
Phenolic Compounds Content Evaluation of Lettuce Grown under Short-Term Preharvest Daytime or Nighttime Supplemental LEDs


 , ,
, ,  , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Results
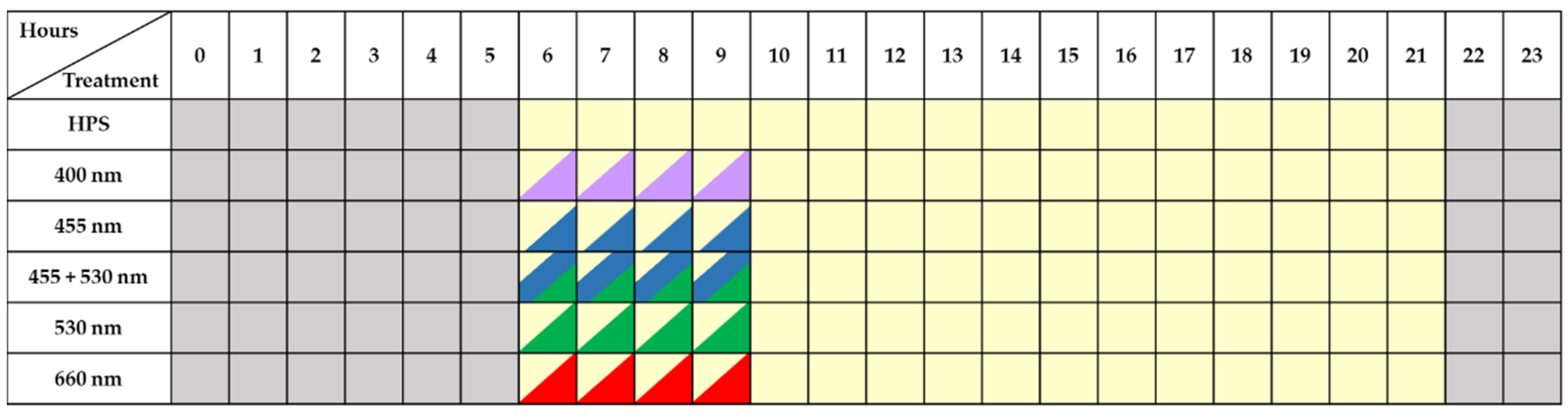
2.1. Effect of Short-Term Daytime Supplemental LEDs to HPS Lighting on Phenolic Compounds Content in Lettuce Cultivated in A Greenhouse during Different Seasons
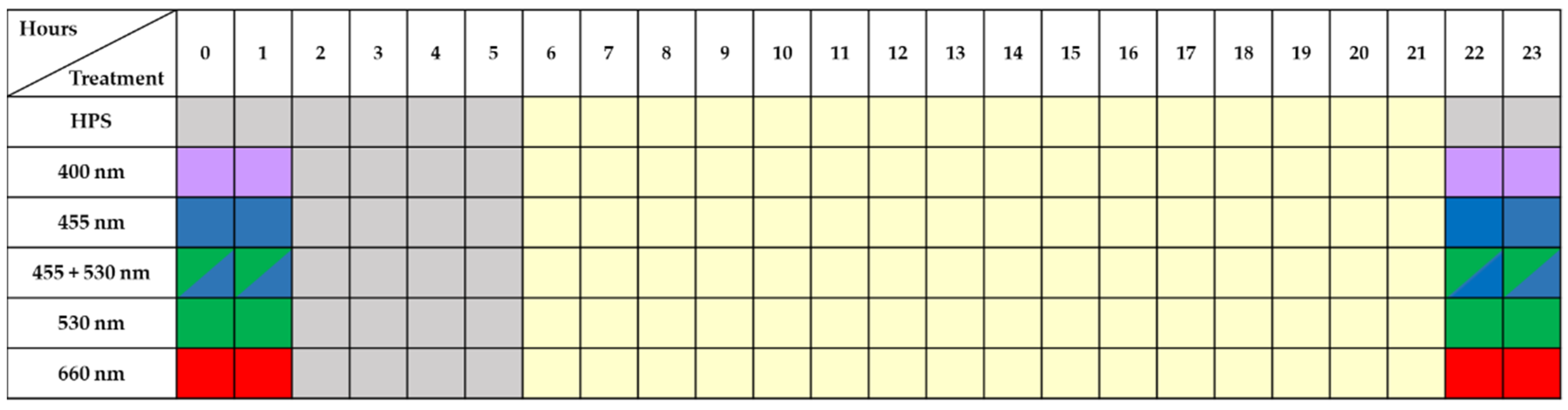
2.2. Effect of Short-Term Nighttime Supplemental LEDs to HPS Lighting on Phenolic Compounds Content in Lettuce Cultivated in A Greenhouse during Different Seasons
3. Discussion
4. Materials and Methods
4.1. Growth Conditions
4.2. Lighting Treatments
4.3. Determination of Individual Phenolic Compounds
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving Vegetable Quality in Controlled Environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of Light on Secondary Metabolites in Selected Leafy Greens: A Review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [CrossRef] [PubMed]
- Appolloni, E.; Pennisi, G.; Zauli, I.; Carotti, L.; Paucek, I.; Quaini, S.; Orsini, F.; Gianquinto, G. Beyond Vegetables: Effects of Indoor LED Light on Specialized Metabolite Biosynthesis in Medicinal and Aromatic Plants, Edible Flowers, and Microgreens. J. Sci. Food Agric. 2022, 102, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Wong, C.E.; Teo, Z.W.N.; Shen, L.; Yu, H. Seeing the Lights for Leafy Greens in Indoor Vertical Farming. Trends Food Sci. Technol. 2020, 106, 48–63. [Google Scholar] [CrossRef]
- Viršilė, A.; Brazaitytė, A.; Jankauskienė, J.; Miliauskienė, J.; Vaštakaitė, V.; Odminytė, I.; Novičkovas, A.; Samuolienė, G. Pre-Harvest LED Lighting Strategies for Reduced Nitrate Contents in Leafy Vegetables. Zemdirb.-Agric. 2018, 105, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.A.; Dzakovich, M.P.; Gomez, C.; Lopez, R.; Burr, J.F.; Hernández, R.; Kubota, C.; Currey, C.J.; Meng, Q.; Runkle, E.S.; et al. Light-Emitting Diodes in Horticulture. In Horticultural Reviews: Volume 43; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–88. ISBN 9781119107781. [Google Scholar]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current Status and Recent Achievements in the Field of Horticulture with the Use of Light-Emitting Diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Zhang, X.; Bian, Z.; Yuan, X.; Chen, X.; Lu, C. A Review on the Effects of Light-Emitting Diode (LED) Light on the Nutrients of Sprouts and Microgreens. Trends Food Sci. Technol. 2020, 99, 203–216. [Google Scholar] [CrossRef]
- Azad, M.O.K.; Kjaer, K.H.; Adnan, M.; Naznin, M.T.; Lim, J.D.; Sung, I.J.; Park, C.H.; Lim, Y.S. The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory. Agriculture 2020, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Field, D.L.; Ahmed, S.M.; Hasan, M.T.; Basher, M.K.; Alameh, K. LED Illumination for High-Quality High-Yield Crop Growth in Protected Cropping Environments. Plants 2021, 10, 2470. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Duchovskis, P. LED Lighting and Seasonality Effects Antioxidant Properties of Baby Leaf Lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Urbonavičiūtė, A.; Duchovskis, P.; Bliznikas, Z.; Vitta, P.; Žukauskas, A. Decrease in Nitrate Concentration in Leafy Vegetables Under a Solid-State Illuminator. HortScience 2009, 44, 1857–1860. [Google Scholar] [CrossRef]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Viršilė, A.; Duchovskis, P. Supplementary Red-LED Lighting and the Changes in Phytochemical Content of Two Baby Leaf Lettuce Varieties During Three Seasons. J. Food. Agric. Environ. 2012, 10, 701–706. [Google Scholar] [CrossRef]
- Zhou, W.L.; Liu, W.K.; Yang, Q.C. Quality Changes in Hydroponic Lettuce Grown under Pre-Harvest Short-Duration Continuous Light of Different Intensities. J. Hortic. Sci. Biotechnol. 2012, 87, 429–434. [Google Scholar] [CrossRef]
- Wanlai, Z.; Wenke, L.; Qichang, Y. Reducing Nitrate Content in Lettuce by Pre-Harvest Continuous Light Delivered by Red and Blue Light-Emitting Diodes. J. Plant Nutr. 2013, 36, 481–490. [Google Scholar] [CrossRef]
- Ouzounis, T.; Parjikolaei, R.B.; Fretté, X.; Rosenqvist, E.; Ottosen, C.-O. Predawn and High Intensity Application of Supplemental Blue Light Decreases the Quantum Yield of PSII and Enhances the Amount of Phenolic Acids, Flavonoids, and Pigments in Lactuca Sativa. Front. Plant Sci. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Oh, M.-M.; Son, K.-H. Short-Term Ultraviolet (UV)-A Light-Emitting Diode (LED) Radiation Improves Biomass and Bioactive Compounds of Kale. Front. Plant Sci. 2019, 10, 1042. [Google Scholar] [CrossRef] [Green Version]
- Kopsell, D.A.; Sams, C.E. Increases in Shoot Tissue Pigments, Glucosinolates, and Mineral Elements in Sprouting Broccoli after Exposure to Short-Duration Blue Light from Light Emitting Diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Li, X.; Tian, J.; Liu, H. Pre-Harvest Supplemental Blue Light Enhanced Antioxidant Activity of Flower Stalk in Chinese Kale during Storage. Plants 2021, 10, 1177. [Google Scholar] [CrossRef] [PubMed]
- Brazaitytė, A.; Sakalauskienė, S.; Viršilė, A.; Jankauskienė, J.; Samuolienė, G.; Sirtautas, R.; Vaštakaitė, V.; Miliauskienė, J.; Duchovskis, P.; Novičkovas, A.; et al. The Effect of Short-Term Red Lighting on Brassicaceae Microgreens Grown Indoors. Acta Hortic. 2016, 1123, 177–184. [Google Scholar] [CrossRef]
- Landi, M.; Zivcak, M.; Sytar, O.; Brestic, M.; Allakhverdiev, S.I. Plasticity of Photosynthetic Processes and the Accumulation of Secondary Metabolites in Plants in Response to Monochromatic Light Environments: A Review. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148131. [Google Scholar] [CrossRef] [PubMed]
- Soto, V.C.; González, R.E.; Galmarini, C.R. Bioactive Compounds in Vegetables, Is There Consistency in the Published Information? A Systematic Review. J. Hortic. Sci. Biotechnol. 2021, 96, 570–587. [Google Scholar] [CrossRef]
- Wojciechowska, R.; Kołton, A.; Długosz-Grochowska, O.; Żupnik, M.; Grzesiak, W. The Effect of LED Lighting on Photosynthetic Parameters and Weight of Lamb’s Lettuce (Valerianella Locusta). Folia Hortic. 2013, 25, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Długosz-Grochowska, O.; Kołton, A.; Wojciechowska, R. Modifying Folate and Polyphenol Concentrations in Lamb’s Lettuce by the Use of LED Supplemental Lighting during Cultivation in Greenhouses. J. Funct. Foods 2016, 26, 228–237. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional Value, Bioactive Compounds and Health Benefits of Lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kim, Y.B.; Li, X.; Seo, J.M.; Kim, S.-J.; Suzuki, T.; Chung, S.-O.; Park, S.U. Effects of Light-Emitting Diodes on Expression of Phenylpropanoid Biosynthetic Genes and Accumulation of Phenylpropanoids in Fagopyrum Tataricum Sprouts. J. Agric. Food Chem. 2014, 62, 4839–4845. [Google Scholar] [CrossRef]
- Nguyen, T.K.L.; Cho, K.M.; Lee, H.Y.; Cho, D.Y.; Lee, G.O.; Jang, S.N.; Lee, Y.; Kim, D.; Son, K.-H. Effects of White LED Lighting with Specific Shorter Blue and/or Green Wavelength on the Growth and Quality of Two Lettuce Cultivars in a Vertical Farming System. Agronomy 2021, 11, 2111. [Google Scholar] [CrossRef]
- Mao, P.; Duan, F.; Zheng, Y.; Yang, Q. Blue and UV-A Light Wavelengths Positively Affected Accumulation Profiles of Healthy Compounds in Pak-choi. J. Sci. Food Agric. 2021, 101, 1676–1684. [Google Scholar] [CrossRef]
- Taulavuori, K.; Pyysalo, A.; Taulavuori, E.; Julkunen-Tiitto, R. Responses of Phenolic Acid and Flavonoid Synthesis to Blue and Blue-Violet Light Depends on Plant Species. Environ. Exp. Bot. 2018, 150, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.; Chivkunova, O.; Zhigalova, T.; Solovchenko, A. In Situ Optical Properties of Foliar Flavonoids: Implication for Non-Destructive Estimation of Flavonoid Content. J. Plant Physiol. 2017, 218, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Y.; Liu, H.; Li, Y.; Liu, Y.; Hao, Y.; Lei, B. Supplemental Blue Light Increases Growth and Quality of Greenhouse Pak Choi Depending on Cultivar and Supplemental Light Intensity. J. Integr. Agric. 2018, 17, 2245–2256. [Google Scholar] [CrossRef] [Green Version]
- Kelly, N.; Vaštakaitė-Kairienė, V.; Runkle, E.S. Indoor lighting effects on plant nutritional compounds. In Plant Factory Basics, Applications and Advances; Kozai, T., Niu, G., Masabni, J., Eds.; Accademic Press: Cambridge, MA, USA, 2022; pp. 329–349. [Google Scholar]
- Vaštakaitė, V.; Viršilė, A.; Brazaitytė, A.; Samuolienė, G.; Jankauskienė, J.; Novičkovas, A.; Duchovskis, P. Pulsed Light-Emitting Diodes for a Higher Phytochemical Level in Microgreens. J. Agric. Food Chem. 2017, 65, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.; Niu, G.; Gu, M. Photosynthesis, Morphology, Yield, and Phytochemical Accumulation in Basil Plants Influenced by Substituting Green Light for Partial Red and/or Blue Light. Horts 2019, 54, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Son, K.-H.; Oh, M.-M. Growth, Photosynthetic and Antioxidant Parameters of Two Lettuce Cultivars as Affected by Red, Green, and Blue Light-Emitting Diodes. Hortic. Environ. Biotechnol. 2015, 56, 639–653. [Google Scholar] [CrossRef]
- Zhang, X.; Bian, Z.; Li, S.; Chen, X.; Lu, C. Comparative Analysis of Phenolic Compound Profiles, Antioxidant Capacities, and Expressions of Phenolic Biosynthesis-Related Genes in Soybean Microgreens Grown under Different Light Spectra. J. Agric. Food Chem. 2019, 67, 13577–13588. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, S.S.; Akbar, F.; Kanwal, F. Correlation of Different Spectral Lights with Biomass Accumulation and Production of Antioxidant Secondary Metabolites in Callus Cultures of Medicinally Important Prunella Vulgaris L. J. Photochem. Photobiol. B Biol. 2016, 159, 1–7. [Google Scholar] [CrossRef]
- Sellaro, R.; Crepy, M.; Trupkin, S.A.; Karayekov, E.; Buchovsky, A.S.; Rossi, C.; Casal, J.J. Cryptochrome as a Sensor of the Blue/Green Ratio of Natural Radiation in Arabidopsis. Plant Physiol. 2010, 154, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Battle, M.W.; Jones, M.A. Cryptochromes Integrate Green Light Signals into the Circadian System. Plant Cell Environ. 2020, 43, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t Ignore the Green Light: Exploring Diverse Roles in Plant Processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Ma, J.-Q.; Ma, C.-L.; Shen, S.-Y.; Liu, Y.-F.; Chen, L. Regulation of Growth and Flavonoid Formation of Tea Plants (Camellia sinensis) by Blue and Green Light. J. Agric. Food Chem. 2019, 67, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Viršilė, A.; Brazaitytė, A.; Sirtautas, R.; Duchovskis, P. Light Spectral Effects on Phenolic Compounds in Perilla frutescens Leaves as Related to the Leaf Age, Color and Duration of Exposure. Acta Hortic. 2017, 1170, 981–988. [Google Scholar] [CrossRef]
- Bliznikas, Z.; Zukauskas, A.; Samuolienė, G.; Viršilė, A.; Brazaitytė, A.; Jankauskienė, J.; Duchovskis, P.; Novičkovas, A. Effect of Supplementary Pre-harvest LED Lighting on the Antioxidant and Nutritional Properties of Green Vegetables. Acta Hortic. 2012, 939, 85–91. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Oroian, M.; Stoleru, T.; Burducea, M.; Pintilie, O.; Zamfirache, M.-M. Blue and Red LED Illumination Improves Growth and Bioactive Compounds Contents in Acyanic and Cyanic Ocimum Basilicum L. Microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Xu, J.; Wang, W.; Rajashekar, C.B. The Effect of Supplemental Blue, Red and Far-Red Light on the Growth and the Nutritional Quality of Red and Green Leaf Lettuce. Am. J. Plant Sci. 2019, 10, 2219–2235. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Zivcak, M.; Bruckova, K.; Brestic, M.; Hemmerich, I.; Rauh, C.; Simko, I. Shift in Accumulation of Flavonoids and Phenolic Acids in Lettuce Attributable to Changes in Ultraviolet Radiation and Temperature. Sci. Hortic. 2018, 239, 193–204. [Google Scholar] [CrossRef]
- Bunning, M.L.; Kendall, P.A.; Stone, M.B.; Stonaker, F.H.; Stushnoff, C. Effects of Seasonal Variation on Sensory Properties and Total Phenolic Content of 5 Lettuce Cultivars. J. Food Sci. 2010, 75, S156–S161. [Google Scholar] [CrossRef]
- Marin, A.; Ferreres, F.; Barberá, G.G.; Gil, M.I. Weather Variability Influences Color and Phenolic Content of Pigmented Baby Leaf Lettuces throughout the Season. J. Agric. Food Chem. 2015, 63, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Oh, I.-N.; Kim, J.; Jung, D.; Cuong, N.P.; Kim, Y.; Lee, J.; Kwon, O.; Park, S.U.; Lim, Y.; et al. Phenolic Compound Profiles and Their Seasonal Variations in New Red-Phenotype Head-Forming Chinese Cabbages. LWT 2018, 90, 433–439. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phenolic Compounds | Lighting | Source of Variance | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HPS | 400 nm | 455 nm | 455 + 530 nm | 530 nm | 660 nm | L | S | L ×S | |||||||
| Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | ||||
| Caffeic a. | 0.068 b | 0.015 gh | 0.033 ef | 0.040 def | 0.047 de | 0.05 cd | 0.029 efg | 0.066 bc | 0.025 fg | 0.007 h | 0.042 def | 0.151 a | * | * | |
| Chicoric a. | 2.57 cd | 3.89 bc | 2.43 d | 5.07 ab | 2.24 d | 3.83 bc | 2.57 cd | 5.51 a | 2.26 d | 3.87 bc | 1.97 d | 2.73 cd | * | * | |
| Chlorogenic a. | 0.81 de | 1.04 bc | 1.07 bc | 0.98 cd | 0.98 cd | 1.16 bc | 0.74 e | 1.46 a | 0.77 e | 0.72 e | 0.75 e | 1.22 b | * | * | |
| Gallic a. | 0.049 a | 0.029 bc | 0.034 abc | 0.020 bc | 0.037 ab | 0.019 bc | 0.031 abc | 0.022 bc | 0.028 bc | 0.018 c | 0.026 bc | 0.020 bc | * | * | |
| o-coumaric a. | 0.038 c | 0.141 b | 0.034 c | 0.230 a | 0.042 c | 0.057 c | 0.056 c | 0.038 c | 0.044 c | 0.043 c | 0.039 c | 0.227 a | * | * | |
| p-coumaric a | 0.082 a | 0.008 f | 0.052 b | 0.016 ef | 0.039 c | 0.016 ef | 0.030 cd | 0.010 ef | 0.030 cd | 0.033 c | 0.021 de | 0.016 ef | * | * | |
| Protocatechuic a. | 0.177 a | 0.071 c | 0.138 b | 0.023 d | 0.133 b | 0.022 d | 0.073 c | 0.023 d | 0.160 ab | 0.020 d | 0.142 ab | 0.093 c | * | * | |
| Rosmarinic a. | 0.641 bc | 0.049 f | 0.596 c | 0.076 f | 0.732 a | 0.520 cd | 0.563 c | 0.424 d | 0.797 a | 0.235 e | 0.601 c | 0.033 f | * | * | |
| Apigenin | 0.844 a | 0.333 e | 0.637 bc | 0.082 f | 0.725 ab | 0.358 e | 0.458 de | 0.044 f | 0.715 b | 0.381 e | 0.521 cd | 0.089 f | * | * | |
| Epicatechin | 0.165 cd | 0.280 b | 0.080 de | 0.432 a | 0.069 de | 0.137 cde | 0.080 de | 0.199 bc | 0.047 e | 0.187 bc | 0.123 cde | 0.515 a | * | * | * |
| Kaempferol | 0.034 ab | 0.020 cd | 0.038 a | 0.013 d | 0.015 cd | 0.026 bc | 0.026 bc | 0.020 cd | 0.035 ab | 0.024 bcd | 0.021 cd | 0.022 cd | * | * | |
| Myricetin | 0.118 a | 0.051 c | 0.089 b | 0.035 c | 0.107 ab | 0.033 c | 0.053 c | 0.038 c | 0.099 ab | 0.037 c | 0.052 c | 0.040 c | * | * | |
| Quercetin | 0.043 d | 0.022 de | 0.036 de | 0.180 a | 0.026 de | 0.015 e | 0.036 de | 0.179 a | 0.071 c | 0.014 e | 0.028 de | 0.139 b | * | * | * |
| Rutin | 0.026 b | 0.042 b | 0.013 c | 0.061 b | 0.026 b | 0.046 b | 0.027 b | 0.049 b | 0.016 c | 0.019 c | 0.012 c | 0.361 a | * | * | * |
| Total | 5.67 cde | 5.99 cd | 5.28 cde | 7.25 ab | 5.21 cde | 6.29 bc | 4.78 de | 8.08 a | 5.10 cde | 5.61 cde | 4.34 e | 5.65 cde | * | * | |
| Phenolic Compounds | Lighting | Source of Variance | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HPS | 400 nm | 455 nm | 455 + 530 nm | 530 nm | 660 nm | L | S | L ×S | |||||||
| Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | Spring | Autumn | ||||
| Caffeic a. | 0.222 b | 0.036 ef | 0.217 b | 0.043 e | 0.269 a | 0.018 ef | 0.116 d | 0.039 e | 0.181 c | 0.098 d | 0.180 c | 0.009 f | * | * | |
| Chicoric a. | 4.47 b | 0.28 e | 2.40 d | 0.40 e | 3.65 c | 0.08 e | 5.42 a | 0.37 e | 1.78 d | 0.29 e | 1.89 d | 0.13 e | * | * | |
| Chlorogenic a. | 2.59 b | 1.11 de | 1.39 d | 2.15 c | 2.04 c | 0.33 f | 5.08 a | 1.44 d | 1.25 d | 1.12 de | 2.01 c | 0.80 e | * | * | * |
| Gallic a. | 0.059 a | 0.012 f | 0.030 cd | 0.036 bc | 0.026 cde | 0.016 def | 0.049 ab | 0.013 ef | 0.044 b | 0.009 f | 0.027 cd | 0.017 def | * | * | |
| o-coumaric a. | 0.303 ab | 0.190 bc | 0.043 c | 0.382 a | 0.070 bc | 0.035 c | 0.087 bc | 0.047 c | 0.035 c | 0.115 bc | 0.036 c | 0.067 bc | * | * | |
| p-coumaric a | 0.021 ab | 0.005 ab | 0.021 ab | 0.027 ab | 0.025 ab | 0.005 ab | 0.052 a | 0.003 b | 0.024 ab | 0.003 b | 0.038 ab | 0.004 ab | * | * | |
| Protocatechuic a. | 0.210 a | 0.012 d | 0.122 bc | 0.022 d | 0.101 c | 0.013 d | 0.148 b | 0.019 d | 0.098 c | 0.022 d | 0.156 b | 0.029 d | * | * | |
| Rosmarinic a. | 0.098 de | 0.006 e | 0.723 c | 0.015 e | 1.153 b | 0.007 e | 2.318 a | 0.005 e | 0.453 cd | 0.006 e | 0.771 c | 0.006 e | * | * | |
| Apigenin | 0.390 b | 0.010 e | 0.428 b | 0.051 e | 0.255 cd | 0.044 e | 0.597 a | 0.010 e | 0.298 c | 0.009 e | 0.196 d | 0.061 e | * | * | |
| Epicatechin | 0.493 a | 0.095 bcd | 0.088 cd | 0.056 d | 0.132 bc | 0.050 d | 0.146 b | 0.067 d | 0.480 a | 0.078 d | 0.143 b | 0.050 d | * | * | * |
| Kaempferol | 0.012 bc | 0.002 de | 0.012 bcd | 0.005 cde | 0.014 b | 0.003 cde | 0.016 b | 0.004 cde | 0.012 bc | 0.001 e | 0.031 a | 0.002 e | * | * | |
| Myricetin | 0.031 abcd | 0.027 bcd | 0.027 bcd | 0.052 a | 0.029 bcd | 0.017 cd | 0.036 abc | 0.014 d | 0.045 ab | 0.011 d | 0.020 cd | 0.015 d | * | ||
| Quercetin | 0.025 ab | 0.005 d | 0.018 c | 0.005 d | 0.017 c | 0.005 d | 0.019 bc | 0.006 d | 0.029 a | 0.005 d | 0.023 abc | 0.005 d | * | * | |
| Rutin | 0.179 f | 2.809 c | 0.041 f | 5.101 a | 0.062 f | 1.299 e | 0.091 f | 3.390 b | 0.053 f | 2.478 cd | 0.027 f | 2.187 d | * | * | |
| Total | 9.11 b | 4.59 ef | 5.56 d | 8.35 bc | 7.85 c | 1.92 g | 14.17 a | 5.43 d | 4.78 de | 4.24 ef | 5.55 d | 3.38 f | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Rasiukevičiūtė, N.; Viršilė, A.; Miliauskienė, J.; Laužikė, K.; Valiuškaitė, A.; Dėnė, L.; Chrapačienė, S.; et al. Phenolic Compounds Content Evaluation of Lettuce Grown under Short-Term Preharvest Daytime or Nighttime Supplemental LEDs. Plants 2022, 11, 1123. https://doi.org/10.3390/plants11091123
Brazaitytė A, Vaštakaitė-Kairienė V, Sutulienė R, Rasiukevičiūtė N, Viršilė A, Miliauskienė J, Laužikė K, Valiuškaitė A, Dėnė L, Chrapačienė S, et al. Phenolic Compounds Content Evaluation of Lettuce Grown under Short-Term Preharvest Daytime or Nighttime Supplemental LEDs. Plants. 2022; 11(9):1123. https://doi.org/10.3390/plants11091123
Chicago/Turabian StyleBrazaitytė, Aušra, Viktorija Vaštakaitė-Kairienė, Rūta Sutulienė, Neringa Rasiukevičiūtė, Akvilė Viršilė, Jurga Miliauskienė, Kristina Laužikė, Alma Valiuškaitė, Lina Dėnė, Simona Chrapačienė, and et al. 2022. "Phenolic Compounds Content Evaluation of Lettuce Grown under Short-Term Preharvest Daytime or Nighttime Supplemental LEDs" Plants 11, no. 9: 1123. https://doi.org/10.3390/plants11091123