1. Introduction
Micronutrients are essential elements for all plants and are required in much smaller amounts than macronutrients [
1,
2]. Multiple experiments have proven the beneficial effects of micronutrients on plant growth and yield [
2,
3,
4]. Plants show different needs for certain micronutrients, but the essential elements for all higher plants are boron (B), chloride (Cl), copper (Cu), iron (Fe), manganese (Mn), molybdenum (Mo), nickel (Ni), and zinc (Zn) [
1]. Micronutrients are involved in virtually all cellular and metabolic functions, such as primary and secondary metabolism, energy metabolism, cell protection, signal transduction, gene regulation, plant defence system, hormone perception, and reproduction [
1,
5,
6,
7,
8,
9]. It should be stressed, however, that when concentrations of the above-mentioned ions are too high, there is the formation of reactive oxygen species, with detrimental consequences to the cells [
1,
6]. Moreover, micronutrients, most notably Cu, Fe, Mn, and Zn [
10], are of great importance not only for plant health but also for animal and human health as key food components that are relevant for the metabolism and maintenance of tissue function [
11].
Several environmental and agronomic factors can impact the availability and uptake of plant nutrients even in nutrient-rich soils, compromising fruit quality and yield [
12,
13,
14]. The deficit in specific micronutrients, such as Fe or Mn, is a global problem [
15,
16]. Certain micronutrients present a challenge, partly because they have narrow dynamic ranges between the minimum requirement and toxicity level [
17,
18] and partly because their concentrations are subject to major fluctuations in the soil [
19]. Thus, balanced plant nutrition, combined with the knowledge of nutrient interactions, is a constantly relevant issue for sustainable and healthy crop growth [
20].
The task of maintaining the right supply of nutrients to plants is especially challenging and important in the case of nitrogen (N), which is in high demand due to its influence on crop yield, but it is also highly mobile [
21,
22,
23]. Given the large amount of N fertilizers used, which impacts plant growth [
23] and the pH of the rhizosphere [
22], additional N supply is expected to influence the uptake of other nutrients, especially micronutrients. For example, a strong correlation between different N levels and micronutrients was discovered in the case of tobacco plants [
12]. These relationships can be synergistic or antagonistic in nature [
12,
24,
25]. Moreover, other factors including weather, soil quality traits, plant species, and variety can also affect nutrient interactions and finally their uptake [
14,
24,
26]. In-depth knowledge of nutrient interactions and the factors accompanying these interactions is essential in order to better understand and control balanced plant feeding as well as improve nutrient use efficiency [
20]. Previous research showed that an increasing N soil supply leads to a rise not only in N but also Mg and Ca concentrations in kiwiberry (
Actinidia arguta [Siebold et Zucc.] Planch. ex Miq.) leaves, while P and K concentrations decrease [
27].
In relation to micronutrients, it has previously been reported that increasing the N supply has a negative effect on Zn and B concentrations in kiwifruit (
Actinidia chinensis var. deliciosa [A.Chev.] A. Chev.) [
28]. To the best of the authors’ knowledge, there have been no other studies on this topic. This report is intended to supplement data on the relationship between an increasing soil N supply and the leaf/fruit Cu, Zn, Mn, Fe, B, and Fe statuses. The analyzed micronutrients are among those that are most frequently deficient in soils, which is usually reflected in the plant’s internal status and external appearance. In turn, based on research and field experiments carried out in Europe and the USA, the kiwiberry plant shows potential to be a future commercial success [
29,
30,
31]. In brief,
A. arguta is the most frost-tolerant of all commercial
Actinidia species: the plants are resistant to biotic stresses, and the fruit are rich in health-promoting compounds and minerals [
32,
33]. Current practices in kiwiberry nutrition management mostly try to replicate the practices used in the cultivation of kiwifruit, which may have differing nutritional needs [
30]. The first steps to set up leaf mineral sufficiency ranges for kiwiberry plants have recently been taken [
34]. New information on the pattern of nutrient changes through two growing seasons in relation to shoot types in male and female plants has been described by Vance and Strik [
34]. However, there are still many knowledge gaps concerning nutritional requirements and especially balanced nutrition for plants.
In light of the above, the aims of this study were: (i) to assess the fluctuation in micronutrients in A. arguta leaves during different growing seasons, which may help indicate the optimum time for leaf sampling for diagnostic purposes, and (ii) to show the relationship between increasing soil N levels and micronutrient accumulation in kiwiberry leaves and fruit. The two A. arguta cultivars, ‘Weiki’ and ‘Geneva’, were selected and tested regarding the above-mentioned issues in 2015 and 2016.
3. Materials and Methods
The results presented in this paper are a supplement to the previously published study on leaf macronutrient status in relation to an increasing soil N level, cultivar, and sampling time [
27], and the experiment design has already been described in detail. Here, therefore, the experimental data are presented as concisely as possible. For a better understanding of the results presented in this manuscript, a short summary of the influence of increasing soil N levels on leaf N content depending on the cultivar in consecutive growing seasons is presented as a background (
Table 1).
3.1. Plant Material, Weather, and Growth Conditions
Tested samples (soil, leaf, and fruit tissues) were collected in 2015 and 2016 from the commercial A. arguta growing orchard (Bodzew, Poland; 51°47′49.9″ N + 20°48′44.0″ E) on a farm that was a partner in the supported project. The female plant tissues were sampled.
Weather data at the experimental site were collected with a field weather station (Vantage Pro 7, Davis Instruments, Hayward, CA, USA). Briefly, compared to the long-term average (1982–2012), the mean monthly temperatures in the examined growing seasons (March–September) were 1.8 and 1.6 °C higher in 2015 and 2016, respectively. In turn, the sums of total precipitation compared to the long-term average were lower by 26% and 6% for 2015 and 2016, respectively. Therefore, 2015 was clearly warmer and drier not only compared to the long-term average but also compared to 2016. More detailed weather data were recently published [
36]. Plants of two kiwiberry cultivars, ‘Weiki’ and ‘Geneva’, which are currently grown in several European countries as cultivars recommended for commercial cultivation [
29], were selected for the study. The plants were planted in a 4 × 4 m spacing in 2011 and trained on a T-bar support. The total male-to-female ratio on the plantation was 1:7. The experiment was set up in a randomized block design. The experiment consisted of three replications where four female plants per repetition were similarly located in relation to male plants. Soil samples were collected in early to mid-March. The N levels under study were 30, 50, and 80 mg N kg
−1 dry soil weight (DW) and were marked as N1, N2, and N3 respectively. The N doses for particular N treatments were calculated based on: (i) the available early spring soil N content (the N content in the soil was subtracted from the respective contents/levels under study), (ii) a topsoil depth of 0.2 m, and (iii) a soil density of 1.5 kg dm
3 on average. Plots were fertilized in three equal doses at the beginning of April, and then four and eight weeks later, using NH
4NO
3 containing 34% N. For detailed ranges of the added N as well as other essential nutrients and the pH kept, please refer to the previous paper [
27].
Soil macronutrients were extracted using 0.03 M CH
3COOH, while in the case of micronutrients, the Lindsey solution was used. The nutrient measurements were then conducted according to Komosa et al. [
41].
3.2. Leaf and Fruit Collection and Analysis
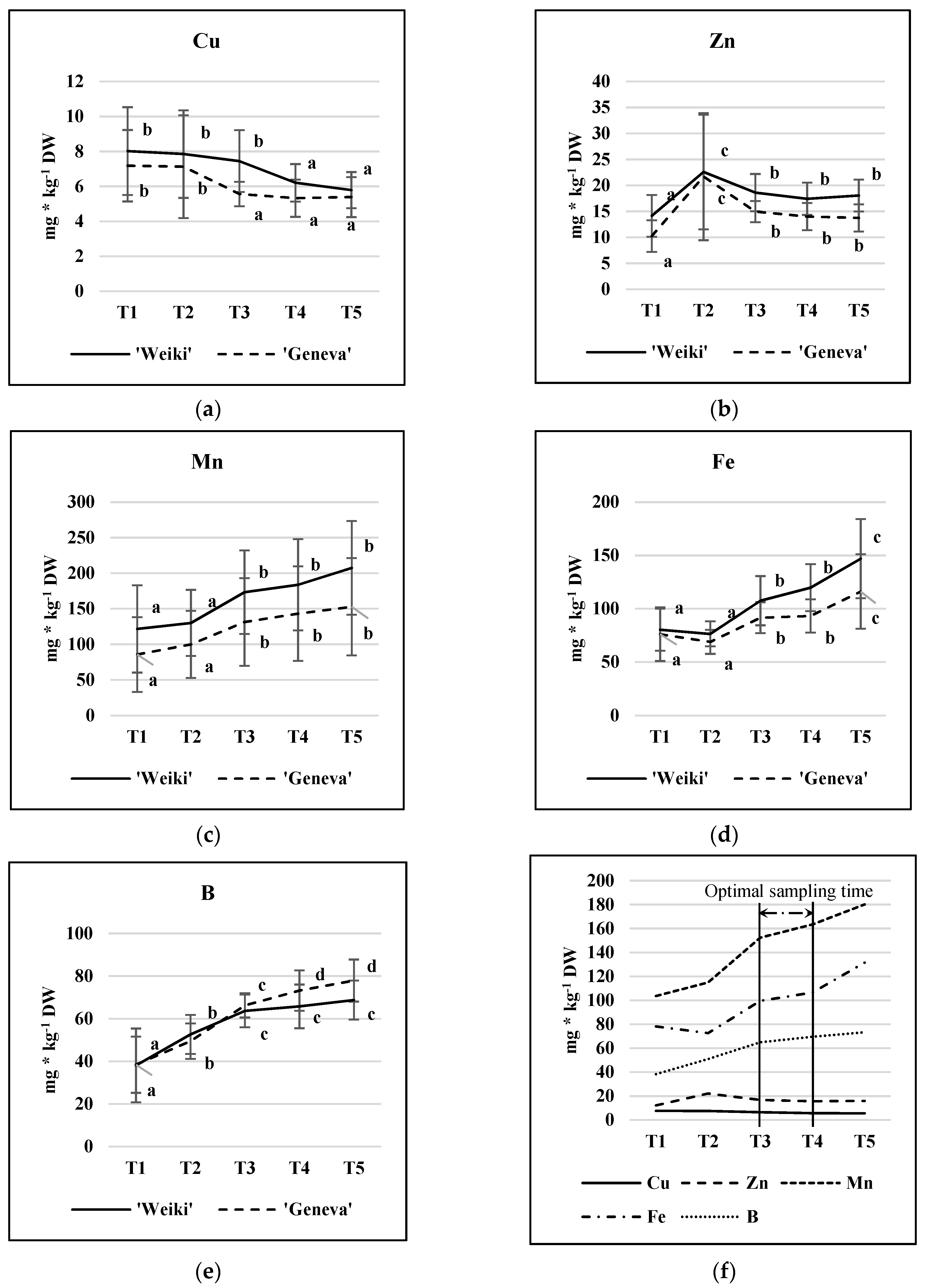
Leaf samples were collected at five time points during the growing season, starting on 28 May in 2015 and on 2 June in 2016 and continuing every three weeks until the end of August, with the time points being marked as T1, T2, T3, T4, and T5, respectively. For each point time, three repetitions per treatment were collected. Each collected repetition consisted of 24 mature leaves, gathered evenly from both sides of every plant from terminating shoots.
Fruit was sampled when their soluble solids concentration (SSC) reached 6.5–7.0%, which is recommended as a suitable time point for kiwiberry harvest [
42]. In 2015, fruit was collected on 3 and 9 September for ‘Geneva’ and ‘Weiki’, respectively, and in 2016, on 30 August and 19 September, respectively. Three repetitions for each N treatment were collected, each consisting of ten similar-sized fruits with no visible deformation or damage.
Leaf and fruit samples were oven-dried at 70 °C and ground to a fine powder, with their dry weights being determined after drying at 105 °C. The micronutrient analysis was conducted in the laboratory of the Chemical and Agricultural Station in Warsaw (Poland, accreditation number AB 312, PN-EN ISO/lEC 17025:2005 standard,
http://www.oschr-warszawa.pl, accessed on 7 July 2017). Standard procedures were used to measure the total micronutrient concentrations in examined kiwiberry tissues. For the mineralization of fruit and leaf samples, phosphoric acid and hydrogen peroxide were used. Flame atomic absorption spectroscopy (FAAS) at 324.8 nm, 248.3 nm, 279.5 nm, and 213.9 nm was used to determine the Cu, Fe, Mn, and Zn concentrations, respectively. The B concentration was evaluated by inductively coupled plasma optical emission spectrometry (ICP-OES) at 249.678 nm. The results were shown as mg kg
−1 of dry weight (DW).
3.3. Statistical Analysis
The obtained data were elaborated using Statistica version 13.0 software (TIBCO Software Inc.,
http://statistica.io—access date 7 July 2017, Palo Alto, CA, USA). The significance of the differences between means of main effects was evaluated using Tukey’s test (HSD) at
p ≤ 0.05. Cultivars were processed separately due to a significant differences in terms of leaf and fruit chemical composition. Leaf analyses included three (year, N-level, time of leaf sampling) main factors to be analyzed, whereas fruit analyses included two (year, N-level). The interactions between main effects were generally not significant, therefore the data were presented as mean values for particular sources of variation, separately for the ‘Geneva’ and ‘Weiki’ cultivars. A summary of statistics data is presented in
Table 1 and
Table 2.
4. Conclusions
The leaf chemical analysis revealed a cultivar-dependent relationship between soil N level and leaf micronutrient concentration. A significant increase in Cu, Zn, Mn, and Fe was noted in ‘Geneva’ leaves when there was a higher N supply, while only the increase in Mn was proven to be significant in ‘Weiki’ leaves. In the case of fruit, increased N fertilization was followed only by a higher Mn content in both tested cultivars. Micronutrient accumulation in A. arguta leaves changed significantly during the growing period. Over time, Mn, Fe, and B concentrations increased markedly, Cu decreased, and Zn remained mostly stable. Between mid-July and the beginning of August, the lowest fluctuations in the micronutrient contents were recorded. The effect of the year on leaf micronutrient accumulation was highly significant; except for Fe, significantly higher micronutrient levels were revealed in 2016, which could be considered as hot and wet compared to both the long-term average and 2015 in particular. Compared to the leaves, the growing season effect was much smaller in the case of fruit micronutrient concentrations. Irrespective of cultivar, the N fertilization increase resulted in a higher fruit Mn concentration and was insignificant in the case of other micronutrients. The results indicate that the N dose may affect the uptake and accumulation of micronutrients within a range depending on the tissue type and the genotype. Interrelationships between essential nutrients are one of the most important issues in terms of effectiveness in plant mineral nutrition.
As a short note, we want to add that the results presented in this manuscript can be read with other reports dealing with N effects on the physiology/biochemistry of kiwiberry plants explored under the same conditions as those described in this study [
33,
36,
43].




{kind=link}