
Application of In Vitro Plant Tissue Culture Techniques to Halophyte Species: A Review
 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Methodology
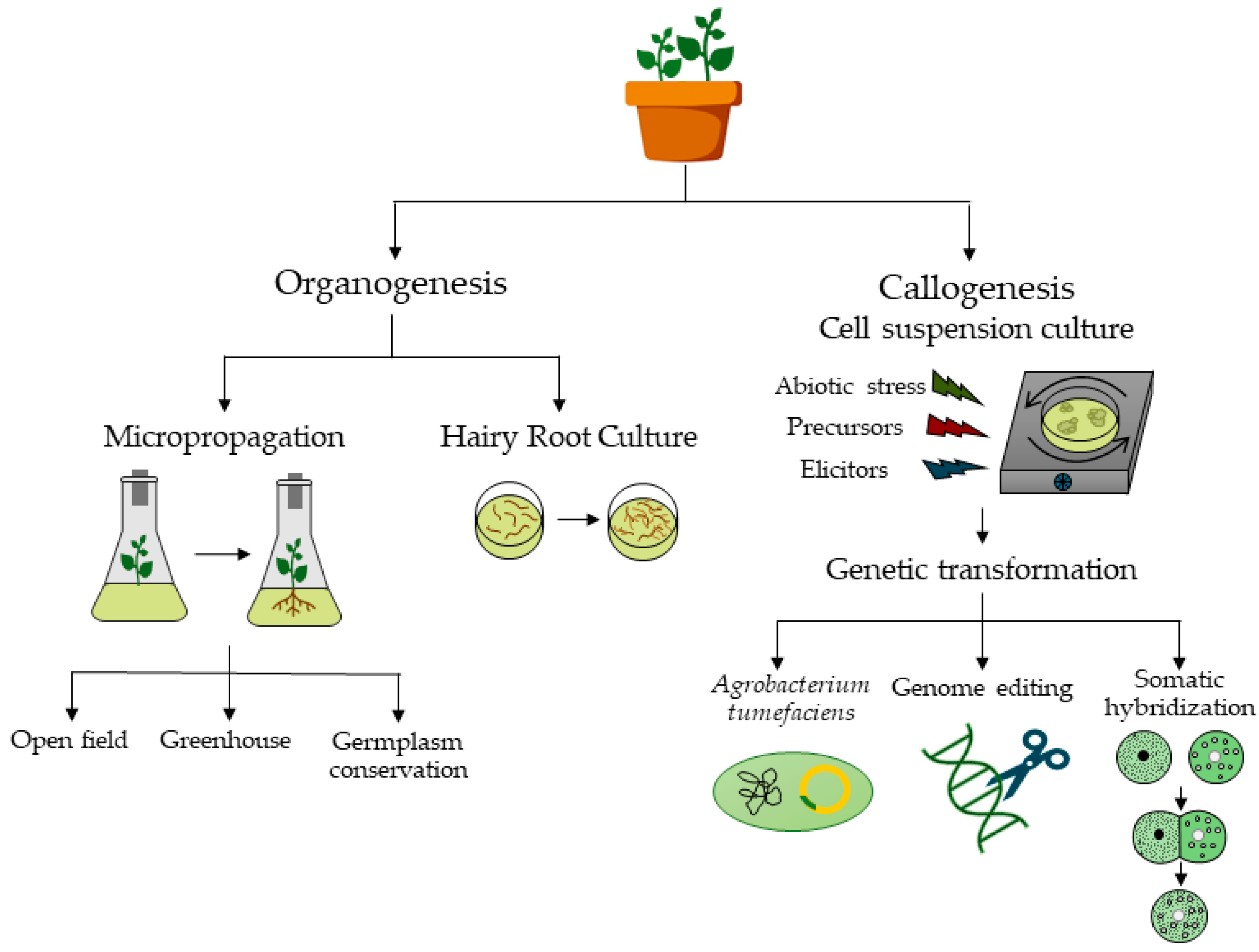
3. In Vitro Plant Tissue Culture
4. Micropropagation of Halophyte Plants
4.1. Material Sources and Decontamination

{kind=link}
| Family/Species | Explant * | Medium ** | Treatment | Morphogenic Response | Growth Regulators *** | Best Results | Acclimatization | References |
|---|---|---|---|---|---|---|---|---|
| Acanthaceae | ||||||||
| Avicennia marina (Forssk.) Vierh. | N | MS + AC | Shoot growth | BAP, NAA | 5 μM BAP + 1 μM NAA | [46] | ||
| MS + AC | Rooting | IBA | 1 μM IBA | + | ||||
| N | MS | Shoot growth | BAP, Kin, IAA | 0.5 mg/L BAP + 1 mg/L Kin + 0.25 mg/L IAA | [47] | |||
| MS | Rooting | IAA, IBA, NAA | 1 mg/L IBA | + | ||||
| Aizoaceae | ||||||||
| Mesembryanthemum crystallinum L. | H, C, L | MS | Caulogenesis | IAA, BAP, NAA | H, C; 1 μM BAP + 1 μM IAA | [48] | ||
| MS | Rooting | MS PGR free | + | |||||
| H | MS with B5 Vit | 80 mM NaCl | Callus induction | 2,4-D, Kin | 1 μM Kin + 5 μM 2,4-D | [49] | ||
| MS | Somatic embryogenesis | 2,4-D, Kin, BAP | 2.5 μM Kin | |||||
| MS | Rooting | PGR free | + | |||||
| Sesuvium portulacastrum (L.) L. | N | MS | Shoot growth | 2iP, BAP, Kin, TDZ | 40 µM 2iP | [50] | ||
| MS | Rooting | IAA, NAA | 5 or 10 µM NAA | + | ||||
| N | MS | 0–600 mM NaCl | Shoot growth | BAP, IBA, GA3 | 200 mM NaCl; 4.44 µM BAP + 0.49 µM IBA + 0.58 µM GA3 | [51] | ||
| Amaranthaceae | ||||||||
| Atriplex canescens (Pursh) Nutt. | N | MS/2 | Shoot growth | BAP, GA3, NAA | 0.01 mg/L NAA + 2 mg/L BAP + 1 mg/L GA3 | [52] | ||
| L | MS/2 | Caulogenesis (direct) | Kin, 2,4-D | 0.01 mg/L 2,4-D + 0.5 mg/L Kin | ||||
| MS | Rooting | IAA, IBA, GA3 | 0.5 mg/L IBA + 0.1 mg/L GA3 | + | ||||
| “, Atriplex torreyi (S. Watson) S. Watson (syn. Atriplex lentiformis ssp. torreyi) | Seed, ST | MS, WPM | Shoot growth | 2iP | WPM with 5 mg/L 2iP | [43] | ||
| WPM | Rooting | PGR free | + | |||||
| Atriplex gmelinii C.A. Mey. ex Bong. | H | LS | Callus induction | BAP, NAA | 1 μM BAP + 5 μM NAA | [53] | ||
| LS | Caulogenesis | NAA, TDZ | 0.1 μM NAA + 20 μM TDZ | |||||
| LS | Rooting | PGR free | n.s. | |||||
| Atriplex halimus L. | ST | MS/2 | 0–1000 mM NaCl | Shoot growth | BAP, IBA, GA3, Kin | 0.1 mg/L GA3; 200 mM NaCl | [54] | |
| MS | Rooting | IBA | PGR free | + | ||||
| N | MS | Shoot growth | BAP, Zea, Kin | 1 mg/L Kin and BAP | [55] | |||
| MS | Shoot multiplication | BAP, Kin, NAA | 0.5 mg/L Kin | |||||
| Beta maritima L. | In | MS | Shoot multiplication | BAP, IAA, NAA, GA3 | 1 mg/L BAP | [56] | ||
| MS/2 | Rooting | NAA | 1 mg/L NAA | + | ||||
| Bienertia sinuspersici Akhani | S | N6, MS + P | 0–200 mM NaCl | Callus induction | 2,4-D | MS + P with 1 mg/L 2,4-D, 50 mM NaCl | [57] | |
| MS + P | CO2, 0–200 mM NaCl | Caulogenesis | BAP | 1.2% CO2, 2 mg/L BAP, 200 mM NaCl | ||||
| MS + P | CO2, 0–200 mM NaCl | Rooting | n.s. | 1.2% CO2, 50 or 200 mM NaCl | + | |||
| Chenopodium quinoa Willd. | H, C, R | MS | Callus induction | 2,4-D | Hypocotyl, 0.45 µM 2,4-D | [58] | ||
| MS | Somatic embryogenesis | - | PGR free | |||||
| ST | MS | Shoot growth | Kin, BAP, NAA | 1 mg/L Kin + 1 mg/L BAP | [44] | |||
| MS | Rooting | IBA | 1 or 2 mg/L IBA | + | ||||
| Halogeton glomeratus (M.Bieb.) Ledeb. | L | MS | Caulogenesis (direct) | BAP, Kin, NAA | 0.5 mg/L BAP + 2 mg/L Kin + 0.2 mg/L NAA | [59] | ||
| Salicornia bigelovii Torr. | ST | MS | Shoot growth | BAP, NAA | 8.89 µM BAP + 0.54 µM NAA | [60] | ||
| MS | Rooting | BAP, NAA | 0.44 µM BAP + 10.74 µM NAA | + | ||||
| Salicornia brachiata Roxb. | ST, N | MS | 0–500 mM NaCl | Shoot multiplication | BAP, Kin, IAA, IBA, NAA, 2,4-D | 250 mM NaCl; 5.37 μM NAA + 44.4 μM BAP | [61] | |
| MS/2 | Rooting | BAP, NAA | 250 or 500 mM NaCl; 5.37 μM NAA + 8.9 or 13.3 μM BAP | + | ||||
| S | MS | Callus induction | 2,4-D | 2 mg/L 2,4-D | [62] | |||
| MS | 80 mM NaCl | Somatic embryogenesis | 2,4-D, IBA | 0.25 mg/L 2,4-D | ||||
| MS | Shoot growth | _ | PGR free | + | ||||
| N | MS * 2 | Shoot growth | BAP, Zea | 3 mg/L BAP + 0.5 mg/L Zea | [63] | |||
| MS * 2 | Shoot multiplication | NAA, TDZ | 1 mg/L NAA + 1 mg/L TDZ | |||||
| MS * 2 | 0–20 g/L MgCl2 | Rooting | IAA, IBA, NAA | 0.5 mg/L NAA + 20 g/L MgCl2 | + | |||
| Salicornia europaea L. | H, ZE, R | MS | 170 mM NaCl | Callus induction | 2,4-D, TDZ | 4.55 µM TDZ | [64] | |
| MS | 0–500 mM NaCl | Caulogenesis | TDZ | 4.55 μM TDZ, 170 mM NaCl | ||||
| MS/2 + AC | Rooting | IBA, Kin | 2.46 μM IBA + 0.46 μM Kin | n.s. | ||||
| ST | MS | 0–100 mM NaCl | Shoot growth | BA, NAA | 100 mM NaCl; 0.5 mg/L NAA + 0.5 mg/L BAP | [65] | ||
| Salsola lanata Pall. (syn. Climacoptera lanata (Pall.) Botsch.) | ZE | MS | Shoot multiplication | Kin | 2.3 μM Kin | [66] | ||
| MS | Shoot growth | BAP, 2iP, IAA | 0.5 μM BAP or 2iP + 0.3 μM IAA | |||||
| L, IS | MS | Callus induction | BAP, Kin, 2iP, IBA, 2,4-D | 9 µM 2,4-D | ||||
| MS | Caulogenesis | BAP | 8 µM BAP | |||||
| MS | Rooting | BAP, 2iP, IAA | 0.5 μM BAP or 2iP + 0.3 μM IAA | n.s. | ||||
| Salsola pestifer A. Nels. (syn. Salsola kali L.) | ZE | MS | Shoot induction | Kin | 2.3 μM Kin | [66] | ||
| MS | Shoot growth | BAP, 2iP, IAA | 0.5 μM BAP or 2iP + 0.3 μM IAA | |||||
| L, IS | MS | Callus induction | BAP, Kin, 2iP, IBA, 2,4-D | 8 µM BAP or 4.9 µM IBA | ||||
| MS | Caulogenesis | BAP | 8 µM BAP | |||||
| MS | Rooting | BAP, 2iP, IAA | 0.5 μM BAP or 2iP + 0.3 μM IAA | n.s. | ||||
| Sarcocornia ambigua (Michx.) M.A. Alonso & M.B. Crespo (syn. Salicornia gaudichaudiana Moq.) | ST, N | MS | 10–30 g/L NaCl, Sediments | Shoot growth | BAP, NAA | ST; 0.5 mg/L NAA + 1 mg/L BAP + 20 g/L NaCl + sediments | [67] | |
| Sarcocornia fruticosa (L.) A.J.Scott | N | H&A * 2 with B5 Vit | 100 mg/L CNH | Shoot growth | CNH + 100 mg/L Vit | [68] | ||
| H&A *2 | 150 mg/L Gln, 100 mg/L CNH | Shoot multiplication | BAP, IAA | PGR free + CNH + Gln | ||||
| H&A * 2 | Gln, CNH | Rooting | GA3 | PGR free + CNH + Gln | n.s. | |||
| Suaeda edulis Flores Olv. & Noguez | N | MS | Shoot growth | BAP | 1 mg/L BAP | [69] | ||
| Suaeda nudiflora (Willd.) Moq. | N | MS | Shoot growth | BAP, Kin | 1 mg/L BAP + 0.2 mg/L Kin | [70] | ||
| MS, MS/2 | Rooting | IAA, IBA, NAA, IPA | MS/2 with IAA + IBA + NAA + IPA (0.5 mg/L each) | + | ||||
| Apiaceae | ||||||||
| Crithmum maritimum L. | ST | B5, MS, WPM | 0–300 mM NaCl | Shoot multiplication | BAP, IBA, NAA | MS with 2.5 µM BAP | [71] | |
| MS | Rooting | IBA, NAA | 2.5 µM IBA or NAA | + | ||||
| Shoot | MS | Shoot growth | BAP, IAA, NAA | 0.5 mg/L BAP + 0.46 mg/L NAA | [72] | |||
| MS | Rooting | IBA | 0.1 mg/L IBA | n.s. | ||||
| Eryngium maritimum L. | N | MS, MS/2 | Shoot growth | BAP, IAA | MS with 1 mg/L BAP + 0.1 mg/L IAA | [73] | ||
| MS, MS/2 | 1.5–5% Sucrose | Rooting | IAA, IBA, NAA | MS/2 + 1.5% Sucrose + 0.1 mg/L IAA | + | |||
| Asteraceae | ||||||||
| Artemisia caerulescens L. | Shoot | MS | Shoot multiplication | BAP | 1 µM BAP | [74] | ||
| Aster tripolium L. (syn. Tripolium pannonicum (Jacq.) Dobrocz.) | C | MS | 0.5 g/L CNH | Callus induction, cell suspen.s.ion | 2,4-D, IAA, NAA, BAP, Kin, Zea, 2iP | 4.9 µM 2iP | [75] | |
| MS | 0.5 g/L CNH | Caulogenesis | NAA, Kin | 5.4 µM NAA + 4.6 µM Kin | ||||
| L | MS | Agar, AgNO3 | Caulogenesis (direct) | 5.4 µM NAA + 4.6 µM Kin | 1.2% agar, 1 g/L AgNO3 | |||
| MS | Rooting | NAA, IBA | 27 µM NAA | + | ||||
| Calendula maritima Guss. (syn. Calendula suffruticosa subsp. maritima (Guss.) Meikle) | L | MS | Caulogenesis (direct) | BAP, NOA, TDZ, IBA | 4.4 µM BAP + 10 µM NOA | [76] | ||
| MS | Rooting | IAA, IBA | 1 µM IAA | + | ||||
| Cineraria maritima Linn. | N | MS with B5 Vit | Shoot growth | BAP, NAA, TDZ | 4.54 µM TDZ | [77] | ||
| MS/2 | Rooting | IBA | 4.92 µM IBA | + | ||||
| Boraginaceae | ||||||||
| Mertensia maritima (L.) Gray | ST, N | MS | Shoot growth | NAA, BAP, Kin, TDZ | 4 µM TDZ + 1 µM NAA | [78] | ||
| MS/2 | Rooting | IAA, IBA, NAA | 4 µM IBA | n.s. | ||||
| Brassicaceae | ||||||||
| Crambe maritima L. | Petiole | MS | Callus induction | BAP, Kin, IAA | 0.5 mg/L IAA + 6 mg/L Kin + 1.5 mg/L BAP | [79] | ||
| MS | Caulogenesis | BAP, Kin | 6 mg/L Kin + 1.5 mg/L BAP | |||||
| MS | Rooting | IBA, NAA | 0.1 mg/L IBA or NAA | + | ||||
| Bromeliaceae | ||||||||
| Dyckia maritima Baker | N | MS | Shoot growth | BAP, Kin | 2 μM BAP + 2 μM Kin | [80] | ||
| MS | Rooting | IBA | 0.5 μM IBA | + | ||||
| Caryophyllaceae | ||||||||
| Honckenya peploides (L.) Ehrh. | ST, N | MS | 0–75 mM NaCl | Shoot growth | BAP, Kin, mT | ST; 25 mM NaCl and 1 mg/L Kin | [81] | |
| MS | 0–75 mM NaCl | Rooting | NAA | 25 mM NaCl and 1.5 mg/L NAA | n.s. | |||
| Ericaceae | ||||||||
| Corema album (L.) D.Don | N | WPM | Shoot growth | 2iP, BAP, Kin, mT | 2 mg/L 2iP + 1 mg/L Kin | [82] | ||
| Soil | ex vitro Rooting | IBA | 2 mg/L IBA | + | ||||
| Euphorbiaceae | ||||||||
| Excoecaria agallocha L. | N | MS, WPM, X | Glutathione | Shoot growth | BAP, Zea, IBA | X medium + 13.3 μM BAP + 4.65 μM Zea + 1.23 μM IBA | [83] | |
| X | Rooting | IBA | 0.23 μM IBA | + | ||||
| N | MS | Shoot growth | BAP, Kin, NAA, 2iP | 3.9 μM BAP + 1.34 μM NAA | [84] | |||
| MS/2 | Rooting | IAA, IBA, NAA | 5.41 μM NAA or 2.85 μM IBA | + | ||||
| Fabaceae | ||||||||
| Alhagi graecorum Boiss. | L, P, S | MS | 0–200 mM NaCl | Somatic embryogenesis | BAP, TDZ, IAA, IBA | L: 1 μM TDZ + 50 mM NaCl | [85] | |
| L, P, S | MS | Caulogenesis (direct) + Shoot growth | BAP, TDZ, IAA, IBA | L: 1 μM TDZ + 0.25 μM IAA | ||||
| MS | Rooting | PGR free | + | |||||
| Pongamia pinnata (L.) Pierre | N | MS | Shoot multiplication | 8.8 μM BAP | [86] | |||
| MS/2 + AC | Rooting | PGR free | + | |||||
| Goodeniaceae | ||||||||
| Scaevola sericea (Gaertn.) Roxb. | N | MS | Shoot multiplication | BAP, Kin, NAA | 1 mg/L BAP + 0.1 mg/L NAA | [18] | ||
| L, R | MS | Callus formation | 2.4-D, BAP, NAA | L; 0.5 mg/L 2,4-D | ||||
| L, R | MS | Somatic embryogenesis | BAP, TDZ | L: 2.5 mg/L BAP; R: 2.5 mg/L TDZ | ||||
| MS/2 | Rooting | NAA | 2.5 mg/L NAA | + | ||||
| Juncaceae | ||||||||
| Juncus roemerianus Scheele | Seed | MS | Callus induction | BAP, NAA, 2,4-D, CW | 2.22 μM BAP + 5.37 μM ANA + 2.26 μM 2,4-D + 5% CW | [87] | ||
| MS | Caulogenesis | BAP, TDZ | 13.3 μM BAP | |||||
| MS | Rooting | IAA, IBA, NAA | 10.7 μM NAA | + | ||||
| Juncus gerardii Loisel. | In | MS | Callus induction | BAP, NAA, 2,4-D, CW | 2.22 μM BAP + 5.37 μM NAA + 2.26 μM 2,4-D + 5% CW | |||
| MS | Caulogenesis | BAP, IAA, TDZ | 0.44 μM BAP + 0.57 μM IAA | |||||
| MS | Rooting | IAA, IBA, NAA | 0.44 μM BAP + 14.8 μM IBA | + | ||||
| Liliaceae | ||||||||
| Urginea maritima (L.) Baker | Bulb scale, L | MS/2, MS | Caulogenesis (direct) + Shoot growth | 2.4-D, BAP, Kin, IAA, NAA | Scale: MS + 2 mg/L BAP; L: MS/2 + 2 mg/L BAP + 2 mg/L NAA | [88] | ||
| MS/2 | Rooting | PGR free | + | |||||
| Bulb scale | MS | Caulogenesis (direct) + Shoot growth | TDZ | 0.55 mg/L TDZ | [89] | |||
| MS | Rooting | IBA | 1 mg/L IBA | + | ||||
| Malvaceae | ||||||||
| Kosteletzkya virginica K. Presl ex Gray (syn. Kosteletzkya pentacarpos (L.) Ledeb.) | S, ZE | MS | Callus | IAA, Kin | 2 mg/L IAA + 1 mg/L 2,4-D | [90] | ||
| MS | Caulogenesis | NAA, Kin | 2 mg/L NAA + 1 mg/L Kin | |||||
| MS/2 | Rooting | IAA, Kin | 0.2 mg/L IAA | + | ||||
| Plantaginaceae | ||||||||
| Bacopa monnieri (L.) Wettst. | L, N | MS, B5 | Caulogenesis, shoot multiplication | BAP, Kin, NAA, 2,4-D | Leaf: MS + 1 mg/L BAP 0.25 mg/L Kin | [91] | ||
| MS/2 | Rooting | IBA | 0.25 mg/L IBA | + | ||||
| Plantago camtschatica Link (syn. Plantago depressa Wild. subsp. camtschatica) | ST | MS | Shoot growth | IAA, BAP, Kin, Zea | 0.6 μM IAA + 8.9 μM BAP | [92] | ||
| H, C, R, L | MS | Caulogenesis (direct) | IAA, BAP, Kin, Zea | H; 0.6 μM IAA + 8.9 μM BAP | ||||
| MS | Rooting | NAA | 0 or 0.5 μM NAA | + | ||||
| Plantago maritima L. | S | MS | Shoot growth | IAA, BAP, Kin | 6.7 μM BAP | [93] | ||
| H, C, R | MS | Caulogenesis (direct) | IAA, BAP, Kin, Zea | Roots; 0.6 μM IAA + 22.8 μM Zea | ||||
| MS | Rooting | IAA, IBA, NAA | 0.5 μM NAA | + | ||||
| Plumbaginaceae | ||||||||
| Armeria maritima (Mill.) Willd. | C | MS with B5 Vit | 88–118 mM sucrose | Somatic embryogenesis | 2.4-D, Kin | 4.5 μM 2.4-D + 0.93 μM Kin, 88 mM sucrose | [94] | |
| MS | Rooting | - | PGR free | + | ||||
| Limoniastrum monopetalum (L.) Boiss. | ST | MS | 0–20 g/L NaCl | Shoot growth | BAP | 0.5 mg/L BAP + 0 g/L NaCl | [95] | |
| MS | 0–20 g/L NaCl | Shoot multiplication | BAP, Zea, Kin, 2iP | 0.5 mg/L BAP + 5 g/L NaCl | ||||
| MS, MS/2 | 0–20 g/L NaCl | Rooting | IBA | MS/2 with 1 mg/L IBA | + | |||
| Limonium bulgaricum Ančev, Limonium gmelinii (Villd.) O. Kuntze, Limonium latifolium (Sm.) O. Kuntze, Limonium meyeri (Boiss.) O. Kuntze, Limonium asterotrichum (Salmon) Salmon, and Limonium vulgare Mill. | In | MS | Shoot multiplication | BAP, IBA, GA3 | BAP + IBA + GA3 (0.1 mg/L each) | [96] | ||
| MS/2 | Rooting | IBA | 1 mg/L IBA | + | ||||
| Limonium aureum (L.) Chaz., Limonium sinuatum (L.) Mill., L. latifolium | C | MS with B5 Vit | 88–118 mM sucrose | Somatic embryogenesis | 2.4-D, Kin | 4.5 μM 2.4-D + 0.93 μM Kin, 88 mM sucrose | [94] | |
| MS | Rooting | - | PGR free | + | ||||
| Limonium bellidifolium (Gouan) Dumort. | C, H, R | MS | 58–117 mM sucrose | Somatic embryogenesis | 2,4-D, Kin | 4.5 µM 2,4-D + 0.5 µM Kin, 117 mM sucrose | [97] | |
| MS | Plantlet growth | Kin | 0.5 µM Kin, 117 mM sucrose | + | ||||
| Limonium bicolor (Bunge) Kuntze | L | MS | Caulogenesis (direct) | BAP, NAA | 4.4 μM BAP + 1.1 μM NAA | [98] | ||
| Rooting | IBA | 4.4 μM IBA | + | |||||
| Limonium perezii (Stapf) F.T.Hubb. ex L.H.Bailey | [99] | |||||||
| Limonium sinuatum | L | MS/2 | Callus induction | Dicamba, picloram, 2,4-D, NAA | 1 mg/L picloram | [100] | ||
| C | MS/2 | Gellan gum, agar | Caulogenesis | BAP, TDZ, Zea | 1 mg/L Zea, 0.25% gellan gum | |||
| MS/2 | Rooting | - | PGR free | + | ||||
| Limonium wrightii (Hance) Kuntze | ST, L, In | MS | Caulogenesis (direct) | BAP, NAA | Shoot tips; 8.87 μM BAP + 17 μM NAA | [101] | ||
| Rooting | IBA, NAA | 4.92 μM IBA | + | |||||
| Plumbago zeylanica L. | ST, In | MS | Callus induction | BAP, IAA, IBA, NAA | 2 mg/L BAP + 1.5 mg/L IAA | [102] | ||
| MS | Caulogenesis | BAP, IAA, NAA, AdS | 0.75 mg/L BAP + 1 mg/L IAA + 1 mg/L NAA + 1 mg/L AdS | |||||
| N | MS | Shoot growth | BAP, IAA, IBA, NAA, AdS | 1 mg/L BAP + 0.5 mg/L IBA + 2 mg/L AdS | ||||
| MS, MS/2 | Rooting | IAA, IBA, NAA | MS/2 + 0.5 mg/L NAA | + | ||||
| Poaceae | ||||||||
| Diplachne fusca (L.) P.Beauv. ex Roem. & Schult. | In-derived callus | MS | Caulogenesis | [45] | ||||
| MS | Shoot multiplication | BAP | 1 mg/L BAP | |||||
| Distichlis spicata (L.) Greene | In | MS | Callus induction | - | PGR free | [103,104] | ||
| MS | Caulogenesis | BAP, NAA, 2,4-D | 0.5 mg/L BAP + 1 mg/L NAA + 0.5 mg/L 2,4-D | |||||
| MS | Shoot regeneration | BAP | 1 mg/L BAP, then 1 mg/L 2,4-D | |||||
| MS | Rooting | - | PGR free | + | ||||
| Hordeum marinum Huds. | ZE | MS | Callus induction | CPA, 2,4-D | 0.5 mg/L CPA or 2,4-D | [105] | ||
| MS | Caulogenesis | IAA, Zea | 1 mg/L IAA + 1 mg/L Zea | + | ||||
| Leymus chinensis (Trin.) Tzvelev | L, Seed | MS | Glu | Callus induction | 2,4-D | Seed; 2 mg/L 2,4-D + 5 mg/L Glu | [106,107] | |
| MS | 2 g/L CNH | Somatic embryogenesis | NAA, Kin | 0.2–0.5 mg/L NAA + 2 mg/L Kin | ||||
| MS/2 | Rooting | - | PGR free | + | ||||
| Puccinellia distans (Jacq.) Parl. | Seed | MS | Callus induction | Kin, 2,4-D | 2 mg/L 2,4-D + 0.5 mg/L Kin | [108] | ||
| MS, N6 | Caulogenesis | Kin, IAA | N6 + 10 mg/L Kin | + | ||||
| Spartina argentinensis Parodi | L, R, In | MS | Callus induction | BAP, NAA, 2,4-D | L: 0.05 mg/L BAP + 0.5 mg/L 2,4-D; In: 0.01 mg/L BAP + 0.1 mg/L 2,4-D | [109] | ||
| MS | Caulogenesis | BAP, NAA | 0.5 mg/L BAP | |||||
| MS | Rooting | NAA | 0.5 mg/L NAA | + | ||||
| Spartina patens Roth) P.M.Peterson & Saarela | S | MS | Callus induction | IAA, 2,4-D | 1 mg/L IAA + 1 mg/L 2,4-D | [110] | ||
| MS | Caulogenesis | BAP, IAA | 3 mg/L BAP | |||||
| MS, MS/2, MS/4 | AC | Rooting | IAA, Kin | MS/4 PGR free | + | |||
| S | MS | Callus induction | IAA, BAP, BL | 0.2 mg/L IAA + 3 mg/L BAP + 0.3 mg/L BL | [111] | |||
| MS | Caulogenesis | IAA, BAP, BL | 0.2 mg/L IAA + 3 mg/L BAP + 0.1 mg/L BL | |||||
| Sporobolus virginicus (L.) Kunth | In | MS | Callus induction | BAP, NAA, 2,4-D, CW | 1 mg/L BAP + 1 mg/L NAA + 0.5 mg/L 2,4-D + 5% CW | [112] | ||
| MS | Caulogenesis | BAP, CW | 1 mg/L BAP + 5% CW | |||||
| Polygonaceae | ||||||||
| Polygonum maritimum L. | N | MS | Shoot multiplication | BAP, Kin, IAA, NAA | 3 mg/L BAP + 0.1 mg/L IAA | [113] | ||
| MS | Rooting | - | PGR free | + | ||||
| Rhizophoraceae | ||||||||
| Bruguiera cylindrica (L.) Blume | H | MS/2 NH4 free | Caulogenesis (direct) | BAP, Kin | PGR free | [114] | ||
| Rooting | PGR free | + | ||||||
| Rhizophora annamalayana Kathiresan | L, ST | MS | Shoot multiplication | BAP, Kin, Zea, CW | ST; 3 mg/L BAP + 3 mg/L Kin + 1% CW | [115] | ||
| Ruppiaceae | ||||||||
| Ruppia maritima L. | Rh | MS/2 | Shoot multiplication | BAP, Kin, 2iP, Zea, TDZ | 10 mg/L Kin or 20 mg/L 2iP + 1 mg/L NAA | [116,117] | ||
| Salvadoraceae | ||||||||
| Salvadora persica L. | N | MS | Shoot growth | BAP, AdS | 8.88 μM BAP + 25 mg/L AdS | [118] | ||
| Shoot multiplication | BAP, Kin, NAA | 1.11 μM BAP + 1.16 μM Kin + 0.54 μM NAA | ||||||
| Rooting | IBA, NOA | 2460.27 μM IBA + 494.56 μM NOA | + | |||||
4.2. Micropropagation from Axillary Buds via Node Culture
4.3. Micropropagation via Direct Shoot Neoformation
4.4. Micropropagation via Indirect Neoformation
4.5. Somatic Embryogenesis
4.6. Androgenesis
4.7. Rooting
4.8. Acclimatization
5. Germplasm Conservation and Cryopreservation
6. Callogenesis and Cell Suspension Cultures
6.1. Biochemical Studies
6.2. Salt-Tolerance Studies
7. Genetic Transformation Studies (Transgenesis)
| Species | Transformed Organ | Gene(s) * | Vector | Procedure | Studied Trait | Reference |
|---|---|---|---|---|---|---|
| Aizoaceae | ||||||
| Mesembryanthemum crystallinum | Callus, Seedling | GUS, NPTII | Agrobacterium tumefaciens | Co-culture | n.s. | [15] |
| Root | GUS | R. rhizogenes | Syringe injection | n.s. | ||
| Seedling | GUS | A. tumefaciens | Co-culture | Stress responses | [167] | |
| Amaranthaceae | ||||||
| Atriplex gmelini | Callus | GUS | A. tumefaciens | Co-culture | n.s. | [53] |
| Sesuvium portulacastrum | Leaf, stem | Ri-TDNA | R. rhizogenes | Co-culture | Phytoremediation | [14] |
| Suaeda salsa (L.) Pall. | Hypocotyl | GUS | A. tumefaciens | Co-culture | n.s. | [171] |
| Brassicaceae | ||||||
| Thellungiella halophila | Flower | FLC | A. tumefaciens | Floral dip | Flowering | [168] |
| Nitrariaceae | ||||||
| Nitraria schoberi L. | Primary leaf | Ri-TDNA | R. rhizogenes | Co-culture | Anti-influenza activity (H5N1, H3N2) | [170] |
| Plumbaginaceae | ||||||
| Limonium bicolor | Leaf segment | GUS | A. tumefaciens | Co-culture | n.s. | [98] |
| Poaceae | ||||||
| Leymus chinensis | Callus | PAT | A. tumefaciens | Particle bombardment | Herbicide resistance | [172] |
| Puccinellia tenuiflora | Cell suspension | AMT1/GFP | A. tumefaciens | Co-culture | Subcellular localization | [173] |
| Callus | GUS, Hyg | A. tumefaciens | Co-culture + US + vacuum | Gene function analysis | [174] |
Somatic Hybridization
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| 2,4-D | 2,4-Dichlorophenoxyacetic acid |
| 2,4,5-T | 2,4,5-Trichlorophenoxyacetic acid |
| BAP | 6-Benzylaminopurine |
| Bl | Brassinolides |
| CMCs | Cambial meristematic cells |
| CNH | Casein hydrolysate (casaminoacids) |
| CW | Coconut water |
| Gln | Glutamine |
| Glu | Glutamic acid |
| H&A | Hoagland and Arnon |
| IAA | Indole-3-acetic acid |
| IBA | Indole-3-butyric acid |
| IPA | Indole-3-propionic acid |
| Kin | Kinetin |
| MeJ | Methyl jasmonate |
| LS | Linsmaier and Skoog |
| MS | Murashige and Skoog |
| mT | meta-Topoline |
| n.s. | Non-specified |
| NAA | 1-Naphthaleneacetic acid |
| PAL | Phenylalanine ammonium lyase |
| PGRs | Plant growth regulators |
| SE | Somatic embryogenesis |
| SMs | Secondary metabolites |
| SOS | Salt Overly Sensitive |
| TDZ | Thidiazuron (1-phenyl-3-(1,2,3-thiadiazol-5-yl) urea) |
| WPM | Woody plant medium |
| ZEA | Zeatin |
References
- Custódio, M.; Villasante, S.; Cremades, J.; Calado, R.; Lillebø, A.I. Unravelling the potential of halophytes for marine integrated multi-trophic aquaculture (IMTA) a perspective on performance, opportunities and challenges. Aquac. Environ. Interact. 2017, 9, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Shabala, S.; Fujita, M. Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; CABI Digital Library: Dhaka, Bangladesh, 2019; p. 408. [Google Scholar]
- Gulzar, B.; Mujib, A.; Qadir Malik, M.; Mamgain, J.; Syeed, R.; Zafar, N. Plant tissue culture: Agriculture and industrial applications. In Transgenic Technology Based Value Addition in Plant Biotechnology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 25–49. [Google Scholar]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.J.; Custódio, L.; Lopes, A.; Oliveira, M.; Neng, N.R.; Nogueira, J.M.F.; Martins, A.; Rauter, A.P.; Varela, J.; Barreira, L. Unlocking the in vitro anti-inflammatory and antidiabetic potential of Polygonum maritimum. Pharm. Biol. 2017, 55, 1348–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.J.; Pereira, C.A.; Oliveira, M.; Neng, N.R.; Nogueira, J.M.F.; Zengin, G.; Mahomoodally, M.F.; Custódio, L. Sea rose (Armeria pungens (Link) Hoffmanns. & Link) as a potential source of innovative industrial products for anti-ageing applications. Ind. Crops Prod. 2018, 121, 250–257. [Google Scholar]
- Rodrigues, M.J.; Monteiro, I.; Placines, C.; Castaneda-Loaiza, V.; Ślusarczyk, S.; Matkowski, A.; Pereira, C.; Pousão-Ferreira, P.; Custodio, L. The irrigation salinity and harvesting affect the growth, chemical profile and biological activities of Polygonum maritimum L. Ind. Crops Prod. 2019, 139, 111510. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Monteiro, I.; Castañeda-Loaiza, V.; Placines, C.; Oliveira, M.C.; Reis, C.; Carpeta, A.D.; Soares, F.; Pousão-Ferreira, P.; Pereira, C.; et al. Growth performance, in vitro antioxidant properties and chemical composition of the halophyte Limonium algarvense Erben are strongly influenced by the irrigation salinity. Ind. Crops Prod. 2020, 143, 111930. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Castañeda-Loaiza, V.; Monteiro, I.; Pinela, J.; Barros, L.; Abreu, R.M.V.; Oliveira, M.C.; Reis, C.; Soares, F.; Pousão-Ferreira, P.; et al. Metabolomic Profile and Biological Properties of Sea Lavender (Limonium algarvense Erben) Plants Cultivated with Aquaculture Wastewaters: Implications for Its Use in Herbal Formulations and Food Additives. Foods 2021, 10, 3104. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Jekő, J.; Cziáky, Z.; Pereira, C.G.; Custódio, L. The Medicinal Halophyte Frankenia laevis L. (Sea Heath) Has In vitro Antioxidant Activity, α-Glucosidase Inhibition, and Cytotoxicity towards Hepatocarcinoma Cells. Plants 2022, 11, 1353. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Y.; Yu, J.; Zhang, H.; Wang, L.; Wang, S.; Guo, S.; Miao, Y.; Chen, S.; Li, Y.; et al. NaCl-responsive ROS scavenging and energy supply in alkaligrass callus revealed from proteomic analysis. BMC Genom. 2019, 20, 990. [Google Scholar] [CrossRef] [PubMed]
- Lokhande, V.H.; Kudale, S.; Nikalje, G.; Desai, N.; Suprasanna, P. Hairy root induction and phytoremediation of textile dye, Reactive green 19A-HE4BD, in a halophyte, Sesuvium portulacastrum L. Biotechnol. Rep. 2015, 8, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.H.; Wang, C.H.; Chen, H.H.; Ho, J.F.; Chi, S.F.; Huang, F.C.; Yen, H.E. Effective Agrobacterium-mediated transformation protocols for callus and roots of halophyte ice plant (Mesembryanthemum crystallinum). Bot. Stud. 2019, 60, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halfaoui, Y.; Kadiri, A.; Ighilhariz, Z. Atriplex halimus (Amaranthaceae) callogenesis induction from different explant type. J. Fund. Appl. Sci. 2018, 10, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.; Kanthaliya, B.; Arora, J. Evaluation of growth and antioxidant activity in Suaeda monoica and Suaeda nudiflora callus cultures under sequential exposure to saline conditions. Curr. Biotechnol. 2019, 8, 42–52. [Google Scholar] [CrossRef]
- Liang, H.; Xiong, Y.; Guo, B.; Yan, H.; Jian, S.; Ren, H.; Zhang, X.; Li, Y.; Zeng, S.; Wu, K.; et al. Shoot organogenesis and somatic embryogenesis from leaf and root explants of Scaevola sericea. Sci. Rep. 2020, 10, 11343. [Google Scholar] [CrossRef]
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Hühn, T.; Eibl, D. Plant cell culture technology in the cosmetics and food industries: Current state and future trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Tanmoy, B. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Academic Press: Haryana, India, 2015; p. 452. [Google Scholar]
- Renna, M. Reviewing the prospects of sea fennel (Crithmum maritimum L.) as emerging vegetable crop. Plants 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ahmed, M.Z.; Hameed, A. Effect of sea salt and L-ascorbic acid on the seed germination of halophytes. J. Arid Environ. 2006, 67, 535–540. [Google Scholar] [CrossRef]
- Oliveira, P.B.; Valdiviesso, T.; Luz, F.R. Melhoramento Genético da camarinha; Seleção e Avaliação de plantas. Actas Port. Hortic. 2020, 30, 338–346. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Akin, B. Tissue culture techniques of medicinal and aromatic plants: History, cultivation and micropropagation. J. Sci. Rep. A 2020, 45, 253–266. [Google Scholar]
- Kumar, N.; Reddy, M.P. In vitro plant propagation: A review. J. For. Environ. Sci. 2011, 27, 61–72. [Google Scholar]
- El Meskaoui, A. Plant cell tissue and organ culture biotechnology and its application in medicinal and aromatic plants. Med. Aromat. Plants 2013, 2, e147. [Google Scholar] [CrossRef]
- Ochoa-Villarreal, M.; Howat, S.; Hong, S.; Jang, M.O.; Jin, Y.W.; Lee, E.K.; Loake, G.J. Plant cell culture strategies for the production of natural products. BMB Rep. 2016, 49, 149. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef]
- Marchev, A.S.; Yordanova, Z.P.; Georgiev, M.I. Green (cell) factories for advanced production of plant secondary metabolites. Crit. Rev. Biotechnol. 2020, 40, 443–458. [Google Scholar] [CrossRef]
- EUMOFA. Available online: https://www.eumofa.eu/documents/20178/84590/blue+bioeconomy.pdf/f5a87949-c541-416b-16e7-521155cdff06?t=1608051570785 (accessed on 17 August 2022).
- Efferth, T. Biotechnology applications of plant callus cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.M.; Ritala, A.; Cardon, F. Hairy root cultures—A versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Wawrosch, C.; Zotchev, S.B. Production of bioactive plant secondary metabolites through in vitro technologies—Status and outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649–6668. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol. 2007, 27, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Mishra, S.; Srivastava, V. Hairy Roots Biotechnology Unzipped: A Journey of Reality and Promises. In Hairy Root Cultures Based Applications; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer: Singapore, 2020; pp. 1–10. [Google Scholar]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- WHO. Available online: https://www.who.int/news-room/fact-sheets/detail/biodiversity-and-health (accessed on 7 September 2022).
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Sheikholeslami, B.; Shukla, M.; Turi, C.; Harpur, C.; Saxena, P.K. Saving threatened plant species: Reintroduction of Hill’s thistle (Cirsium hillii (Canby) Fernald) to its natural habitat. PLoS ONE 2020, 15, e0231741. [Google Scholar] [CrossRef] [Green Version]
- Edesi, J.; Tolonen, J.; Ruotsalainen, A.L.; Aspi, J.; Häggman, H. Cryopreservation enables long-term conservation of critically endangered species Rubus humulifolius. Biodivers. Conserv. 2020, 29, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Vera, I.; Lucero, M.; Barrow, J. An improved protocol for micropropagation of saltbush (Atriplex) species. NPJ 2010, 11, 52–56. [Google Scholar] [CrossRef]
- Regalado Gonzalez, J.J.; Tossi, V.; Burrieza, H.; Encina, C.; Pitta, S. Micropropagation protocol for coastal quinoa. Plant Cell Tissue Organ Cult. 2020, 142, 213–219. [Google Scholar] [CrossRef]
- Nanakorn, M.; Jiamjetjaroon, W.; Suwanawong, S.; Wongwattana, C.; Shim, I.S. In vitro selection of salt-tolerant cell lines in kallar grass [Diplachne fusca (L.) Beauv.]. Weed Biol. Manag. 2003, 3, 49–52. [Google Scholar] [CrossRef]
- Alatar, A.; Faisal, M.; Hegazy, A.; Alwathnani, H.; Okla, M. Clonal in vitro multiplication of grey mangrove and assessment of genetic fidelity using single primer amplification reaction (SPAR) methods. Biotechnol. Biotechnol. Equ. 2015, 29, 1069–1074. [Google Scholar] [CrossRef]
- Mangrio, A.; Rafiq, M.; Sheng, Z.; Rind, N.; Farzana, M.; Korejo, B.; Habib, S.; Naqvi, S.; Mangrio, S. Comparative study of growth and biochemical attributes of Avicennia marina (Forssk.) Vierh of Indus delta and in vitro raised plants established at Jamshoro, Sindh Pakistan. Pak. J. Bot. 2021, 53, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Meiners, M.S.; Thomas, J.C.; Bohnert, H.J.; Cushman, J.C. Regeneration of multiple shoots and plants from Mesembryanthemum crystallinum. Plant Cell Rep. 1991, 9, 563–566. [Google Scholar] [CrossRef]
- Cushman, J.C.; Wulan, T.; Kuscuoglu, N.; Spatz, M.D. Efficient plant regeneration of Mesembryanthemum crystallinum via somatic embryogenesis. Plant Cell Rep. 2020, 19, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Lokhande, V.H.; Nikam, T.D.; Ghane, S.G.; Suprasanna, P. In vitro culture, plant regeneration and clonal behaviour of Sesuvium portulacastrum (L.) L.: A prospective halophyte. Physiol. Mol. Biol. Plants 2010, 16, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Lokhande, V.H.; Nikam, T.D.; Patade, V.Y.; Ahire, M.L.; Suprasanna, P. Effects of optimal and supra-optimal salinity stress on antioxidative defence, osmolytes and in vitro growth responses in Sesuvium portulacastrum L. Plant Cell Tissue Organ Cult. 2011, 104, 41–49. [Google Scholar] [CrossRef]
- Mei, B.; No, E.G.; Mcwilliams, E.L.; Gould, J.H.; Newton, R.J. In vitro regeneration of fourwing saltbush [Atriplex canescens (Pursh) Nutt.]. J. Range Manag. 1997, 50, 413–418. [Google Scholar] [CrossRef]
- Uchida, A.; Nagamiya, K.; Takabe, T. Transformation of Atriplex gmelini plants from callus lines using Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult. 2003, 75, 151–157. [Google Scholar] [CrossRef]
- Aldahhak, O.; Zaid, S.; Abdul-Kader, A.M. Development of in vitro propagation system for Atriplex halimus L. Int. J. Hortic. Sci. 2014, 20, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Papafotiou, M.; Majumder, D.A.N.; Martini, A.N.; Bertsouklis, K.F. Micropropagation of Atriplex halimus L. Acta Hortic. 2016, 1113, 207–210. [Google Scholar] [CrossRef]
- Zhong, Z.; Smith, H.G.; Thomas, T.H. Micropropagation of wild beet (Beta maritima) from inflorescence pieces. Plant Growth Regul. 1993, 12, 53–57. [Google Scholar] [CrossRef]
- Rosnow, J.; Offermann, S.; Park, J.; Okita, T.W.; Tarlyn, N.; Dhingra, A.; Edwards, G.E. In vitro cultures and regeneration of Bienertia sinuspersici (Chenopodiaceae) under increasing concentrations of sodium chloride and carbon dioxide. Plant Cell Rep. 2011, 30, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Eisa, S.; Koyro, H.; Kogel, K.H.; Imani, J. Induction of somatic embryogenesis in cultured cells of Chenopodium quinoa. Plant Cell Tissue Organ Cult. 2005, 81, 243–246. [Google Scholar] [CrossRef]
- Yao, L.; Wang, J.; Yang, K.; Li, B.; Meng, Y.; Ma, X.; Lai, Y.; Si, E.; Ren, P.; Shang, X.; et al. In vitro regeneration system of Halogeton glomeratus: An important halophyte. In Vitro Cell. Dev. Biol.-Plant 2021, 57, 332–340. [Google Scholar] [CrossRef]
- Lee, C.W.; Glenn, E.P.; O’Leary, J.W. In Vitro Propagation of Salicornia bigelovii by Shoot-tip Cultures. Hort. Sci. 1992, 27, 472. [Google Scholar] [CrossRef]
- Joshi, M.; Mishra, A.; Jha, B. NaCl plays a key role for in vitro micropropagation of Salicornia brachiata, an extreme halophyte. Ind. Crops Prod. 2012, 35, 313–316. [Google Scholar] [CrossRef]
- Rathore, M.S.; Paliwal, N.; Anand, K.G.V.; Agarwal, P.K. Somatic embryogenesis and in vitro plantlet regeneration in Salicornia brachiata Roxb. Plant Cell Tissue Organ Cult. 2015, 120, 355–360. [Google Scholar] [CrossRef]
- Singh, A.; Kruti, J.; Priyanka, K.; Pradeep Kumar, A. Effect of MgCl2 and double concentration of Murashige and Skoog medium on in vitro plantlet and root cultures generation in halophytic grasswort Salicornia brachiata. Plant Cell Tissue Organ Cult. 2015, 120, 563–570. [Google Scholar] [CrossRef]
- Shi, X.L.; Han, H.P.; Shi, W.L.; Li, Y.X. NaCl and TDZ are two key factors for the improvement of in vitro regeneration rate of Salicornia europaea L. J. Integr. Plant Biol. 2006, 48, 1185–1189. [Google Scholar] [CrossRef]
- Mahmoudi, A.; Danesh, M. Assessment of Salinity Effects on Some Morphological and Physiological Traits and In Vitro Culture of Halophyte Plant (Salicornia europaea). J. Crop Breed. 2019, 11, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Stefaniak, B.; Wozny, A.; Li, V. Plant micropropagation and callus induction of some annual Salsola species. Biol. Plant. 2003, 46, 305–308. [Google Scholar] [CrossRef]
- Gonçalves, J.R.; Zaffari, G.R. In vitro morphogenetic response of Sarcocornia ambigua. Plant Cell Cult. Micropropag. 2019, 14, 56–63. [Google Scholar]
- Raposo, M.F.J.; de Morais, R.M.S.C. Micropropagation of the Halophyte Sarcocornia fruticosa (L.) A. J. Scott. J. Basic Appl. Sci. 2014, 10, 53–59. [Google Scholar]
- Cerrillo-Rojas, G.V.; Tiscareño-Andrade, M.; Ochoa-Alfaro, A.E.; Pérez-Molphe Balch, E.; Soria-Guerra, R.E.; Morales-Domínguez, J.F. In Vitro Propagation, Isolation and Expression Studies of Suaeda edulis Genes Involved in the Osmoprotectants Biosynthesis. Phyton-Int. J. Exp. Bot. 2020, 89, 715–726. [Google Scholar] [CrossRef]
- Cherian, S.; Reddy, M.P.M. Micropropagation of the halophyte Suaeda nudiflora MOQ. through axillary bud culture. Indian J. Plant Physiol. 2002, 7, 40–43. [Google Scholar]
- Grigoriadou, K.; Maloupa, E. Micropropagation and salt tolerance of in vitro grown Crithmum maritimum L. Plant Cell Tissue Organ Cult. 2008, 94, 209–217. [Google Scholar] [CrossRef]
- Pistelli, L.; Noccioli, C.; D’Angiolillo, F.; Pistelli, L. Composition of volatile in micropropagated and field grown aromatic plants from Tuscany Islands. Acta Biochim. Pol. 2013, 60, 43–50. [Google Scholar] [CrossRef]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Oleszek, W. The Effect of Nutritional Factors and Plant Growth Regulators on Micropropagation and Production of Phenolic Acids and Saponins from Plantlets and Adventitious Root Cultures of Eryngium maritimum L. J. Plant Growth Regul. 2014, 33, 809–819. [Google Scholar] [CrossRef]
- Pieracci, Y.; Vento, M.; Pistelli, L.; Lombardi, T.; Pistelli, L. Halophyte Artemisia caerulescens L.: Metabolites from In Vitro Shoots and Wild Plants. Plants 2022, 11, 1081. [Google Scholar] [CrossRef]
- Uno, Y.; Nakao, S.; Yamai, Y.; Koyama, R.; Kanechi, M.; Inagaki, N. Callus formation, plant regeneration, and transient expression in the halophyte sea aster (Aster tripolium L.). Plant Cell Tissue Organ Cult. 2009, 98, 303–309. [Google Scholar] [CrossRef]
- Carra, A.; Bambina, M.; Pasta, S.; Garfì, G.; Badalamenti, O.; Catalano, C.; Carimi, F.; Sajeva, M. In-vitro regeneration of Calendula maritima guss. (Asteraceae), a threatened plant endemic to Western Sicily. Pak. J. Bot. 2016, 48, 589–593. [Google Scholar]
- Banerjee, S.; Tripathi, J.; Verma, P.C.; Dwivedi, P.D.; Khanuja, S.P.S.; Bagchi, G.D. Thidiazuron-induced high-frequency shoot proliferation in Cineraria maritima Linn. Curr. Sci. 2004, 87, 1287–1290. [Google Scholar]
- Park, H.; Kim, D.H.; Saini, R.K.; Gopal, J.; Keum, Y.-S.; Sivanesan, I. Micropropagation and Quantification of Bioactive Compounds in Mertensia maritima (L.) Gray. Int. J. Mol. Sci. 2019, 20, 2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peron, J.; Regnier, E. In vitro propagation of Crambe maritima. Can. J. Bot. 2011, 65, 72–75. [Google Scholar] [CrossRef]
- Lopes da Silva, A.L.; Franco, E.; Dornelles, E.; Gesing, J. Micropropagation of Dyckia maritima Baker—Bromeliaceae. Iheringia Ser. Bot. 2008, 63, 135–138. [Google Scholar]
- Kulpa, D.; Wrobel, M.; Bednarek, M. Type of Explant Affects In Vitro Development and Multiplication Success of the Rare Halophyte Plant Honckenya peploides L. Ehrh. Plants 2020, 9, 1526. [Google Scholar] [CrossRef]
- Alves, V.; Pinto, R.; Debiasi, C.; Conceição Santos, M.; Carlos Gonçalves, J.; Domingues, J. Micropropagation of Corema album from adult plants in semisolid medium and temporary immersion bioreactor. Plant Cell Tissue Organ Cult. 2021, 145, 641–648. [Google Scholar] [CrossRef]
- Rao, C.S.; Eganathan, P.; Anand, A.; Balakrishna, P.; Reddy, T.P. Protocol for in vitro propagation of Excoecaria agallocha L., a medicinally important mangrove species. Plant Cell Rep. 1998, 17, 861–865. [Google Scholar]
- Manickam, A.; Ramachandra, U.P.; Rajaram, P. A Micropropagation Protocol for a Critically Endangered Mangrove Excoecaria agallocha L. Int. J. Conserv. Sci. 2012, 3, 119–126. [Google Scholar]
- Zobayed, S.M.A.; Murch, S.J.; El-Demerdash, M.A.; Saxena, P.K. NaCl enhances growth and morphogenesis potential of Alhagi graecorum. In Vitro Cell. Dev. Biol.-Plant 2006, 42, 607. [Google Scholar] [CrossRef]
- Sujatha, R.; Hazra, S. In Vitro Regeneration of Pongamia pinnata Pierre. J. Plant Biotechnol. 2006, 33, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Seliskar, D.M.; Gallagher, J.L. Tissue Culture and Plant Regeneration of the Salt Marsh Monocots Juncus roemerianus and Juncus gerardi. In Vitro Cell. Dev. Biol.-Plant 2005, 41, 274–280. [Google Scholar] [CrossRef]
- Stojakowska, A. Micropropagation of Urginea maritima (L.) Baker s. str. Acta Soc. Bot. Pol. 1993, 62, 11–15. [Google Scholar] [CrossRef]
- Aasim, M.; Khawar, K.M.; Özcan, S. In Vitro Regeneration of Red Squill Urginea maritima (L.) Baker Using Thidiazuron. Biotechnol. Biotechnol. Equ. 2014, 22, 925–928. [Google Scholar] [CrossRef]
- Cook, D.A.; Decker, D.M.; Gallagher, J.L. Regeneration of Kosteletzkya virginica (L.) Presl. (Seashore Mallow) from callus cultures. Plant Cell Tissue Organ Cult. 1989, 17, 111–119. [Google Scholar] [CrossRef]
- Rahe, A.; Mollika, S.R.; Khan, S.; Banu, T.A.; Amin, G.M.; Habib, A.; Akter, S.; Islam, M.; Sharmin, R.A. In vitro Micropropagation of Bacopa monnieri (L.) Penn.—An Important Medicinal Plant. Plant Tissue Cult. Biotechnol. 2020, 30, 57–63. [Google Scholar] [CrossRef]
- Andrzejewska-Golec, E.; Makowczynska, J. Micropropagation of Plantago camtschatica Link. Acta Soc. Bot. Pol. 2008, 77, 269–273. [Google Scholar]
- Makowczyñska, J.; Andrzejewska-Golec, E.; Sliwinska, E. Nuclear DNA content in different plant materials of Plantago maritima L. cultured in vitro. Plant Cell Tissue Organ Cult. 2008, 94, 65–71. [Google Scholar] [CrossRef]
- Aly, M.A.M.; Rathinasabapathi, B.; Bhalsod, S. Somatic embryogenesis in members of the Plumbaginaceae ornamental statice Limonium and sea thrift Armeria maritima. Hort. Sci. 2002, 37, 1122–1123. [Google Scholar] [CrossRef] [Green Version]
- Martini, A.N.; Papafotiou, M. In Vitro Propagation and NaCl Tolerance of the Multipurpose Medicinal Halophyte Limoniastrum monopetalum. HortScience 2020, 55, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Kaninski, A.; Ivanova, I.; Bistrichanov, S.; Zaryanova, N.; Atanassova, B.; Iakimova, E.T. Ex situ conservation of endangered Limonium species in the Bulgarian flora. J. Fruit Ornam. Plant Res. 2012, 20, 115–129. [Google Scholar] [CrossRef] [Green Version]
- Aly, M.A.M.; Rathinasabapathi, B.; Kelley, K. Somatic embryogenesis in perennial statice Limonium bellidifolium, Plumbaginaceae. Plant Cell Tissue Organ Cult. 2002, 68, 127–135. [Google Scholar] [CrossRef]
- Yuan, F.; Chen, M.; Yang, J.; Leng, B.; Wang, B. A system for the transformation and regeneration of the recretohalophyte Limonium bicolor. In Vitro Cell Dev. Biol. Plant 2014, 50, 610–617. [Google Scholar] [CrossRef]
- Kunitake, H.; Mii, M. Plant regeneration from cell culture-derived protoplasts of statice (Limonium perezii Hubbard). Plant Sci. 1990, 70, 115–119. [Google Scholar] [CrossRef]
- Igawa, T.; Hoshino, Y.; Mii, M. Efficient plant regeneration from cell cultures of ornamental statice, Limonium sinuatum Mill. In Vitro Cell. Dev. Biol.-Plant 2002, 38, 157–162. [Google Scholar] [CrossRef]
- Huang, C.L.; Hsieh, M.T.; Hsieh, W.C.; Sagare, A.P.; Tsay, H.S. In vitro propagation of Limonium wrightii (Hance) Ktze. (Plumbaginaceae), an ethnomedicinal plant, from shoot-tip, leaf- and inflorescence-node explants. In Vitro Cell. Dev. Biol.-Plant 2000, 36, 220–224. [Google Scholar] [CrossRef]
- Sivanesan, I.; Jeong, B.R. Micropropagation of Plumbago zeylanica L. Afr. J. Biotechnol. 2009, 8, 3761–3768. [Google Scholar]
- Straub, P.F.; Decker, D.M.; Gallagher, J.L. Tissue culture and regeneration of Distichlis spicata (Gramineae). Am. J. Bot. 1989, 76, 1448–1451. [Google Scholar] [CrossRef]
- Seliskar, D.M.; Gallagher, J.L. Exploiting wild population diversity and somaclonal variation in the salt marsh grass Distichlis spicata (Poaceae) for marsh creation and restoration. Am. J. Bot. 2000, 87, 141–146. [Google Scholar]
- Rotem-Abarbanell, D.; Breiman, A. Plant regeneration from immature and mature embryo derived calli of Hordeum marinum. Plant Cell Tissue Organ Cult. 1989, 16, 207–216. [Google Scholar] [CrossRef]
- Sun, Y.; Hong, S. Somatic embryogenesis and in vitro plant regeneration from various explants of the halophyte Leymus chinensis (Trin.). J. Plant Biotechnol. 2009, 36, 236–243. [Google Scholar] [CrossRef]
- Sun, Y.L.; Hong, S.K. Effects of plant growth regulators and L-glutamic acid on shoot organogenesis in the halophyte Leymus chinensis (Trin.). Plant Cell Tissue Organ Cult. 2010, 100, 317–328. [Google Scholar] [CrossRef]
- Binh, D.Q.; Heszky, L.E.; Gyulai, G. Plant regeneration from callus of Puccinellia distans (L.) Parl. Plant Cell Tissue Organ Cult. 1989, 18, 195–200. [Google Scholar] [CrossRef]
- Bueno, M.; Sorrequieta, J.; Feldman, S.; Ortiz, J.P.A. Regeneración in vitro de Spartina argentinensis Parodi. Rev. Colomb. Biotecnol. 2012, 14, 61–69. [Google Scholar]
- Li, X.G.; Seliskar, D.M.; Moga, J.A.; Gallagher, J.L. Plant regeneration from callus cultures of salt marsh hay, Spartina patens, and its cellular-based salt tolerance. Aquat. Bot. 1995, 51, 103–113. [Google Scholar] [CrossRef]
- Lu, Z.; Huang, M.; Ge, D.P.; Yang, Y.H.; Cai, X.N.; Qin, P.; She, J.M. Effect of brassinolide on callus growth and regeneration in Spartina patens (Poaceae). Plant Cell Tissue Organ Cult. 2003, 73, 87–89. [Google Scholar] [CrossRef]
- Straub, P.F.; Decker, D.M.; Gallagher, J.L. Characterization of tissue culture initiation and plant regeneration in Sporobolus virginicus (Gramineae). Am. J. Bot. 1992, 79, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Custódio, L.; Slusarczyk, S.; Matkowski, A.; Castañeda-Loaiza, V.; Fernandes, E.; Pereira, C.G.; Rodrigues, M. A first approach for the micropropagation of the medicinal halophyte Polygonum maritimum L. and phenolic profile of acclimatized plants. Front. Plant Sci. 2022, 13, 960306. [Google Scholar] [CrossRef]
- Vartak, V.; Shindikar, M. Micropropagation of rare mangrove Bruguiera cylindrica L. towards conservation. Indian J. Biotechnol. 2008, 7, 255–259. [Google Scholar]
- Kathiresan, K.; Singh, C.R. Preliminary conservation effort on Rhizophora annamalayana Kathir. The only endemic mangrove to India through in vitro method. J. Plant Dev. 2013, 20, 57–61. [Google Scholar]
- Koch, E.; Durako, M. In vitro studies of the submerged Angiosperm Ruppia maritima: Auxin and cytokinin effects on plant growth and development. Mar. Biol. 1991, 110, 1–6. [Google Scholar] [CrossRef]
- Bird, K.T.; Jewett-Smith, J.; Fonseca, M.S. Use of in vitro Propagated Ruppia maritima for seagrass meadow restoration. J. Coast. Res. 1994, 10, 732–737. [Google Scholar]
- Phulwaria, M.; Ram, K.; Gahlot, P.; Shekhawat, N.S. Micropropagation of Salvadora persica—A tree of arid horticulture and forestry. New For. 2011, 42, 317–327. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Murashige, T.; Thorpe, T.A.; Vasil, I.K. Plant Tissue Culture Media. In Vitro 1976, 12, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1938, 347, 32. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- McCown, B.H.; Lloyd, G. Woody Plant Medium (WPM)—A Mineral Nutrient Formulation for Microculture of Woody Plant Species. HortScience 1981, 16, 453. [Google Scholar]
- Ayuso, M.; Landín, M.; Gallego, P.P.; Barreal, M.E. Artificial intelligence tools to better understand seed dormancy and germination. In Seed Dormancy and Germination; Jimenes-Lopez, J.C., Ed.; IntechOpen: London, UK, 2019; pp. 55–71. [Google Scholar]
- Khan, M.A.; Gul, B. Halophyte seed germination. In Ecophysiology of High Salinity Tolerant Plants; Springer: Dordrecht, The Netherlands, 2006; pp. 11–30. [Google Scholar]
- Gul, B.; Ansari, R.; Flowers, T.J.; Ajmal Khan, M. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Copetta, A.; Bazzicalupo, M.; Cassetti, A.; Marchioni, I.; Mascarello, C.; Cornara, L.; Pistelli, L.; Ruffoni, B. Plant production and leaf anatomy of Mertensia maritima (L.) Gray: Comparison of in vitro culture methods to improve acclimatization. Horticulturae 2021, 7, 111. [Google Scholar] [CrossRef]
- Aldahhak, O.; Zaid, S.; da Silva, J.A.T.; Abdul-Kader, A.M. In vitro approach to the multiplication of a halophyte species forage shrub Atriplex halimus L. and in vitro selection for salt tolerance. Int. J. Plant Dev. Biol. 2010, 4, 8–14. [Google Scholar]
- Von Hedenström, H.; Breckle, S.-W. Obligate halophytes? A test with tissue culture methods. Z. Pflanzenphysiol. 1974, 74, 183–185. [Google Scholar] [CrossRef]
- Blazquez, S.; Olmos, E.; Hernández, J.A.; Fernández-García, N.; Fernández, J.; Piqueras, A. Somatic embryogenesis in saffron (Crocus sativus L.). Histological differentiation and implication of some components of the antioxidant enzymatic system. Plant Cell Tissue Organ Cult. 2009, 97, 49–57. [Google Scholar] [CrossRef]
- Zang, Q.L.; Jiao, Y.L.; Guo, X.M.; Fei, Z.; Yeh, K.; Lin, X. Callus induction and plant regeneration from lateral shoots of herbaceous bamboo Mniochloa abersend. J. Hortic. Sci. Biotechnol. 2016, 92, 168–174. [Google Scholar] [CrossRef]
- Chand, S.; Sahrawat, A.K. Stimulatory effect of partial desiccation on plant regeneration in indica rice (Oryza sativa L). J. Plant Biochem. Biotechnol. 2001, 10, 43–47. [Google Scholar] [CrossRef]
- Othmani, A.; Bayoudh, C.; Drira, N.; Marrakchi, M.; Trifi, M. Somatic embryogenesis and plant regeneration in date palm Phoenix dactylifera L., cv. Boufeggous is significantly improved by fine chopping and partial desiccation of embryogenic callus. Plant Cell Tissue Organ Cult. 2009, 97, 71–79. [Google Scholar] [CrossRef]
- Kenny, L.; Caligari, P.D. Androgenesis of the salt tolerant shrub Atriplex glauca. Plant Cell Rep. 1996, 15, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.R.; Kathiresan, K. In vitro callus induction from Ceriops decandra—A true mangrove viviparous. Int. J. Adv. Multidiscip. Res. 2015, 2, 86052945. [Google Scholar]
- Barraco, G.; Sylvestre, I.; Iapichino, G.; Engelmann, F. Cryopreservation of Limonium serotinum apical meristems from in vitro plantlets using droplet-vitrification. Sci. Hortic. 2011, 130, 309–313. [Google Scholar] [CrossRef]
- Ravinder Singh, C.; Kandasamy, K.; Sekar, A.; Kanagaraj, S. Antioxidant and antibacterial activity of field grown and tissue cultured root callus of mangrove species. Eur. J. Med. Plants 2014, 4, 723–742. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Ogita, S.; Kawana, Y.; Oyanagi, T.; Hasegawa, A.; Sasamoto, H. Relation between amino acids profiles and recalcitrancy of cell growth or salt tolerance in tissue and protoplast cultures of three mangrove species, Avicennia alba, Bruguiera sexangula, and Sonneratia alba. Am. J. Plant Sci. 2013, 4, 1366–1374. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, A.; Kurita, A.; Hayashi, S.; Fukumoto, T.; Sasamoto, H. Halophilic and salt tolerant protoplast cultures of mangrove plants, Sonneratia alba and Avicennia marina. Plant Biotechnol. Rep. 2013, 7, 205–209. [Google Scholar] [CrossRef]
- Treichel, S. The influence of NaCl on 1-pyrroline-5-carboxylate reductase in proline-accumulating cell suspension cultures of Mesembryanthemum nodiflorum and other halophytes. Physiol. Plant. 1986, 67, 173–181. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Nikam, T.D.; Suprasanna, P. Biochemical, physiological and growth changes in response to salinity in callus cultures of Sesuvium portulacastrum L. Plant Cell Tissue Organ Cult. 2010, 102, 17–25. [Google Scholar]
- Sharma, V.; Ramawat, K.G. Salt stress enhanced antioxidant response in callus of three halophytes (Salsola baryosma, Trianthema triquetra, Zygophyllum simplex) of Thar desert. Biologia 2014, 69, 178–185. [Google Scholar] [CrossRef]
- El-Amery, E.M.; Matsuda, R.; El-Khatib, A.; Takechi, K.; Takano, H.; Takio, S. Differential tolerance to high salt with regard to cell growth and superoxide dismutase (SOD) activity in calluses of the halophyte Suaeda maritima from Japan and Egypt. Plant Omics J. 2016, 9, 81–89. [Google Scholar]
- Zhao, S.Z.; Sun, H.Z.; Chen, M.; Wang, B.S. Light-regulated betacyanin accumulation in euhalophyte Suaeda salsa calli. Plant Cell Tissue Organ Cult. 2010, 102, 99–107. [Google Scholar] [CrossRef]
- Uno, Y.; Kanechi, M.; Inagaki, N.; Taki, N.; Maekawa, S. Growth and protein profile responses in the halophyte sea aster (Aster tripolium L.) suspension-cultured cells to salinity. J. Plant Res. 1996, 109, 409–414. [Google Scholar] [CrossRef]
- Bucchini, A.; Giamperi, L.; Ricci, D. Total polyphenol content, in vitro antifungal and antioxidant activities of callus cultures from Inula crithmoides. Nat. Prod. Comm. 2013, 8, 1587–1590. [Google Scholar]
- Ben Hamed, I.B.; Biligui, B.; Arbelet-Bonnin, D.; Abdelly, C.; Ben Hamed, K.; Bouteau, F. Establishment of a cell suspension culture of the halophyte Cakile maritima. Adv. Hortic. Sci. 2014, 28, 43–48. [Google Scholar]
- Hamed-Laouti, I.B.; Arbelet-Bonnin, D.; De Bont, L.; Biligui, B.; Gakière, B.; Abdelly, C.; Ben Hamed, K.; Bouteau, F. Comparison of NaCl-induced programmed cell death in the obligate halophyte Cakile maritima and the glycophyte Arabidospis thaliana. Plant Sci. 2016, 247, 49–59. [Google Scholar] [CrossRef]
- Arbelet-Bonnin, D.; Hamed-Laouti, I.B.; Laurenti, P.; Abdelly, C.; Ben Hamed, K.; Bouteau, F. Cellular mechanisms to survive salt in the obligate halophyte Cakile maritima. Plant Sci. 2018, 272, 173–178. [Google Scholar] [CrossRef]
- Zhao, X.; Tan, H.J.; Liu, Y.B.; Li, X.R.; Chen, G.X. Effect of salt stress on growth and osmotic regulation in Thellungiella and Arabidopsis callus. Plant Cell Tissue Organ Cult. 2009, 98, 97–103. [Google Scholar] [CrossRef]
- Akatu, M.; Hosoi, Y.; Sasamoto, H.; Ashihara, H. Purine metabolism in cells of a mangrove plant, Sonneratia alba, in tissue culture. J. Plant Physiol. 1996, 149, 133–137. [Google Scholar] [CrossRef]
- Yasumoto, E.; Adachi, K.; Kato, M.; Sano, H.; Sasamoto, H.; Baba, S.; Ashii-Iar, H. Uptake of inorganic ions and compatible solutes in cultured mangrove cells during salt stress. In Vitro Cell. Dev. Biol. Plant 1999, 35, 82–85. [Google Scholar] [CrossRef]
- Blits, K.C.; Cook, D.A.; Gallagher, J.L. Salt tolerance in cell suspension cultures of the halophyte Kosteletzkya virginica. J. Exp. Bot. 1993, 44, 681–686. [Google Scholar] [CrossRef]
- Gourguillon, L.; Rustenholz, C.; Lobstein, A.; Gondet, L. Callus induction and establishment of cell suspension cultures of the halophyte Armeria maritima (Mill.) Willd. Sci. Hortic. 2018, 233, 407–411. [Google Scholar] [CrossRef]
- Warren, R.S.; Gould, A.R. Salt tolerance expressed as a trait in suspension cultures developed from the halophytic grass Distichlis spicata. Z. Pflanzenphysiol. 1982, 107, 347–356. [Google Scholar] [CrossRef]
- Wu, J.D.; Seliskar, M. Salinity adaptation of plasma membrane H+-ATPase in the salt marsh plant Spartina patens: ATP hydrolysis and enzyme kinetics. J. Exp. Bot. 1998, 49, 1005–1013. [Google Scholar] [CrossRef]
- Warren, R.S.; Baird, L.M.; Thompson, A.K. Salt tolerance in cultured cells of Spartina pectinata. Plant Cell Rep. 1985, 4, 84–87. [Google Scholar] [CrossRef]
- Mimura, T.; Mimura, M.; Washitani-Nemoto, S.; Sakano, K.; Shimmen, T.; Siripatanadilok, S. Efficient callus initiation from leaf of mangrove plant, Bruguiera sexangula in amino acid medium: Effect of NaCl on callus initiation. J. Plant Res. 1997, 110, 25–29. [Google Scholar] [CrossRef]
- Kura-Hotta, M.; Mimura, M.; Tsujimura, T.; Washitani-Nemoto, S.; Mimura, T. High salt-treatment-induced Na+ extrusion and low salt-treatment-induced Na+ accumulation in suspension-cultured cells of the mangrove plant, Bruguiera sexangula. Plant Cell Environ. 2001, 24, 1105–1112. [Google Scholar] [CrossRef]
- Fitriana, D.; Prihastanti, E.; Nurchayati, Y.; Hastuti, R.B. Effect of combination explant difference leaf part and concentration of active charcoal on callus initiation mangrove (Rhizophora apiculata BI) by in-vitro. J. Phys. 2019, 1217, 012166. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Ramawat, K.G. Salinity-induced modulation of growth and antioxidant activity in the callus cultures of miswak (Salvadora persica). 3 Biotech 2013, 3, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wei, X.; Shi, R.; Fan, Q.; An, L. Salinity-induced physiological modification in the callus from halophyte Nitraria tangutorum Bobr. J. Plant Growth Regul. 2010, 29, 465–476. [Google Scholar] [CrossRef]
- Linsmaier, E.M.; Skoog, F. Organic Growth Factor Requirements of Tobacco Tissue Cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Javed, F.; Ashraf, M. Iso-osmotic effect of NaCl and PEG on growth, cations and free proline accumulation in callus tissue of two indica rice (Oryza sativa L.) genotypes. Plant Growth Regul. 2007, 53, 53–63. [Google Scholar] [CrossRef]
- Tonon, G.; Kevers, C.; Faivre-Rampant, O.; Grazianil, M.; Gaspar, T. Effect of NaCl and mannitol iso-osmotic stresses onproline and free polyamine levels in embryogenic Fraxinus angustifolia callus. J. Plant Physiol. 2004, 161, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Arzani, A. Improving salinity tolerance in crop plants: A biotechnological view. In Vitro Cell. Dev. Biol. Plant 2008, 44, 373–383. [Google Scholar] [CrossRef]
- Wu, J.; Seliskar, D.M.; Gallagher, J.L. The response of plasma membrane lipid composition in callus of the halophyte Spartina patens (Poaceae) to salinity stress. Am. J. Bot. 2005, 92, 852–858. [Google Scholar] [CrossRef]
- Ishimaru, K. Transformation of a CAM plant, the facultative halophyte Mesembryanthemum crystallinum by Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult. 1999, 57, 61–63. [Google Scholar] [CrossRef]
- Fang, Q.; Xu, Z.; Song, R. Cloning, characterization and genetic engineering of FLC homolog in Thellungiella halophila. Biochem. Biophys. Res. Commun. 2006, 347, 707–714. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Martínez-Jiménez, C.; Izquierdo-Martínez, A.; Acosta-Motos, J.R.; Hernández, J.A.; Díaz-Vivancos, P. H2O2-elicitation of black carrot hairy roots induces a controlled oxidative burst leading to increased anthocyanin production. Plants 2021, 10, 2753. [Google Scholar] [CrossRef] [PubMed]
- Zheleznichenko, T.; Banaev, E.; Asbaganov, S.; Voronkova, M.; Kukushkina, T.; Filippova, E.; Mazurkova, N.; Shishkina, L.; Novikova, T. Nitraria schoberi L. hairy root culture as a source of compounds with antiviral activity against influenza virus subtypes A(H5N1) and A(H3N2). 3 Biotech 2018, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.Z.; Ruan, Y.; Sun, H.Z.; Wang, B.S. Highly efficient Agrobacterium-based transformation system for callus cells of the C3 halophyte Suaeda salsa. Acta Physiol. Plant. 2008, 30, 729–736. [Google Scholar] [CrossRef]
- Shu, Q.Y.; Liu, G.S.; Xu, S.X.; Li, X.F.; Li, H.J. Genetic transformation of Leymus chinensis with the PAT gene through microprojectile bombardment to improve resistance to the herbicide Basta. Plant Cell Rep. 2005, 24, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Zhao, M.; Sun, B.; Zhang, X.; Takano, T.; Liu, S. An efficient method for stable protein targeting in grasses (Poaceae): A case study in Puccinellia tenuiflora. BMC Biotechnol. 2014, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Qin, C.; Liu, S.; Xu, Y.; Li, Y.; Zhang, Y.; Song, Y.; Sun, M.; Fu, C.; Qin, Z.; et al. Establishment of an efficient Agrobacterium-mediated genetic transformation system in halophyte Puccinellia tenuiflora. Mol. Breed. 2021, 41, 55. [Google Scholar] [CrossRef]
- Xia, G.M.; Zhou, A.F.; Xiang, F.; Chen, H.M. Asymmetric somatic hybridization between Triticum aestivum L. (wheat) and Leymus chinensis (Trin.) Tzvel. In Somatic Hybridization in Crop Improvement II. Biotechnology in Agriculture and Forestry; Nagata, T., Bajaj, Y.P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 49, pp. 65–77. [Google Scholar]
- Chen, X.L.; Xia, G.M.; Chen, H.M. Nuclear and cytoplasmic genome analysis of somatic hybrid of Triticum aestivum L. and Leymus chinensis (Trin.) Tzvel. J. Plant Physiol. Mol. Biol. 2004, 30, 379–386. [Google Scholar]
- Xia, G.; Xiang, F.; Zhou, A.; Wang, H.; Chen, H. Asymmetric somatic hybridization between wheat (Triticum aestivum L.) and Agropyron elongatum (Host) Nevishi. Theor. Appl. Genet. 2003, 107, 299–305. [Google Scholar] [CrossRef]
- Wei, Y.; Guangmin, X.; Daying, Z.; Huimin, C. Transfer of salt tolerance from Aeluropus littoralis sinensis to wheat (Triticum aestivum L.) via asymmetric somatic hybridization. Plant Sci. 2001, 161, 259–266. [Google Scholar] [CrossRef]

| Family/Species | Explant Source * | Medium ** | Conditions | Growth Regulators *** | Optimal Conditions | Result | Aim/Application | Reference |
|---|---|---|---|---|---|---|---|---|
| Acanthaceae | ||||||||
| Acanthus ilicifolius L. | R | MS | 2,4-D, IAA, NAA, BAP, Kin | 0.3 mg/L 2,4-D + 0.5 mg/L BAP | Callus | Biological activities | [136] | |
| Avicennia alba Blume | C, H | AAM | 2,4-D, TDZ | 1 μM 2,4-D + 1 μM TDZ | Callus, Protoplasts | Salt tolerance | [137] | |
| Avicennia marina | R | MS | 2,4-D, IAA, NAA, BAP, Kin | 0.3 mg/L 2,4-D + 0.5 mg/L BAP | Protoplasts | Salt tolerance | [138] | |
| Aizoaceae | ||||||||
| Mesembryanthemum crystallinum | H | LS | Kin, 2,4-D | 0.5 μM Kin + 2.3 μM 2,4-D | Cell suspension | Salt responses | [139] | |
| Sesuvium portulacastrum | N | MS | BAP | 20 μM BAP, then 10 μM 2,4-D + 5 μM BAP | Callus | Salt tolerance | [140] | |
| Trianthema triquetra Willd. | n.s. | MS, MS/2 | 0–200 mM NaCl | 2,4,5-T, Kin | 1 mg/L 2,4,5-T + 0.1 mg/L Kin; 50 or 100 mM NaCl | Callus | Antioxidant activities | [141] |
| Amaranthaceae | ||||||||
| Atriplex halimus | C, H, IS, L, ST | MS/2, B5/2 | 2,4-D, Kin | S, H; B5/2 + 0.5 mg/L 2,4-D + 0.5 mg/L Kin | Callus | Micropropagation | [16] | |
| Salicornia europaea | H | B5 | IAA, Kin | 1 mg/L IAA + 10 mg/L Kin | Callus | Salt tolerance | [128] | |
| Salsola baryosma (Roem. & Schult.) Dandy (syn. Caroxylon imbricatum (Forssk.) Akhani & Roalson) | n.s. | MS, MS/2 | 0–200 mM NaCl | 2,4,5-T, Kin | 1 mg/L 2,4,5-T + 0.1 mg/L Kin; 0–100 mM NaCl | Callus | Antioxidant activities | [141] |
| Salsola lanata | IS, L | MS | BAP, Kin, 2iP, IBA, 2,4-D | 9 µM 2,4-D | Callus | Salt tolerance | [66] | |
| Salsola pestifer | IS, L | MS | BAP, Kin, 2iP, IBA, 2,4-D | 8.8 µM BAP or 4.9 µM IBA | Callus | Salt tolerance | [66] | |
| Suaeda maritima (L.) Dumort. | H | MS | 0–400 mM NaCl | 2,4-D, Kin | 1 μM 2,4-D + 1 μM Kin; 0 or 200 mM NaCl | Callus | Salt tolerance | [142] |
| H | B5 | IAA, Kin | 1 mg/L IAA + 10 mg/L Kin | Callus | Salt tolerance | [124] | ||
| Suaeda monoica Forssk. ex J.F.Gmel. | H | MS | 0–1000 mM NaCl | 2,4-D, BAP, NAA, Kin | 1 mg/L 2,4-D + 0.5 mg/L BAP; 500 mM NaCl | Callus | Salt tolerance | [17] |
| Suaeda nudiflora | Ep | MS | 0–1000 mM NaCl | 2,4-D, BAP, NAA, Kin | 0.5 mg/L 2,4-D + 0.25 Kin; 0 mM NaCl | Callus | Salt tolerance | [17] |
| Suaeda salsa | H | MS | 2,4-D, BAP | 0.2 mg/L 2,4-D + 0.5 mg/L BAP | Callus | Betacyanin synthesis | [143] | |
| Asteraceae | ||||||||
| Aster tripolium (syn. Tripolium pannonicum) | C | MS | 2,4-D, Zea, 2iP | 0.5 mg/L 2,4-D + 0.1 mg/L Zea, transferred to 0.1 mg/L 2,4-D + 1 mg/L 2iP | Callus, cell suspension | Salt responses | [144] | |
| Inula crithmoides L. | L | MS | 2,4-D, IBA, NAA | 1 mg/L 2,4-D | Callus | Biological activities | [145] | |
| Brassicaceae | ||||||||
| Cakile maritima Scop. | IS | B5 | 2,4-D, Kin | 9.06 μM 2,4-D + 0.46 μM Kin | Callus | Salt tolerance | [146] | |
| B5 | 0–800 mM NaCl | 2,4-D, Kin | 0.2 μM 2,4-D + 0.45 μM Kin | Cell suspension | Salt responses | |||
| n.s. | MS | 50–400 mM NaCl | 2,4-D | 0.2 mg/L 2,4-D | Cell suspension | Salt responses | [147] | |
| n.s. | MS | 50–400 mM NaCl | 2,4-D | 0.2 mg/L 2,4-D | Cell suspension | Salt responses | [148] | |
| Thellungiella halophila (Bayanaul) (syn. Eutrema halophilum (C.A.Mey.) Al-Shehbaz & Warwick) | L | MS | 2,4-D, Kin | 1 mg/L 2,4-D + 0.05 mg/L Kin | Callus | Salt responses | [149] | |
| Clusiaceae | ||||||||
| Calophyllum inophyllum L. | R | MS | 2,4-D, IAA, NAA, BAP, Kin | 0.3 mg/L 2,4-D + 0.5 mg/L BAP | Callus | Biological activities | [136] | |
| Euphorbiaceae | ||||||||
| Excoecaria agallocha L. | R | MS | 2,4-D, IAA, NAA, BAP, Kin | 0.3 mg/L 2,4-D + 0.5 mg/L BAP | Callus | Biological activities | [136] | |
| Lythraceae | ||||||||
| Sonneratia alba Sm. | Pistil | MS | 2,4-D, phenylurea | 0.1 µM 2,4-D + 0.1 µM Phenylurea | Callus | n.s. | [150] | |
| MS | 0–500 mM NaCl | 50 mM NaCl | Callus | Salt responses | [151] | |||
| Malvaceae | ||||||||
| Kosteletzkya virginica (syn. Kosteletzkya pentacarpos) | Callus | MS | 0–255 mM NaCl | 85 mM NaCl | Cell suspension | Salt tolerance | [152] | |
| Plumbaginaceae | ||||||||
| Armeria maritima | C, L, R, YL | MS | 2,4-D, NAA, Kin, BAP | 4.5 μM 2.4-D + 0.93 μM Kin | Callus, cell suspension | Bioproduction | [153] | |
| Poaceae | ||||||||
| Diplachne fusca | In | MS | 2,4-D | 1 mg/L 2,4-D | Callus | Salt tolerance | [45] | |
| Distichlis spicata | ST | MS | 2,4-D | 4 mg/L 2,4-D | Callus, cell suspension | Salt tolerance | [154] | |
| Puccinellia tenuiflora (Griseb.) Scribn. & Merr. | Seed | MS | 2,4-D | 4 mg/L 2,4-D | Callus | Salt responses | [13] | |
| Spartina patens | Seedling | MS | BAP, NAA, 2,4-D, CW | 0.5 mg/L 2,4-D + 0.5 mg/L BAP + 1 mg/L NAA + 5% CW | Callus | Salt tolerance | [110,155] | |
| Spartina pectinata Link (syn. Sporobolus michauxianus (Hitchc.) P.M.Peterson & Saarela) | In | MS | 2,4-D | 2 mg/L 2,4-D | Cell suspension | Salt tolerance | [156] | |
| Rhizophoraceae | ||||||||
| Bruguiera sexangula (Lour.) Poir. | L, seedling | MS, AAM | 2,4-D, Phenylurea | AAM + 2 µM 2,4-D + 2 µM Phenylurea | Callus, cell suspension | Salt responses | [157,158] | |
| Ceriops decandra (Griff.) W.Theob. | R | MS | BAP, IAA, IBA, NAA | 0.5 mg/L BAP + 2.5 mg/L NAA | Callus | Bioproduction | [134] | |
| Rhizophora apiculata Blume | L | MS | 4–20 g/L AC | BAP, NAA | 12 g/L AC; 0.3 mg/L BAP + 1 mg/L NAA | Callus | n.s. | [159] |
| Salvadoraceae | ||||||||
| Salvadora persica | N | MS | 2,4,5-T, BAP | 0.5 mg/L 2,4,5-T + 0.5 mg/L BAP | Callus | Salt responses | [160] | |
| Zygophyllaceae | ||||||||
| Nitraria tangutorum Bobr. | C | MS | NAA, BAP | 0.3 mg/L BAP + 1 mg/L NAA | Callus | Salt responses | [161] | |
| Tetraena simplex (L.) Beier & Thulin | n.s. | MS, MS/2 | 0–200 mM NaCl | 2,4,5-T, Kin | 0.5 mg/L 2,4,5-T + 0.1 mg/L Kin; 50 and 100 mM NaCl | Callus | Antioxidant activities | [141] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Custódio, L.; Charles, G.; Magné, C.; Barba-Espín, G.; Piqueras, A.; Hernández, J.A.; Ben Hamed, K.; Castañeda-Loaiza, V.; Fernandes, E.; Rodrigues, M.J. Application of In Vitro Plant Tissue Culture Techniques to Halophyte Species: A Review. Plants 2023, 12, 126. https://doi.org/10.3390/plants12010126
Custódio L, Charles G, Magné C, Barba-Espín G, Piqueras A, Hernández JA, Ben Hamed K, Castañeda-Loaiza V, Fernandes E, Rodrigues MJ. Application of In Vitro Plant Tissue Culture Techniques to Halophyte Species: A Review. Plants. 2023; 12(1):126. https://doi.org/10.3390/plants12010126
Chicago/Turabian StyleCustódio, Luísa, Gilbert Charles, Christian Magné, Gregorio Barba-Espín, Abel Piqueras, José A. Hernández, Karim Ben Hamed, Viana Castañeda-Loaiza, Eliana Fernandes, and Maria João Rodrigues. 2023. "Application of In Vitro Plant Tissue Culture Techniques to Halophyte Species: A Review" Plants 12, no. 1: 126. https://doi.org/10.3390/plants12010126