [CrossRef]
- Grieve, C.M.; Poss, J.A.; Grattan, S.; Shouse, P.J.; Lieth, J.H.; Zeng, L. Productivity and Mineral Nutrition of Limonium Species Irrigated with Saline Wastewaters. HortScience 2005, 40, 654–658. [Google Scholar] [CrossRef]
- Zia, S.; Egan, T.; Khan, M. Growth and Selective Ion Transport of Limonium stocksii Plumbaginacea under Saline Conditions. Pak. J. Bot. 2008, 40, 697–709. [Google Scholar]
- Al Hassan, M.; Estrelles, E.; Soriano, P.; López-Gresa, M.P.; Bellés, J.M.; Boscaiu, M.; Vicente, O. Unraveling Salt Tolerance Mechanisms in Halophytes: A Comparative Study on Four Mediterranean Limonium Species with Different Geographic Distribution Patterns. Front. Plant Sci. 2017, 8, 1438. [Google Scholar] [CrossRef] [Green Version]
- Li, Y. Kinetics of the Antioxidant Response to Salinity in the Halophyte Limonium Bicolor. Plant Soil Environ. 2008, 54, 493–497. [Google Scholar] [CrossRef]
- González-Orenga, S.; Grigore, M.N.; Boscaiu, M.; Vicente, O. Constitutive and Induced Salt Tolerance Mechanisms and Potential Uses of Limonium Mill. Species. Agronomy 2021, 11, 413. [Google Scholar] [CrossRef]
- González Orenga, S.; Al Hassan, M.; Llinares, J.; Lisón, P.; López-Gresa, M.P.; Verdeguer, M.; Vicente, O.; Boscaiu, M. Qualitative and Quantitative Differences in Osmolytes Accumulation and Antioxidant Activities in Response to Water Deficit in Four Mediterranean Limonium Species. Plants 2019, 8, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laguna, E.; Fos, S.; Ferrando-Pardo, I.; Ferrer-Gallego, P.P. Contents Endangered Halophytes and Their Conservation. Lessons from Eastern Spain, 1st ed.; Grigore, M.-N., Ed.; Springer: Cham, Switzerland, 2021; ISBN 9783030576356. [Google Scholar]
- Erben, M. Gattung Limonium Im Sudwestmediterranen Raum. Mitt. Bot. Staatssamml. München 1978, 14, 361–631. [Google Scholar]
- Moreno, J.; Terrones, A.; Juan, A.; Alonso, M.Á. Halophytic Plant Community Patterns in Mediterranean Saltmarshes: Shedding Light on the Connection between Abiotic Factors and the Distribution of Halophytes. Plant Soil 2018, 430, 185–204. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering Plant Memory: Intersections between Stress Recovery, RNA Turnover, and Epigenetics. Sci. Adv. 2022, 2, e1501340. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic Shifts in Ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress Memory and the Inevitable Effects of Drought: A Physiological Perspective. Front. Plant Sci. 2016, 7, 143. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.D.F.B.; Menezes-Silva, P.E.; Sousa, L.F.; Loram-Lourenço, L.; Silva, M.L.F.; Almeida, S.E.S.; Silva, F.G.; Perez de Souza, L.; Fernie, A.R.; Farnese, F.S. Evidence of Drought Memory in Dipteryx Alata Indicates Differential Acclimation of Plants to Savanna Conditions. Sci. Rep. 2020, 10, 16455. [Google Scholar] [CrossRef]
- Hamed, K.B.; Ellouzi, H.; Talbi, O.Z.; Hessini, K.; Slama, I.; Ghnaya, T.; Bosch, S.M.; Savouré, A.; Abdelly, C. Physiological Response of Halophytes to Multiple Stresses. Funct. Plant Biol. 2013, 40, 883–896. [Google Scholar] [CrossRef]
- Rasheed, A.; Koyro, H.-W.; Hameed, A.; Gul, B. Physiological Responses of the Xero-Halophyte Salsola Drummondii to Seasonal Alterations of Environmental Conditions in a Salt Desert. Ecol. Res. 2022, 37, 738–752. [Google Scholar] [CrossRef]
- Mir, R.; Romero, I.; González-Orenga, S.; Ferrer-Gallego, P.P.; Laguna, E.; Boscaiu, M.; Oprică, L.; Grigore, M.N.; Vicente, O. Constitutive and Adaptive Traits of Environmental Stress Tolerance in the Threatened Halophyte Limonium angustebracteatum Erben (Plumbaginaceae). Plants 2022, 11, 1137. [Google Scholar] [CrossRef] [PubMed]
- González-Orenga, S.; Donat-Torres, M.P.; Llinares, J.V.; Navarro, A.; Collado, F.; Ferrer-Gallego, P.P.; Laguna, E.; Vicente, O.; Boscaiu, M. Multidisciplinary Studies Supporting Conservation Programmes of Two Rare, Endangered Limonium Species from Spain. Plant Soil 2021, 466, 505–524. [Google Scholar] [CrossRef]
- Liu, X.; Duan, D.; Li, W.; Tadano, T.; Khan, M.A. A. A Comparative Study On Responses Of Growth And Solute Composition in Halophytes Suaeda Salsa and Limonium Bicolor to Salinity. In Ecophysiology of High Salinity Tolerant Plants; Springer: Dordrecht, The Netherlands, 2006; pp. 135–143. [Google Scholar] [CrossRef]
- Tabot, P.T.; Adams, J.B. Salt Secretion, Proline Accumulation and Increased Branching Confer Tolerance to Drought and Salinity in the Endemic Halophyte Limonium linifolium. S. Afr. J. Bot. 2014, 94, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Blanco, M.J.; Álvarez, S.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Root System Response to Drought and Salinity: Root Distribution and Water Transport. In Root Engineering; Asunción Morte, A.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 325–352. ISBN 9783642542763. [Google Scholar]
- Yang, Y.; Guo, Y.; Zhong, J.; Zhang, T.; Li, D.; Ba, T.; Xu, T.; Chang, L.; Zhang, Q.; Sun, M. Root Physiological Traits and Transcriptome Analyses Reveal That Root Zone Water Retention Confers Drought Tolerance to Opisthopappus taihangensis. Sci. Rep. 2020, 10, 2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, H.J.; Baek, D.; Cho, H.M.; Jung, H.S.; Jeong, M.S.; Jung, W.H.; Choi, C.W.; Lee, S.H.; Jin, B.J.; Park, M.S.; et al. Metabolic Adjustment of Arabidopsis Root Suspension Cells during Adaptation to Salt Stress and Mitotic Stress Memory. Plant Cell Physiol. 2019, 60, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Alegre, L. Changes in Carotenoids, Tocopherols and Diterpenes during Drought and Recovery, and the Biological Significance of Chlorophyll Loss in Rosmarinus officinalis Plants. Planta 2000, 210, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Molinari, H.B.C.; Marur, C.J.; Daros, E.; De Campos, M.K.F.; De Carvalho, J.F.R.P.; Filho, J.C.B.; Pereira, L.F.P.; Vieira, L.G.E. Evaluation of the Stress-Inducible Production of Proline in Transgenic Sugarcane (Saccharum Spp.): Osmotic Adjustment, Chlorophyll Fluorescence and Oxidative Stress. Physiol. Plant. 2007, 130, 218–229. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Virlouvet, L.; Fromm, M. Physiological and Transcriptional Memory in Guard Cells during Repetitive Dehydration Stress. New Phytol. 2015, 205, 596–607. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple Exposures to Drought ‘train’ transcriptional Responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef] [PubMed]
- Virlouvet, L.; Avenson, T.J.; Du, Q.; Zhang, C.; Liu, N.; Fromm, M.; Avramova, Z.; Russo, S.E. Dehydration Stress Memory: Gene Networks Linked to Physiological Responses during Repeated Stresses of Zea mays. Front. Plant Sci. 2018, 9, 1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calone, R.; Mircea, D.M.; González-Orenga, S.; Boscaiu, M.; Lambertini, C.; Barbanti, L.; Vicente, O. Recovery from Salinity and Drought Stress in the Perennial Sarcocornia fruticosa vs. the Annual Salicornia europaea and S. veneta. Plants 2022, 11, 1058. [Google Scholar] [CrossRef] [PubMed]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of Increased Seawater Salinity Irrigation on Growth and Quality of the Edible Halophyte Mesembryanthemum crystallinum L. under Field Conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium Signaling and Salt Tolerance Are Diversely Entwined in Plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef]
- Bachani, J.; Mahanty, A.; Aftab, T.; Kumar, K. Insight into Calcium Signalling in Salt Stress Response. S. Afr. J. Bot. 2022, 151, 1–8. [Google Scholar] [CrossRef]
- Shabbir, R.; Javed, T.; Hussain, S.; Ahmar, S.; Naz, M.; Zafar, H.; Pandey, S.; Chauhan, J.; Siddiqui, M.H.; Pinghua, C. Calcium Homeostasis and Potential Roles in Combatting Environmental Stresses in Plants. S. Afr. J. Bot. 2022, 148, 683–693. [Google Scholar] [CrossRef]
- Knight, H.; Brandt, S.; Knight, M.R. A History of Stress Alters Drought Calcium Signalling Pathways in Arabidopsis. Plant J. 1998, 16, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Polavarapu, K.K.B.; Nese, S. Is Proline Accumulation per Se Correlated with Stress Tolerance or Is Proline Homeostasis a More Critical Issue? Plant. Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and Its Actions during the Drought Stress in Plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Liu, L.; Cao, X.; Zhai, Z.; Ma, S.; Tian, Y.; Cheng, J. Direct Evidence of Drought Stress Memory in Mulberry from a Physiological Perspective: Antioxidative, Osmotic and Phytohormonal Regulations. Plant Physiol. Biochem. 2022, 186, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, H.; Wang, L.; Liu, H.; Huo, H.; Zhang, C.; Liu, A.; Zhu, A.; Hu, J.; Lin, Y.; et al. Physiological and Transcriptome Analyses Reveal Short-Term Responses and Formation of Memory Under Drought Stress in Rice. Front. Genet. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auler, P.A.; do Amaral, M.N.; Rodrigues, G.S.; Benitez, L.C.; da Maia, L.C.; Souza, G.M.; Braga, E.J.B. Molecular Responses to Recurrent Drought in Two Contrasting Rice Genotypes. Planta 2017, 246, 899–914. [Google Scholar] [CrossRef] [PubMed]
- Derbali, W.; Manaa, A.; Goussi, R.; Derbali, I.; Abdelly, C.; Koyro, H.W. Post-Stress Restorative Response of Two Quinoa Genotypes Differing in Their Salt Resistance after Salinity Release. Plant Physiol. Biochem. 2021, 164, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Mohsin, S.M.; Fujita, M. Comparative Physiological and Biochemical Changes in Tomato (Solanum lycopersicum L.) under Salt Stress and Recovery: Role of Antioxidant Defense and Glyoxalase Systems. Antioxidants 2019, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar Sensing and Signaling in Plants: Conserved and Novel Mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Ellouzi, H.; Ben Hamed, K.; Asensi-Fabado, M.A.; Müller, M.; Abdelly, C.; Munné-Bosch, S. Drought and Cadmium May Be as Effective as Salinity in Conferring Subsequent Salt Stress Tolerance in Cakile maritima. Planta 2013, 237, 1311–1323. [Google Scholar] [CrossRef]
- Carvalho, F.E.L.; Silveira, J.A.G. H2O2-Retrograde Signaling as a Pivotal Mechanism to Understand Priming and Cross Stress Tolerance in Plants; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128178935. [Google Scholar]
- Wahid, A.; Perveen, M.; Gelani, S.; Basra, S.M.A. Pretreatment of Seed with H2O2 Improves Salt Tolerance of Wheat Seedlings by Alleviation of Oxidative Damage and Expression of Stress Proteins. J. Plant Physiol. 2007, 164, 283–294. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Rolim Medeiros, J.-V.; Gomes-Filho, E. Hydrogen Peroxide Pre-Treatment Induces Salt-Stress Acclimation in Maize Plants. J. Plant Physiol. 2005, 162, 1114–1122. [Google Scholar] [CrossRef]
- Wojtyla, Ł.; Paluch-Lubawa, E.; Sobieszczuk-Nowicka, E.; Garnczarska, M. Drought Stress Memory and Subsequent Drought Stress Tolerance in Plants. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D.J., Fujita, M., Huang, B., Eds.; Academic Press: New York, NY, USA, 2020; pp. 115–131. ISBN 978-0-12-817892-8. [Google Scholar]
- Auler, P.A.; Souza, G.M.; da Silva Engela, M.R.G.; do Amaral, M.N.; Rossatto, T.; da Silva, M.G.Z.; Furlan, C.M.; Maserti, B.; Braga, E.J.B. Stress Memory of Physiological, Biochemical and Metabolomic Responses in Two Different Rice Genotypes under Drought Stress: The Scale Matters. Plant Sci. 2021, 311, 110994. [Google Scholar] [CrossRef]
- Virlouvet, L.; Ding, Y.; Fujii, H.; Avramova, Z.; Fromm, M. ABA Signaling Is Necessary but Not Sufficient for RD29B Transcriptional Memory during Successive Dehydration Stresses in Arabidopsis thaliana. Plant J. 2014, 79, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Fleta-Soriano, E.; Pintó-Marijuan, M.; Munné-Bosch, S. Evidence of Drought Stress Memory in the Facultative CAM, Aptenia Cordifolia: Possible Role of Phytohormones. PLoS ONE 2015, 10, e0135391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Kumar, P.; Verma, V.; Sharma, R.; Bhargava, B.; Irfan, M. Understanding Plant Stress Memory Response for Abiotic Stress Resilience: Molecular Insights and Prospects. Plant Physiol. Biochem. 2022, 179, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Wellbur, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Weimberg, R. Solute Adjustments in Leaves of Two Species of Wheat at Two Different Stages of Growth in Response to Salinity. Physiol. Plant. 1987, 70, 381–388. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Taulavuori, E.; Hellström, E.; Taulavuori, K.; Laine, K. Comparison of Two Methods Used to Analyse Lipid Peroxidation from Vaccinium myrtillus (L.) during Snow Removal, Reacclimation and Cold Acclimation. J. Exp. Bot. 2002, 52, 2375–2380. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Velikova, V. Isoprene Produced by Leaves Protects the Photosynthetic Apparatus against Ozone Damage, Quenches Ozone Products, and Reduces Lipid Peroxidation of Cellular Membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.; Mello, J. Application and Analysis of the Folin Ciocalteu Method for the Determination of the Total Phenolic Content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity Effects on Polyphenol Content and Antioxidant Activities in Leaves of the Halophyte Cakile Maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, O.F.; Shukla, M.K.; Stringam, B.; Picchioni, G.A.; Gard, C. Irrigation with Brackish Water Changes Evapotranspiration, Growth and Ion Uptake of Halophytes. Agric. Water Manag. 2018, 195, 142–153. [Google Scholar] [CrossRef]
- Heidari-Sharifabad, H.; Mirzaie-Nodoushan, H. Salinity-Induced Growth and Some Metabolic Changes in Three Salsola Species. J. Arid Environ. 2006, 67, 715–720. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications, Inc.: Newbury Park, CA, USA, 2019. [Google Scholar]
- Lenth, R.V.; Buerkner, P.; Herve, M.; Love, J.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, Aka LeastSquares Means. R Pack. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 12 December 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis. Available online: https://ggplot2.tidyverse.org/reference/ (accessed on 12 December 2021).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T0 | First Stress (T30) | Recovery (T45) | Second Stress (T60) | |
|---|---|---|---|---|
| Control | ||||
| total leaf no. | 14 ± 1.20 aA | 17 ± 1.40 aA | 19 ± 1.96 aA | 19 ± 1.64 aA |
| wilted leaf % | 0 C | 13 ± 2.70 bB | 22 ± 2.68 aA | 21 ± 2.39 abAB |
| SLA (cm2 g−1) | 20.78 ± 3.59 aA | 27.11 ± 3.08 aA | 25.12 ± 5.59 aA | |
| 0.5 M NaCl | ||||
| total leaf no. | 14 ± 1.21 aA | 19 ± 2.09 aA | 21 ± 3.30 aA | 20 ± 3.43 aA |
| wilted leaf % | 0 B | 32 ± 6.80 aA | 22 ± 2.94 aB | 26 ± 2.67 aA |
| SLA (cm2 g−1) | 37.14 ± 13.97 aA | 25.46 ± 4.38 aA | 31.17 ± 7.66 aA | |
| 1 M NaCl | ||||
| total leaf no. | 13 ± 0.96 aB | 16 ± 0.97 aA | 15 ± 1.32 aA | 28 ± 7.73 aA |
| wilted leaf % | 0 C | 41 ± 1.92 aA | 38 ± 1.70 abA | 19 ± 1.65 bA |
| SLA (cm2 g−1) | 16.90 ± 3.15 aA | 25.37 ± 4.14 aA | 52.89 ± 18.08 aA | |
| Water Stress | ||||
| total leaf no. | 13 ± 1.02 Aa | 16 ± 0.83 aA | 16 ± 2.27 aA | 18 ± 2.84 aA |
| wilted leaf % | 0 B | 38 ± 4.88 aA | 50 ± 12.21 aA | 26 ± 2.97 aA |
| SLA (cm2 g−1) | 25.48 ± 8.21 aA | 39.56 ± 9.43 aA | 36.90 ± 4.60 aA |
| Control | 0.5 M NaCl | 1 M NaCl | Water Stress | |
|---|---|---|---|---|
| First stress—30 days | 1.17 | 1.17 | 1.17 | 0 |
| Recovery—15 days | 0.65 | 1.5 | 1.5 | 1.25 |
| Second stress—15 days | 0.65 | 0.65 | 0.65 | 0 |
| Total (L pot−1) | 2.47 | 3.32 | 3.32 | 1.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calone, R.; Mircea, D.-M.; González-Orenga, S.; Boscaiu, M.; Zuzunaga-Rosas, J.; Barbanti, L.; Vicente, O. Effect of Recurrent Salt and Drought Stress Treatments on the Endangered Halophyte Limonium angustebracteatum Erben. Plants 2023, 12, 191. https://doi.org/10.3390/plants12010191
Calone R, Mircea D-M, González-Orenga S, Boscaiu M, Zuzunaga-Rosas J, Barbanti L, Vicente O. Effect of Recurrent Salt and Drought Stress Treatments on the Endangered Halophyte Limonium angustebracteatum Erben. Plants. 2023; 12(1):191. https://doi.org/10.3390/plants12010191
Chicago/Turabian StyleCalone, Roberta, Diana-Maria Mircea, Sara González-Orenga, Monica Boscaiu, Javier Zuzunaga-Rosas, Lorenzo Barbanti, and Oscar Vicente. 2023. "Effect of Recurrent Salt and Drought Stress Treatments on the Endangered Halophyte Limonium angustebracteatum Erben" Plants 12, no. 1: 191. https://doi.org/10.3390/plants12010191