
Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
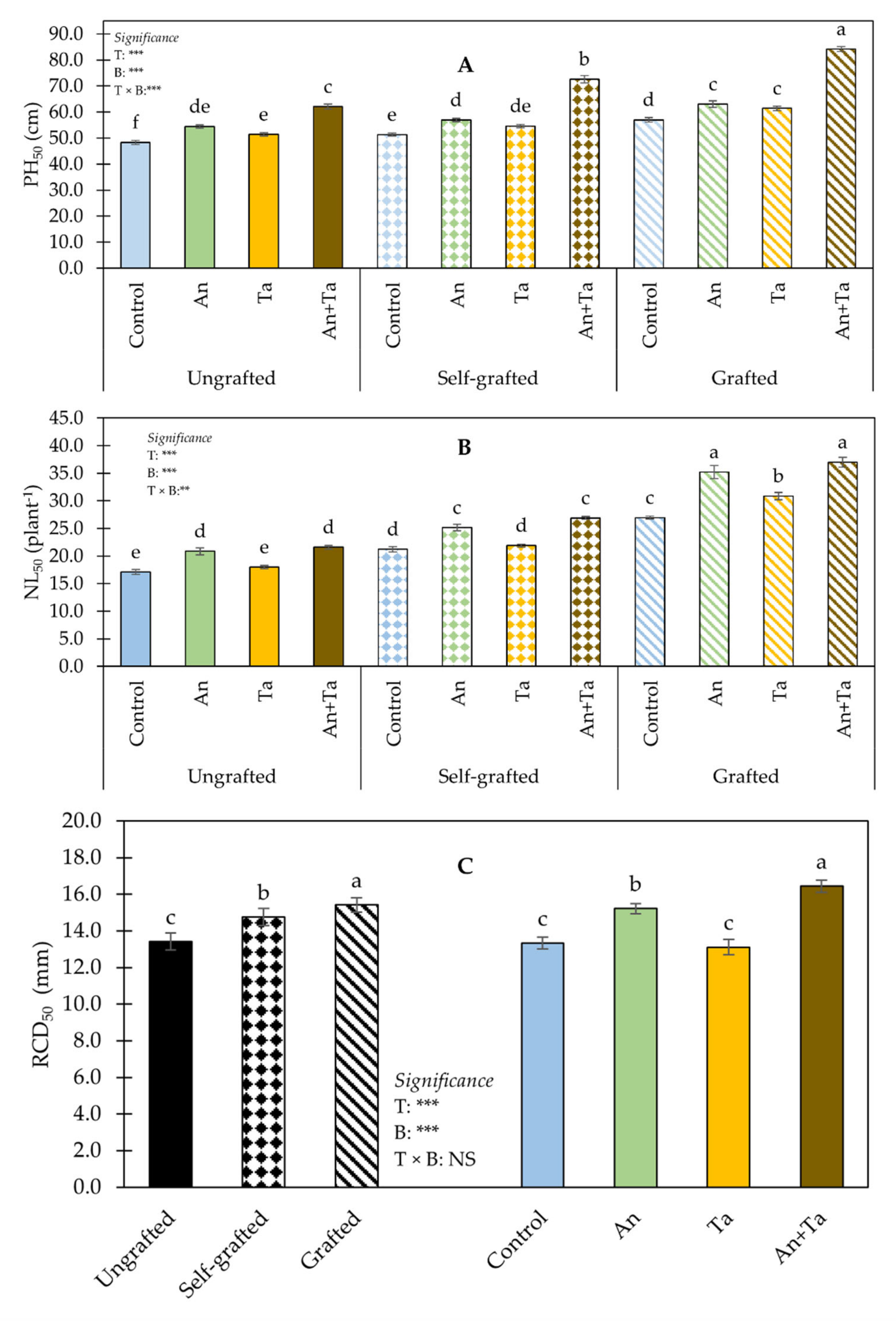
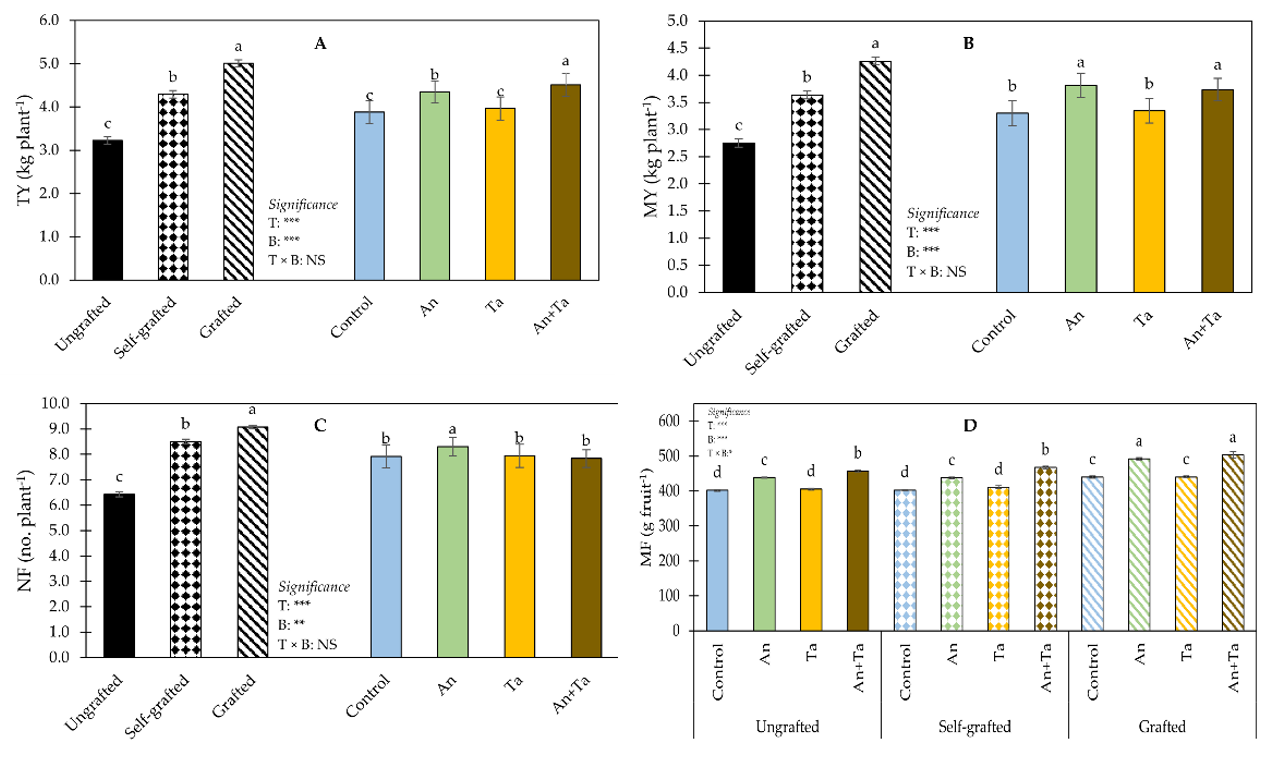
2.1. Plant Performance and Production
2.2. Nutritional and Functional Fruit Traits
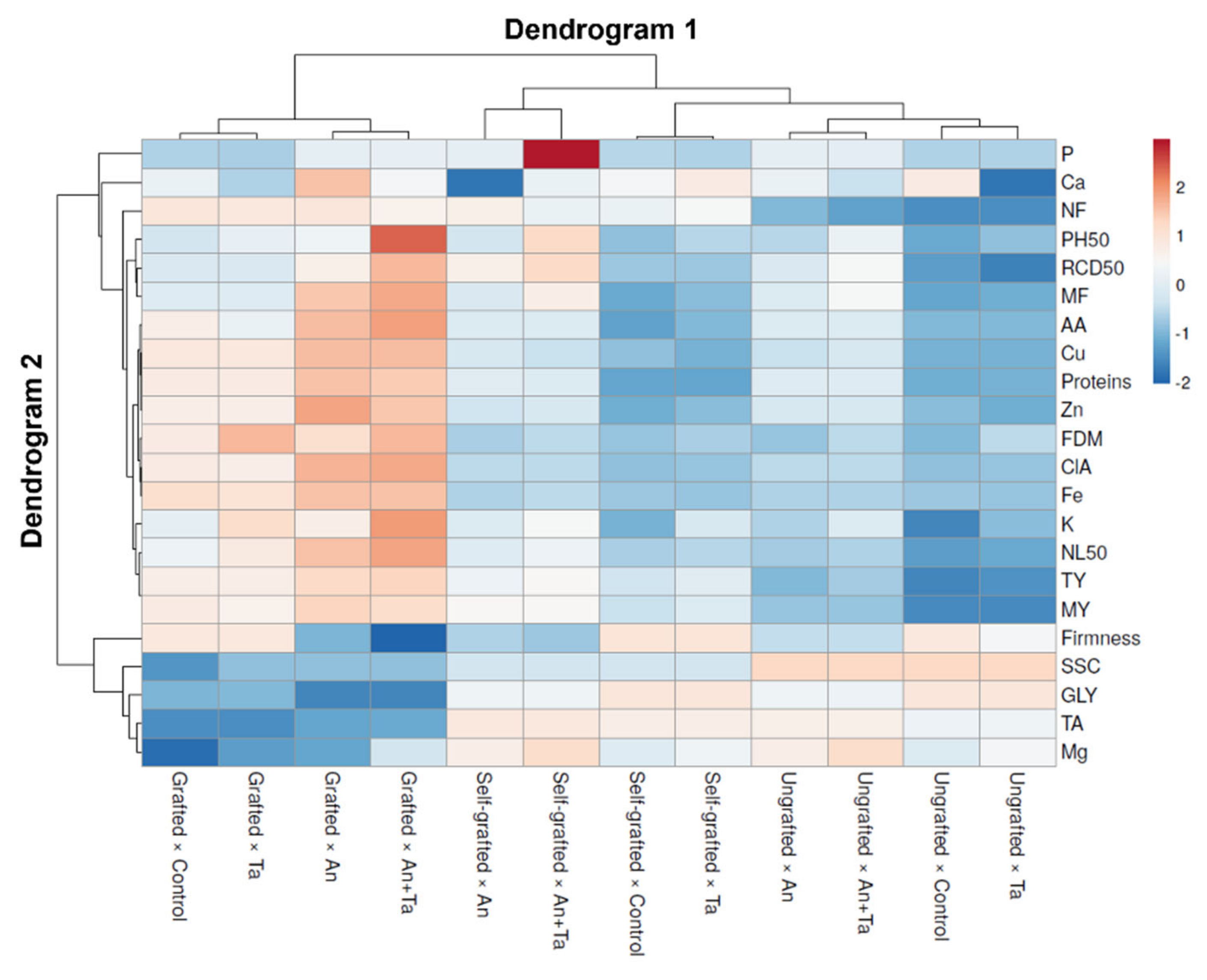
2.3. Heat Map Analysis of All Eggplant Traits
3. Discussion
4. Materials and Methods
4.1. Experimental Site, Plant Material, and Cultivation Conditions
4.2. Biostimulants Application
4.3. Plant Growth, Yield, and Yield Components
4.4. Fruit Quality Traits
4.5. Proteins and Mineral Profile in Fruit Tissue
4.6. Experimental Design and Statistics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. Global Agriculture towards 2050, High-Level Expert Forum, How to Feed the World 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- FAO. The Future of Food and Agriculture—Alternative Pathways to 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Lassaletta, L.; Billen, G.; Garnier, J.; Bouwman, L.; Velazquez, E.; Mueller, N.D.; Gerber, J.S. Nitrogen use in the global food system: Past trends and future trajectories of agronomic performance, pollution, trade, and dietary demand. Environ. Res. Lett. 2016, 11, 095007. [Google Scholar] [CrossRef]
- Savci, S. Investigation of Effect of Chemical Fertilizers on Environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Miceli, A.; Vetrano, F.; Sabatino, L.; D’Anna, F.; Moncada, A. Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket. Horticulturae 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- La Bella, S.; Virga, G.; Iacuzzi, N.; Licata, M.; Sabatino, L.; Consentino, B.B.; Leto, C.; Tuttolomondo, T. Effects of Irrigation, Peat-Alternative Substrate and Plant Habitus on the Morphological and Production Characteristics of Sicilian Rosemary (Rosmarinus officinalis L.) Biotypes Grown in Pot. Agriculture 2021, 11, 13. [Google Scholar] [CrossRef]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; Consentino, B.B.; Rouphael, Y. Selenium Biofortification and Grafting Modulate Plant Performance and Functional Features of Cherry Tomato Grown in a Soilless System. Sci. Hortic. 2021, 285, 110095. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein Hydrolysates and Mo-Biofortification Interactively Modulate Plant Performance and Quality of ‘Canasta’ Lettuce Grown in a Protected Environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 196, 1–134. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Consentino, B.B.; Virga, G.; La Placa, G.G.; Sabatino, L.; Rouphael, Y.; Ntatsi, G.; Iapichino, G.; La Bella, S.; Mauro, R.P.; D’Anna, F.; et al. Celery (Apium graveolens L.) Performances as Subjected to Different Sources of Protein Hydrolysates. Plants 2020, 9, 1633. [Google Scholar] [CrossRef]
- Consentino, B.B.; Sabatino, L.; Mauro, R.P.; Nicoletto, C.; De Pasquale, C.; Iapichino, G.; La Bella, S. Seaweed Extract Improves Lagenaria siceraria Young Shoot Production, Mineral Profile and Functional Quality. Horticulturae 2021, 7, 549. [Google Scholar] [CrossRef]
- Di Mola, I.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, L.; Rouphael, Y.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- La Bella, S.; Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Sabatino, L. Impact of Ecklonia maxima Seaweed Extract and Mo Foliar Treatments on Biofortification, Spinach Yield, Quality and NUE. Plants 2021, 10, 1139. [Google Scholar] [CrossRef] [PubMed]
- Consentino, B.B.; Aprile, S.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Alibrandi, P.; Sabatino, L. Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agronomy 2022, 12, 236. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Woo, S.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Stewart, A.; Hill, R. Applications of Trichoderma in plant growth promotion. In Biotechnology and Biology of Trichoderma; Chapter 31; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 415–428. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Bletsos, F.; Thanassoulopoulos, C.; Roupakias, D. Effect of Grafting on Growth, Yield, and Verticillium Wilt of Eggplant. HortScience 2003, 38, 183–186. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Rotino, G.L.; Palazzolo, E.; Mennella, G.; D’Anna, F. Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy 2019, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Kumar, P.; Kumar, A.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Grafting Tomato as a Tool to Improve Salt Tolerance. Agronomy 2020, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; Iapichino, G.; Consentino, B.B.; D’Anna, F.; Rouphael, Y. Rootstock and Arbuscular Mycorrhiza Combinatorial Effects on Eggplant Crop Performance and Fruit Quality under Greenhouse Conditions. Agronomy 2020, 10, 693. [Google Scholar] [CrossRef]
- Turhan, A.; Ozmen, N.; Serbeci, M.S.; Seniz, V. Effects of Grafting on Different Rootstocks on Tomato Fruit Yield and Quality. Hortic. Sci. 2011, 38, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Daunay, M.C. Eggplant. In Handbook of Plant Breeding-Vegetables II; Fabeceae, Liliaceae, Solanaceae and Umbrelliferae; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 163–220. [Google Scholar]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping Quantitative Trait Loci Affecting Biochemical and Morphological Fruit Properties in Eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Tesi, R. Orticoltura Mediterranea Sostenibile; Pàtron Editore: Bologna, Italy, 2010. [Google Scholar]
- D’Anna, F.; Sabatino, L. Morphological and agronomical characterization of eggplant genetic resources from the Sicily area. J. Food Agri. Environ. 2013, 11, 401–404. [Google Scholar]
- Woo, S.L.; Scala, F.; Ruocco, M.; Lorito, M. The molecular biology of the interactions between Trichoderma spp., phytopathogenic fungi, and plants. Phytopathology 2006, 96, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manganiello, G.; Sacco, A.; Ercolano, M.R.; Vinale, F.; Lanzuise, S.; Pascale, A.; Napolitano, M.; Lombardi, N.; Lorito, M.; Woo, S.L. Modulation of tomato response to Rhizoctonia solani by Trichoderma harzianum and its secondary metabolite harzianic acid. Front. Microbiol. 2018, 9, 1966. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatino, L.; Iapichino, G.; D’Anna, F.; Palazzolo, E.; Mennella, G.; Rotino, G.L. Hybrids and allied species as potential rootstocks for eggplant: Effect of grafting on vigour, yield and overall fruit quality traits. Sci. Hortic. 2018, 228, 81–90. [Google Scholar] [CrossRef]
- Mauro, R.P.; Agnello, M.; Distefano, M.; Sabatino, L.; San Bautista Primo, A.; Leonardi, C.; Giuffrida, F. Chlorophyll Fluorescence, Photosynthesis and Growth of Tomato Plants as Affected by Long-Term Oxygen Root Zone Deprivation and Grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S.; Sabatino, L. Agronomic Performance and Fruit Quality in Greenhouse Grown Eggplant Are Interactively Modulated by Iodine Dosage and Grafting. Sci. Hortic. 2022, 295, 110891. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Manna, D.; Sarkar, A.; Maity, T.K. Impact of biozyme on growth, yield and quality of chilli (Capsicum annuum L.). J. Crop Weed 2012, 8, 40–43. [Google Scholar]
- Abd El-Gawad, H.G.; Osman, H.S. Effect of exogenous application of boric acid and seaweed extract on growth, biochemical content and yield of eggplant. J. Hortic. Sci. Ornam. Plants 2014, 6, 133–143. [Google Scholar]
- Rolland, F.; Moore, B.; Sheen, J. Sugar sensing and signaling in plants. Plant Cell 2002, 14 (Suppl. 1), S185–S205. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Francioso, O.; Tinti, A.; Schiavon, M.; Pizzeghello, D.; Nardi, S. Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 2018, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Blunden, G.; Gordon, S.M. Betaines and their sulphono analogues in marine algae. Prog. Phycol. Res. 1986, 4, 39–80. [Google Scholar]
- Molla, A.H.; Haque, M.; Haque, A.; Ilias, G.N.M. Trichoderma-enriched biofertilizer enhances production and nutritional quality of tomato (Lycopersicon esculentum Mill.) and minimizes NPK fertilizer use. Agric. Res. 2012, 1, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Altomare, C.; Norvell, W.A.; Bjorkman, T.; Harman, G.E. Solubilization of phosphates and micronutrients by the plant-growth-promoting and biocontrol fungus Trichoderma harzianum Rifai 1295-22. Appl. Environ. Microbiol. 1999, 65, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E. Overview of new insights into mechanisms and uses of Trichoderma based products. Phytopathology 2004, 94, 138. [Google Scholar]
- Lee, J.M. Cultivation of grafted vegetables I: Current status, grafting methods and benefits. HortScience 1994, 29, 235–239. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E. Effect of salinity on yield, fruit quality, leaf gas exchange, and mineral composition of grafted watermelon plants. HortScience 2006, 41, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Farrell, A.; Ramsubhag, A.; Jayaraman, J. The effect of Ascophyllum nodosum extract on the growth, yield and fruit quality of tomato grown under tropical conditions. J. Appl. Psychol. 2016, 28, 1353–1362. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Hodges, D.M.; Zhang, J.; Kirby, C.W.; Ji, X.; Locke, S.J.; Critchley, A.T.; Prithiviraj, B. Commercial extract of brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis against oxidative and thermal stress. Food Chem. 2011, 124, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of Spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Darré, M.; Valerga, L.; Zaro, M.J.; Lemoine, M.L.; Concellón, A.; Vicente, A.R. Eggplant Grafting on a Cold-Tolerant Rootstock Reduces Fruit Chilling Susceptibility and Improves Antioxidant Stability during Storage. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, L.; Iapichino, G.; Maggio, A.; D’Anna, E.; Bruno, M.; D’Anna, F. Grafting affects yield and phenolic profile of Solanum melongena L. landraces. J. Int. agric. 2016, 15, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Hoyos Echevarria, P.; Morra, L.; Oda, A.M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Maršič, N.K.; Mikulič-Petkovšek, M.; Štampar, F. Grafting influences phenolic profile and carpometric traits of fruits of greenhouse-grown eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 10504–10514. [Google Scholar] [CrossRef] [PubMed]
- Oztekin, G.B.; Tuzel, Y.; Tuzel, I.H. Does mycorrhiza improve salinity tolerance in grafted plants? Sci. Hortic 2013, 149, 55–60. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef] [PubMed]
- Roussos, P.A.; Denaxa, N.K.; Damvakaris, T. Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci. Hortic. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Pohl, A.; Grabowska, A.; Kalisz, A.; Sękara, A. Biostimulant Application Enhances Fruit Setting in Eggplant—An Insight into the Biology of Flowering. Agronomy 2019, 9, 482. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awad, M.A.; de Jager, A.; van der Plas, L.H.W.; van der Krol, A.R. Flavonoid and chlorogenic acid changes in skin of ‘Elstar’ and ‘Jonagold’ apples during development and ripening. Sci. Hortic. 2001, 90, 69–83. [Google Scholar] [CrossRef]
- Mugnai, S.; Azzarello, E.; Pandolfi, C.; Salamagne, S.; Briand, X.; Mancuso, S. Enhancement of ammonium and potassium root influxes by the application of marine bioactive substances positively affects Vitis vinifera plant growth. J. Appl. Phycol. 2008, 20, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro-Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Riga, P. Effect of rootstock on growth, fruit production and quality of tomato plants grown under low temperature and light conditions. Hortic. Environ. Biotechnol. 2015, 56, 626–638. [Google Scholar] [CrossRef]
- Mystkowska, I. Reduction of glycoalkaloids in potato under the influence of biostimulators. Appl. Ecol. Environ. Res. 2019, 17, 3567–3574. [Google Scholar] [CrossRef]
- Papathanasiou, F.; Mitchell, S.H.; Watson, S.; Harvey, B.M.R. Effect of environmental stress during tuber development on accumulation of glycoalkaloids in potato (Solanum tuberosum L.). J. Sci. Food Agric. 1999, 79, 1183–1189. [Google Scholar] [CrossRef]
- Sabatino, L.; D’Anna, F.; Iapichino, G.; Moncada, A.; D’Anna, E.; De Pasquale, C. Interactive Effects of Genotype and Molybdenum Supply on Yield and Overall Fruit Quality of Tomato. Front. Plant Sci. 2019, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Stommel, J.R.; Whitaker, B.D. Phenolic Acid Content and Composition of Eggplant Fruit in a Germplasm Core Subset. J. Am. Soc. Hortic. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Mennella, G.; Scalzo, R.L.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; De Almeida, A.E.; Rotino, G.L. Chemical and Bioactive Quality Traits During Fruit Ripening in Eggplant (S. melongena L.) and Allied Species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef] [PubMed]
- Birner, J. A method for the determination of total steroid bases. J. Pharm. Sci. 1969, 58, 258–259. [Google Scholar] [CrossRef]
- Fogg, D.N.; Wilkinson, A.N. The colorimetric determination of phosphorus. Analyst 1958, 83, 406–414. [Google Scholar] [CrossRef]
- Subramanian, R.; Subbramaniyan, P.; Raj, V. Determination of some minerals and trace elements in two tropical medicinal plants. Asian Pac. J. Trop. Biomed. 2012, 2, S555–S558. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | FDM (%) | Firmness (N) | ClA (mg 100 g−1 dw) | TA (mg 100 g−1 dw) | Proteins (g 100 g−1 dw) | K (mg 100 g−1 dw) | Fe (µg g−1 dw) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T × B | ||||||||||||||
| Ungrafted × Control | 4.2 | e | −46.2 | a | 735.6 | d | 7531.8 | c | 12.0 | d | 338.0 | f | 24.80 | d |
| Ungrafted × An | 4.3 | e | −47.8 | b | 768.7 | c | 7607.8 | b | 13.9 | c | 353.0 | e | 25.37 | c |
| Ungrafted × Ta | 4.5 | d | −46.8 | a | 739.0 | d | 7536.9 | c | 12.1 | d | 348.9 | e | 24.57 | d |
| Ungrafted × An+Ta | 4.5 | d | −47.8 | b | 769.3 | c | 7606.6 | b | 13.9 | c | 360.9 | d | 25.40 | c |
| Self-grafted × Control | 4.3 | e | −46.1 | a | 733.6 | d | 7614.8 | b | 11.9 | d | 346.2 | e | 24.87 | d |
| Self-grafted × An | 4.4 | d | −48.0 | b | 768.8 | c | 7647.7 | a | 14.0 | c | 360.8 | d | 25.50 | c |
| Self-grafted × Ta | 4.4 | d | −46.1 | a | 736.3 | d | 7615.7 | b | 11.9 | d | 358.9 | d | 24.57 | d |
| Self-grafted × An+Ta | 4.5 | d | −48.2 | b | 769.8 | c | 7648.1 | a | 13.8 | c | 369.3 | cd | 25.67 | c |
| Grafted × Control | 5.3 | c | −46.2 | a | 895.3 | b | 7228.5 | e | 15.4 | b | 363.6 | d | 31.80 | b |
| Grafted × An | 5.5 | b | −48.5 | b | 987.5 | a | 7284.7 | d | 16.8 | a | 373.2 | c | 33.20 | a |
| Grafted × Ta | 5.8 | a | −46.2 | a | 892.4 | b | 7228.4 | e | 15.4 | b | 380.3 | b | 31.50 | b |
| Grafted × An+Ta | 5.8 | a | −49.7 | c | 995.7 | a | 7289.9 | d | 16.6 | a | 391.2 | a | 33.23 | a |
| Significance | ||||||||||||||
| T | *** | *** | *** | *** | *** | *** | *** | |||||||
| B | *** | *** | *** | *** | *** | *** | *** | |||||||
| T × B | ** | *** | *** | *** | *** | ** | ** | |||||||
| Treatments | SSC (°Brix) | AA (mg 100 g−1 fw) | GLY (mg 100 g−1 dw) | P (mg 100 g−1 dw) | Ca (mg 100 g−1 dw) | Mg (mg 100 g−1 dw) | Cu (µg g−1 dw) | Zn (µg g−1 dw) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T | ||||||||||||||||
| Ungrafted | 4.3 | a | 6.55 | b | 76.67 | a | 513.40 | 104.04 | 17.68 | a | 2.68 | b | 9.61 | b | ||
| Self-grafted | 4.0 | b | 6.53 | b | 77.09 | a | 540.08 | 104.14 | 17.68 | a | 2.72 | b | 9.55 | b | ||
| Grafted | 3.9 | c | 7.13 | a | 46.64 | b | 513.98 | 104.41 | 15.75 | b | 3.57 | a | 10.57 | a | ||
| B | ||||||||||||||||
| Control | 4.1 | 6.58 | b | 71.73 | a | 502.12 | 104.42 | 16.27 | d | 2.82 | b | 9.68 | b | |||
| An | 4.1 | 6.90 | a | 61.84 | b | 526.36 | 104.20 | 17.17 | b | 3.17 | a | 10.14 | a | |||
| Ta | 4.1 | 6.52 | b | 72.06 | a | 501.32 | 103.93 | 16.86 | c | 2.81 | b | 9.68 | b | |||
| An+Ta | 4.0 | 6.94 | a | 61.57 | b | 560.14 | 104.23 | 17.86 | a | 3.14 | a | 10.13 | a | |||
| Significance | ||||||||||||||||
| T | *** | *** | *** | NS | NS | *** | *** | *** | ||||||||
| B | NS | *** | *** | NS | NS | *** | *** | *** | ||||||||
| T × B | NS | NS | NS | NS | NS | NS | NS | NS | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabatino, L.; Consentino, B.B.; Ntatsi, G.; La Bella, S.; Baldassano, S.; Rouphael, Y. Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant. Plants 2022, 11, 1175. https://doi.org/10.3390/plants11091175
Sabatino L, Consentino BB, Ntatsi G, La Bella S, Baldassano S, Rouphael Y. Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant. Plants. 2022; 11(9):1175. https://doi.org/10.3390/plants11091175
Chicago/Turabian StyleSabatino, Leo, Beppe Benedetto Consentino, Georgia Ntatsi, Salvatore La Bella, Sara Baldassano, and Youssef Rouphael. 2022. "Stand-Alone or Combinatorial Effects of Grafting and Microbial and Non-Microbial Derived Compounds on Vigour, Yield and Nutritive and Functional Quality of Greenhouse Eggplant" Plants 11, no. 9: 1175. https://doi.org/10.3390/plants11091175