
Physiological and Growth Responses of Potato (Solanum Tuberosum L.) to Air Temperature and Relative Humidity under Soil Water Deficits

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
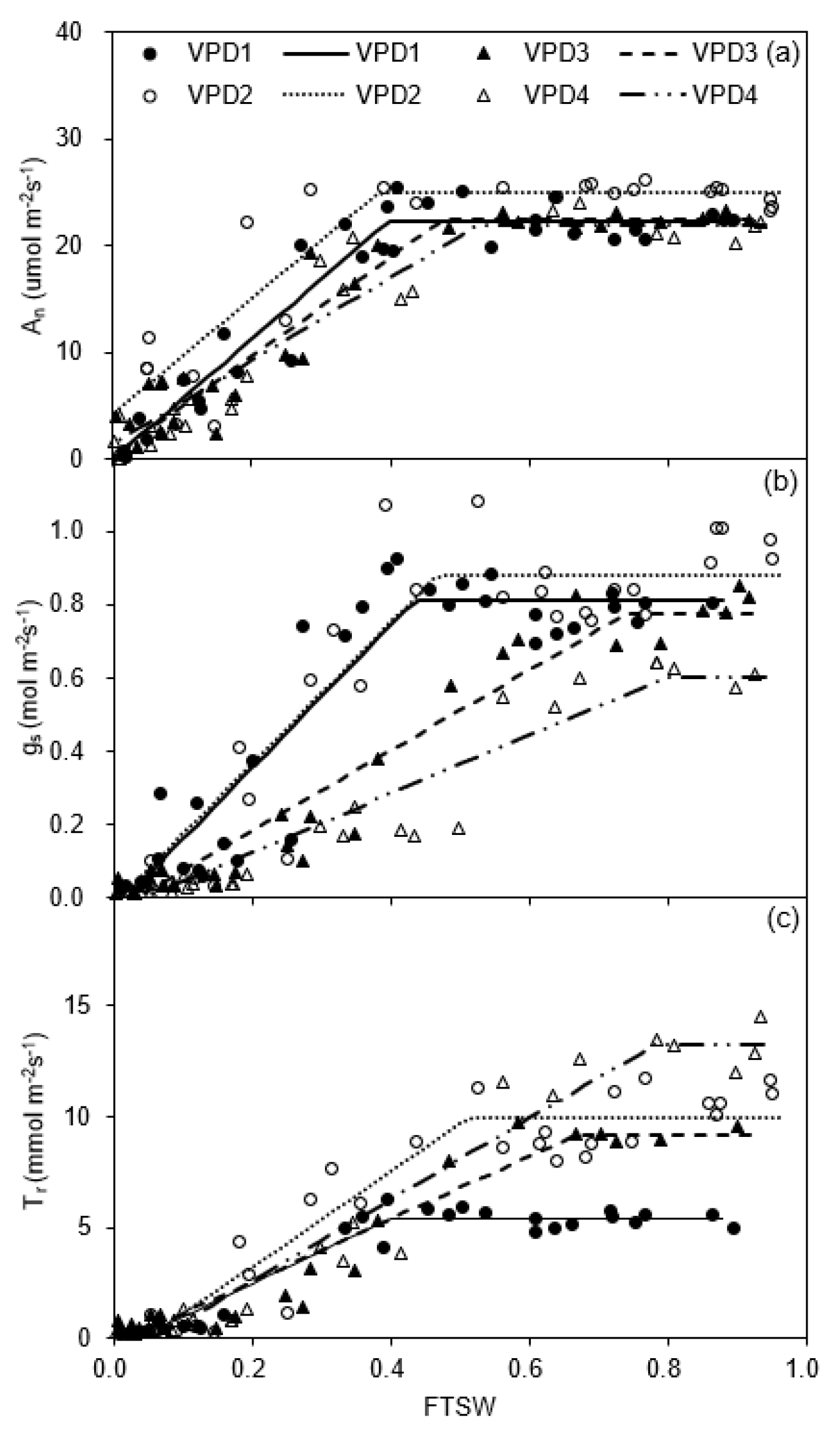
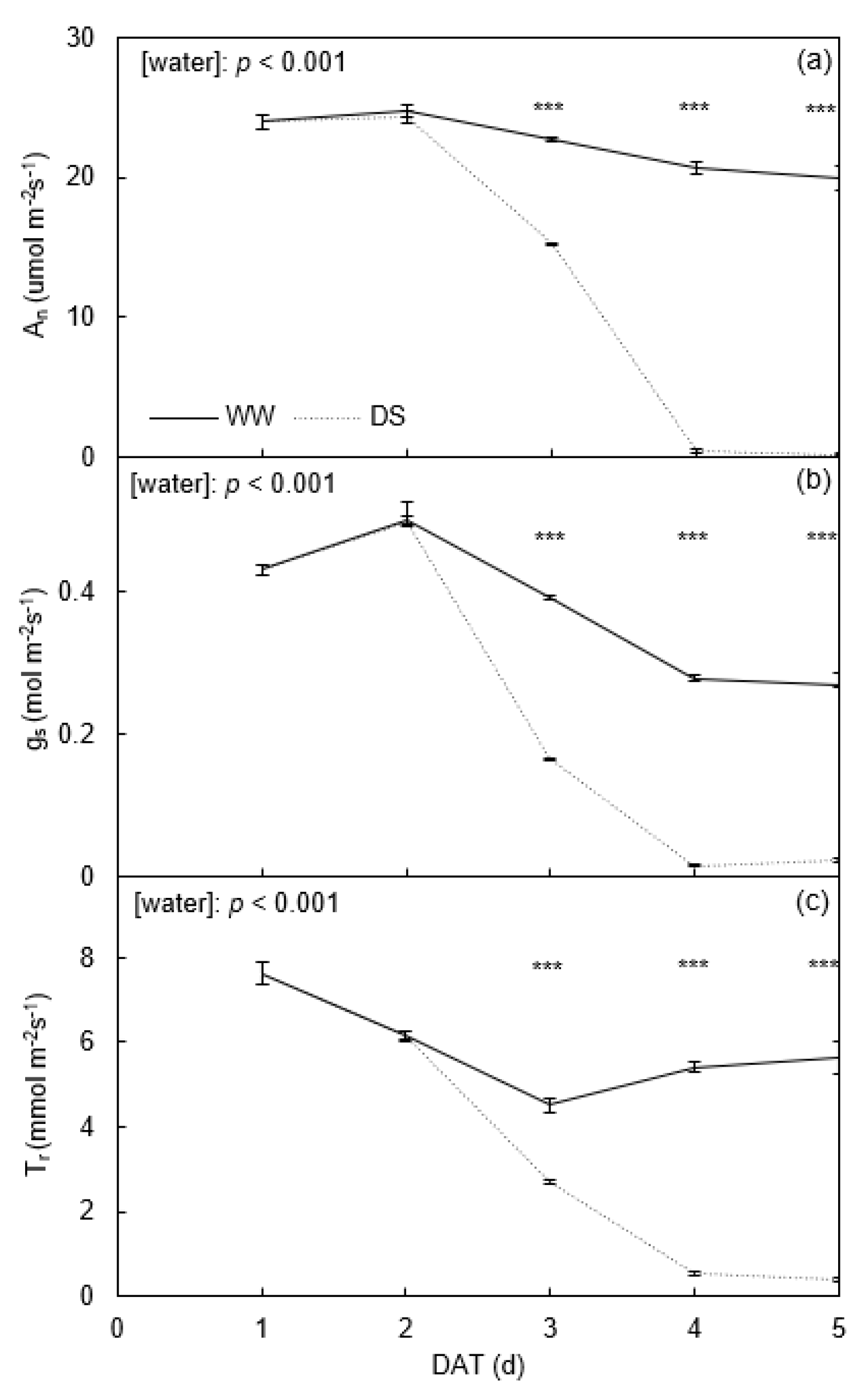
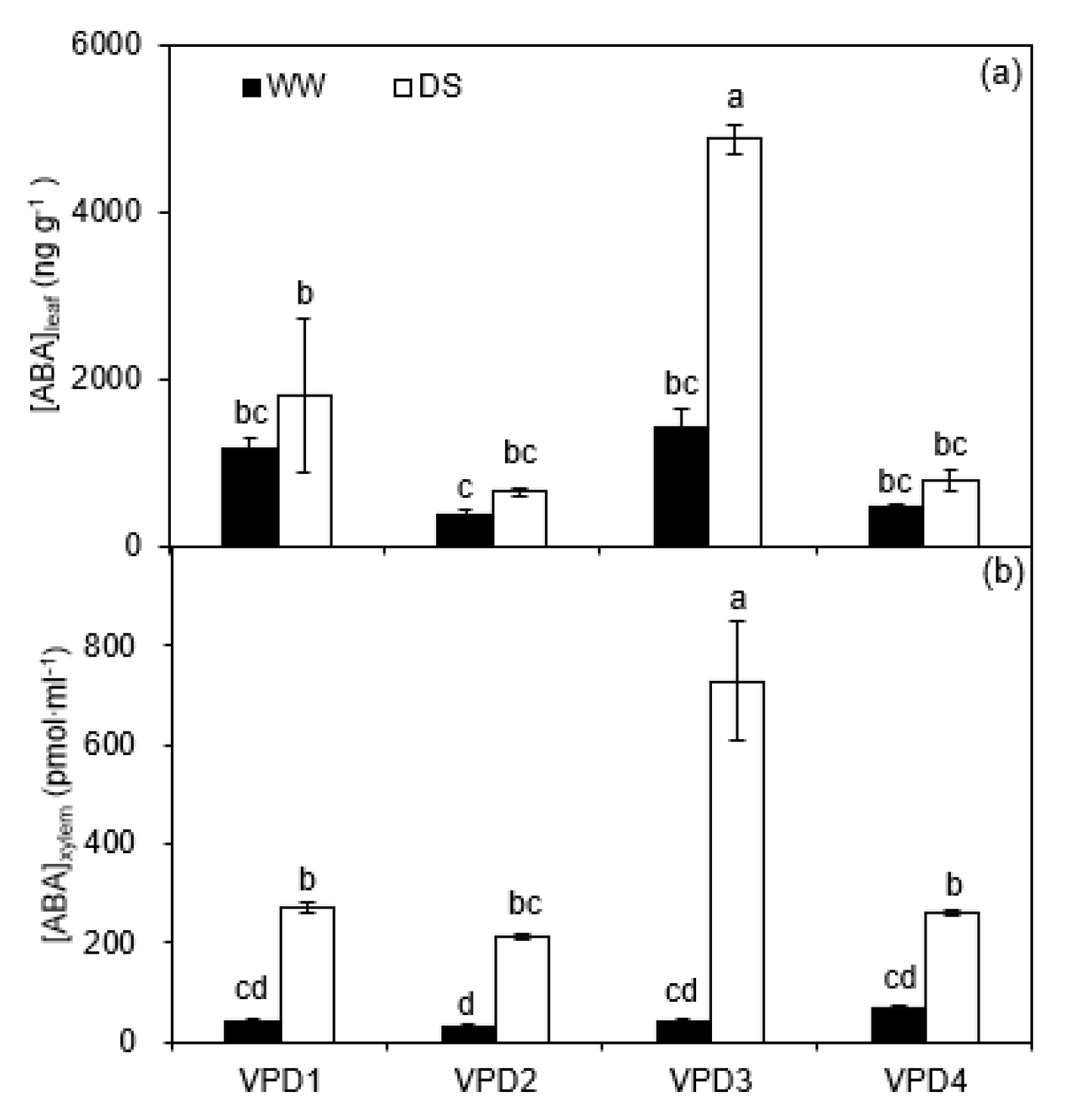
2.1. Leaf Gas Exchange and Chemical Signals
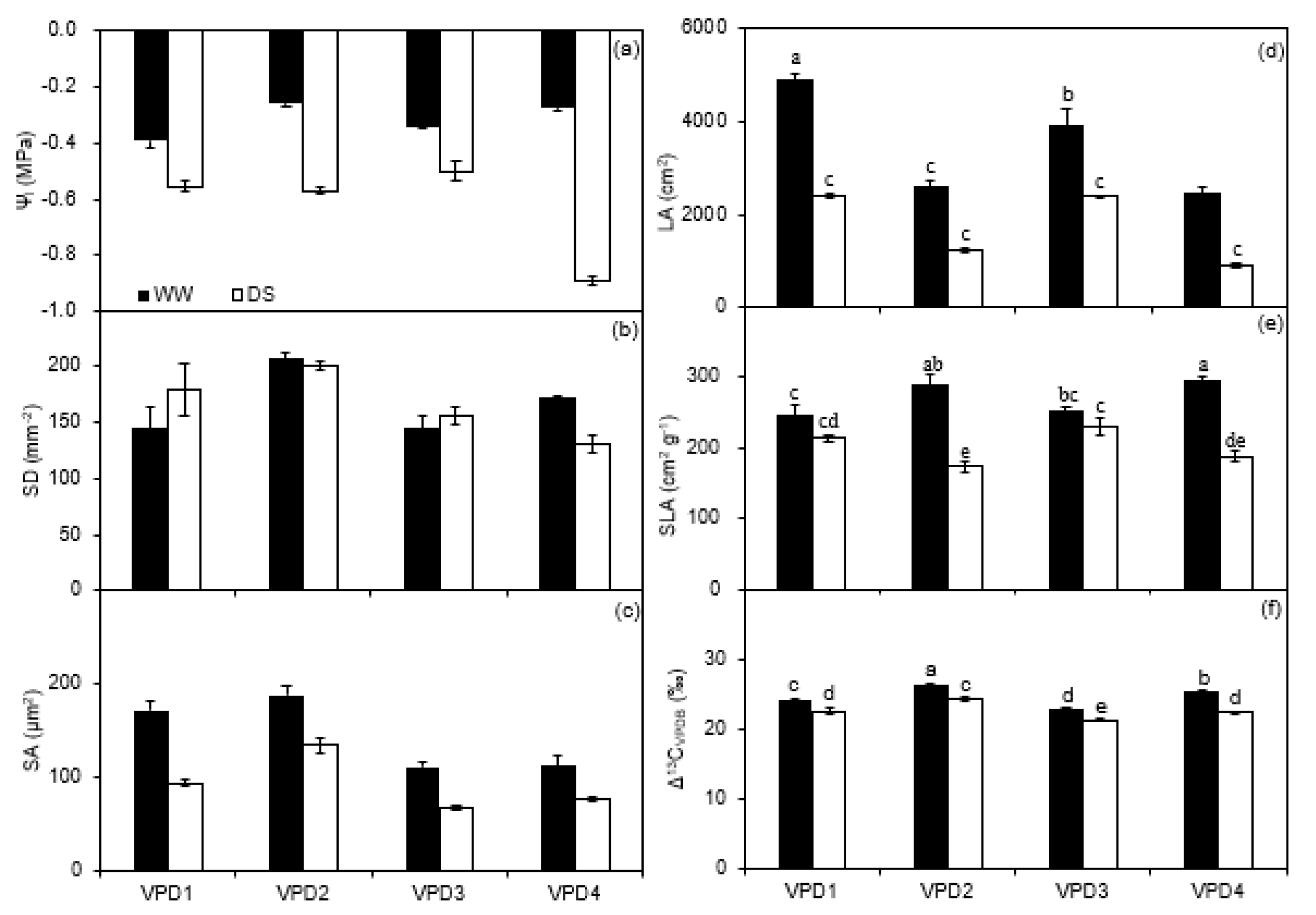
2.2. Leaf Water Potential, Stomatal Morphology, Leaf Area, and Specific Leaf Area
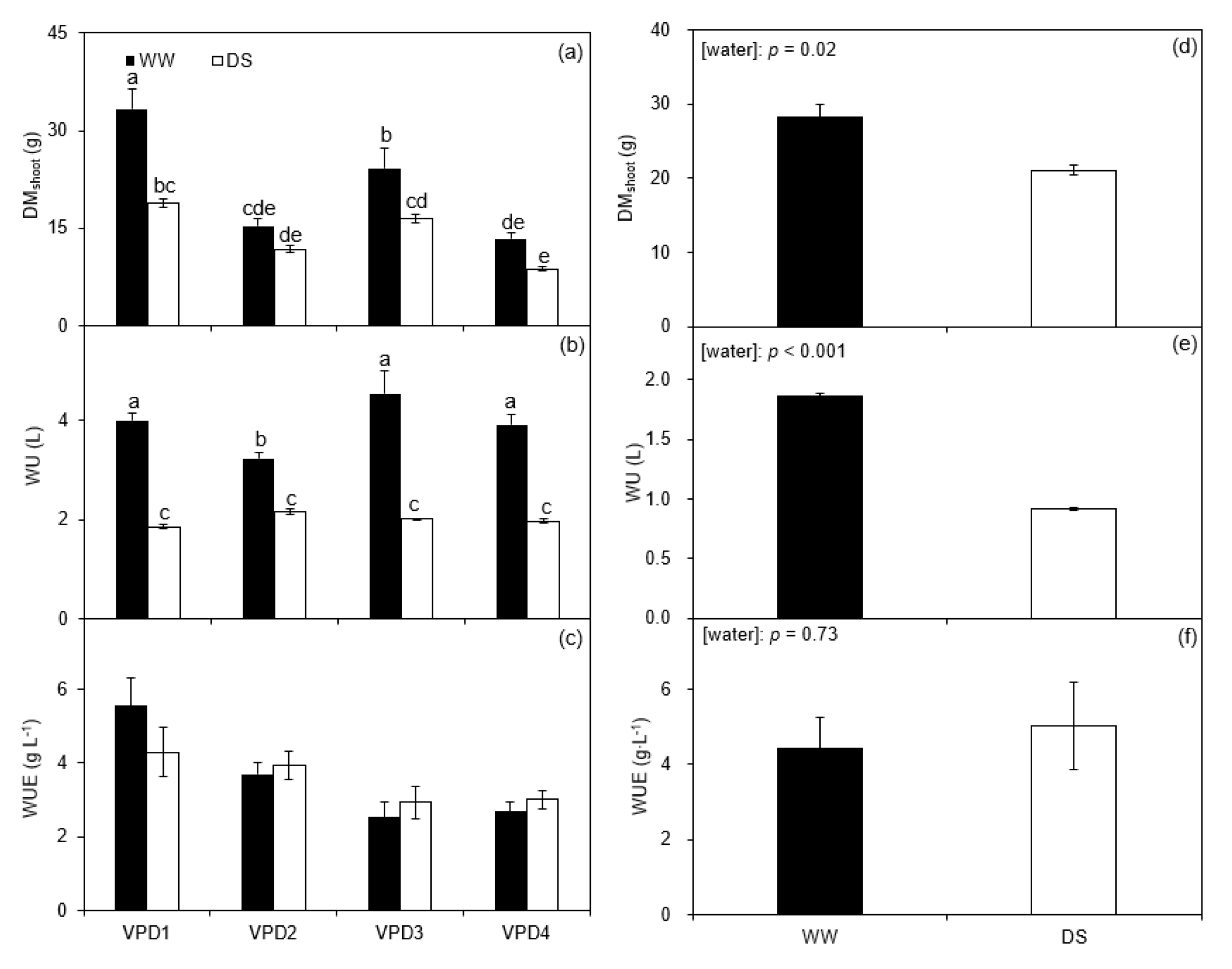
2.3. Shoot Dry Matter, Water Use, and Water Use Efficiency
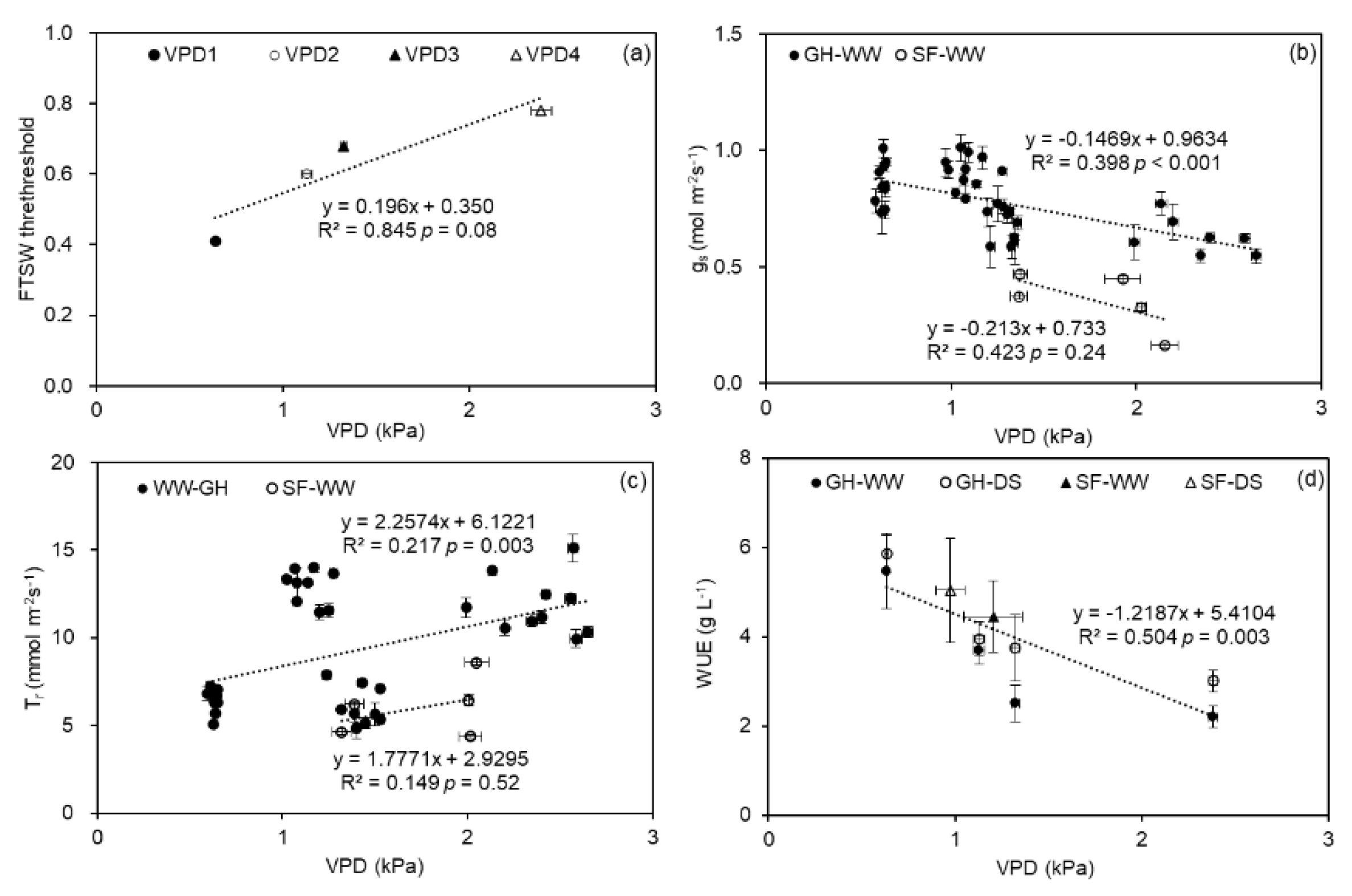
2.4. Relationships between FTSW Threshold, gs, Tr, WUE, and VPD
3. Discussion
3.1. Leaf Gas Exchange and Chemical Signaling
3.2. Leaf Water Relations, Stomatal Morphology, Leaf Area, and Specific Leaf Area
3.3. Shoot Dry Matter, Water Use, and Water Use Efficiency
3.4. Relationships between FTSW Threshold, gs, Tr, WUE, and VPD
4. Materials and Methods
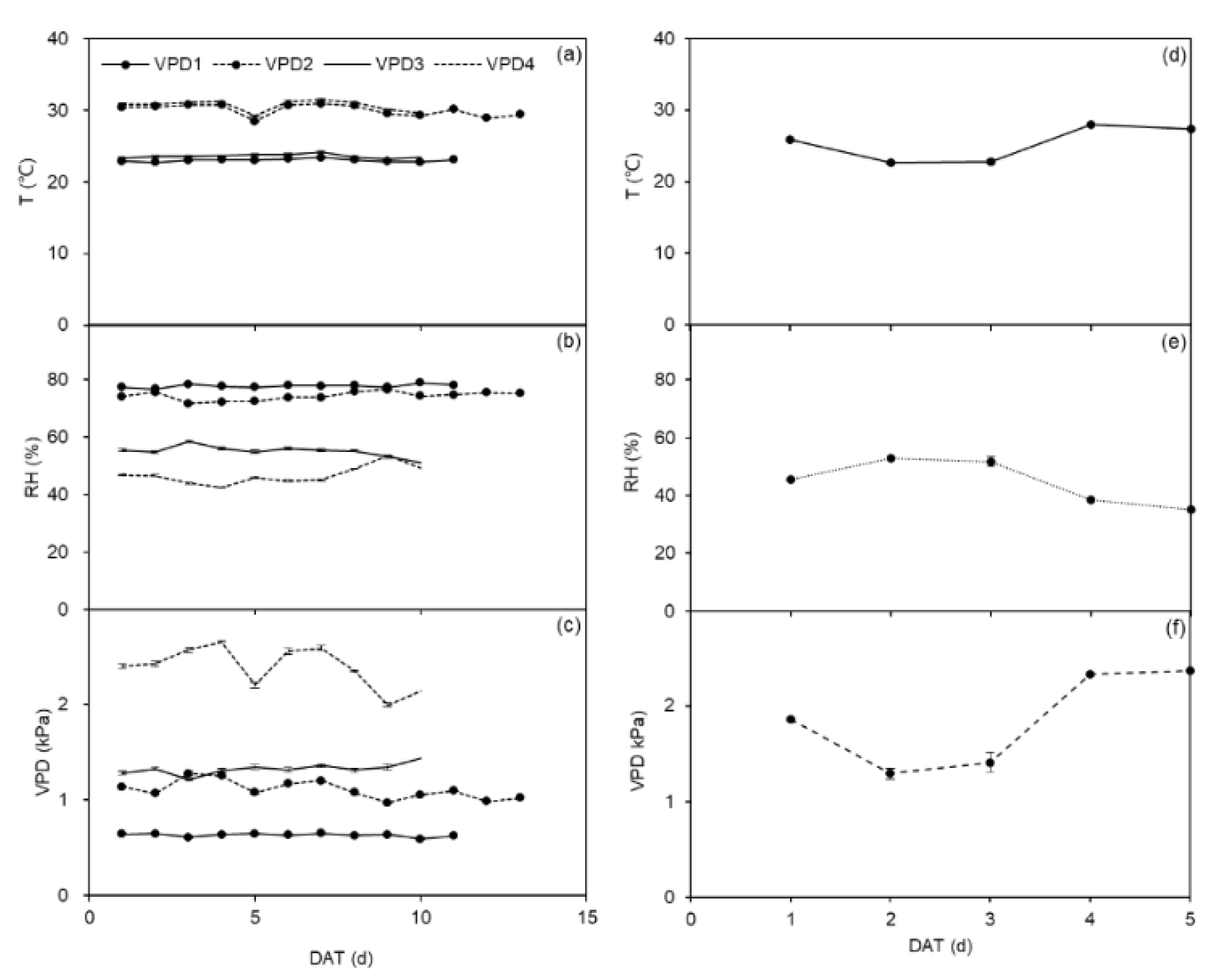
4.1. Greenhouse and Field Experiments
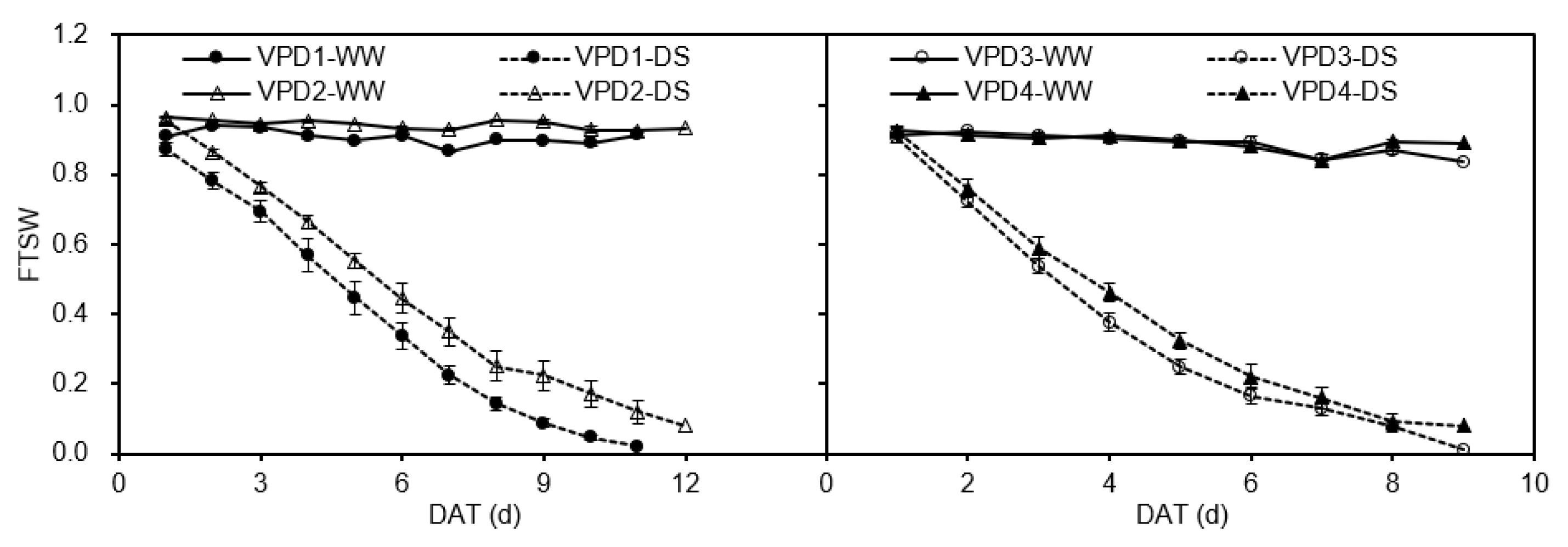
4.2. Irrigation Treatments
4.3. Variables Evalated
4.4. Leaf Gas Exchange and Chemical Signals
4.5. Leaf Water Potential, Stomatal Morphology, Leaf Area, and Specific Leaf Area
4.6. Shoot Dry Matter, Water Use, and Water Use Efficiency
4.7. Data Analyses and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trenberth, K.E.; Dai, A.; Van, D.; Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Wang, D.B.; Hejazi, M.I.; Cai, X.M.; Valocchi, A.J. Climate change impact on meteorological, hydrological, and agricultural drought: A case study of central illinois. Water Resour. Res. 2011, 47, W09527. [Google Scholar] [CrossRef] [Green Version]
- Strzepek, K.; Yohe, G.; Neumann, J.; Boehlert, B. Characterizing changes in drought risk for the United States from climate change. Environ. Res. Lett. 2010, 5, 44012–44019. [Google Scholar] [CrossRef]
- Patanè, C.; Cosentino, S.L. Effects of soil water deficit on yield and quality of processing tomato under a Mediterranean climate. Agric. Water Manag. 2010, 97, 131–138. [Google Scholar] [CrossRef]
- Liu, F.L.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.E. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- López, J.; Way, A.D.; Sadok, W. Systemic effects of rising atmospheric vapor pressure deficit on plant physiology and productivity. Glob. Change Biol. 2021, 27, 1704–1720. [Google Scholar] [CrossRef]
- Yang, Z.; Sinclair, T.R.; Zhu, M.; Messina, C.D.; Cooper, M.; Hammer, G.L. Temperature effect on transpiration response of maize plants to vapour pressure deficit. Environ. Exp. Bot. 2012, 78, 157–162. [Google Scholar] [CrossRef]
- Kaminski, K.P.; Kørup, K.; Nielsen, K.L.; Liu, F.L.; Topbjerg, H.B.; Kirk, H.G.; Andersen, M.N. Gas-exchange, water use efficiency and yield responses of elite potato (Solanum tuberosum L.) cultivars to changes in atmospheric carbon dioxide concentration, temperature and relative humidity. Agric. For. Meteorol. 2014, 187, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Buckley, T.N. The control of stomata by water balance. New Phytol. 2005, 168, 275–292. [Google Scholar] [CrossRef]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA transport and plant water stress responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef]
- Liu, F.L.; Shahnazari, A.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J. Exp. Bot. 2006, 57, 3727–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, W.J.; Zhang, J. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. Optimizing nitrogen fertilization to improve qualitative performances and physiological and yield responses of potato (Solanum Tuberosum L.). Agronomy 2020, 10, 352. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, S.; Abbate, C.; Pandino, G.; Parisi, B.; Mauromicale, G. Productive and physiological response of organic potato grown under highly calcareous soils to fertilization and mycorrhization management. Agronomy 2020, 10, 1200. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The mineral profile in organically and conventionally grown “early” crop potato tubers. Sci. Hortic. 2014, 167, 169–173. [Google Scholar] [CrossRef]
- Rykaczewska, K. The impact of high temperature during growing season on potato cultivars with different response to environmental stresses. Am. J. Plant Sci. 2013, 4, 2386–2393. [Google Scholar] [CrossRef] [Green Version]
- Elferjani, R.; Soolanayakanahally, R. Canola responses to drought, heat, and combined stress: Shared and specific effects on carbon assimilation, seed yield, and oil composition. Front. Plant Sci. 2018, 9, 01224. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Wu, L.F.; Fiona, F.; Yang, X.X.; Fu, D.H. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant. 2017, 39, 125. [Google Scholar] [CrossRef]
- Rykaczewska, K. Impact of heat and drought stresses on size and quality of the potato yield. Plant Soil Environ. 2017, 63, 40–46. [Google Scholar] [CrossRef] [Green Version]
- McAdam, S.A.; Brodribb, T.J. The evolution of mechanisms driving the stomatal response to vapor pressure deficit. Plant Physiol. 2015, 167, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Wang, Y.; Williamson, L.; Holroyd, G.H.; Tagliavia, C.; Murchie, E.; Theobald, J.; Knight, M.R.; Davies, W.J.; Leyser, H.O.; et al. The identification of genes involved in the stomatal response to reduced atmospheric relative humidity. Curr. Biol. 2006, 16, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Nukaya, T.; Kamimura, T.; Zhang, D.; Kurimoto, I.; Takagaki, M.; Maruo, T.; Kozai, T.; Yamori, W. Control of vapor pressure deficit (VPD) in greenhouse enhanced tomato growth and productivity during the winter season. Sci. Hortic. 2015, 197, 17–23. [Google Scholar] [CrossRef]
- Grange, R.I.; Hand, D.W. A review of the effects of atmospheric humidity on the growth of horticultural crops. J. Pomol. Hortic. Sci. 1987, 62, 125–134. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hammer, G.L.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Chang. 2013, 3, 497–501. [Google Scholar] [CrossRef]
- Nejad, A.R.; Meeteren, U. Stomatal response characteristics of tradescantia virginiana grown at high relative air humidity. Physiol. Plant. 2003, 125, 324–332. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, X.; Chen, Y.T.; Wei, Z.H.; Liu, F.L. Dissecting the combined effects of air temperature and relative humidity on water-use efficiency of barley under drought stress. J. Agron. Crop Sci. 2021, 207, 606–617. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.E.; Power, S.A.; Duursma, R.A.; Medlyn, B.E.; Aspinwall, M.J.; Choat, B.; Creek, D.; Eamus, D.; Maier, C.; Pfautsch, S. Stomatal and non-stomatal limitations of photosynthesis for four tree species under drought: A comparison of model formulations. Agric. For. Meteorol. 2017, 247, 454–466. [Google Scholar] [CrossRef]
- Shirke, P.A.; Pathre, U.V. Influence of leaf-to-air vapour pressure deficit (VPD) on the biochemistry and physiology of photosynthesis in Prosopis juliflora. J. Exp. Bot. 2004, 55, 2111–2120. [Google Scholar] [CrossRef]
- Prange, R.K.; Mcrae, K.B.; Midmore, D.J.; Deng, R. Reduction in potato growth at high temperature: Role of photosynthesis and dark respiration. Am. Potato J. 1990, 67, 357–369. [Google Scholar] [CrossRef]
- Wolf, S.; Marani, A.; Rudich, J. Effects of temperature and photoperiod on assimilate partitioning in potato plants. Ann. Bot. 1990, 66, 513–520. [Google Scholar] [CrossRef]
- Liu, F.L.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ. Exp. Bot. 2005, 54, 33–40. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signaling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Merilo, E.; Yarmolinsky, D.; Jalakas, P.; Parik, H.; Tulva, I.; Rasulov, B.; Kilk, K.; Kollist, H. Stomatal VPD response: There is more to the story than ABA. Plant Physiol. 2018, 176, 851–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamshiri, R.R.; Jones, J.W.; Thorp, K.R.; Ahmad, D.; Che, M.H.; Taheri, S. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: A review. Int. Agrophysics 2018, 32, 287–302. [Google Scholar] [CrossRef]
- Denmead, O.T.; Shaw, R.H. Availability of soil water to plants as affected by soil moisture content and meteorological conditions. Agron. J. 1962, 54, 385–390. [Google Scholar] [CrossRef]
- Fanourakis, D.; Bouranis, D.; Giday, H.; Carvalho, D.R.; Nejad, A.R.; Ottosen, C.O. Improving stomatal functioning at elevated growth air humidity: A review. J. Plant Physiol. 2016, 207, 51–60. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Kurepin, L.V.; Reid, D.M. Effects of temperature and watering regime on growth, gas exchange and abscisic acid content of canola (Brassica napus) seedlings. Environ. Exp. Bot. 2012, 75, 107–113. [Google Scholar] [CrossRef]
- Arve, L.E.; Terfa, M.T.; Gislerød, H.R.; Olsen, J.E.; Torre, S. High relative air humidity and continuous light reduce stomata functionality by affecting the ABA regulation in rose leaves. Plant Cell Environ. 2013, 36, 382–392. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Hinckley, T.M.; Ceulemans, R. Apparent responses of stomata to transpiration and humidity in a hybrid poplar canopy. Plant Cell Environ. 1997, 20, 1301–1308. [Google Scholar] [CrossRef]
- Montero, J.I.; Antón, A.; Munoz, P.; Lorenzo, P. Transpiration from geranium grown under high temperatures and low humilities in greenhouses. Agric. For. Meteorol. 2001, 107, 323–332. [Google Scholar] [CrossRef]
- Crawford, A.J.; McLachlan, D.H.; Hetherington, A.M.; Franklin, K.A. High temperature exposure increases plant cooling capacity. Curr. Biol. 2012, 22, 396–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143. [Google Scholar] [CrossRef]
- Brilli, F.; Barta, C.; Fortunati, A.; Lerdau, M.; Loreto, F.; Centritto, M. Response of isoprene emission and carbon metabolism to drought in white poplar (Populus alba) saplings. New Phytol. 2007, 175, 244–254. [Google Scholar] [CrossRef]
- Gholipoor, M.; Sinclair, T.R.; Raza, M.A.; Löffler, C.; Cooper, M.; Messina, C.D. Maize hybrid variability for transpiration decreases with progressive soil drying. J. Agron. Crop Sci. 2012, 199, 23–29. [Google Scholar] [CrossRef]
- Devi, M.J.; Taliercio, E.W.; Sinclair, T.R. Leaf expansion of soybean subjected to high and low atmospheric vapour pressure deficits. J. Exp. Bot. 2015, 66, 1845–1850. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R.; Ludlow, M.M. Influence of soil water supply on the plant water balance of four tropical grain legumes. Funct. Plant Biol. 1986, 13, 329–341. [Google Scholar] [CrossRef]
- Tsukaguchi, T.; Kawamitsu, Y.; Takeda, H.; Suzuki, K.; Egawa, Y. Water status of flower buds and leaves as affected by high temperature in heat-tolerant and heat-sensitive cultivars of snap bean (Phaseolus vulgaris L.). Plant Prod. Sci. 2003, 6, 24–27. [Google Scholar] [CrossRef]
- Marino, G.; Brunetti, C.; Tattini, M.; Romano, A.; Biasioli, F.; Tognetti, R.; Loreto, F.; Ferrini, F.; Centritto, M. Dissecting the role of isoprene and stress-related hormones (ABA and ethylene) in Populus nigra exposed to unequal root zone water stress. Tree Physiol. 2017, 37, 1637–1647. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.; Guerin, G.; Hill, B.; Watling, J. Temperature influences stomatal density and maximum potential water loss through stomata of dodonaea viscosa subsp. angustissima along a latitude gradient in southern Australia. Aust. J. Bot. 2015, 62, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Jumrani, K.; Bhatia, V.S.; Pandey, G.P. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean. Photosynth. Res. 2017, 131, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, D.R.; Bond, B.J.; Meinzer, F.C. Does turgor limit growth in tall trees? Plant Cell Environ. 2004, 27, 229–236. [Google Scholar] [CrossRef]
- Feller, U.; Anders, I.; Mae, T. Rubiscolytics: Fate of Rubisco after its enzymatic function in a cell is terminated. J. Exp. Bot. 2007, 59, 1615–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carins Murphy, M.R.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Stuerz, S.; Asch, F. Responses of rice growth to day and night temperature and relative air humidity—Dry matter, leaf area, and partitioning. Plants 2019, 8, 521. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.L.; Stützel, H. Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci. Hortic. 2004, 102, 15–27. [Google Scholar] [CrossRef]
- Stewart, J.J.; Demmig-Adams, B.; Cohu, C.M.; Wenzl, C.A.; Muller, O.; Adams, W.W. Growth temperature impact on leaf form and function in arabidopsis thaliana ecotypes from northern and southern Europe. Plant Cell Environ. 2016, 39, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Beerling, D.J.; Woodward, F.I. Leaf stable carbon isotope composition records increased water-use efficiency of C3 plants in response to atmospheric CO2 enrichment. Funct. Ecol. 1995, 9, 394–401. [Google Scholar] [CrossRef]
- Liu, J.; Hu, T.; Fang, L.; Peng, X.; Liu, F.L. CO2 elevation modulates the response of leaf gas exchange to progressive soil drying in tomato plants. Agric. For. Meteorol. 2019, 268, 181–188. [Google Scholar] [CrossRef]
- Rashid, M.A.; Andersen, M.N.; Wollenweber, B.; Zhang, X.; Olesen, J.E. Acclimation to higher VPD and temperature minimized negative effects on assimilation and grain yield of wheat. Agric. For. Meteorol. 2018, 248, 119–129. [Google Scholar] [CrossRef]
- Cunningham, S.C. Stomatal sensitivity to vapour pressure deficit of temperate and tropical evergreen rainforest trees of Australia. Trees 2004, 18, 399–407. [Google Scholar] [CrossRef]
- Ray, J.D.; Sinclair, T.R. The effect of pot size on growth and transpiration of maize and soybean during water deficit stress. J. Exp. Bot. 1998, 49, 1381–1386. [Google Scholar] [CrossRef]
- Ray, J.D.; Gesch, R.W.; Sinclair, T.R.; Allen, L.H. The effect of vapor pressure deficit on maize transpiration response to a drying soil. Plant Soil 2002, 239, 113–121. [Google Scholar] [CrossRef]
- Asch, F. Determination of Abscisic Acid by Indirect Enzyme Linked Immuno Sorbent Assay (ELISA); Technical Report; Laboratory for Agro-hydrology and Bioclimatology, Department of Agricultural Sciences, The Royal Veterinary and Agricultural University: Taastrup, Denmark, 2000; pp. 12–56. [Google Scholar]
- Su, Y.; Li, W.; Huang, Z.; Wang, R.; Luo, W.; Liu, Q.; Tong, J.; Xiao, L. Sensitive and high throughput quantification of abscisic acid based on quantitative real time immuno-PCR. Plant Methods 2018, 14, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, S.E.; Liu, F.L.; Jensen, C.R. Does root-sourced aba play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa willd.). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | An An max (μmol m−2 s−1) | CA | gs gs max (mol m−2 s−1) | Cg | Tr Tr max (mmol m−2 s−1) | CT |
|---|---|---|---|---|---|---|
| VPD1 | 22.23 (21.04–23.42) | 0.40 (0.35–0.44) | 0.81 (0.75–0.87) | 0.43 (0.37–0.50) | 4.96 (4.57–5.35) | 0.41 (0.35–0.47) |
| VPD2 | 24.94 (23.21–26.66) | 0.38 (0.28–0.49) | 0.88 (0.81–0.95) | 0.47 (0.38–0.55) | 10.69 (9.93–11.46) | 0.60 (0.46–0.75) |
| VPD3 | 22.38 (20.78–23.98) | 0.47 (0.39–0.56) | 0.77 (0.70–0.85) | 0.74 (0.64–0.84) | 9.35 (8.63–10.06) | 0.68 (0.60–0.77) |
| VPD4 | 21.91 (19.97–23.86) | 0.53 (0.43–0.62) | 0.60 (0.52–0.69) | 0.80 (0.66–0.94) | 14.99 (13.16–16.81) | 0.78 (0.64–0.91) |
| Factor | [ABA]leaf (ng g−1) | [ABA]xylem (pmol ml−1) | Ψl (MPa) | SD (mm−2) | SA (μm2) | LA (cm2) | SLA (cm2 g−1) | Δ13C (‰) | DMshoot (g) | WU (L) | WUE (g L−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| [T] | *** | ** | * | ** | * | *** | ns | *** | *** | ns | * |
| [RH] | ** | ** | * | ns | *** | * | ns | *** | * | ns | *** |
| [Irr] | ** | *** | *** | ns | *** | *** | *** | *** | *** | *** | ns |
| [T*RH] | * | * | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| [T*Irr] | * | ** | ns | ns | ns | ns | *** | * | * | ** | ns |
| [RH*Irr] | * | ** | ns | ns | * | ns | ns | ns | ns | ns | ns |
| [T*RH*Irr] | * | ** | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Greenhouse Cells | VPD Treatments | T (°C) | RH (%) | VPD (kPa) | CO2 (ppm) |
|---|---|---|---|---|---|
| I | VPD1 | 23 | 75 | 0.70 | 400 |
| II | VPD2 | 30 | 75 | 1.06 | 400 |
| III | VPD3 | 23 | 50 | 1.40 | 400 |
| IV | VPD4 | 30 | 50 | 2.12 | 400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Yang, X.; Manevski, K.; Li, S.; Wei, Z.; Andersen, M.N.; Liu, F. Physiological and Growth Responses of Potato (Solanum Tuberosum L.) to Air Temperature and Relative Humidity under Soil Water Deficits. Plants 2022, 11, 1126. https://doi.org/10.3390/plants11091126
Zhang P, Yang X, Manevski K, Li S, Wei Z, Andersen MN, Liu F. Physiological and Growth Responses of Potato (Solanum Tuberosum L.) to Air Temperature and Relative Humidity under Soil Water Deficits. Plants. 2022; 11(9):1126. https://doi.org/10.3390/plants11091126
Chicago/Turabian StyleZhang, Peng, Xin Yang, Kiril Manevski, Shenglan Li, Zhenhua Wei, Mathias Neumann Andersen, and Fulai Liu. 2022. "Physiological and Growth Responses of Potato (Solanum Tuberosum L.) to Air Temperature and Relative Humidity under Soil Water Deficits" Plants 11, no. 9: 1126. https://doi.org/10.3390/plants11091126