
The Multiple Role of Silicon Nutrition in Alleviating Environmental Stresses in Sustainable Crop Production




Abstract
:1. Introduction
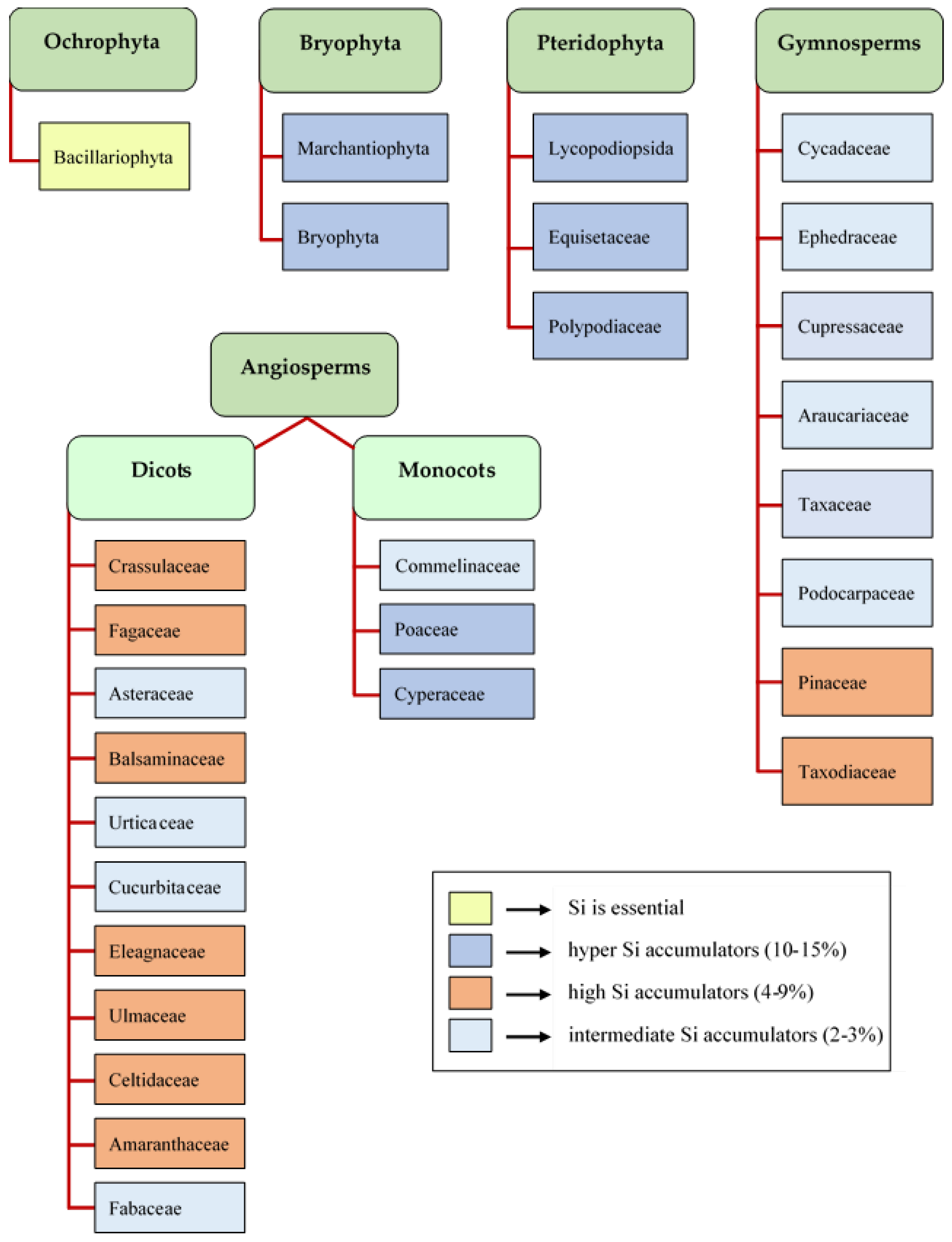
2. Analysis of Silicon in Plants
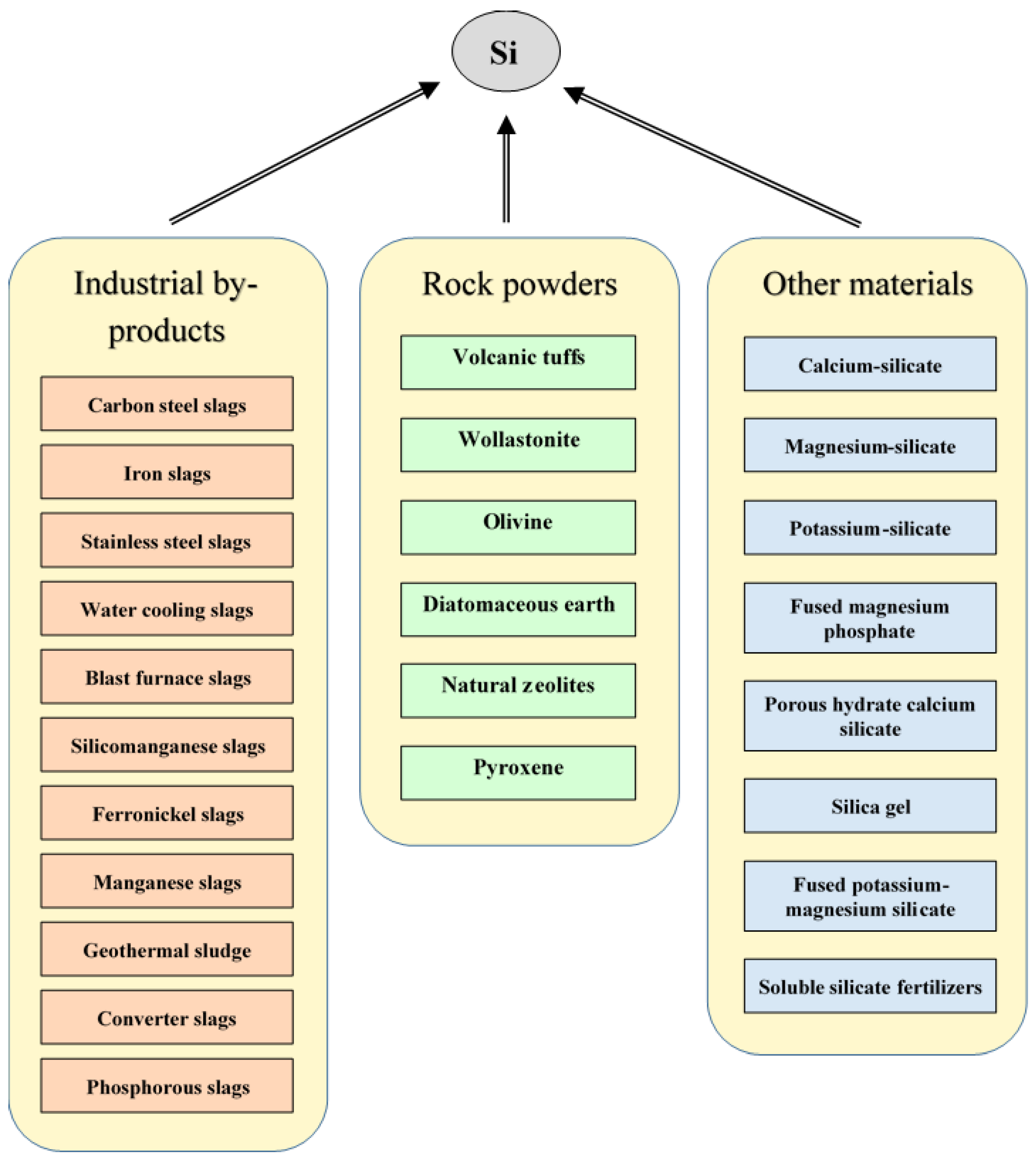
3. Availability of Silicon for Plants
4. Silicon Uptake and Transport in Plants
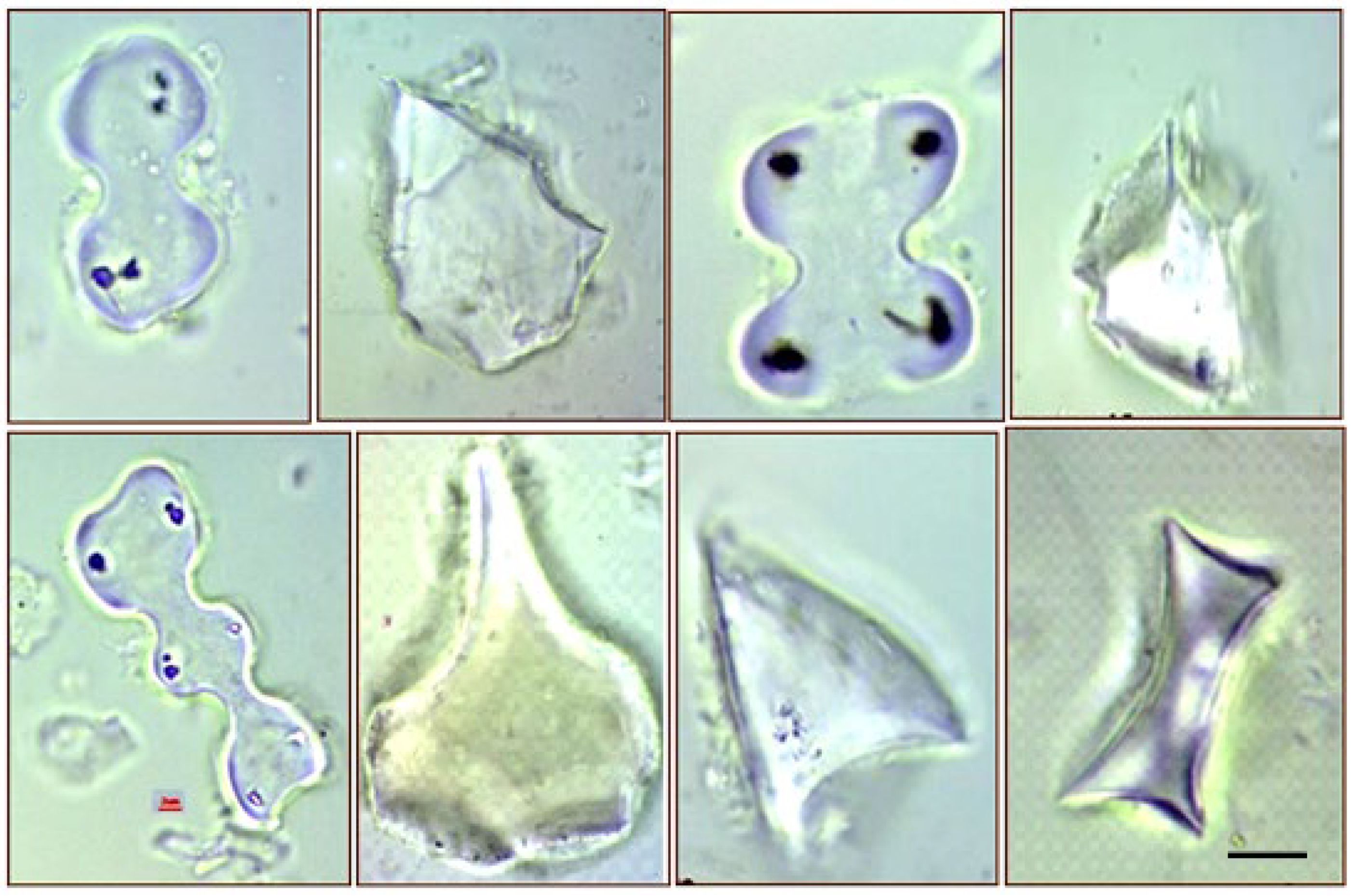
5. Phytoliths
Areas of Practical Use for Phytoliths
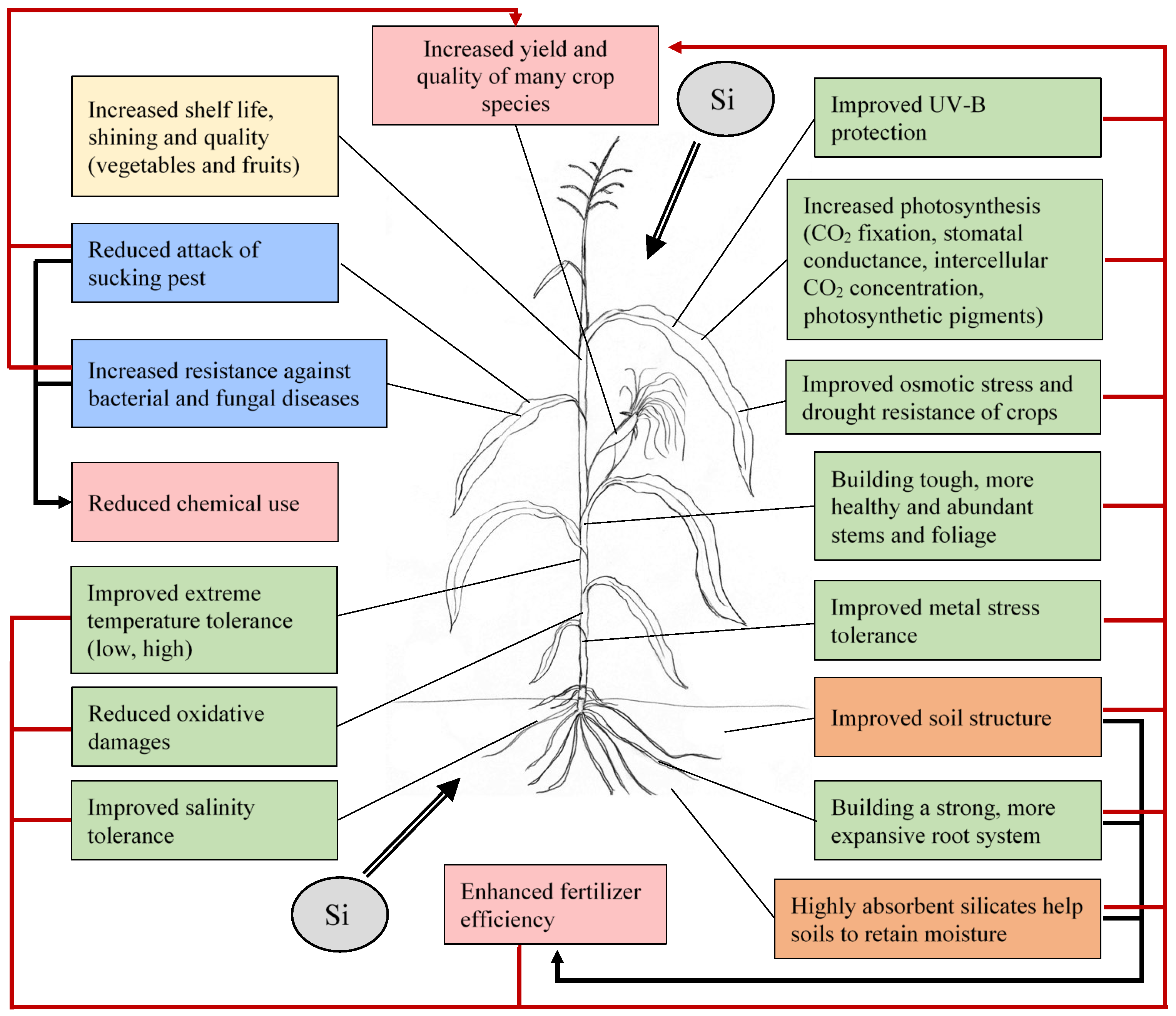
6. Effects and Possibilities of Silicon Fertilization
7. Silicon—A Valuable Tool for Regulating Biological and Abiological Stresses in Crop Production
7.1. Abiotic Stresses
7.1.1. Function of Silicon in Mitigating Plant Nutrient Imbalance and Improving Nutrient Use Efficiency
7.1.2. Function of Silicon in Mitigating Metal and Heavy Metal Stress
7.1.3. Function of Silicon in Mitigating the Negative Effect of Heat and Drought Stress
7.1.4. Function of Silicon in Mitigating the Negative Effect of Salt Stress
7.1.5. Function of Silicon in Carbon Sequestration
7.2. Biotic Stresses
7.2.1. Disease Resistance
7.2.2. Pest Resistance
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 1–294. ISBN 9780080525761. [Google Scholar]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Onodera, I. Chemical studies on rice blast. J. Sci. Agric. Soc. 1917, 180, 606–617. [Google Scholar]
- Savant, N.K.; Datnoff, L.E.; Snyder, G.H. Depletion of plant-available silicon in soils: A possible cause of declining rice yields. Commun. Soil Sci. Plant Anal. 1997, 28, 1245–1252. [Google Scholar] [CrossRef]
- Datnoff, L.; Rodrigues, F. The Role of Silicon in Suppressing Rice Diseases. APSnet Feature Artic. 1997, 58. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture from Theory to Practice; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Role of siucon in rice nutrition. Soil Sci. Plant Nutr. 1959, 5, 127–133. [Google Scholar] [CrossRef]
- Snyder, G.H. Methods for silicon analysis in plants, soils, and fertilizers. In Studies in Plant Science Volume 8 Silicon in Agriculture, 1st ed.; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 185–196. [Google Scholar]
- Meyer, M.L.; Bloom, P.R. Lithium metaborate fusion for silicon, calcium, magnesium, and potassium analysis of wild rice. Plant Soil 1993, 153, 281–285. [Google Scholar] [CrossRef]
- Elliott, C.L.; Snyder, G.H. Autoclave-induced digestion for the colorimetric determination of silicon in rice straw. J. Agric. Food Chem. 1991, 39, 1118–1119. [Google Scholar] [CrossRef]
- Saito, K.; Yamamoto, A.; Sa, T.; Saigusa, M. Rapid, micro-methods to estimate plant silicon content by dilute hydrofluoric acid extraction and spectrometric molybdenum method. I. Silicon in rice plants and molybdenum yellow method. Soil Sci. Plant Nutr. 2005, 51, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Kraska, J.E.; Breitenbeck, G.A. Simple, Robust Method for Quantifying Silicon in Plant Tissue. Commun. Soil Sci. Plant Anal. 2010, 41, 2075–2085. [Google Scholar] [CrossRef]
- Hallmark, C.T.; Wilding, L.P.; Smeck, N.E. Silicon. In Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties, 2nd ed.; Agronomy monograph no. 9; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; The America Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982; pp. 263–273. [Google Scholar]
- Eaton, A.D.; Clesceri, L.S.; Greenberg, A.E. Standard Methods for the Examination of Water and Waste-Water, 19th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Jones, R.L.; Dreher, G.B. Silicon. In Methods of Soil Analysis, Part 3: Chemical Methods. Soil Science Society of America; Book series No. 5; Bigham, J.M., Ed.; The Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 627–638. [Google Scholar]
- Reidinger, S.; Ramsey, M.H.; Hartley, S.E. Rapid and accurate analyses of silicon and phosphorus in plants using a portable X-ray fluorescence spectrometer. New Phytol. 2012, 195, 699–706. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Lisztes-Szabó, Z.; Filep, A.F.; Csík, A.; Pető, Á.; Kertész, T.G.; Braun, M. pH-dependent silicon release from phytoliths of Norway spruce (Picea abies). J. Paleolimnol. 2019, 63, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Fraysse, F.; Pokrovsky, O.S.; Schott, J.; Meunier, J.D. Surface chemistry and reactivity of plant phytoliths in aqueous solu-tions. Chem. Geol. 2009, 258, 97–206. [Google Scholar] [CrossRef]
- Raturi, G.; Sharma, Y.; Rana, V.; Thakral, V.; Myaka, B.; Salvi, P.; Singh, M.; Dhar, H.; Deshmukh, R. Exploration of silicate solubilizing bacteria for sustainable agriculture and silicon biogeochemical cycle. Plant Physiol. Biochem. 2021, 166, 827–838. [Google Scholar] [CrossRef]
- Barker, W.W.; Welch, S.A.; Banfield, J.F. Biogeochemical weathering of silicate minerals. In Geomicrobiology: Interactions between Microbes and Minerals; Banfield, J.F., Nealson, K.H., Eds.; Mineralogical Society of America: Chantilly, VA, USA, 1997; Volume 35, pp. 391–428. ISBN 0-939950-45-6. [Google Scholar]
- Chen, W.; Yang, F.; Zhang, L.; Wang, J. Organic Acid Secretion and Phosphate Solubilizing Efficiency of Pseudomonas sp. PSB12: Effects of Phosphorus Forms and Carbon Sources. Geomicrobiol. J. 2016, 33, 870–877. [Google Scholar] [CrossRef]
- Lee, K.-E.; Adhikari, A.; Kang, S.-M.; You, Y.-H.; Joo, G.-J.; Kim, J.-H.; Kim, S.-J.; Lee, I.-J. Isolation and Characterization of the High Silicate and Phosphate Solubilizing Novel Strain Enterobacter ludwigii GAK2 that Promotes Growth in Rice Plants. Agronomy 2019, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Rogers, J.R.; Bennett, P.C. Mineral stimulation of subsurface microorganisms: Release of limiting nutrients from silicates. Chem. Geol. 2004, 203, 91–108. [Google Scholar] [CrossRef]
- Wang, R.R.; Wang, Q.; He, L.Y.; Qiu, G.; Sheng, X.F. Isolation and the interaction between a mineral-weathering Rhizobium tropici Q34 and silicate minerals. World J. Microbiol. Biotechnol. 2015, 31, 747–753. [Google Scholar] [CrossRef]
- Kang, S.-M.; Waqas, M.; Shahzad, R.; You, Y.-H.; Asaf, S.; Khan, M.A.; Lee, K.-E.; Joo, G.-J.; Kim, S.-J.; Lee, I.-J. Isolation and characterization of a novel silicate-solubilizing bacterial strain Burkholderia eburnea CS4-2 that promotes growth of japonica rice (Oryza sativa L. cv. Dongjin). Soil Sci. Plant Nutr. 2017, 63, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Kumawat, N.; Kumar, R.; Kumar, S.; Meena, V.S. Nutrient Solubilizing Microbes (NSMs): Its Role in Sustainable crop pro-duction. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V., Mishra, P., Bisht, J., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 25–61. [Google Scholar] [CrossRef]
- Chandrakala, C.; Voleti, S.R.; Bandeppa, S.; Kumar, N.S.; Latha, P.C. Silicate Solubilization and Plant Growth Promoting Potential of Rhizobium Sp. Isolated from Rice Rhizosphere. Silicon 2019, 11, 2895–2906. [Google Scholar] [CrossRef]
- Bist, V.; Niranjan, A.; Ranjan, M.; Lehri, A.; Seem, K.; Srivastava, S. Silicon-Solubilizing Media and Its Implication for Characterization of Bacteria to Mitigate Biotic Stress. Front. Plant Sci. 2020, 11, 28. [Google Scholar] [CrossRef]
- Vasanthi, N.; Saleena, L.M.; Raj, S.A. Silica Solubilization Potential of Certain Bacterial Species in the Presence of Different Silicate Minerals. Silicon 2016, 10, 267–275. [Google Scholar] [CrossRef]
- David, S.R.; Geoffroy, V.A. A Review of Asbestos Bioweathering by Siderophore-Producing Pseudomonas: A Potential Strategy of Bioremediation. Microorganisms 2020, 8, 1870. [Google Scholar] [CrossRef]
- Jones, L.H.P.; Handreck, K.A. Silica in Soils, Plants and Animals. Adv. Agron. 1967, 19, 107–149. [Google Scholar]
- Thummel, R.V.; Brightly, W.H.; Strömberg, C.A.E. Evolution of phytolith deposition in modern bryophytes, and implications for the fossil record and influence on silica cycle in early land plant evolution. New Phytol. 2018, 221, 2273–2285. [Google Scholar] [CrossRef]
- Kaur, H.; Greger, M. A Review on Si Uptake and Transport System. Plants 2019, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Hodson, M.; White, P.; Mead, A.; Broadley, M.R. Phylogenetic Variation in the Silicon Composition of Plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Ma, J.F. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivaraj, S.M.; Mandlik, R.; Bhat, J.A.; Raturi, G.; Elbaum, R.; Alexander, L.; Tripathi, D.K.; Deshmukh, R.; Sonah, H. Outstanding Questions on the Beneficial Role of Silicon in Crop Plants. Plant Cell Physiol. 2021, 63, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Ma, J.F. Linking transport system of silicon with its accumulation in different plant species. Soil Sci. Plant Nutr. 2020, 67, 10–17. [Google Scholar] [CrossRef]
- Faisal, S.; Callis, K.L.; Slot, M.; Kitajima, K. Transpiration-dependent passive silica accumulation in cucumber (Cucumis sativus) under varying soil silicon availability. Botany 2012, 90, 1058–1064. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Si, J.; Römheld, V. Silicon uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Wang, H.; Yu, C.; Fan, P.; Bao, B.; Li, T.; Zhu, Z. Identification of two cucumber putative silicon transporter genes in Cu-cumis sativus. J. Plant Growth Regul. 2015, 34, 332–338. [Google Scholar] [CrossRef]
- Deshmukh, R.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. HvLsi1 is a silicon influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of Maize Silicon Influx Transporters. Plant Cell Physiol. 2008, 50, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]
- Vivancos, J.; Deshmukh, R.; Grégoire, C.; Rémus-Borel, W.; Belzile, F.; Bélanger, R.R. Identification and characterization of silicon efflux transporters in horsetail (Equisetum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef]
- Sun, H.; Duan, Y.; Mitani-Ueno, N.; Che, J.; Jia, J.; Liu, J.; Guo, J.; Ma, J.F.; Gong, H. Tomato roots have a functional silicon influx transporter but not a functional silicon efflux transporter. Plant Cell Environ. 2019, 43, 732–744. [Google Scholar] [CrossRef]
- Liang, Y.; Hua, H.; Zhu, Y.; Zhang, J.; Cheng, C.; Römheld, V. Importance of plant species and external silicon concentration to active silicon uptake and transport. New Phytol. 2006, 172, 63–72. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A Transporter Regulating Silicon Distribution in Rice Shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Prychid, C.J.; Rudall, P.; Gregory, M. Systematics and Biology of Silica Bodies in Monocotyledons. Bot. Rev. 2003, 69, 377–440. [Google Scholar] [CrossRef]
- Lisztes-Szabó, Z. Complex environmental research: Do we need exact knowledge of plant anatomy? A critical discussion of Rashid et al. (2019). Earth-Sci. Rev. 2019, 198, 102920. [Google Scholar] [CrossRef]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Casey, W.H.; Kinrade, S.D.; Knight, C.T.G.; Rains, D.W.; Epstein, E. Aqueous silicate complexes in wheat, (Triticum aestivum L.). Plant Cell Environ. 2004, 27, 51–54. [Google Scholar] [CrossRef]
- Jarvis, S.C. The uptake and transport of silicon by perennial ryegrass and wheat. Plant Soil 1987, 97, 429–437. [Google Scholar] [CrossRef]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a beneficial element for crop plants. In Silicon in Agriculture; Datnoff, L., Snyder, G., Korndorfer, G., Eds.; Elsevier Science: New York, NY, USA, 2001; Volume 8, pp. 17–39. [Google Scholar]
- Raven, J.A. Cycling silicon—the role of accumulation in plants. New Phytol. 2003, 158, 419–421. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon Improves Water Use Efficiency in Maize Plants. J. Plant Nutr. 2005, 27, 1457–1470. [Google Scholar] [CrossRef]
- Kumar, S.; Adiram-Filiba, N.; Blum, S.; Sanchez-Lopez, J.A.; Tzfadia, O.; Omid, A.; Volpin, H.; Heifetz, Y.; Goobes, G.; Elbaum, R. Siliplant1 protein precipitates silica in sorghum silica cells. J. Exp. Bot. 2020, 71, 6830–6843. [Google Scholar] [CrossRef] [PubMed]
- Piperno, D.R. Phytolith Analysis. In An Archaeological and Geological Perspective; Academic Press: Cambridge, MA, USA; Harcourt Brace Jovanovich Publishers: San Diego, CA, USA, 1988; p. 280. ISBN 0-12-557175-5. [Google Scholar]
- Bartoli, F.; Wilding, L.P. Dissolution of Biogenic Opal as a Function of its Physical and Chemical Properties. Soil Sci. Soc. Am. J. 1980, 44, 873–878. [Google Scholar] [CrossRef]
- Carnelli, A.; Madella, M.; Theurillat, J.P.; Ammann, B. Aluminum in the opal silica reticule of phytoliths: A new tool in pal-aeoecological studies. Am. J. Bot. 2002, 89, 346–351. [Google Scholar] [CrossRef]
- Kameník, J.; Mizera, J.; Řanda, Z. Chemical composition of plant silica phytoliths. Environ. Chem. Lett. 2013, 11, 189–195. [Google Scholar] [CrossRef]
- Marschner, H.; Oberle, H.; Cakmak, I.; Römheld, V. Growth enhancement by silicon in cucumber (Cucumis sativus) plants depends on imbalance in phosphorus and zinc supply. Plant Soil 1990, 124, 211–219. [Google Scholar] [CrossRef]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Sangster, A.G. Silicon in the roots of higher plants. Am. J. Bot. 1978, 65, 929–935. [Google Scholar] [CrossRef]
- Hodson, M.J.; Sangster, A.G. Silica deposition in the inflorescence bracts of wheat (Triticum aestivum). II. X-ray microanalysis and backscattered electron imaging. Can. J. Bot. 1989, 67, 281–287. [Google Scholar] [CrossRef]
- Lisztes-Szabó, Z.; Kovács, S.; Pető, Á. Phytoliths analysis of Poa pratensis (Poaceae) leaves. Turk. J. Bot. 2014, 38, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R. Phytoliths. In A Comprehensive Guide for Archaeologists and Palaeoecologists; Altamira Press: New York, NY, USA, 2006; p. 248. ISBN 978-0759103849. [Google Scholar]
- Pethő, Á. Phytolith analysis of modern soil profile as a tool to demonstrate land-use changes and anthropogenic impacts. Cereal Res. Commun. 2008, 36, 955–958. [Google Scholar]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Hunt, J.W.; Dean, A.P.; Webster, R.E.; Johnson, G.N.; Ennos, A.R. A Novel Mechanism by which Silica Defends Grasses Against Herbivory. Ann. Bot. 2008, 102, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Cherif, M.; Asselin, A.; Belanger, R.R. Defense responses induced by soluble silicon in cucumber roots infected by Pythium spp. Phytopathology 1994, 84, 236–242. [Google Scholar] [CrossRef]
- Parr, J.; Sullivan, L. Soil carbon sequestration in phytoliths. Soil Biol. Biochem. 2005, 37, 117–124. [Google Scholar] [CrossRef]
- Rajendiran, S.; Coumar, V.; Kundu, S.; Dotaniya, M.L.; Rao, A.S. Role of phytolith occluded carbon of crop plants for en-hancing soil carbon sequestration in agro-ecosystems. Curr. Sci. 2012, 103, 911–920. [Google Scholar]
- Ning, D.; Song, A.; Fan, F.; Li, Z.; Liang, Y. Effects of Slag-Based Silicon Fertilizer on Rice Growth and Brown-Spot Resistance. PLoS ONE 2014, 9, e102681. [Google Scholar] [CrossRef] [Green Version]
- Meena, V.D.; Dotaniya, M.L.; Coumar, V.; Rajendiran, S.; Kundu, S.; Rao, A.S. A Case for Silicon Fertilization to Improve Crop Yields in Tropical Soils. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2013, 84, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Korndörfer, G.H.; Lepsch, I. Effect of silicon on plant growth and crop yield. In Silicon in Agriculture, Studies in Plant Science 8, 1st ed.; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 133–147. [Google Scholar]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis, and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of Kentucky bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Manivannan, A.; Soundararajan, P.; Jeong, B.R. Physiological and Proteomic Analysis in Chloroplasts of Solanum lycopersicum L. under Silicon Efficiency and Salinity Stress. Int. J. Mol. Sci. 2014, 15, 21803–21824. [Google Scholar] [CrossRef] [PubMed]
- Moussa, H.; Galad, M. Comparative response of salt tolerant and salt sensitive maize (Zea mays L.) cultivars to silicon. Eur. J. Appl. Econ. 2015, 2, 1–5. [Google Scholar]
- Du, Q.S.; Fan, X.W.; Wang, C.H.; Huang, R.B. A Possible CO2 Conducting and Concentrating Mechanism in Plant Stomata SLAC1 Channel. PLoS ONE 2011, 6, e24264. [Google Scholar] [CrossRef]
- Tian, W.; Hou, C.; Ren, Z.; Pan, Y.; Jia, J.; Zhang, H.; Bai, F.; Zhang, P.; Zhu, Y.H.; Luo, L.; et al. A molecular pathway for CO2 response in Arabidopsis guard cells. Nat. Commun. 2015, 6, 6057. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, M.; Mueller, H.M.; Bauer, H.; Peirats-Llobet, M.; Rodriguez, P.L.; Geilfus, C.-M.; Carpentier, S.C.; Al Rasheid, K.A.S.; Kollist, H.; Merilo, E.; et al. The role of Arabidopsis ABA receptors from the PYR/PYL/RCAR family in stomatal acclimation and closure signal integration. Nat. Plants 2019, 26, 1002–1011. [Google Scholar] [CrossRef]
- Rastogi, A.; Yadav, S.; Hussain, S.; Kataria, S.; Hajihashemi, S.; Kumari, P.; Yang, X.; Brestic, M. Does silicon really matter for the photosynthetic machinery in plants…? Plant Physiol. Biochem. 2021, 169, 40–48. [Google Scholar] [CrossRef]
- Sánchez-Navarro, J.F.; González-García, Y.; Benavides-Mendoza, A.; Morales-Díaz, A.B.; González-Morales, S.; Cadenas-Pliego, G.; García-Guillermo, M.D.S.; Juárez-Maldonado, A. Silicon Nanoparticles Improve the Shelf Life and Antioxidant Status of Lilium. Plants 2021, 10, 2338. [Google Scholar] [CrossRef]
- Schaller, J.; Puppe, D.; Kaczorek, D.; Ellerbrock, R.; Sommer, M. Silicon Cycling in Soils Revisited. Plants 2021, 10, 295. [Google Scholar] [CrossRef]
- Pereira, H.S.; Korndörfer, G.H.; Moura, W.F.; Corrêa, G.F. Extratores de silício disponível em escórias e fertilizantes. Rev. Bras. Cienc. Solo 2003, 27, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Buck, G.B.; Korndörfer, G.H.; Datnoff, L.E. Extractors for estimating plant available silicon from potential silicon fertilizer sources. J. Plant Nutr. 2010, 34, 272–282. [Google Scholar] [CrossRef]
- Harley, A.; Gilkes, R. Factors influencing the release of plant nutrient elements from silicate rock powders: A geochemical overview. Nutr. Cycl. Agroecosyst. 2000, 56, 11–36. [Google Scholar] [CrossRef]
- Makabe-Sasaki, S.; Kakuda, K.-I.; Sasaki, Y.; Ando, H. Effects of slag silicate fertilizer on silicon content of rice plants grown in paddy fields on the Shounai Plain, Yamagata, Japan. Soil Sci. Plant Nutr. 2014, 60, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Kim, S.Y.; Villamil, M.B.; Pramanik, P.; Hong, C.O.; Kim, P.J. Different response of silicate fertilizer having electron acceptors on methane emission in rice paddy soil under green manuring. Biol. Fertil. Soils 2011, 48, 435–442. [Google Scholar] [CrossRef]
- Song, A.; Ning, D.; Fan, F.; Li, Z.; Provance-Bowley, M.; Liang, Y. The potential for carbon bio-sequestration in China’s paddy rice (Oryza sativa L.) as impacted by slag-based silicate fertilizer. Sci. Rep. 2015, 5, 17354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocharnikova, E.A.; Loginov, S.V.; Matychenkov, V.V.; Storozhenko, P.A. Silicon fertilizer efficiency. Russ. Agric. Sci. 2010, 36, 446–448. [Google Scholar] [CrossRef]
- Agostinho, F.B.; Tubana, B.S.; Martins, M.S.; Datnoff, L.E. Effect of Different Silicon Sources on Yield and Silicon Uptake of Rice Grown under Varying Phosphorus Rates. Plants 2017, 6, 35. [Google Scholar] [CrossRef]
- Cook, G.W. The effect of some silicate slags on the utilization of soil and fertilizer phosphorous. J. Sci. Food Agric. 1956, 7, 56–61. [Google Scholar] [CrossRef]
- El-Ramady, H.; Verma, K.K.; Rajput, V.D.; Minkina, T.; Elbehery, F.; Elbasiony, H.; Elsakhawy, T.; Omara, A.E.-D.; Amer, M. Chapter 1—Sources of silicon and nano-silicon in soils and plants. In Silicon and Nano-silicon in Environmental Stress Management and Crop Quality Improvement; Etesami, H., Al Saeedi, A.H., El-Ramady, H., Fujita, M., Pessarakli, M., Hossain, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 1–15. [Google Scholar] [CrossRef]
- Abro, S.A.; Qureshi, R.; Soomro, F.M.; Mirbahar, A.A.; Jakhar, G.S. Effects of silicon levels on growth and yield of wheat in silty loam soil. Pak. J. Bot. 2009, 41, 1385–1390. [Google Scholar]
- Neu, S.; Schaller, J.; Dudel, E.G. Silicon availability modifies nutrient use efficiency and content, C:N:P stoichiometry, and productivity of winter wheat (Triticum aestivum L.). Sci. Rep. 2017, 7, 40829. [Google Scholar] [CrossRef]
- Xie, Z.; Song, F.; Xu, H.; Shao, H.; Song, R. Effects of Silicon on Photosynthetic Characteristics of Maize (Zea mays L.) on Alluvial Soil. Sci. World J. 2014, 2014, 718716. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of Silicon Fertilization on Maize Performance Under Limited Water Supply. Silicon 2016, 10, 177–183. [Google Scholar] [CrossRef]
- Shwethakumari, U.; Prakash, N. Effect of Foliar Application of Silicic Acid on Soybean Yield and Seed Quality under Field Conditions. J. Indian Soc. Soil Sci. 2018, 66, 406. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, Y.; Chu, G. Applying silicate fertilizer increases both yield and quality of table grape (Vitis vinifera L.) grown on calcareous grey desert soil. Sci. Hortic. 2017, 225, 757–763. [Google Scholar] [CrossRef]
- Radkowski, A.; Sosin-Bzducha, E.; Radkowska, I. Effects of silicon foliar fertilization of meadow plants on the nutritional value of silage fed to dairy cows. J. Elem. 2017, 22, 1311–1322. [Google Scholar] [CrossRef]
- Jayawardana, H.A.R.K.; Weerahewa, H.L.D.; Saparamadu, M.D.J.S. Effect of root or foliar application of soluble silicon on plant growth Trop. Agric. Res. 2014, 26, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Muljani, S.; Wahyudi, B.; Sumada, K. Potassium Silicate Foliar Fertilizer Grade from Geothermal Sludge and Pyrophyllite. In Proceedings of the MATEC Web of Conferences, Amsterdam, The Netherlands, 23–25 March 2016; Volume 58, p. 1021. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Liu, M. Slow-Release Potassium Silicate Fertilizer with the Function of Superabsorbent and Water Retention. Ind. Eng. Chem. Res. 2007, 46, 6494–6500. [Google Scholar] [CrossRef]
- Yao, Y.; Yoneyama, T.; Hayashi, H. Potassium uptake by Chinese cabbage (Brassica pekinensis Rupy.) from fused potassium silicate, a slow-releasing fertilizer. Plant Soil 2003, 249, 279–286. [Google Scholar] [CrossRef]
- Górecki, R.; Danielski-Busch, W. Effect of silicate fertilizers on yielding of greenhouse cucumber (Cucumis sativus L.) in container culitivation. J. Elementol. 2009, 14, 71–77. [Google Scholar] [CrossRef]
- Elawad, S.H.; Street, J.J.; Gascho, G.J. Response of Sugarcane to Silicate Source and Rate. II. Leaf Freckling and Nutrient Content. Agron. J. 1982, 74, 484–487. [Google Scholar] [CrossRef]
- Anderson, D.L. Soil and leaf nutrient interactions following application of calcium silicate slag to sugarcane. Fertil. Res. 1991, 30, 9–18. [Google Scholar] [CrossRef]
- Bokhtiar, S.M.; Huang, H.-R.; Li, Y.-R. Response of Sugarcane to Calcium Silicate on Yield, Gas Exchange Characteristics, Leaf Nutrient Concentrations, and Soil Properties in Two Different Soils. Commun. Soil Sci. Plant Anal. 2012, 43, 1363–1381. [Google Scholar] [CrossRef]
- Zargar, S.M.; Mahajan, R.; Bhat, J.A.; Nazir, M.; Deshmukh, R. Role of silicon in plant stress tolerance: Opportunities to achieve a sustainable cropping system. 3 Biotech 2019, 9, 73. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.-P.; Tian, D.-D.; Guo, D.-J.; Chen, Z.-L.; Zhong, C.-S.; Nikpay, A.; Singh, M.; Rajput, V.D.; Singh, R.K.; et al. Influence of Silicon on Biocontrol Strategies to Manage Biotic Stress for Crop Protection, Performance, and Improvement. Plants 2021, 10, 2163. [Google Scholar] [CrossRef]
- Hu, A.Y.; Xu, S.N.; Ni Qin, D.; Li, W.; Zhao, X.Q. Role of Silicon in Mediating Phosphorus Imbalance in Plants. Plants 2020, 10, 51. [Google Scholar] [CrossRef]
- Galindo, F.; Pagliari, P.; Rodrigues, W.; Fernandes, G.; Boleta, E.; Santini, J.; Jalal, A.; Buzetti, S.; Lavres, J.; Filho, M.T. Silicon Amendment Enhances Agronomic Efficiency of Nitrogen Fertilization in Maize and Wheat Crops under Tropical Conditions. Plants 2021, 10, 1329. [Google Scholar] [CrossRef]
- Réthoré, E.; Ali, N.; Yvin, J.-C.; Hosseini, S.A. Silicon Regulates Source to Sink Metabolic Homeostasis and Promotes Growth of Rice Plants under Sulfur Deficiency. Int. J. Mol. Sci. 2020, 21, 3677. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Rad, S.N.; Ali, N.; Yvin, J.-C. The Ameliorative Effect of Silicon on Maize Plants Grown in Mg-Deficient Conditions. Int. J. Mol. Sci. 2019, 20, 969. [Google Scholar] [CrossRef] [Green Version]
- Lozano-González, J.M.; Valverde, C.; Hernández, C.D.; Martin-Esquinas, A.; Hernández-Apaolaza, L. Beneficial Effect of Root or Foliar Silicon Applied to Cucumber Plants under Different Zinc Nutritional Statuses. Plants 2021, 10, 2602. [Google Scholar] [CrossRef]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Saeed, D.A.; Nawaz, M.A.; Mehmood, S.; Tu, S. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manag. 2016, 183, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, N.; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Zaman, Q.U.; Rashid, M.; Nawaz, R.; Hussain, A.; Ashraf, K.; Latif, M.; Heile, A.O.; Mehmood, F.; Salahuddin, S.; Chen, Y. Silicon Fertilization: A Step towards Cadmium-Free Fragrant Rice. Plants 2021, 10, 2440. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zou, Q.; Xue, S.; Mo, J.; Pan, W.; Lou, L.; Wong, M.H. Effects of silicon (Si) on arsenic (As) accumulation and speciation in rice (Oryza sativa L.) genotypes with different radial oxygen loss (ROL). Chemosphere 2015, 138, 447–453. [Google Scholar] [CrossRef]
- Ali, N.; Réthoré, E.; Yvin, J.-C.; Hosseini, S.A. The Regulatory Role of Silicon in Mitigating Plant Nutritional Stresses. Plants 2020, 9, 1779. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The Effect of Silicon on Photosynthesis and Expression of Its Relevant Genes in Rice (Oryza sativa L.) under High-Zinc Stress. PLoS ONE 2014, 9, e113782. [Google Scholar] [CrossRef]
- Ahmad, M.; El-Saeid, M.H.; Akram, M.A.; Ahmad, H.R.; Haroon, H.; Hussain, A. Silicon fertilization—A tool to boost up drought tolerance in wheat (Triticum aestivum L.) crop for better yield. J. Plant Nutr. 2015, 39, 1283–1291. [Google Scholar] [CrossRef]
- Ciecierski, W. Effect of silicon on biotic and abiotic stress mitigation in horticultural and field crops. In Proceedings of the In-ternational Symposium “Mikroelementy w rolnictwie i’srodowisku”, Kudowa-Zdrój, Poland, 21–24 June 2016; p. 25. [Google Scholar]
- Kowalska, J.; Tyburski, J.; Bocianowski, J.; Krzymińska, J.; Matysiak, K. Methods of Silicon Application on Organic Spring Wheat (Triticum aestivum L. spp. vulgare) Cultivars Grown across Two Contrasting Precipitation Years. Agronomy 2020, 10, 1655. [Google Scholar] [CrossRef]
- Parveen, A.; Mumtaz, S.; Saleem, M.H.; Hussain, I.; Perveen, S. Thind, Chapter 11—Silicon and nanosilicon mediated heat stress tolerance in plants. In Silicon and Nano-Silicon in Environmental Stress Management and Crop Quality Improvement; Etesami, H., Al Saeedi, A.H., El-Ramady, H., Fujita, M., Pessarakli, M., Hossain, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 153–159. [Google Scholar] [CrossRef]
- Thorne, S.; Hartley, S.; Maathuis, F. The Effect of Silicon on Osmotic and Drought Stress Tolerance in Wheat Landraces. Plants 2021, 10, 814. [Google Scholar] [CrossRef]
- Kutasy, E.; Buday-Bódi, E.; Virág, I.C.; Forgács, F.; Melash, A.A.; Zsombik, L.; Nagy, A.; Csajbók, J. Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants 2021, 11, 30. [Google Scholar] [CrossRef]
- Ning, D.; Qin, A.; Liu, Z.; Duan, A.; Xiao, J.; Zhang, J.; Liu, Z.; Zhao, B.; Liu, Z. Silicon-Mediated Physiological and Agronomic Responses of Maize to Drought Stress Imposed at the Vegetative and Reproductive Stages. Agronomy 2020, 10, 1136. [Google Scholar] [CrossRef]
- Crusciol, C.A.C.; Pulz, A.L.; Lemos, L.B.; Soratto, R.P.; Lima, G.P.P. Effects of Silicon and Drought Stress on Tuber Yield and Leaf Biochemical Characteristics in Potato. Crop Sci. 2009, 49, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Gugała, M.; Sikorska, A.; Zarzecka, K.; Kapela, K.; Mystkowska, I. The effect of sowing method and biostimulators on au-tumn development and overwintering of winter rape. Acta Sci. Pol. Agric. 2017, 16, 111–120. [Google Scholar]
- Saja-Garbarz, D.; Ostrowska, A.; Kaczanowska, K.; Janowiak, F. Accumulation of Silicon and Changes in Water Balance under Drought Stress in Brassica napus var. napus L. Plants 2021, 10, 280. [Google Scholar] [CrossRef]
- Čermelj, A.M.; Golob, A.; Vogel-Mikuš, K.; Germ, M. Silicon Mitigates Negative Impacts of Drought and UV-B Radiation in Plants. Plants 2021, 11, 91. [Google Scholar] [CrossRef]
- Sabir, F.; Noreen, S.; Malik, Z.; Kamran, M.; Riaz, M.; Dawood, M.; Parveen, A.; Afzal, S.; Ahmad, I.; Ali, M. Chapter 8—Silicon improves salinity tolerance in crop plants: Insights into photosynthesis, defense system, and production of phytohormones. In Silicon and Nano-Silicon in Environmental Stress Management and Crop Quality Improvement; Etesami, H., Al Saeedi, A.H., El-Ramady, H., Fujita, M., Pessarakli, M., Hossain, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 91–103. [Google Scholar] [CrossRef]
- Liu, B.; Soundararajan, P.; Manivannan, A. Mechanisms of Silicon-Mediated Amelioration of Salt Stress in Plants. Plants 2019, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A.A.; Mazrou, Y.S.; Hafez, Y.M. Silicon Foliar Application Mitigates Salt Stress in Sweet Pepper Plants by Enhancing Water Status, Photosynthesis, Antioxidant Enzyme Activity and Fruit Yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Gong, H.-J.; Yin, J.-L. Role of Silicon in Mediating Salt Tolerance in Plants: A Review. Plants 2019, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Schaller, J.; Heimes, R.; Ma, J.F.; Meunier, J.-D.; Shao, J.F.; Fujii-Kashino, M.; Knorr, K.H. Silicon accumulation in rice plant aboveground biomass affects leaf carbon quality. Plant Soil 2019, 444, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Sun, Y.; Wang, H.; Wang, H. Advances in Understanding Silicon Transporters and the Benefits to Silicon-Associated Disease Resistance in Plants. Appl. Sci. 2022, 12, 3282. [Google Scholar] [CrossRef]
- Buck, G.B.; Korndörfer, G.H.; Nolla, A.; Coelho, L. Potassium Silicate as Foliar Spray and Rice Blast Control. J. Plant Nutr. 2008, 31, 231–237. [Google Scholar] [CrossRef]
- Datnoff, L.; Deren, C.; Snyder, G. Silicon fertilization for disease management of rice in Florida. Crop Prot. 1997, 16, 525–531. [Google Scholar] [CrossRef]
- Ciecierski, W.; Korbas, M.; Horoszkiewicz-Janka, J. Effectiveness of silicon application on mycotoxins reduction in maize. In Proceedings of the 7th International Conference on Silicon in Agriculture, Bengaluru, India, 24–28 October 2017; p. 96. [Google Scholar]
- Win, K.; Maeda, S.; Kobayashi, M.; Jiang, C.-J. Silicon Enhances Resistance to Red Crown Rot Caused by Calonectria ilicicola in Soybean. Agronomy 2021, 11, 899. [Google Scholar] [CrossRef]
- Njenga, K.W.; Nyaboga, E.; Wagacha, J.M.; Mwaura, F.B. Silicon Induces Resistance to Bacterial Blight by Altering the Physiology and Antioxidant Enzyme Activities in Cassava. World J. Agric. Res. 2017, 5, 42–51. [Google Scholar] [CrossRef]
- Katz, O. Silicon and Plant–Animal Interactions: Towards an Evolutionary Framework. Plants 2020, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.R.; Dagg, V.; Waterman, J.M.; Johnson, S.N. Silicon Alters Leaf Surface Morphology and Suppresses Insect Herbivory in a Model Grass Species. Plants 2020, 9, 643. [Google Scholar] [CrossRef]
- Cotterill, J.V.; Watkins, R.W.; Brennon, C.B.; Cowan, D.P. Boosting silica levels in wheat leaves reduces grazing by rabbits. Pest Manag. Sci. 2006, 63, 247–253. [Google Scholar] [CrossRef]
- Alhousari, F.; Greger, M. Silicon and Mechanisms of Plant Resistance to Insect Pests. Plants 2018, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Fawe, A.; Menzies, J.G.; Chérif, M.; Bélanger, R.R. Chapter 9 Silicon and disease resistance in dicotyledons. In Studies in Plant Science; Elsevier: Amsterdam, The Netherlands, 2001; pp. 159–169. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Zakharenko, A.M.; Zemchenko, I.V.; Haider, M.S.; Ali, M.A.; Imtiaz, M.; Chung, G.; Tsatsakis, A.; Sun, S.; Golokhvast, K.S. Phytolith Formation in Plants: From Soil to Cell. Plants 2019, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Parrella, M.; Costamagna, T.; Kaspi, R. The addition of potassium silicate to the fertilizer mix to suppress liriomyza leafminers attacking chrysanthemums. Acta Hortic. 2007, 747, 365–370. [Google Scholar] [CrossRef]
- Almeida, G.D.; Pratissoli, D.; Zanuncio, J.C.; Vicentini, V.B.; Holtz, A.M.; Serrão, J.E. Calcium silicate and organic mineral fertilizer increase the resistance of tomato plants to Frankliniella schultzei. Phytoparasitica 2009, 37, 225–230. [Google Scholar] [CrossRef]
- Hou, M.; Han, Y. Silicon-mediated rice plant resistance to the Asiatic rice borer (Lepidoptera: Crambidae): Effects of silicon amendment and rice varietal resistance. J. Econ. Entomol. 2010, 103, 1412–1419. [Google Scholar] [CrossRef]
- França, L.L.; Dierings, C.A.; Almeida, A.C.D.S.; Araújo, M.D.S.; Heinrichs, E.A.; Da Silva, A.R.; Barrigossi, J.A.F.; De Jesus, F.G. Resistance in Rice to Tibraca limbativentris (Hemiptera: Pentatomidae) Influenced by Plant Silicon Content. Fla. Entomol. 2018, 101, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Jeer, M.; Suman, K.; Maheswari, T.U.; Voleti, S.; Padmakumari, A. Rice husk ash and imidazole application enhances silicon availability to rice plants and reduces yellow stem borer damage. Field Crop. Res. 2018, 224, 60–66. [Google Scholar] [CrossRef]
- Villegas, J.M.; Way, M.O.; Pearson, R.A.; Stout, M.J. Integrating Soil Silicon Amendment into Management Programs for Insect Pests of Drill-Seeded Rice. Plants 2017, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Radkowski, A.; Radkowska, I. Effects of Silicate Fertilizer on Seed Yield in Timothy-Grass (Phleum pratense L.). Ecol. Chem. Eng. S 2018, 25, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Mir, R.A.; Bhat, B.A.; Yousuf, H.; Islam, S.T.; Raza, A.; Rizvi, M.A.; Charagh, S.; Albaqami, M.; Sofi, P.A.; Zargar, S.M. Multidimensional Role of Silicon to Activate Resilient Plant Growth and to Mitigate Abiotic Stress. Front. Plant Sci. 2022, 13, 819658. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.; Reynolds, O.L.; Gurr, G. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [Green Version]
- Meunier, J.D.; Barboni, D.; Anwar-Ul-Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J.; et al. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Wallis, C.M.; Uddin, W. Silicon-Induced Systemic Defense Responses in Perennial Ryegrass Against Infection by Magnaporthe oryzae. Phytopathology 2015, 105, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Zhao, W.; Zhu, X. Silicon improves photosynthesis and strengthens enzyme activities in the C 3 succulent xerophyte Zygophyllum xanthoxylum under drought stress. J. Plant Physiol. 2016, 199, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, J.; Samardžić, J.; Kostic, L.; Laursen, K.H.; Natić, M.; Timotijevic, G.; Schjoerring, J.K.; Nikolic, M. Silicon enhances leaf remobilization of iron in cucumber under limited iron conditions. Ann. Bot. 2016, 118, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to Promote Direct and Indirect Effects on Plant Defense Against Arthropod Pests in Agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Leroy, N.; de Tombeur, F.; Walgraffe, Y.; Cornélis, J.-T.; Verheggen, F.J. Silicon and Plant Natural Defenses against Insect Pests: Impact on Plant Volatile Organic Compounds and Cascade Effects on Multitrophic Interactions. Plants 2019, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Puppe, D.; Kaczorek, D.; Schaller, J. Chapter 22—Biological Impacts on Silicon Availability and Cycling in Agricultural Plant-Soil Systems; Etesami, H., Al Saeedi, A.H., El-Ramady, H., Fujita, M., Pessarakli, M., Hossain, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 309–324. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, S.; Kutasy, E.; Csajbók, J. The Multiple Role of Silicon Nutrition in Alleviating Environmental Stresses in Sustainable Crop Production. Plants 2022, 11, 1223. https://doi.org/10.3390/plants11091223
Kovács S, Kutasy E, Csajbók J. The Multiple Role of Silicon Nutrition in Alleviating Environmental Stresses in Sustainable Crop Production. Plants. 2022; 11(9):1223. https://doi.org/10.3390/plants11091223
Chicago/Turabian StyleKovács, Szilvia, Erika Kutasy, and József Csajbók. 2022. "The Multiple Role of Silicon Nutrition in Alleviating Environmental Stresses in Sustainable Crop Production" Plants 11, no. 9: 1223. https://doi.org/10.3390/plants11091223