
Breeding Diploid F1 Hybrid Potatoes for Propagation from Botanical Seed (TPS): Comparisons with Theory and Other Crops


Abstract
:1. Introduction
2. Mendel, Darwin, and Fisher
3. Diploid Germplasm
4. Breeding Methods for Diploid Potatoes
5. Successful Self-Pollination
Self-Pollination in Other Outbreeding Crops
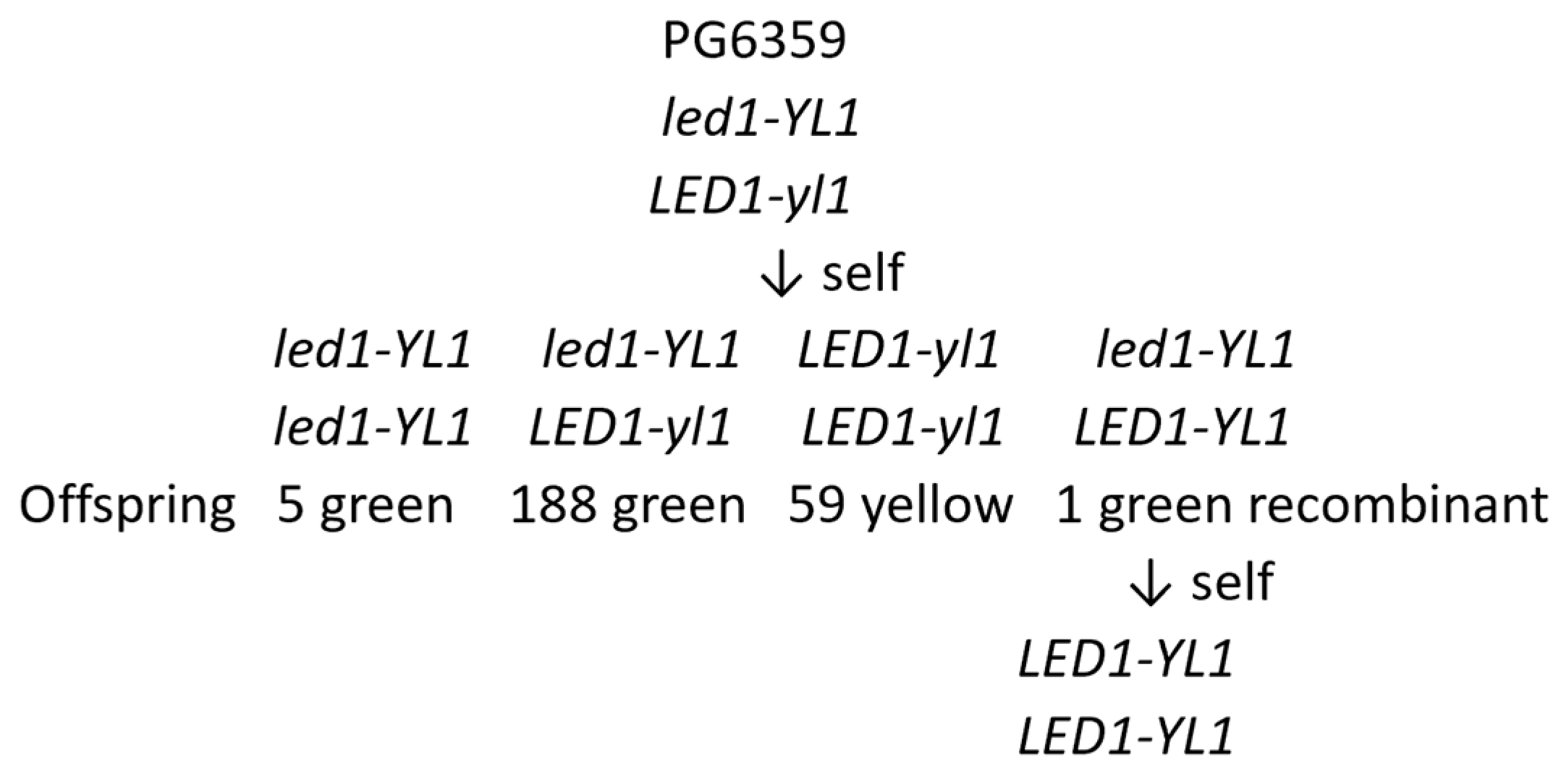
6. Self-Compatible Diploid Potatoes
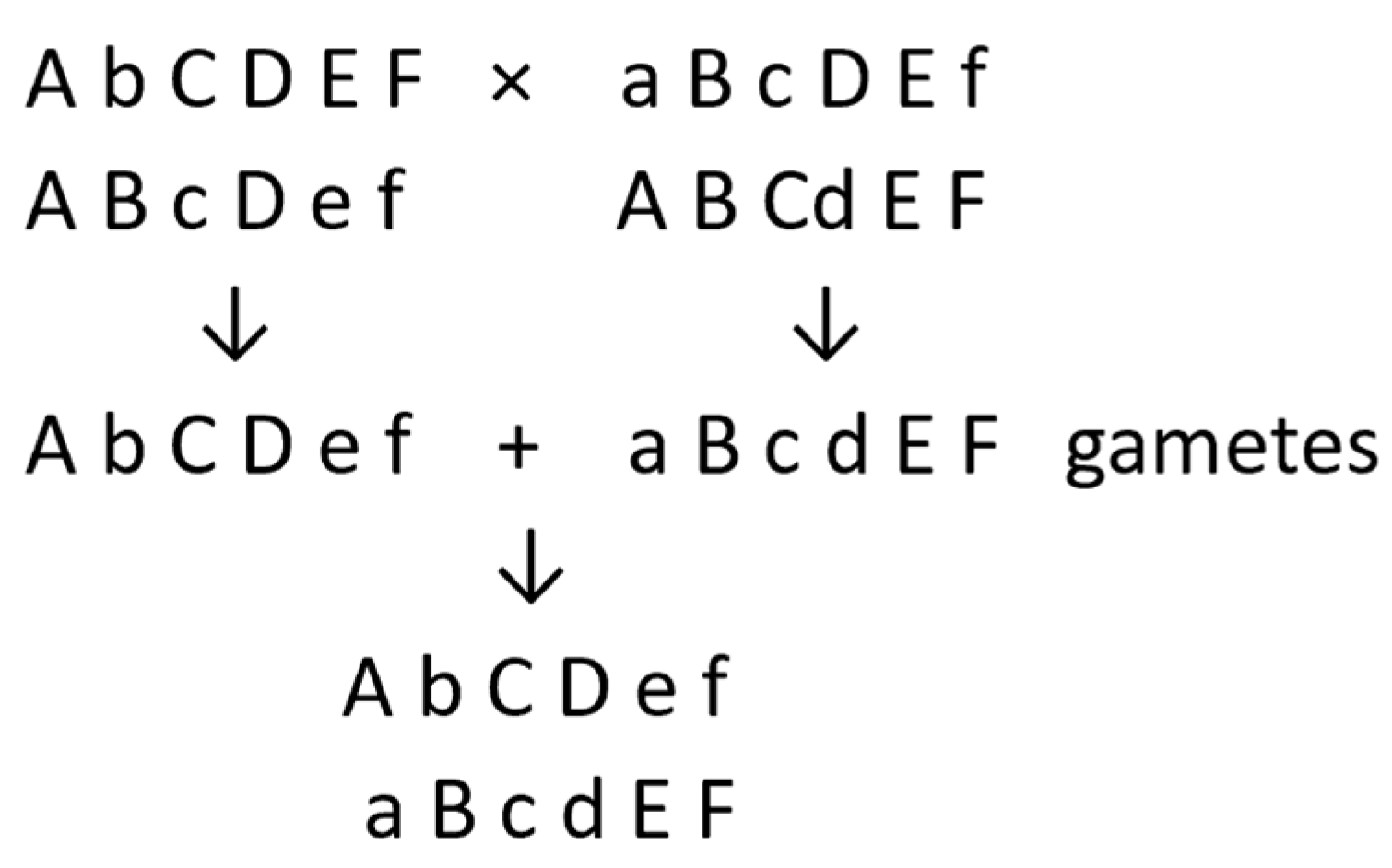
7. Ability to Produce Homozygous Inbred Lines
7.1. Theory
7.2. Diploid Potatoes in Practice
7.3. Maize in Practice
7.4. Brassica Vegetables in Practice
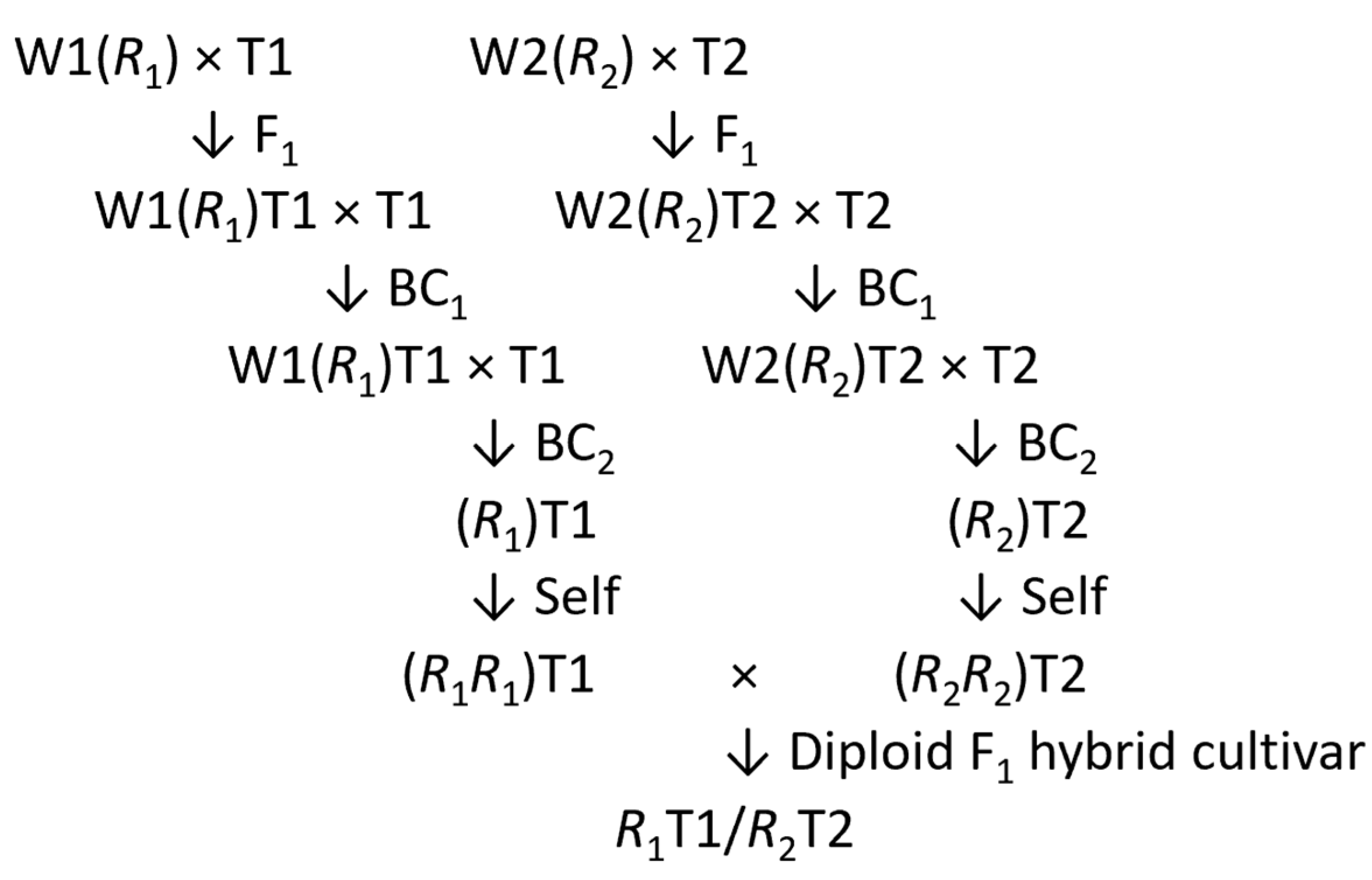
8. Cycles of Inbreeding and Crossbreeding to Produce Hybrids
8.1. Introgression (Backcross) Breeding and Chromosome Segment Substitution Lines
8.2. Theory for Quantitative Traits
| Genotypes of F1 hybrids | AA | Aa | aa | |
| Genotype frequency | p2 | 2pq | q2 | |
| Frequency of A in F1 hybrids | 1 | ½ | 0 | |
| F1 hybrid means | a | d | −a | |
| Genotypes of inbred lines | AA | AA | aa | aa |
| Genotype frequency | p2 | pq | pq | q2 |
| Frequency of A in inbred lines | 1 | 1 | 0 | 0 |
| Inbred line means | a | a | −a | −a |
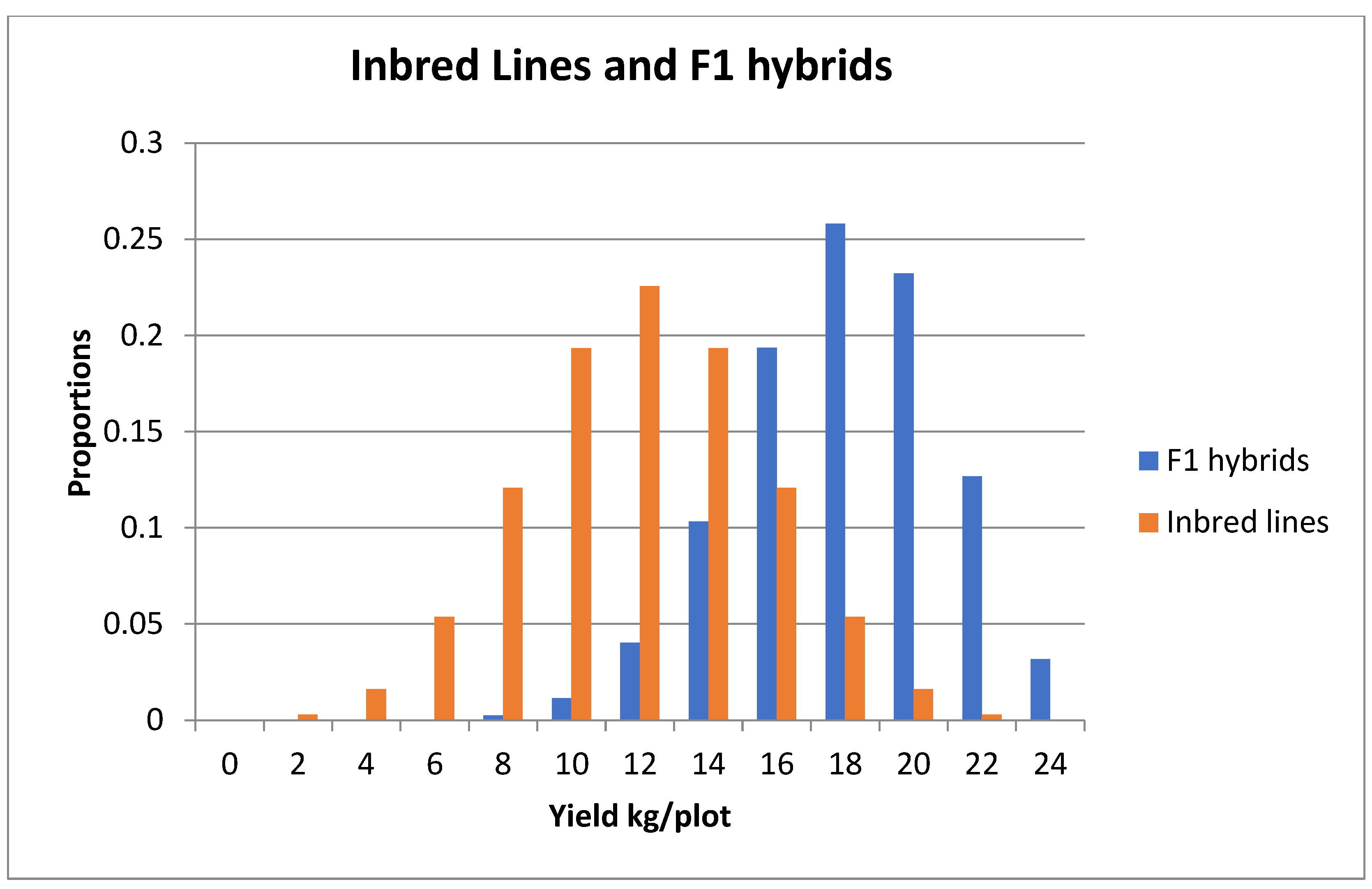
8.3. Simple Example of Inbreeding and Crossbreeding
8.4. Likely Practice for Diploid F1 Hybrid Potatoes
9. Seed Production for Diploid F1 Hybrid Potatoes
10. Progress and Success in Diploid F1 Hybrid Potato Breeding
10.1. Registration, Multiplication, and Protection of New Cultivars
10.2. Planting Material for Farmers
11. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spooner, D.M.; McLean, K.; Ramsay, G.; Waugh, R.; Bryan, G.J. A single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proc. Natl. Acad. Sci. USA 2005, 102, 14694–14699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkes, J.G. The Potato: Evolution, Biodiversity and Genetic Resources; Belhaven Press: London, UK, 1990. [Google Scholar]
- Golmirzaie, A.M.; Malagamba, P.; Pallais, N. Breeding potatoes based on true seed propagation. In Potato Genetics; Bradshaw, J.E., Mackay, G.R., Eds.; CAB International: Wallingford, UK, 1994; pp. 499–513. [Google Scholar]
- Chilver, A.; Walker, T.S.; Khatana, V.; Fano, R.; Suherman, R.; Rizk, A. On-farm profitability and prospects for true potato seed (TPS). In Genetic Improvement of Solanaceous Crops, Volume I: Potato; Razdan, M.K., Mattoo, A.K., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2005; pp. 39–63. [Google Scholar]
- Lindhout, P.; Meijer, D.; Schotte, T.; Hutten, R.C.B.; Visser, R.G.F.; van Eck, H.J. Towards F1 hybrid seed potato breeding. Potato Res. 2011, 54, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Jansky, S.H.; Charkowski, A.O.; Douches, D.S.; Gusmini, G.; Richael, C.; Bethke, P.C.; Spooner, D.M.; Novy, R.G.; De Jong, H.; De Jong, W.S.; et al. Reinventing potato as a diploid inbred line-based crop. Crop Sci. 2016, 56, 1412–1422. [Google Scholar] [CrossRef] [Green Version]
- Jansky, S.H.; Spooner, D.M. The evolution of potato breeding. Plant Breed. Rev. 2018, 41, 169–214. [Google Scholar]
- Bachem, C.W.B.; van Eck, H.J.; de Vries, M.E. Understanding genetic load in potato for hybrid diploid breeding. Mol. Plant 2019, 12, 896–898. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Plant Breeding: Past, Present and Future; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Darwin, C. The Effects of Cross and Self Fertilisation in the Vegetable Kingdom; John Murray: London, UK, 1876. [Google Scholar]
- Mendel, G. Experiments in Plant Hybridisation; Oliver & Boyd: Edinburgh, UK, 1965. [Google Scholar]
- Bateson, W.; Mendel, G. Mendel’s Principles of Heredity; Cambridge University Press: Cambridge, UK, 1902. [Google Scholar]
- Bradshaw, J.E. A brief history of the impact of potato genetics on the breeding of tetraploid potato cultivars for tuber propagation. Potato Res. 2022, 1–41. [Google Scholar] [CrossRef]
- Darwin, C. The Different Forms of Flowers on Plants of the Same Species; John Murray: London, UK, 1877. [Google Scholar]
- Fisher, R.A. The Theory of Inbreeding, 2nd ed.; Oliver and Boyd: London, UK, 1965. [Google Scholar]
- Watanabe, K.N.; Orrillo, M.; Perez, S.; Crusadol, J.; Watanabe, J.A. Testing yield of diploid potato breeding lines for cultivar development. Jpn. J. Breed. 1996, 46, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Burgos, G.; Amoros, W.; Morote, M.; Stangoulis, J.; Bonierbale, M. Iron and zinc concentration of native Andean potato cultivars from a human nutrition perspective. J. Sci. Food Agric. 2007, 87, 668–675. [Google Scholar] [CrossRef]
- Paget, M.; Amoros, W.; Salas, E.; Eyzaguirre, R.; Alspach, P.; Apiolaza, L.; Noble, A.; Bonierbale, M. Genetic evaluation of micronutrient traits in diploid potato from a base population of Andean landrace cultivars. Crop Sci. 2014, 54, 1949–1959. [Google Scholar] [CrossRef]
- Amoros, W.; Salas, E.; Hualla, V.; Burgos, G.; De Boeck, B.; Eyzaguirre, R.; zum Felde, T.; Bonierbale, M. Heritability and genetic gains for iron and zinc concentration in diploid potato. Crop Sci. 2020, 60, 1884–1896. [Google Scholar] [CrossRef]
- Carroll, C.P. A mass-selection method for the acclimatization and improvement of edible diploid potatoes in the United Kingdom. J. Agric. Sci. Camb. 1982, 99, 631–640. [Google Scholar] [CrossRef]
- Haynes, K.G.; Lu, W. Improvement at the diploid species level. In Genetic Improvement of Solanaceous Crops, Volume I: Potato; Razdan, M.K., Mattoo, A.K., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2005; pp. 101–114. [Google Scholar]
- Haynes, K.G.; Guedes, M.L. Self-compatibility in a diploid hybrid population of Solanum phureja—S. stenotomum. Am. J. Potato Res. 2018, 95, 729–734. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato breeding at the Scottish Plant Breeding Station and the Scottish Crop Research Institute: 1920–2008. Potato Res. 2009, 52, 141–172. [Google Scholar] [CrossRef]
- Hougas, R.W.; Peloquin, S.J. The potential of potato haploids in breeding and genetic research. Am. Potato J. 1958, 35, 701–707. [Google Scholar] [CrossRef]
- Hougas, R.W.; Peloquin, S.J.; Ross, R.W. Haploids of the common potato. J. Hered. 1958, 49, 103–107. [Google Scholar] [CrossRef]
- Hougas, R.W.; Peloquin, S.J.; Gabert, A.C. Effect of seed-parent and pollinator on the frequency of haploids in Solanum tuberosum. Crop Sci. 1964, 4, 593–595. [Google Scholar] [CrossRef]
- Ortiz, R.; Mihovilovich, E. Genetics and cytogenetics of the potato. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 219–247. [Google Scholar]
- Busse, J.S.; Jansky, S.H.; Agha, H.I.; Carley, C.A.S.; Shannon, L.M.; Bethke, P.C. A high throughput method for generating dihaploids from tetraploid potato. Am. J. Potato Res. 2021, 98, 304–314. [Google Scholar] [CrossRef]
- Peloquin, S.; Hougas, R. Fertility in two haploids of Solanum tuberosum. Science 1958, 128, 1340–1342. [Google Scholar] [CrossRef]
- Peloquin, S.J.; Hougas, R.W. Genetic variation among haploids of the common potato. Am. Potato J. 1960, 37, 289–297. [Google Scholar] [CrossRef]
- Cipar, M.S.; Peloquin, S.J.; Hougas, R.W. Inheritance of incompatibility in hybrids between Solanum tuberosum haploids and diploid species. Euphytica 1964, 13, 163–172. [Google Scholar] [CrossRef]
- Dodds, K.S. The history and relationships of cultivated potatoes. In Essays in Crop Plant Evolution; Hutchinson, J.B., Ed.; Cambridge University Press: Cambridge, UK, 1965; pp. 123–141. [Google Scholar]
- Gebhardt, C.; Ritter, E.; Barone, A.; Debener, T.; Walkemeier, B.; Schachtschabel, U.; Kaufmann, H.; Thompson, R.D.; Bonierbale, M.W.; Ganal, M.W.; et al. RFLP maps of potato and their alignment with the homoeologous tomato genome. Theor. Appl. Genet. 1991, 83, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, H.; Salamini, F.; Thompson, R.D. Sequence variability and gene structure at the self-incompatibility locus of Solanum tuberosum. Mol. Gen. Genet. 1991, 226, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.E. Potato Breeding: Theory and Practice; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Hanneman, R.E., Jr. Self-fertility in Solanum chacoense. Am. Potato J. 1985, 62, 428–429. [Google Scholar]
- Hosaka, K.; Hanneman, R.E., Jr. Genetics of self-compatibility in a self-incompatible wild diploid potato species Solanum chacoense. 1. Detection of an S-locus inhibitor (Sli) gene. Euphytica 1998, 99, 191–197. [Google Scholar] [CrossRef]
- Hosaka, K.; Hanneman, R.E., Jr. Genetics of self-compatibility in a self-incompatible wild diploid potato species Solanum chacoense. 2. Localization of an S locus inhibitor (Sli) gene on the potato genome using DNA markers. Euphytica 1998, 103, 265–271. [Google Scholar] [CrossRef]
- Jansky, S.; Chung, Y.S.; Kittipadukal, P. M6: A diploid potato inbred line for use in breeding and genetics research. J. Plant Regist. 2014, 8, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Leisner, C.P.; Hamilton, J.P.; Crisovan, E.; Crisovan, E.; Manrique-Carpintero, N.C.; Marand, A.P.; Newton, L.; Pham, G.M.; Jiang, J.; Douches, D.S.; et al. Genome sequence of M6, a diploid inbred clone of the high-glycoalkaloid-producing tuber-bearing potato species Solanum chacoense, reveals residual heterozygosity. Plant J. 2018, 94, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.P.; Hansey, C.N.; Whitty, B.R.; Stoffel, K.; Massa, A.N.; Deynze, A.V.; De Jong, W.S.; Douches, D.S.; Buell, C.R. Single nucleotide polymorphism discovery in elite North American potato germplasm. BMC Genom. 2011, 12, 302. [Google Scholar] [CrossRef] [Green Version]
- Endelman, J.B.; Jansky, S.H. Genetic mapping with an inbred line-derived F2 population in potato. Theor. Appl. Genet. 2016, 129, 935–943. [Google Scholar] [CrossRef]
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.R.; Billings, G.; Coombs, J.; Buell, C.R.; Enciso-Rodríguez, F.; Douches, D.S. Self-fertility and resistance to the Colorado potato beetle (Leptinotarsa decemlineata) in a diploid Solanum chacoense recombinant inbred line population. Crop Sci. 2021, 61, 3392–3414. [Google Scholar] [CrossRef]
- Lindhout, P.; de Vries, M.; ter Maat, M.; Ying, S.; Viquez-Zamora, M.; van Heusden, S. Hybrid potato breeding for improved varieties. In Achieving Sustainable Cultivation of Potatoes Volume 1: Breeding Improved Varieties; Wang-Pruski, G., Ed.; Burleigh Dodds: Cambridge, UK, 2018; pp. 99–122. [Google Scholar]
- Clot, C.R.; Polzer, C.; Prodhomme, C.; Schuit, C.; Engelen, C.J.M.; Hutten, R.C.B.; van Eck, H.J. The origin and widespread occurrence of Sli-based self-compatibility in potato. Theor. Appl. Genet. 2020, 133, 2713–2728. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, C.E. The origination and test culture of seedling potatoes. Trans. N. Y. State Agric. Soc. 1863, 23, 89–134. [Google Scholar]
- Kaiser, N.R.; Jansky, S.; Coombs, J.J.; Collins, P.; Alsahlany, M.; Douches, D.S. Assessing the contribution of Sli to self-compatibility in North American diploid potato germplasm using KASP™ markers. Am. J. Potato Res. 2021, 98, 104–113. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, C.; Zhang, B.; Tang, F.; Li, F.; Liao, Q.; Tang, D.; Peng, Z.; Jia, Y.; Gao, M.; et al. A non-S-locus F-box gene breaks self-incompatibility in diploid potatoes. Nat. Commun. 2021, 12, 4142. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Enciso-Rodriguez, F.; Manrique-Carpintero, N.C.; Nadakuduti, S.S.; Buell, C.R.; Zarka, D.; Douches, D.S. Overcoming self-incompatibility in diploid potato using CRISPR-Cas9. Front Plant Sci. 2019, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- East, E.M. Inbreeding in corn. Conn. Agric. Exp. Stn. Rep. 1907, 419–428. [Google Scholar]
- Shull, G.H. The composition of a field of maize. Am. Breeders’ Assoc. Rep. 1908, 4, 296–301. [Google Scholar] [CrossRef]
- Shull, G.H. A pure-line method of corn breeding. Am. Breeders’ Assoc. Rep. 1909, 5, 51–59. [Google Scholar] [CrossRef]
- Jones, D.F. The effects of inbreeding and crossbreeding upon development. Conn. Agric. Exp. Stn. Bull. 1918, 207, 5–100. [Google Scholar]
- Troyer, A.F. Adaptedness and heterosis in corn and mule hybrids. Crop Sci. 2006, 46, 528–543. [Google Scholar] [CrossRef]
- Shigyo, M.; Kik, C. Onion. In Vegetables II, Handbook of Plant Breeding 2; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 121–159. [Google Scholar]
- Singh, H.; Khar, A. Perspectives of onion hybrid breeding in India: An overview. Indian J. Agric. Sci. 2021, 91, 1426–1432. [Google Scholar]
- Simon, P.W.; Freeman, R.E.; Vieira, J.V.; Boiteux, L.S.; Briard, M.; Nothnagel, T.; Michalik, B.; Kwon, Y.-S. Carrot. In Vegetables II, Handbook of Plant Breeding 2; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 327–357. [Google Scholar]
- Thompson, K.F. Self-incompatibility in marrow-stem kale, Brassica oleracea var. acephala. I. Demonstration of a sporophytic system. J. Genet. 1957, 55, 45–60. [Google Scholar] [CrossRef]
- Thompson, K.F. Breeding problems in kale (Brassica oleracea) with particular reference to marrow-stem kale. Rep. Plant Breed. Inst. 1965, 66, 7–34. [Google Scholar]
- Lundqvist, A. Self-incompatibility in rye. I. Genetic control in the diploid. Hereditas 1956, 42, 293–348. [Google Scholar] [CrossRef]
- Ossent, H.P. Zehn Jahre Roggenzüchtung in Müncheberg. Züchter 1938, 10, 255–261. [Google Scholar] [CrossRef]
- Wricke, G. Untersuchungen zur Vererbung der Selbstfertilität beim Roggen (Secale cereale). Theor. Appl. Genet. 1969, 39, 371–378. [Google Scholar] [CrossRef]
- Wolski, T. Studies on breeding of rye. Genet. Pol. Warszawa/Poznań 1970, 11, 1–26. [Google Scholar]
- Geiger, H.H.; Schnell, F.W. Cytoplasmic male sterility in rye (Secale cereale L.). Crop Sci. 1970, 10, 590–593. [Google Scholar] [CrossRef]
- Geiger, H.H.; Miedaner, T. Rye Breeding. In Cereals, Handbook of Plant Breeding 3; Carena, M.J., Ed.; Springer: New York, NY, USA, 2009; pp. 157–181. [Google Scholar]
- Savitsky, V.F. Monogerm sugar beets in the United States. Proc. ASSBT 1950, 6, 156–159. [Google Scholar]
- Biancardi, E.; McGrath, J.M.; Panella, L.W.; Lewellen, R.T.; Stevanato, P. Sugar Beet. In Root and Tuber Crops, Handbook of Plant Breeding 7; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 173–219. [Google Scholar]
- Alsahlany, M.; Enciso-Rodriguez, F.; Lopez-Cruz, M.; Coombs, J.; Douches, D.S. Developing self-compatible diploid potato germplasm through recurrent selection. Euphytica 2021, 217, 47. [Google Scholar] [CrossRef]
- Ortiz, R. Breeding for potato production from true seed. Plant Breed. Abstr. 1997, 67, 1355–1360. [Google Scholar]
- Simmonds, N.W. A review of potato propagation by means of seed, as distinct from clonal propagation by tubers. Potato Res. 1997, 40, 191–214. [Google Scholar] [CrossRef]
- Gopal, J.; Ortiz, R. True potato seed. In Handbook of Potato Production, Improvement and Postharvest Management; Gopal, J., Khurana, S.M.P., Eds.; The Haworth Press: Binghampton, NY, USA, 2006; pp. 557–585. [Google Scholar]
- Hayman, B.I.; Mather, K. The progress of inbreeding when homozygotes are at a disadvantage. Heredity 1953, 7, 165–183. [Google Scholar] [CrossRef]
- Mather, K. Genetical Structure of Populations; Chapman and Hall: London, UK, 1973. [Google Scholar]
- Gale, J.S. Population Genetics.; Blackie: Glasgow, UK, 1980. [Google Scholar]
- Bennett, J.H. Junctions in inbreeding. Genetica 1953, 26, 392–406. [Google Scholar] [CrossRef]
- Dobzhansky, T. Genetics of the Evolutionary Process; Columbia University Press: New York, NY, USA, 1970. [Google Scholar]
- Lian, Q.; Tang, D.; Bai, Z.; Qi, J.; Lu, F.; Huang, S.; Zhang, C. Acquisition of deleterious mutations during potato polyploidization: Variations of deleterious mutations in potato. J. Integr. Plant Biol. 2019, 61, 7–11. [Google Scholar] [CrossRef]
- Hoopes, G.; Meng, X.; Hamilton, J.P.; Achakkagari, S.R.; de Alves Freitas Guesdes, F.; Bolger, M.E.; Coombs, J.J.; Esselink, D.; Kaiser, N.R.; Kodde, L.; et al. Phased, chromosome-scale genome assemblies of tetraploid potato reveals a complex genome, transcriptome, and predicted proteome landscape underpinning genetic diversity. Mol. Plant 2022, 15, 520–536. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, P.; Tang, D.; Yang, Z.; Lu, F.; Qi, J.; Tawari, N.R.; Shang, Y.; Li, C.; Huang, S. The genetic basis of inbreeding depression in potato. Nat. Genet. 2019, 51, 374–378. [Google Scholar] [CrossRef]
- Phumichai, C.; Mori, M.; Kobayashi, A.; Kamijima, O.; Hosaka, K. Toward the development of highly homozygous diploid potato lines using the self-compatibility controlling Sli gene. Genome 2005, 48, 977–984. [Google Scholar] [CrossRef] [Green Version]
- van Lieshout, N.; van der Burgt, A.; de Vries, M.E.; ter Maat, M.; Eickholt, D.; Esselink, D.; van Kaauwen, M.P.W.; Kodde, L.P.; Visser, R.G.F.; Lindhout, P.; et al. Solyntus, the new highly contiguous reference genome for potato (Solanum tuberosum). G3 Genes Genomes Genet. 2020, 10, 3489–3495. [Google Scholar] [CrossRef] [PubMed]
- Pham, G.M.; Hamilton, J.P.; Wood, J.C.; Burke, J.T.; Zhao, H.; Vaillancourt, B.; Ou, S.; Jiang, J.; Buell, C.R. Construction of a chromosome-scale long-read reference genome assembly for potato. GigaScience 2020, 9, giaa100. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Tang, D.; Huang, W.; Yang, Z.; Zhang, Y.; Hamilton, J.P.; Visser, R.G.F.; Bachem, C.W.B.; Buell, C.R.; Zhang, Z.; et al. Haplotype-resolved genome analyses of a heterozygous diploid potato. Nat. Genet. 2020, 52, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Z.; Tang, D.; Zhu, Y.; Wang, P.; Li, D.; Zhu, G.; Xiong, X.; Shang, Y.; Li, C.; et al. Genome design of hybrid potato. Cell 2021, 184, 3873–3883. [Google Scholar] [CrossRef]
- Veilleux, R.E. Cell and tissue culture of potato (Solanaceae). In Genetic Improvement of Solanaceous Crops, Volume I: Potato; Razdan, M.K., Mattoo, A.K., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2005; pp. 185–208. [Google Scholar]
- Paz, M.M.; Veilleux, R.E. Genetic diversity based on randomly amplified polymorphic DNA (RAPD) and its relationship with the performance of diploid potato hybrids. J. Am. Soc. Hortic. Sci. 1997, 122, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Paz, M.M.; Veilleux, R.E. Influence of culture medium and in vitro conditions on shoot regeneration in Solanum phureja monoploids and fertility of regenerated doubled monoploids. Plant Breed. 1999, 118, 53–57. [Google Scholar] [CrossRef]
- Betrán, J. Hybrid breeding in maize. In Principles of Plant Genetics and Breeding, 3rd ed.; Acquaah, G., Ed.; Wiley Blackwell: Chichester, UK, 2021; pp. 680–685. [Google Scholar]
- Jones, D.F. Continued inbreeding in maize. Genetics 1939, 24, 462–473. [Google Scholar] [CrossRef]
- Lindstrom, E.W. Analysis of Modern Maize Breeding Principles and Methods. In Proceedings of the Seventh International Genetical Congress, Edinburgh, UK, 23–30 August 1939; Volume 7, pp. 191–196. [Google Scholar]
- Harrington, J. Pioneer Hi-Bred, a DuPont business bringing seed value to the grower. In Principles of Plant Genetics and Breeding, 2nd ed.; Acquaah, G., Ed.; Wiley Blackwell: Chichester, UK, 2012; pp. 357–360. [Google Scholar]
- Parisseaux, B.; Bernardo, R. In silico mapping of quantitative trait loci in maize. Theor. Appl. Genet. 2004, 109, 508–514. [Google Scholar] [CrossRef]
- Duvick, D.N. Heterosis: Feeding people and protecting natural resources. In The Genetics and Exploitation of Heterosis in Crops; Coors, J.G., Pandey, S., Eds.; ASA: Madison, WI, USA, 1999; pp. 19–29. [Google Scholar]
- Nagaru, U. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Bateman, A.J. Self-incompatibility systems in Angiosperms: III. Cruciferae. Heredity 1955, 9, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.A.; Thurling, N. The genetics of self-incompatibility in Brassica campestris L. ssp. oleifera Metzg. I. Characteristics of S-locus. Control of self-incompatibility. Genetica 1973, 44, 428–438. [Google Scholar] [CrossRef]
- Mackay, G.R. A diallel cross method for the recognition of S-allele homozygotes in turnip, Brassica campestris L. ssp. rapifera. Heredity 1977, 38, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.G. Inbreeding and the production of commercial F1 hybrid seed of Brussels sprouts. Euphytica 1966, 15, 68–79. [Google Scholar] [CrossRef]
- Dixon, G.R. Vegetable Brassicas and Related Crucifers, Crop Production Science in Horticulture 14; CABI: Wallingford, UK, 2007. [Google Scholar]
- Werner, C.P.; Smith, B.M.; Godwin, M.; Kearsey, M.J. The feasibility of producing inbred rather than F1 hybrid cultivars in Brussels sprouts (Brassica oleracea var. gemmifera): An assessment of recombinant inbred lines. Ann. Appl. Biol. 1991, 119, 331–337. [Google Scholar] [CrossRef]
- Bradshaw, J.E.; Titley, M.; Wilson, R.N. Single seed descent as a breeding method for swedes (Brassica napus L. var. napobrassica Peterm). Euphytica 2009, 169, 387–401. [Google Scholar] [CrossRef]
- Ramsay, L.D.; Bradshaw, J.E.; Kearsey, M.J. The inheritance of quantitative traits in swedes (Brassica napus L. spp. rapifera). Diallel analysis of dry matter yield. J. Genet. Breed. 1994, 49, 253–258. [Google Scholar]
- Gemmell, D.J.; Bradshaw, J.E.; Hodgkin, T.; Gowers, S. Self-incompatibility in Brassica napus: Seed set on crossing 19 self-incompatible lines. Euphytica 1989, 42, 71–77. [Google Scholar] [CrossRef]
- Gowers, S. The production of F1 hybrid swedes (Brassica napus ssp. rapifera) by the utilisation of self-incompatibility. Euphytica 1974, 23, 205–208. [Google Scholar] [CrossRef]
- Gowers, S. Methods of producing F1 hybrid swedes (Brassica napus ssp. rapifera). Euphytica 1975, 24, 537–541. [Google Scholar] [CrossRef]
- Gowers, S. Swedes and turnips. In Root and Tuber Crops, Handbook of Plant Breeding 7; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 245–289. [Google Scholar]
- Bradshaw, J.E.; Wilson, R.N. Inbred line versus F1 hybrid breeding in swedes (Brassica napus L. var. napobrassica Peterm). Ann. Appl. Biol. 1993, 123, 657–665. [Google Scholar] [CrossRef]
- Lewis, D. The physiology of incompatibility in plants. III. Autopolyploids. J. Genet. 1943, 44, 171–185. [Google Scholar] [CrossRef]
- Lewis, D. Comparative incompatibility in angiosperms and fungi. Adv. Genet. 1954, 6, 235–285. [Google Scholar] [PubMed]
- Glendinning, D.R. Neo-Tuberosum: New potato breeding material. 4. The breeding system of Neo-Tuberosum, and the structure and composition of the Neo-Tuberosum gene pool. Potato Res. 1976, 19, 27–36. [Google Scholar] [CrossRef]
- Brown, C.R. Outcrossing rate in cultivated autotetraploid potato. Am. Potato J. 1993, 70, 725–734. [Google Scholar] [CrossRef]
- Kempthorne, O. An Introduction to Genetic Statistics; Wiley: New York, NY, USA, 1957. [Google Scholar]
- Bradshaw, J.E. Quantitative genetics theory for tetrasomic inheritance. In Potato Genetics; Bradshaw, J.E., Mackay, G.R., Eds.; CAB International: Wallingford, UK, 1994; pp. 71–99. [Google Scholar]
- Hospital, F. Marker-assisted breeding. In Plant Molecular Breeding; Newbury, H.J., Ed.; Blackwell: Oxford, UK, 2003; pp. 30–59. [Google Scholar]
- Bethke, P.C.; Halterman, D.A.; Jansky, S.H. Potato germplasm enhancement enters the genomics era. Agronomy 2019, 9, 575. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Viquez-Zamora, M.; den Uil, D.; Sinnige, J.; Kruyt, H.; Vossen, J.; Lindhout, P.; van Heusden, S. Introgression of genes for resistance against Phytophthora infestans in diploid potato. Am. J. Potato Res. 2020, 97, 33–42. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Vossen, J.H.; Visser, R.G.F. Durable late blight resistance in potato through dynamic varieties obtained by cisgenesis: Scientific and societal advances in the DuRPh project. Potato Res. 2016, 59, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Foster, S.J.; Park, T.-H.; Pel, M.A.; Brigneti, G.; Sliwka, J.; Jagger, L.; van der Vossen, E.; Jones, J.D.G. Rpi-vnt1.1, a Tm-22 homolog from Solanum venturii, confers resistance to potato late blight. Mol. Plant-Microbe Interact. 2009, 22, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Pel, M.A.; Foster, S.J.; Park, T.H.; Rietman, H.; van Arkel, G.; Jones, J.D.G.; Van Eck, H.J.; Jacobsen, E.; Visser, R.G.F.; Van der Vossen, E.A.G. Mapping and cloning of late blight resistance genes from Solanum venturii using an interspecific candidate gene approach. Mol. Plant-Microbe Interact. 2009, 22, 601–615. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Zhang, Y.; Zhu, Z.; Chen, T.; Zhao, Q.; Yao, S.; Zhao, L.; Zhu, W.; Wang, C. Substitutional mapping the cooked rice elongation by using chromosome segment substitution lines in rice. Mol. Plant Breed. 2013, 4, 107–115. [Google Scholar] [CrossRef]
- Hofvander, P.; Andreasson, E.; Anderson, M. Potato trait development going fast-forward with genome editing. Trends Genet. 2022, 38, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Acquaah, G. Principles of Plant Genetics and Breeding, 3rd ed.; Wiley Blackwell: Chichester, UK, 2021. [Google Scholar]
- Hallauer, A.R.; Carena, M.J. Maize Breeding. In Cereals, Handbook of Plant Breeding 3; Carena, M.J., Ed.; Springer: New York, NY, USA, 2009; pp. 3–98. [Google Scholar]
- Hallauer, A.R.; Carena, M.J.; Miranda, F.J.B. Quantitative Genetics in Maize Breeding, 3rd ed.; Handbook of Plant Breeding 6; Springer: New York, NY, USA, 2010. [Google Scholar]
- van Dijk, L.C.M.; Lommen, W.J.M.; de Vries, M.E.; Kacheyo, O.C.; Struik, P.C. Hilling of transplanted seedlings from novel hybrid true potato seeds does not enhance tuber yield but can affect tuber size distribution. Potato Res. 2021, 64, 353–374. [Google Scholar] [CrossRef]
- Bai, Y.; Lindhout, P. Domestication and breeding of tomatoes: What have we gained and what can we gain in the future? Ann. Bot. 2007, 100, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Horneburg, B.; Myers, J.R. Tomato: Breeding for improved disease resistance in fresh market and home garden varieties. In Organic Crop Breeding; Lammerts van Bueren, E.T., Myers, J.R., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 239–249. [Google Scholar]
- Díez, M.J.; Nuez, F. Tomato. In Vegetables II, Handbook of Plant Breeding 2; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 249–323. [Google Scholar]
- Craig, W.F. Production of Hybrid Seed Corn. In Corn and Corn Improvement, Agronomy 18; Sprague, G.F., Ed.; ASA, Inc.: Madison, WI, USA, 1977; pp. 671–719. [Google Scholar]
- Tatum, L.A. The southern corn leaf blight epidemic. Science 1971, 171, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Ullstrup, A.J. The impacts of the southern corn leaf blight epidemic of 1970–1971. Ann. Rev. Phytopathol. 1972, 10, 37–50. [Google Scholar] [CrossRef]
- Jones, J.N.; Clarke, A.E. Inheritance of Male Sterility in the Onion and the Production of Hybrid Seed. Proc. Am. Soc. Hortic. Sci. 1943, 43, 189–194. [Google Scholar]
- Anisimova, I.N.; Gavrilenko, T.A. Cytoplasmic male sterility and prospects for its utilization in potato breeding, genetic studies and hybrid seed production. Russ. J. Genet. Appl. Res. 2017, 7, 721–735. [Google Scholar] [CrossRef]
- van Heerwaarden, J.; Hufford, M.B.; Ross-Ibarra, J. Historical genomics of North American maize. Proc. Natl. Acad. Sci. USA 2012, 109, 12420–12425. [Google Scholar] [CrossRef] [Green Version]
- Troyer, A.F. Champaign County, Illinois, and the Origin of Hybrid Corn. Plant Breed. Rev. 2004, 24, 41–59. [Google Scholar]
- Stockem. J.; de Vries, M.; van Nieuwenhuizen, E.; Lindhout, P.; Struik, P.C. Contribution and stability of yield components of diploid hybrid potatoes. Potato Res. 2020, 63, 345–366. [Google Scholar] [CrossRef]
- De Vries, M.; ter Maat, M.; Lindhout, P. The potential of hybrid potato for East-Africa. Open Agric. 2016, 1, 151–156. [Google Scholar] [CrossRef]
- Beumer, K.; Stemerding, D.; Swart, J.A.A. Innovation and the commons: Lessons from the governance of genetic resources in potato breeding. Agric. Hum. Values 2021, 38, 525–539. [Google Scholar] [CrossRef]
- Beumer, K.; Stemerding, D. A breeding consortium to realize the potential of hybrid diploid potato for food security. Nat. Plants 2021, 7, 1530–1532. [Google Scholar] [CrossRef] [PubMed]
- Forbes, G.A.; Charkowski, A.; Andrade-Piedra, J.; Parker, M.L.; Schulte-Geldermann, E. Potato seed systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 431–447. [Google Scholar]
- Priegnitz, U.; Lommen, W.J.M.; van der Vlugt, R.A.A.; Struik, P.C. Impact of positive selection on incidence of different viruses during multiple generations of potato seed tubers in Uganda. Potato Res. 2019, 62, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Priegnitz, U.; Lommen, W.J.M.; van der Vlugt, R.A.A.; Struik, P.C. Potato yield and yield components as affected by positive selection during several generations of seed multiplication in southwestern Uganda. Potato Res. 2020, 63, 507–543. [Google Scholar] [CrossRef]
- Jin, L. Potato production and breeding in China. In Achieving Sustainable Cultivation of Potatoes, Volume 1: Breeding Improved Varieties; Wang-Pruski, G., Ed.; Burleigh Dodds: Cambridge, UK, 2018; pp. 259–270. [Google Scholar]
- Singh, R.K.; Buckseth, T.; Sharma, A.K.; Tiwari, J.K.; Chakrabarti, S.K. Potato seed certification in India. Indian Hortic. 2019, 64, 54–58. [Google Scholar]
- Buckseth, T.; Singh, R.K.; Sharma, A.K.; Tiwari, J.K.; Chakrabarti, S.K. Hi-tech seed potato production system. Indian Hortic. 2019, 64, 18–21. [Google Scholar]
- Kreuze, J.F.; Souza-Dias, J.A.C.; Jeevalatha, A.; Figueira, A.R.; Valkonen, J.P.T.; Jones, R.A.C. Viral diseases in potato. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 389–430. [Google Scholar]
- Parker, M.L.; Reddy, R. Apical cuttings to boost potato seed systems. In Global Potato Conclave 2020; Sharma, S., Tiwari, J.K., Kumar, R., Kardile, H.B., Bhardwaj, V., Chakrabarti, S.K., Dua, V.K., Eds.; Malhotra Publishing House: New Delhi, India, 2020; pp. 276–278. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation (n) | Mean Number of Heterozygous Tracts (m) | Proportion of Lines Homozygous at All Loci |
|---|---|---|
| 0 | 12.000000 | 0 |
| 1 | 16.000000 | very small |
| 2 | 13.000000 | very small |
| 3 | 9.000000 | small |
| 4 | 5.750000 | small |
| 5 | 3.500000 | 0.0302 |
| 6 | 2.062500 | 0.1271 |
| 7 | 1.187500 | 0.3050 |
| 8 | 0.671875 | 0.5108 |
| 9 | 0.375000 | 0.6873 |
| 10 | 0.207031 | 0.8130 |
| 11 | 0.113281 | 0.8929 |
| 12 | 0.061523 | 0.9403 |
| 13 | 0.033203 | 0.9673 |
| 14 | 0.017822 | 0.9823 |
| 15 | 0.009521 | 0.9905 |
| 16 | 0.005066 | 0.9949 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradshaw, J.E. Breeding Diploid F1 Hybrid Potatoes for Propagation from Botanical Seed (TPS): Comparisons with Theory and Other Crops. Plants 2022, 11, 1121. https://doi.org/10.3390/plants11091121
Bradshaw JE. Breeding Diploid F1 Hybrid Potatoes for Propagation from Botanical Seed (TPS): Comparisons with Theory and Other Crops. Plants. 2022; 11(9):1121. https://doi.org/10.3390/plants11091121
Chicago/Turabian StyleBradshaw, John E. 2022. "Breeding Diploid F1 Hybrid Potatoes for Propagation from Botanical Seed (TPS): Comparisons with Theory and Other Crops" Plants 11, no. 9: 1121. https://doi.org/10.3390/plants11091121