


Enhancing the Production of Hydrolytic Enzymes in Elicited Tamarillo (Solanum betaceum Cav.) Cell Suspension Cultures


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
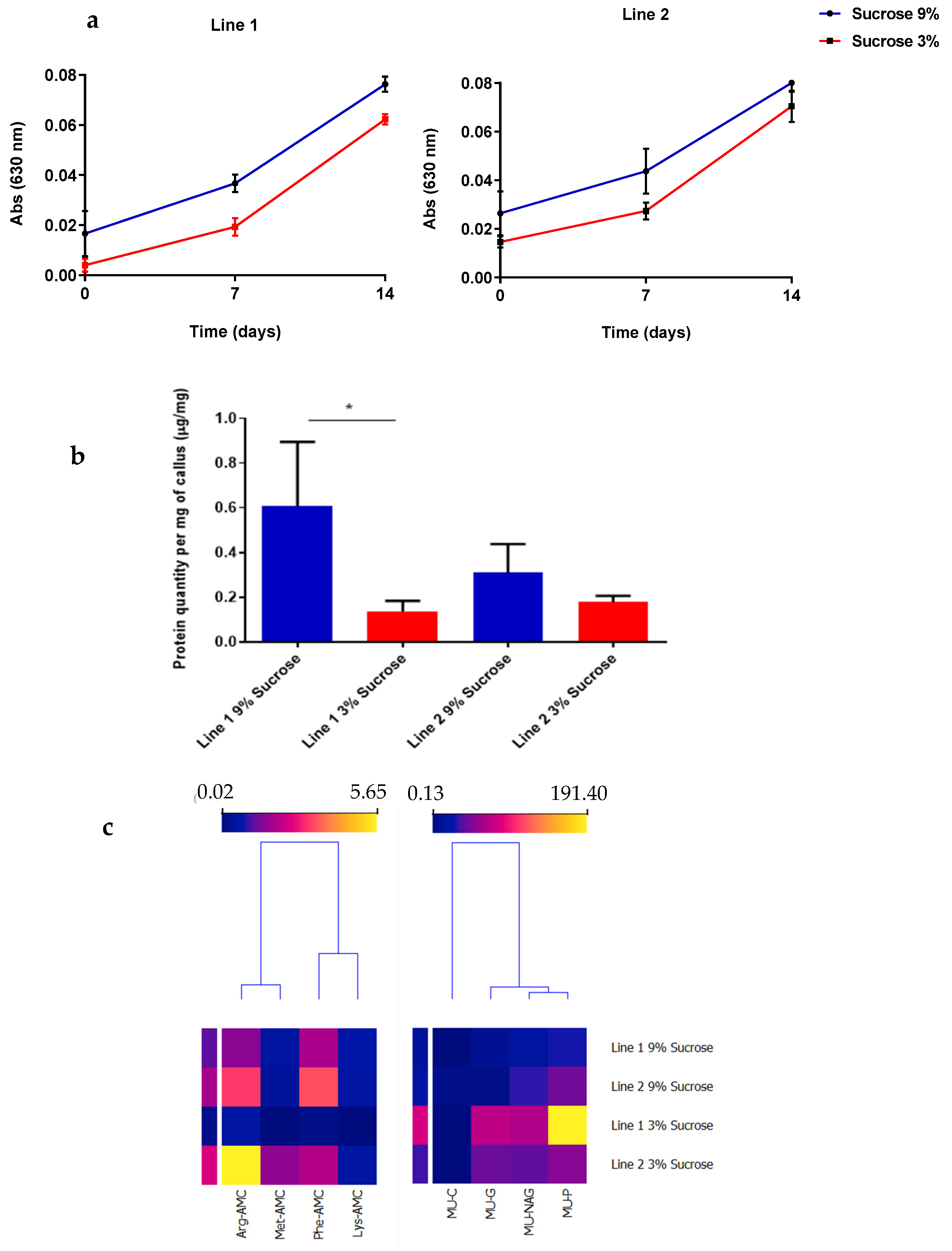
2.1. Cell Suspension Cultures Growth and Enzymatic Activity under Different Sucrose Concentrations
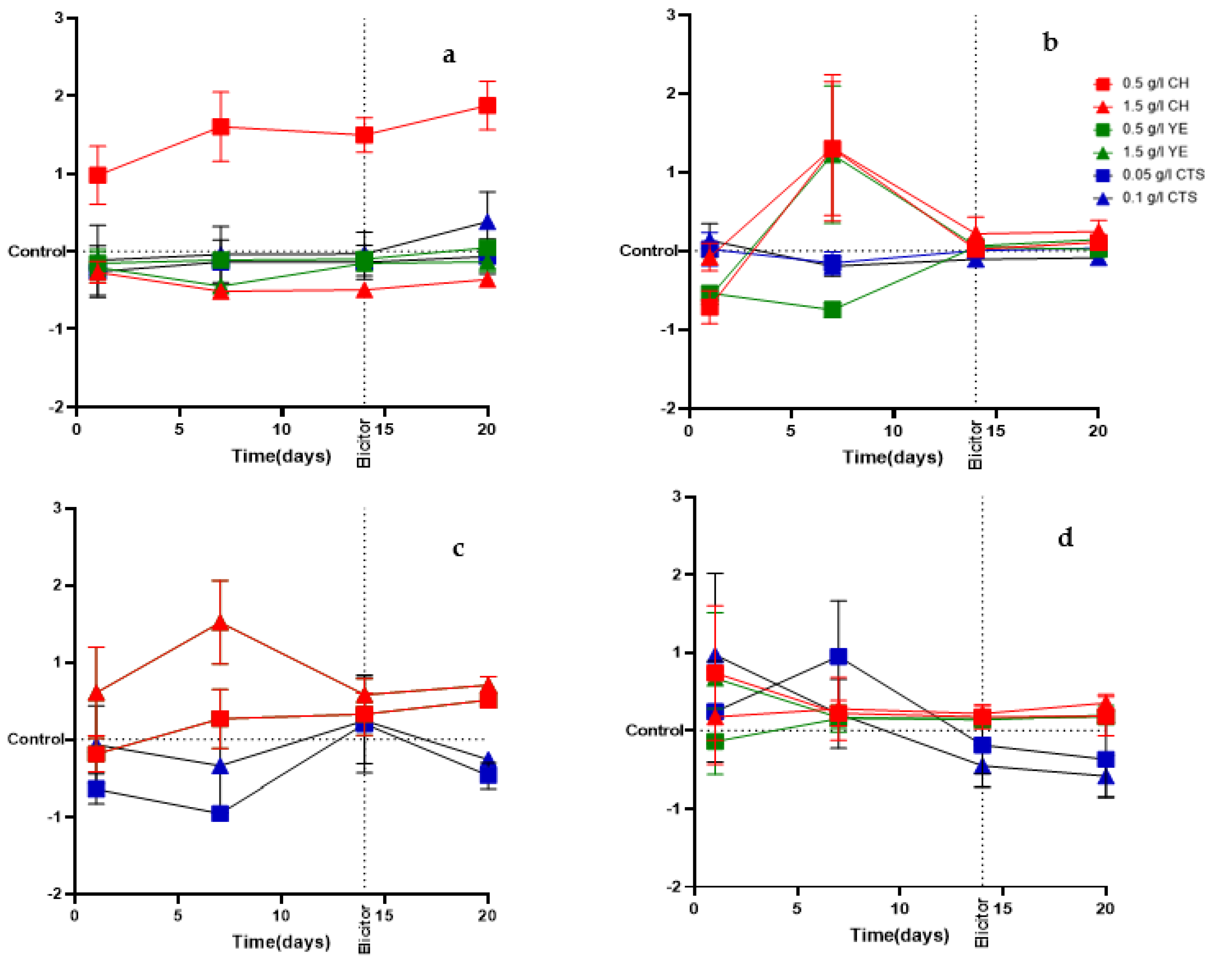
2.2. Effect of Elicitors on the Growth of Cells Suspension Cultures
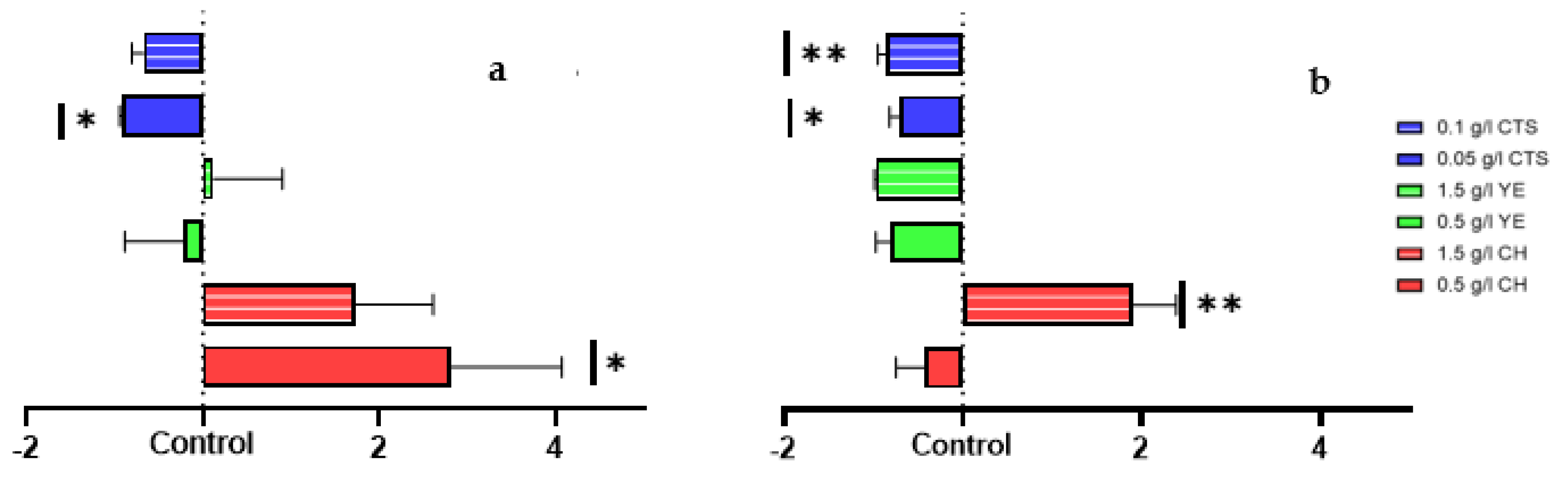
2.3. Elicited Cell Suspension Total Protein Production
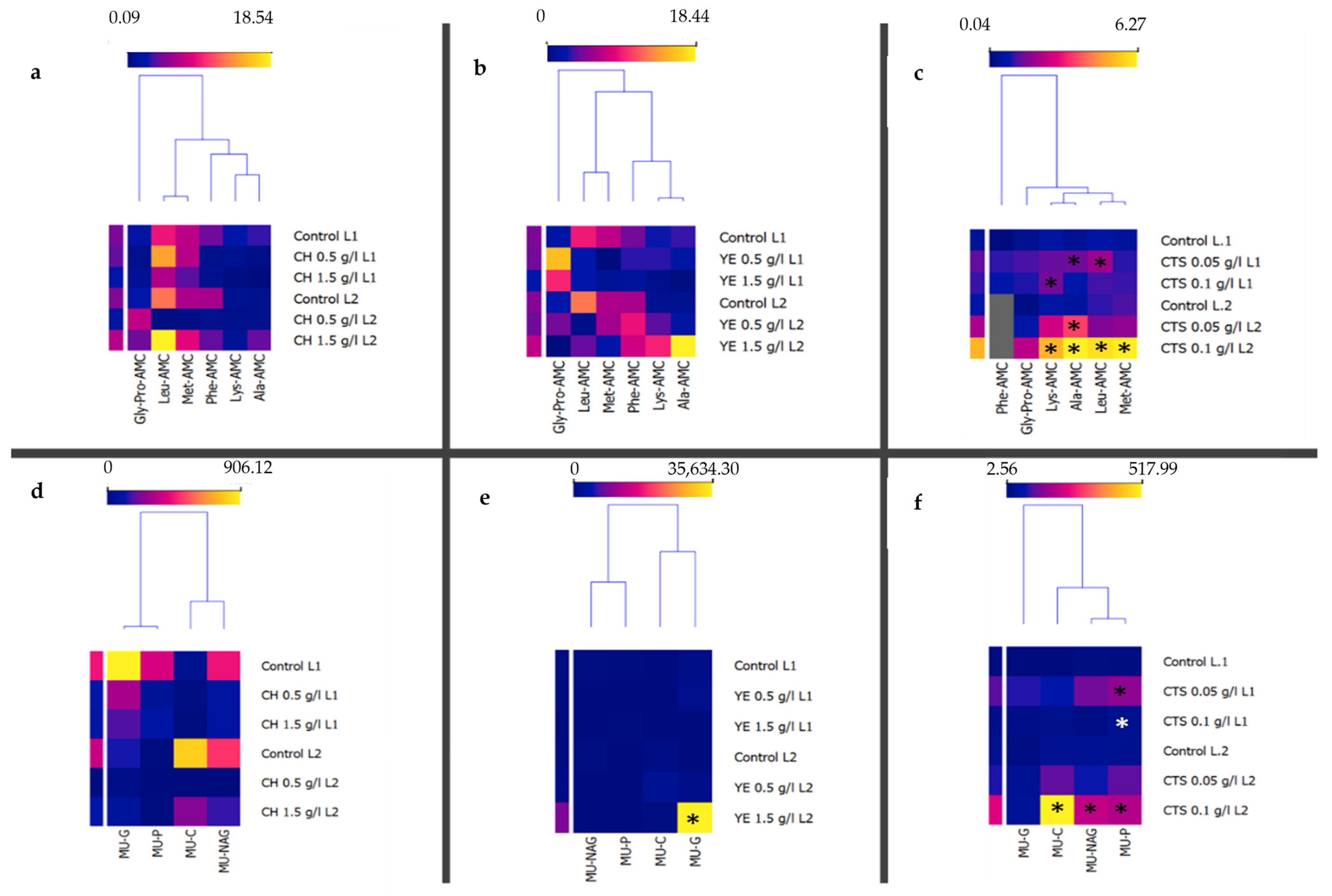
2.4. Specific Enzymatic Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.1.1. Callus Induction and Maintenance
4.1.2. Establishment of Cell Suspension Cultures and Growth Kinetics
4.1.3. Effect of Biotic Elicitors on Cell Growth
4.2. Protein Extraction
4.3. Protein Quantification
4.4. Enzymatic Activity Assays
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Moon, K.-B.; Park, J.-S.; Park, Y.-I.; Song, I.-J.; Lee, H.-J.; Cho, H.S.; Jeon, J.-H.; Kim, H.-S. Development of systems for the production of plant-derived biopharmaceuticals. Plants 2019, 9, 30. [Google Scholar] [CrossRef] [Green Version]
- Tusé, D.; Nandi, S.; McDonald, K.A.; Buyel, J.F. The emergency response capacity of plant-based biopharmaceutical manufacturing-what it is and what it could be. Front. Plant Sci. 2020, 11, 1573. [Google Scholar] [CrossRef]
- Lindsay, P.; Ackerman, A.; Jian, Y.; Artz, O.; Rosado, D.; Skopelitis, T.; Kitagawa, M.; Pedmale, U.V.; Jackson, D. Rapid Expression of COVID-19 proteins by transient expression in tobacco. Author Contributions. bioRxiv 2020, 1, 1–17. [Google Scholar] [CrossRef]
- Hellwig, S.; Drossard, J.; Twyman, R.M.; Fischer, R. Plant cell cultures for the production of recombinant proteins. Nat. Biotechnol. 2004, 22, 1415–1422. [Google Scholar] [CrossRef]
- Xu, J.; Ge, X.; Dolan, M.C. Towards high-yield production of pharmaceutical proteins with plant cell suspension cultures. Biotechnol. Adv. 2011, 29, 278–299. [Google Scholar] [CrossRef]
- Bosch, D.; Castilho, A.; Loos, A.; Schots, A.; Steinkellner, H. N-glycosylation of plant-produced recombinant proteins. Curr. Pharm. Des. 2013, 19, 5503–5512. [Google Scholar] [CrossRef]
- Santos, R.B.; Abranches, R.; Fischer, R.; Sack, M.; Holland, T. Putting the spotlight back on plant suspension cultures. Front. Plant Sci. 2016, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Reuter, L.J.; Bailey, M.J.; Joensuu, J.J.; Ritala, A. Scale-up of hydrophobin-assisted recombinant protein production in tobacco BY-2 suspension cells. Plant Biotechnol. J. 2014, 12, 402–410. [Google Scholar] [CrossRef]
- Martin, D.; Lopes, T.; Correia, S.; Canhoto, J.; Marques, M.P.M.; Batista de Carvalho, L.A.E. Nutraceutical properties of tamarillo fruits: A vibrational study. Spectrochim. Acta-Part A Mol. Biomol. Spectrosc. 2021, 252, 119501. [Google Scholar] [CrossRef]
- Correia, S.; Canhoto, J.M. Somatic Embryogenesis of Tamarillo (Solanum betaceum Cav.); Jain, S., Gupta, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 171–179. [Google Scholar]
- Correia, S.; Vinhas, R.; Manadas, B.; Lourenço, A.S.; Veríssimo, P.; Canhoto, J.M. Comparative proteomic analysis of auxin-induced embryogenic and nonembryogenic tissues of the solanaceous tree Chyphomandra betacea (Tamarillo). J. Proteome Res. 2012, 11, 1666–1675. [Google Scholar] [CrossRef]
- Correia, S.; Alhinho, A.T.; Casimiro, B.; Miguel, C.M.; Oliveira, M.; Veríssimo, P.; Canhoto, J. NEP-TC a rRNA methyltransferase involved on somatic embryogenesis of tamarillo (Solanum betaceum Cav.). Front. Plant Sci. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Caeiro, A.; Correia, S.I.; Veríssimo, P.; Canhoto, J. Establishment and biochemical characterization of tamarillo (Solanum betaceum Cav.) embryogenic cell suspension cultures. Vitr. Cell. Dev. Biol.-Plant 2017, 53, 606–618. [Google Scholar] [CrossRef]
- Chavanke, S.N.; Penna, S.; Dalvi, S.G. β-glucan and its nanocomposites in sustainable agriculture and environment: An overview of mechanisms and applications. Environ. Sci. Pollut. Res. 2022, 29, 80062–80087. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef]
- Caseiro, C.; Dias, J.N.R.; de Andrade Fontes, C.M.G.; Bule, P. From cancer therapy to winemaking: The molecular structure and applications of β-glucans and β-1, 3-glucanases. Int. J. Mol. Sci. 2022, 23, 3156. [Google Scholar] [CrossRef]
- Sandhu, J.S.; Sidhu, M.K.; Yadav, I.S. Control of fungal diseases in agricultural crops by chitinase and glucanase transgenes. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2017; Volume 22, pp. 163–212. [Google Scholar] [CrossRef]
- Javed, S.; Ahmad, M.; Ahmad, M.; Abdin, M.; Hamid, R.; Khan, M.; Musarrat, J. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21. [Google Scholar] [CrossRef]
- Moravčíková, J.; Libantová, J.; Heldák, J.; Salaj, J.; Bauer, M.; Matušíková, I.; Gálová, Z.; Mlynárová, Ľ. Stress-induced expression of cucumber chitinase and Nicotiana plumbaginifolia β-1,3-glucanase genes in transgenic potato plants. Acta Physiol. Plant. 2007, 29, 133–141. [Google Scholar] [CrossRef]
- Mamo, J.; Assefa, F. The role of microbial aspartic protease enzyme in food and beverage industries. J. Food Qual. 2018, 2018, 7957269. [Google Scholar] [CrossRef] [Green Version]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Bhaskar, R.; Xavier, L.S.E.; Udayakumaran, G.; Kumar, D.S.; Venkatesh, R.; Nagella, P. Biotic elicitors:a boon for the in-vitro production of plant secondary metabolites. Plant Cell Tissue Organ Cult. 2022, 149, 7–24. [Google Scholar] [CrossRef]
- Boller, T. Chemoperception of microbial signals in plant cells. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 189–214. [Google Scholar] [CrossRef]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Bera, T. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780128024980. [Google Scholar]
- Malerba, M.; Cerana, R. Chitosan effects on plant systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, M.; Yadav, G.; Sonigra, P.; Nagda, A.; Mehta, T.; Swapnil, P.; Harish; Marwal, A. Role of elicitors to initiate the induction of systemic resistance in plants to biotic stress. Plant Stress 2022, 5, 100103. [Google Scholar] [CrossRef]
- Ahmad, Z.; Shahzad, A.; Sharma, S. Chitosan versus yeast extract driven elicitation for enhanced production of fragrant compound 2-hydroxy-4-methoxybenzaldehyde (2H4MB) in root tuber derived callus of Decalepis salicifolia (Bedd. Ex Hook.f.) Venter. Plant Cell. Tissue Organ Cult. 2019, 136, 29–40. [Google Scholar] [CrossRef]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Curr. Res. Microb. Sci. 2021, 2, 100054. [Google Scholar] [CrossRef]
- Iula, G.; Miras-Moreno, B.; Rouphael, Y.; Lucini, L.; Trevisan, M. The complex metabolomics crosstalk triggered by four molecular elicitors in tomato. Plants 2022, 11, 678. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, P.; Miras-Moreno, B.; Lucini, L.; Gallego, P.P. The metabolomics reveals intraspecies variability of bioactive compounds in elicited suspension cell cultures of three Bryophyllum species. Ind. Crops Prod. 2021, 163, 113322. [Google Scholar] [CrossRef]
- Curtis, W.R.; Emery, A.H. Plant cell suspension culture rheology. Biotechnol. Bioeng. 1993, 42, 520–526. [Google Scholar] [CrossRef]
- Huang, T.-K.; McDonald, K.A. Bioreactor engineering for recombinant protein production in plant cell suspension cultures. Biochem. Eng. J. 2009, 45, 168–184. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M.; Ali, S. Sucrose induced osmotic stress and photoperiod regimes enhanced the biomass and production of antioxidant secondary metabolites in shake-flask suspension cultures of Prunella vulgaris L. Plant Cell. Tissue Organ Cult. 2016, 124, 573–581. [Google Scholar] [CrossRef]
- Kim, S.I.; Choi, H.K.; Kim, J.H.; Lee, H.S.; Hong, S.S. Effect of osmotic pressure on paclitaxel production in suspension cell cultures of Taxus chinensis. Enzyme Microb. Technol. 2001, 28, 202–209. [Google Scholar] [CrossRef]
- Narayan, M.S.; Venkataraman, L.V. Effect of sugar and nitrogen on the production of anthocyanin in cultured carrot (Daucus carota) Cells. J. Food Sci. 2002, 67, 84–86. [Google Scholar] [CrossRef]
- Lee, E.J.; Mobin, M.; Hahn, E.J.; Paek, K.Y. Effects of sucrose, inoculum density, auxins, and aeration volume on cell growth of Gymnema sylvestre. J. Plant Biol. 2006, 49, 427–431. [Google Scholar] [CrossRef]
- Dantas, L.A.; Faria, P.S.A.; Dário, B.M.M.; Arantes, A.L.M.; Silva, F.G.; Avila, R.G.; Pereira, P.S.; Neto, A.R. The impact of carbon source on cell growth and the production of bioactive compounds in cell suspensions of Hancornia speciosa Gomes. Sci. Rep. 2021, 11, 24315. [Google Scholar] [CrossRef]
- Malik, S.; Cusidó, R.M.; Mirjalili, M.H.; Moyano, E.; Palazón, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Process Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Tanaka, H.; Aoyagi, H.; Jitsufuchi, T. Turbidimetric measurement of cell biomass of plant cell suspensions. J. Ferment. Bioeng. 1992, 73, 130–134. [Google Scholar] [CrossRef]
- Salehi, M.; Karimzadeh, G.; Naghavi, M.R. Synergistic effect of coronatine and sorbitol on artemisinin production in cell suspension culture of Artemisia annua L. Cv. Anamed. Plant Cell. Tissue Organ Cult. 2019, 137, 587–597. [Google Scholar] [CrossRef]
- Khaleda, L.; Al-Forkan, M. Stimulatory Effects of casein hydrolysate and proline in in vitro callus induction and plant regeneration from five deep water rice (Oryza sativa L.). Biotechnology 2006, 5, 379–384. [Google Scholar] [CrossRef]
- Salehi, M.; Moieni, A.; Safaie, N. A novel medium for enhancing callus growth of hazel (Corylus avellana L.). Sci. Rep. 2017, 7, 15598. [Google Scholar] [CrossRef] [Green Version]
- Bayraktar, M.; Naziri, E.; Akgun, I.H.; Karabey, F.; Ilhan, E.; Akyol, B.; Bedir, E.; Gurel, A. Elicitor induced stevioside production, in vitro shoot growth, and biomass accumulation in micropropagated Stevia rebaudiana. Plant Cell. Tissue Organ Cult. 2016, 127, 289–300. [Google Scholar] [CrossRef]
- Krstić-Milošević, D.; Janković, T.; Uzelac, B.; Vinterhalter, D.; Vinterhalter, B. Effect of elicitors on xanthone accumulation and biomass production in hairy root cultures of Gentiana dinarica. Plant Cell. Tissue Organ Cult. 2017, 130, 631–640. [Google Scholar] [CrossRef]
- Malerba, M.; Crosti, P.; Cerana, R. Defense/stress responses activated by chitosan in sycamore cultured cells. Protoplasma 2012, 249, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Fernandez, M.; Marhuenda-Egea, F.C.; Lopez-Moya, F.; Arnao, M.B.; Cabrera-Escribano, F.; Nueda, M.J.; Gunsé, B.; Lopez-Llorca, L.V. Chitosan induces plant hormones and defenses in tomato root exudates. Front. Plant Sci. 2020, 11, 1677. [Google Scholar] [CrossRef] [PubMed]
- Vasconsuelo, A.; Boland, R. Molecular aspects of the early stages of elicitation of secondary metabolites in plants. Plant Sci. 2007, 172, 861–875. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, J.K.; Park, S.U. Yeast extract improved biosynthesis of astragalosides in hairy root cultures of Astragalus membranaceus. Prep. Biochem. Biotechnol. 2021, 51, 467–474. [Google Scholar] [CrossRef]
- Trong, T.T.; Truong, D.H.; Nguyen, H.C.; Tran, D.T.; Nguyen Thi, H.T.; Dang, G.D.; Huu, H.N. Biomass accumulation of Panax vietnamensis in cell suspension cultures varies with addition of plant growth regulators and organic additives. Asian Pac. J. Trop. Med. 2017, 10, 907–915. [Google Scholar] [CrossRef]
- Kamalipourazad, M.; Sharifi, M.; Maivan, H.Z.; Behmanesh, M.; Chashmi, N.A. Induction of aromatic amino acids and phenylpropanoid compounds in Scrophularia striata Boiss. Cell Culture in Response to Chitosan-Induced Oxidative Stress. Plant Physiol. Biochem. 2016, 107, 374–384. [Google Scholar] [CrossRef]
- Fooladi Vanda, G.; Shabani, L.; Razavizadeh, R. Chitosan enhances rosmarinic acid production in shoot cultures of Melissa officinalis L. through the Induction of Methyl Jasmonate. Bot. Stud. 2019, 60, 26. [Google Scholar] [CrossRef]
- Katiyar, D.; Hemantaranjan, A.; Singh, B. Chitosan as a promising natural compound to enhance potential physiological responses in plant: A Review. Indian J. Plant Physiol. 2015, 20, 1–9. [Google Scholar] [CrossRef]
- Stasińska-Jakubas, M.; Hawrylak-Nowak, B. Protective, biostimulating, and eliciting effects of chitosan and its derivatives on crop plants. Molecules 2022, 27, 2801. [Google Scholar] [CrossRef] [PubMed]
- Dalvi, S.; Suprasanna, P. Studies on radiation processed polymers mediated biochemical and molecular responses and relevance to enhancing plant productivity. In Radiation-Processed Polysaccharides; Elsevier: Amsterdam, The Netherlands, 2022; pp. 355–367. ISBN 9780323856720. [Google Scholar]
- Kaku, H.; Shibuya, N.; Xu, P.; Aryan, A.P.; Fincher, G.B. N-acetylchitooligosaccharides elicit expression of a single (13)-beta-glucanase gene in suspension-cultured cells from barley (Hordeum vulgare). Physiol. Plant. 1997, 100, 111–118. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Baig, M.M.V. Biotic elicitor enhanced production of psoralen in suspension cultures of Psoralea corylifolia L. Saudi J. Biol. Sci. 2014, 21, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy-Hernández, G.; Vázquez-Flota, F.A. Growth measurements: Estimation of cell division and cell expansion. Methods Mol. Biol. 2006, 318, 51–58. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gee, K.R.; Sun, W.-C.; Bhalgat, M.K.; Upson, R.H.; Klaubert, D.H.; Latham, K.A.; Haugland, R.P. Fluorogenic substrates based on fluorinated umbelliferones for continuous assays of phosphatases and β-galactosidases. Anal. Biochem. 1999, 273, 41–48. [Google Scholar] [CrossRef]
- Harris, J.L.; Backes, B.J.; Leonetti, F.; Mahrus, S.; Ellman, J.A.; Craik, C.S. Rapid and general profiling of protease specificity by using combinatorial fluorogenic substrate libraries. Proc. Natl. Acad. Sci. USA 2000, 97, 7754–7759. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casimiro, B.; Mota, I.; Veríssimo, P.; Canhoto, J.; Correia, S. Enhancing the Production of Hydrolytic Enzymes in Elicited Tamarillo (Solanum betaceum Cav.) Cell Suspension Cultures. Plants 2023, 12, 190. https://doi.org/10.3390/plants12010190
Casimiro B, Mota I, Veríssimo P, Canhoto J, Correia S. Enhancing the Production of Hydrolytic Enzymes in Elicited Tamarillo (Solanum betaceum Cav.) Cell Suspension Cultures. Plants. 2023; 12(1):190. https://doi.org/10.3390/plants12010190
Chicago/Turabian StyleCasimiro, Bruno, Inês Mota, Paula Veríssimo, Jorge Canhoto, and Sandra Correia. 2023. "Enhancing the Production of Hydrolytic Enzymes in Elicited Tamarillo (Solanum betaceum Cav.) Cell Suspension Cultures" Plants 12, no. 1: 190. https://doi.org/10.3390/plants12010190