
The Delay of Raphanus raphanistrum subsp. sativus (L.) Domin Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism



Abstract
:1. Introduction
2. Results
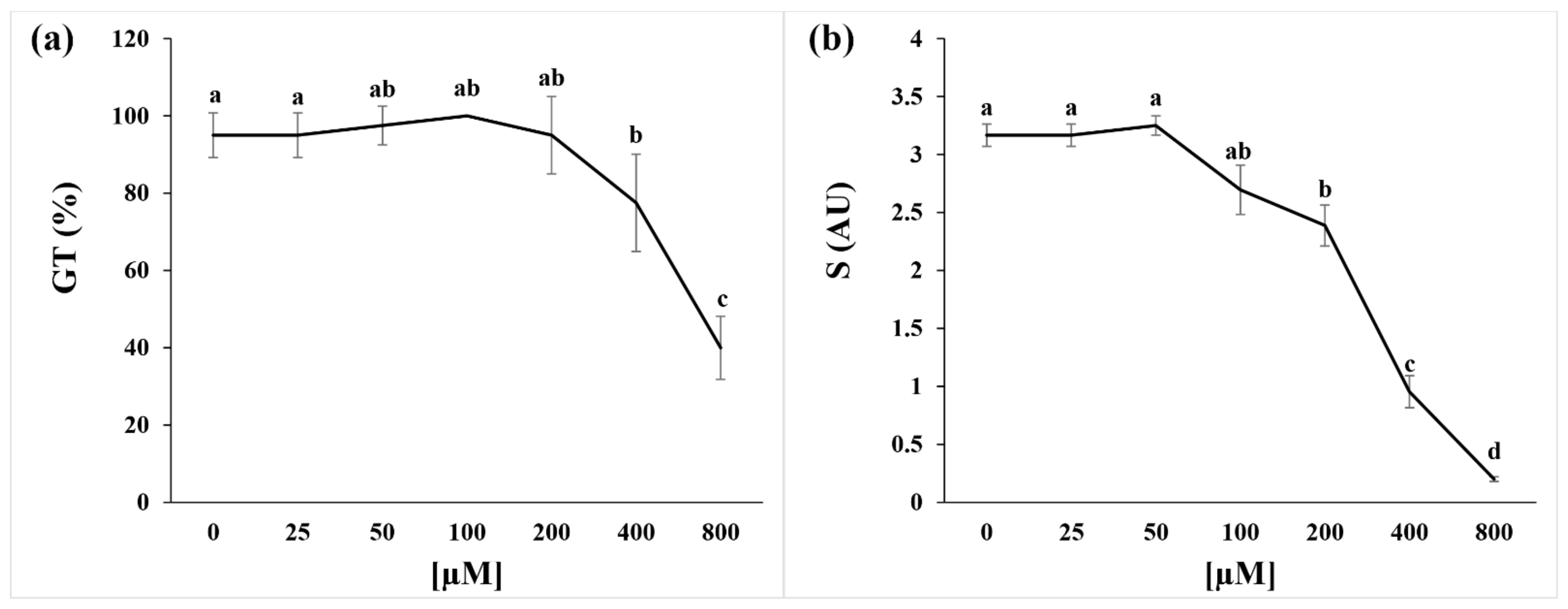
2.1. Germination Index and Seed Respiration
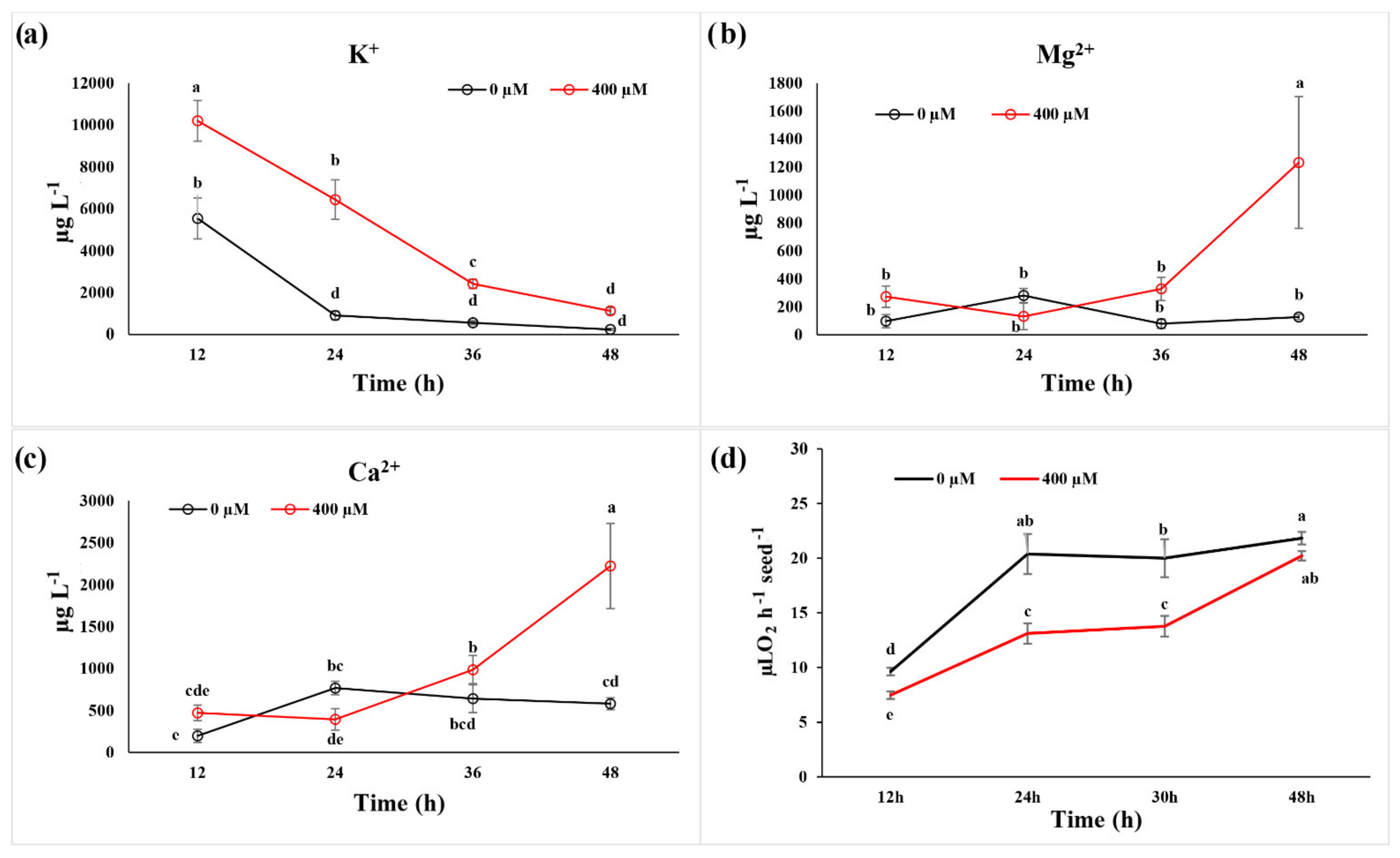
2.2. Ion Leakage and Reabsorption and Seed Respiration
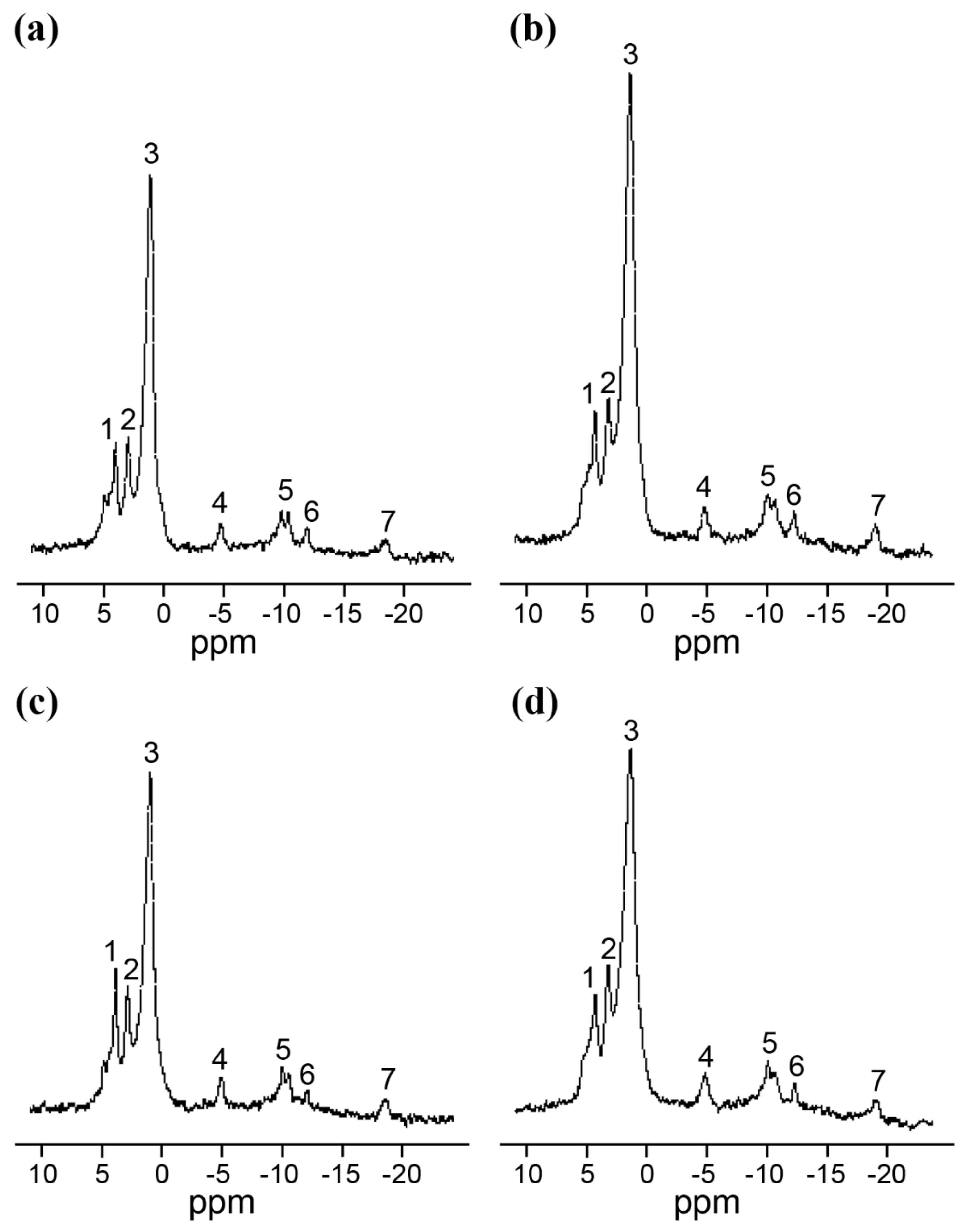
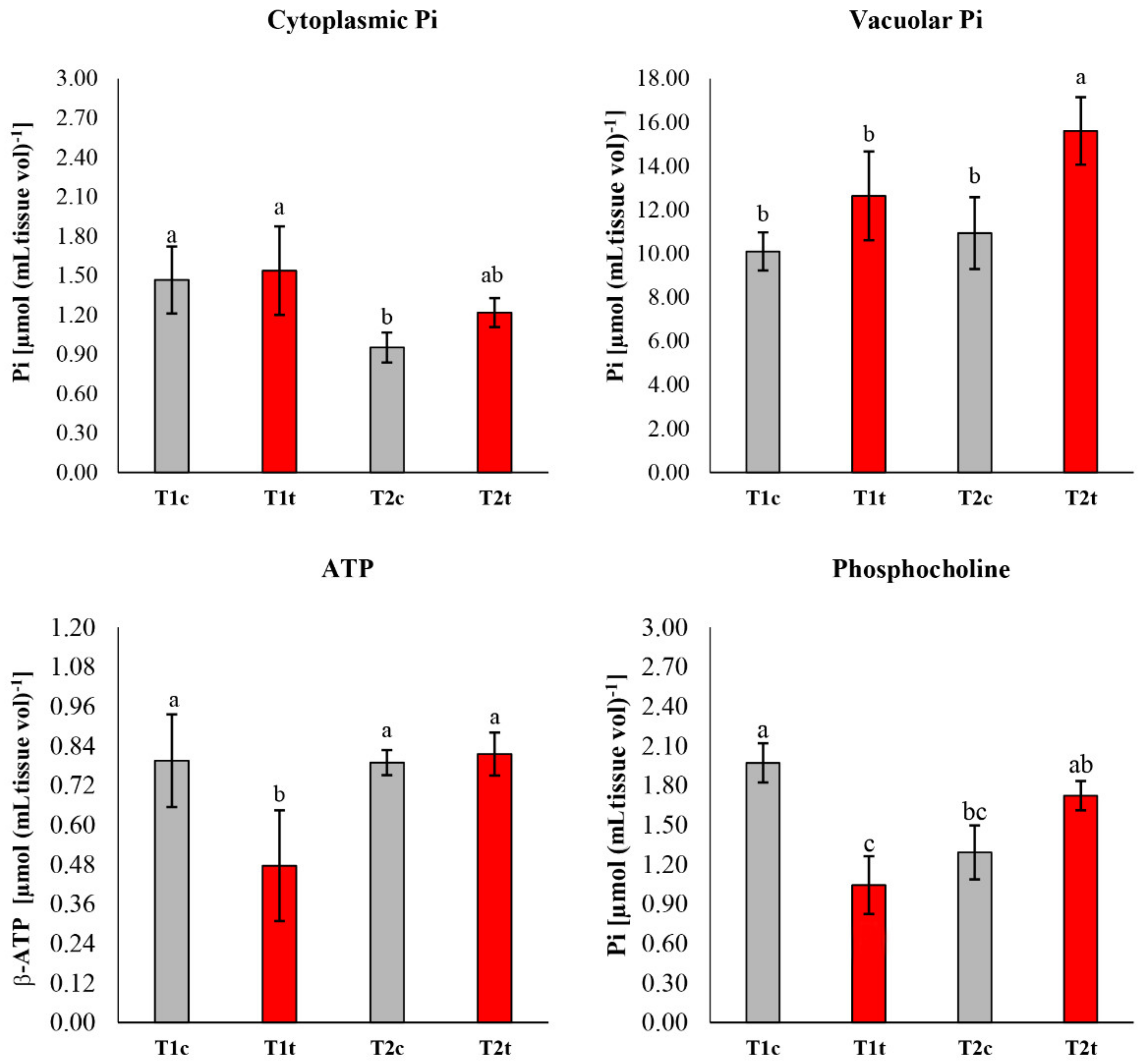
2.3. In Vivo NMR Analysis
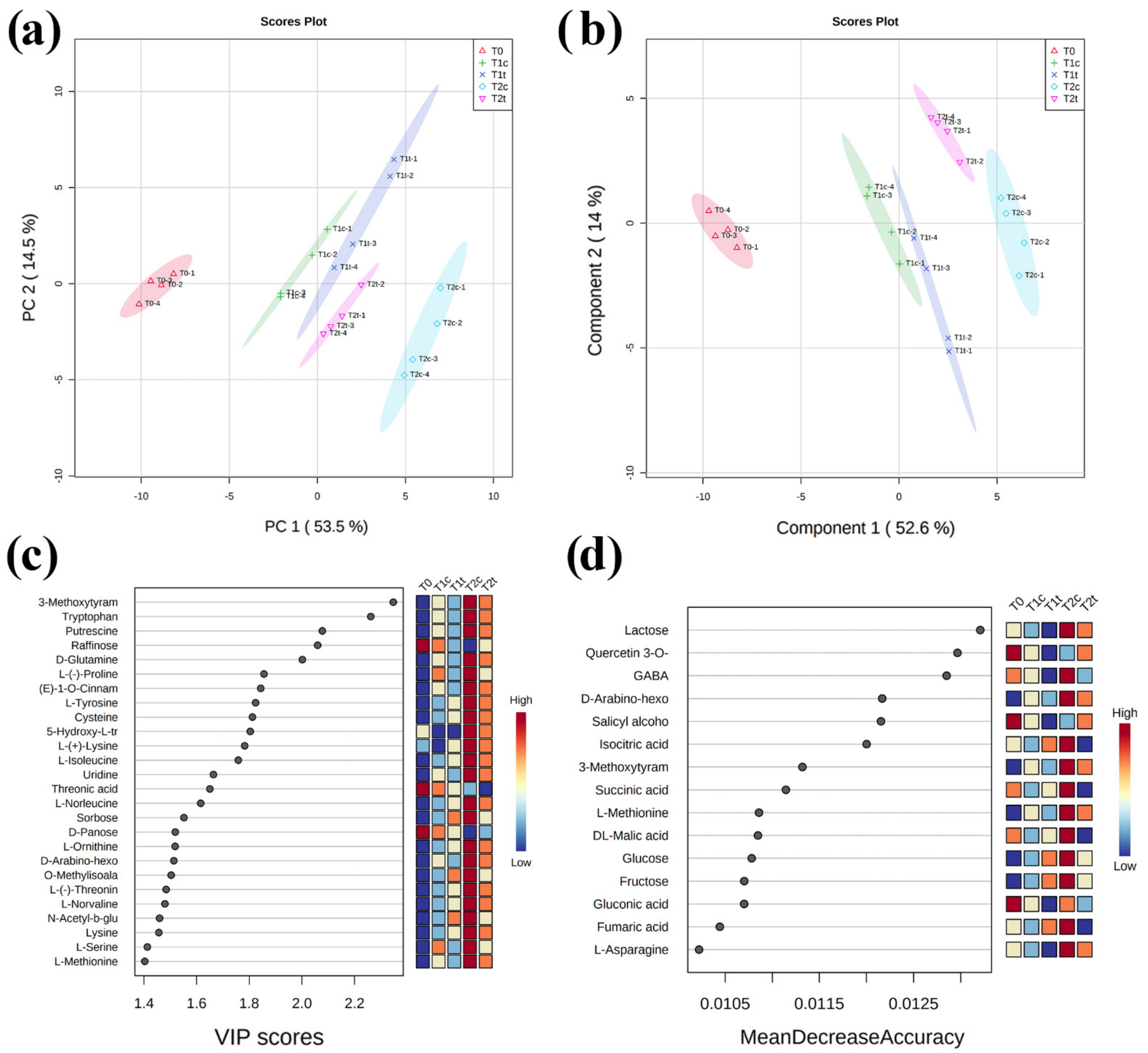
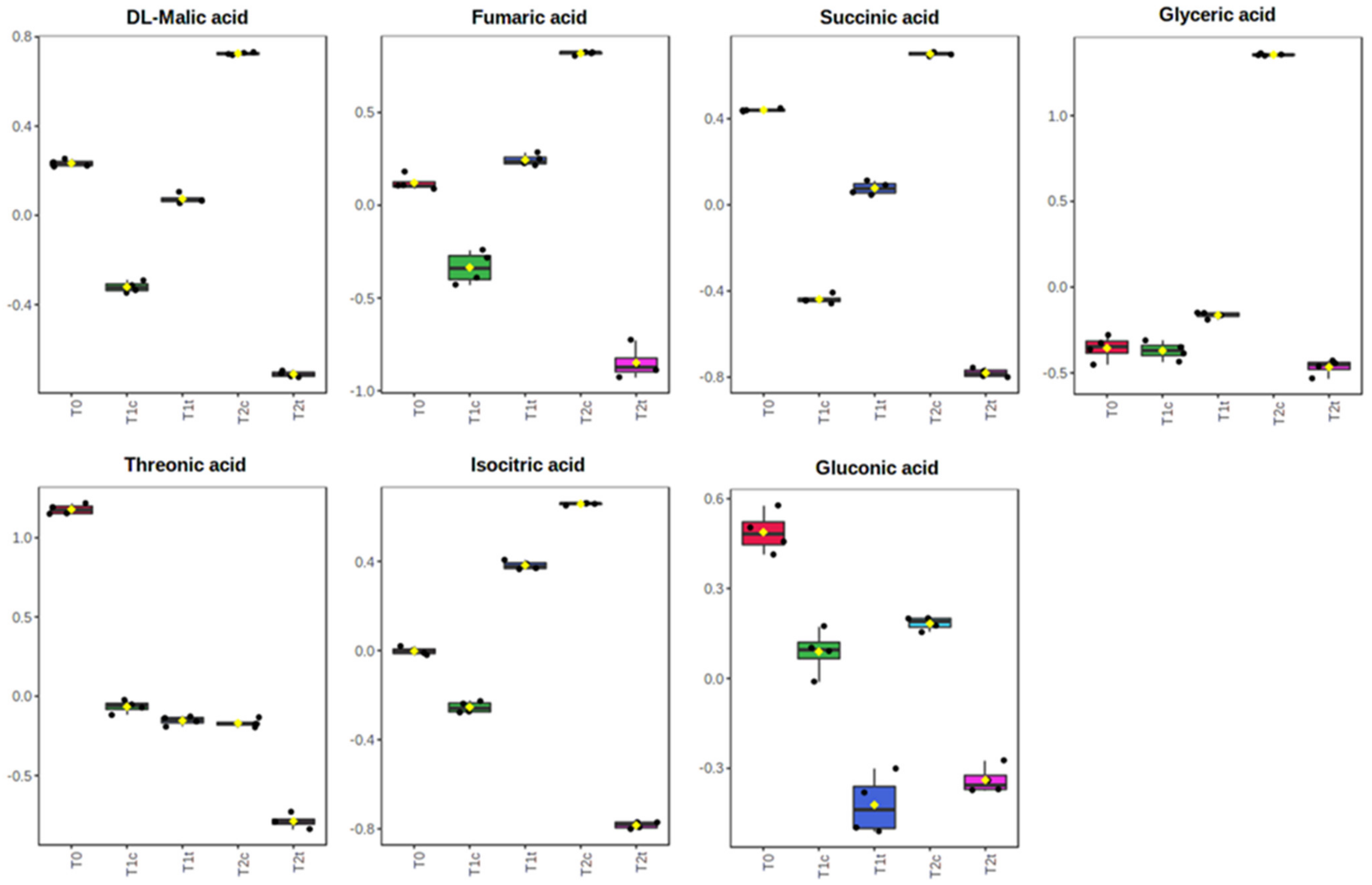
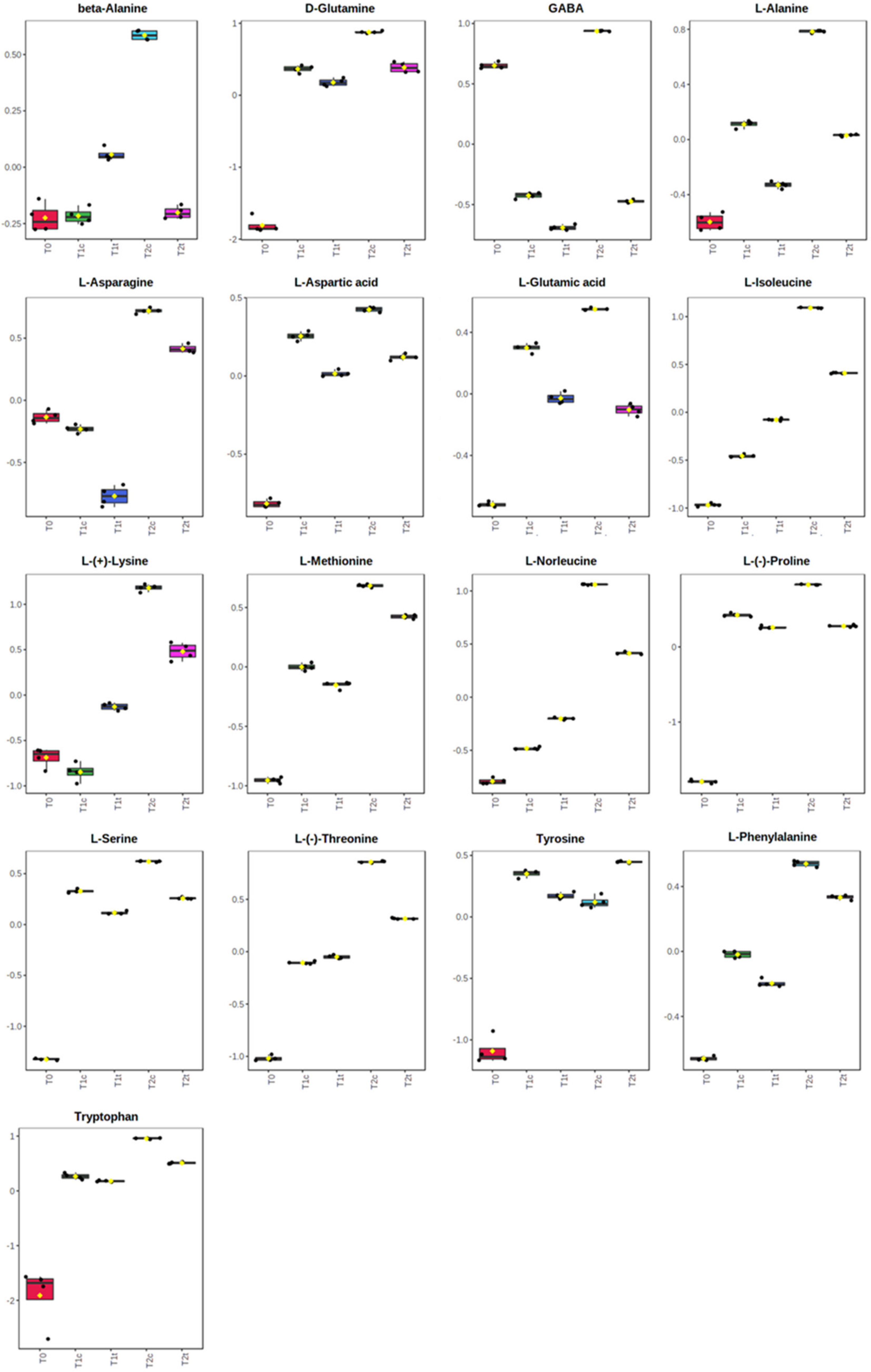
2.4. GC-MS Untargeted Metabolomic
3. Discussion
4. Materials and Methods
4.1. Plant Material, Treatment, and Germination Indexes
4.2. Ion Concentration in the Incubation Medium
4.3. Oxygen-Uptake Rate Evaluation
4.4. Nuclear Magnetic Resonance Spectroscopy
4.5. Untargeted Metabolomic Analysis
4.6. GC-Quadrupole/MS Analysis
4.7. GC/MS Data Analysis Using MS-DIAL
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duke, S.O.; Pan, Z.; Bajsa-Hirschel, J. Proving the mode of action of phytotoxic phytochemicals. Plants 2020, 9, 1756. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirumurugan, D.; Cholarajan, A.; Raja, S.; Vijayakumar, R. An introductory chapter: Secondary metabolites. Second. Metab. Sources Appl. 2018, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.; Owens, D.; Dayan, F. Natural product-based chemical herbicides. In Weed Control; CRC Press: Boca Raton, FL, USA, 2018; pp. 153–165. [Google Scholar]
- Reigosa, M.; Pazos-Malvido, E. Phytotoxic effects of 21 plant secondary metabolites on Arabidopsis thaliana germination and root growth. J. Chem. Ecol. 2007, 33, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Pergo, É.M.; Abrahim, D.; Soares da Silva, P.C.; Kern, K.A.; Da Silva, L.J.; Voll, E.; Ishii-Iwamoto, E.L. Bidens pilosa L. exhibits high sensitivity to coumarin in comparison with three other weed species. J. Chem. Ecol. 2008, 34, 499–507. [Google Scholar] [CrossRef]
- Chen, B.-X.; Peng, Y.-X.; Yang, X.-Q.; Liu, J. Delayed germination of Brassica parachinensis seeds by coumarin involves decreased GA4 production and a consequent reduction of ROS accumulation. Seed Sci. Res. 2021, 31, 224–235. [Google Scholar] [CrossRef]
- Chen, B.-X.; Peng, Y.-X.; Gao, J.-D.; Zhang, Q.; Liu, Q.-J.; Fu, H.; Liu, J. Coumarin-induced delay of rice seed germination is mediated by suppression of abscisic acid catabolism and reactive oxygen species production. Front. Plant Sci. 2019, 10, 828. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, M.; Cacco, G.; Sorgona, A.; Marabottini, R.; Paolacci, A.; Ciaffi, M.; Badiani, M. The inhibitory effects of coumarin on the germination of durum wheat (Triticum turgidum ssp. durum, cv. Simeto) seeds. J. Chem. Ecol. 2006, 32, 489–506. [Google Scholar] [CrossRef]
- Aliotta, G.; Cafiero, G. Seed bioassay and microscopy in the study of allelopathy: Radish and purslane responses. In Handbook of Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2001; pp. 1–20. [Google Scholar]
- Aliotta, G.; Cafiero, G.; Fiorentino, A.; Strumia, S. Inhibition of radish germination and root growth by coumarin and phenylpropanoids. J. Chem. Ecol. 1993, 19, 175–183. [Google Scholar] [CrossRef]
- Aliotta, G.; Fuggi, A.; Strumia, S. Coat-imposed dormancy by coumarin in radish seeds: The influence of light. Plant Biosyst. 1992, 126, 631–637. [Google Scholar] [CrossRef]
- Bruno, L.; Talarico, E.; Cabeiras-Freijanes, L.; Madeo, M.L.; Muto, A.; Minervino, M.; Lucini, L.; Miras-Moreno, B.; Sofo, A.; Araniti, F. Coumarin interferes with polar auxin transport altering microtubule cortical array organization in Arabidopsis thaliana (L.) Heynh. root apical meristem. Int. J. Mol. Sci. 2021, 22, 7305. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Scognamiglio, M.; Chambery, A.; Russo, R.; Esposito, A.; D’Abrosca, B.; Fiorentino, A.; Lupini, A.; Sunseri, F.; Abenavoli, M.R. Highlighting the effects of coumarin on adult plants of Arabidopsis thaliana (L.) Heynh. by an integrated-omic approach. J. Plant Physiol. 2017, 213, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Mancuso, R.; Lupini, A.; Giofrè, S.V.; Sunseri, F.; Gabriele, B.; Abenavoli, M.R. Phytotoxic potential and biological activity of three synthetic coumarin derivatives as new natural-like herbicides. Molecules 2015, 20, 17883–17902. [Google Scholar] [CrossRef] [Green Version]
- McCalla, T.; Norstard, F.A. Toxicity problems in mulch tillage. Agric. Environ. 1974, 1, 153–174. [Google Scholar] [CrossRef]
- McCalla, T. Studies on phytotoxic substances from soil microorganisms and crop residues at Lincoln, Nebraska. In Biochemical Interactions among Plants; National Academy of Sciences: Washington, DC, USA, 1971; pp. 39–43. [Google Scholar]
- Dornbos, D.L., Jr.; Spencer, G.F.; Miller, R.W. Medicarpin delays alfalfa seed germination and seedling growth. Crop Sci. 1990, 30, 162–166. [Google Scholar] [CrossRef]
- Schnute, M.E. The allelopathic aspects of Melilotus alba through coumarin. J. Wash. Acad. Sci. 1984, 74, 117–120. [Google Scholar]
- Nutile, G. Inducing dormancy in lettuce seed with coumarin. Plant Physiol. 1945, 20, 433. [Google Scholar] [CrossRef]
- Mayer, A.; Evenari, M. The relation between the structure of coumarin and its derivatives, and their activity as germination inhibitors. J. Exp. Bot. 1952, 3, 246–252. [Google Scholar] [CrossRef]
- Mayer, A.; Poljakoff-Mayber, A. Coumarins and their role in growth and germination. In Plant Growth Regulation; Klein, R.M., Ed.; Iowa State University Press: Ames, IA, USA, 1961; pp. 735–750. [Google Scholar]
- Khan, A.; Tolbert, N. Light-controlled cycocel reversal of coumarin inhibition of lettuce seed germination and root growth. Physiol. Plant. 1966, 19, 76–80. [Google Scholar] [CrossRef]
- Berrie, A.; Parker, W.; Knights, B.; Hendrie, M. Studies on lettuce seed germination-I. Coumarin induced dormancy. Phytochemistry 1968, 7, 567–573. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreiras, A.M.; Graña, E.; Díaz-Tielas, C.; López-González, D.; Araniti, F.; Celeiro, M.; Teijeira, M.; Verdeguer, M.; Reigosa, M.J. Elucidating the phytotoxic potential of natural compounds. In Advances in Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2018; pp. 363–378. [Google Scholar]
- Williams, R.; Bartholomew, P. Radish (Raphanus sativus) seed size affects germination response to coumarin. Allelopath. J. 2012, 30, 271–279. [Google Scholar]
- Bernhard, R.A. Some studies of coumarin and coumarin analogues as germination inhibitors of radish seeds. Bot. Gaz. 1959, 121, 17–21. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef] [Green Version]
- Cocucci, S.; Cocucci, M. Effect of ABA, GA3 and FC on the development of potassium uptake in germinating radish seeds. Plant Sci. Lett. 1977, 10, 85–95. [Google Scholar] [CrossRef]
- Espen, L.; Pirovano, L.; Cocucci, S.M. Effects of Ni2+ during the early phases of radish (Raphanus sativus) seed germination. Environ. Exp. Bot. 1997, 38, 187–197. [Google Scholar] [CrossRef]
- Li, M.; Chen, X.; He, D.; Yang, P. Proteomic analysis reveals that calcium channel blockers affect radicle protrusion during rice seed germination. Plant Growth Regul. 2020, 90, 393–407. [Google Scholar] [CrossRef]
- Negrini, N.; Rivetta, A.; Cocucci, M. Calmodulin levels in radish (Raphanus sativus L.) seeds germinating at low calcium availability induced by EGTA treatments. Plant Cell Environ. 1995, 18, 159–167. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Araguirang, G.E.; Ngo, A.H.; Lin, K.-T.; Angkawijaya, A.E.; Nakamura, Y. The four Arabidopsis choline/ethanolamine kinase isozymes play distinct roles in metabolism and development. Plant Physiol. 2020, 183, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Lord, J.; Kagawa, T.; Beevers, H. Intracellular distribution of enzymes of the cytidine diphosphate choline pathway in castor bean endosperm. Proc. Natl. Acad. Sci. USA 1972, 69, 2429–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craddock, C.P.; Adams, N.; Bryant, F.M.; Kurup, S.; Eastmond, P.J. Phosphatidic acid phosphohydrolase regulates phosphatidylcholine biosynthesis in Arabidopsis by phosphatidic acid-mediated activation of CTP:Phosphocholine cytidylyltransferase activity. Plant Cell 2015, 27, 1251–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupidlowska, E.; Dobrzynska, K.; Parys, E.; Zobel, A.M. Effect of coumarin and xanthotoxin on mitochondrial structure, oxygen uptake, and succinate dehydrogenase activity in onion root cells. J. Chem. Ecol. 1994, 20, 2471–2480. [Google Scholar] [CrossRef] [PubMed]
- Cloetens, P.; Mache, R.; Schlenker, M.; Lerbs-Mache, S. Quantitative phase tomography of Arabidopsis seeds reveals intercellular void network. Proc. Natl. Acad. Sci. USA 2006, 103, 14626–14630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botha, F.; Potgieter, G.; Botha, A.-M. Respiratory metabolism and gene expression during seed germination. Plant Growth Regul. 1992, 11, 211–224. [Google Scholar] [CrossRef]
- Spoelstra, P.; Joosen, R.V.; Van der Plas, L.H.; Hilhorst, H.W. The distribution of ATP within tomato (Lycopersicon esculentum Mill.) embryos correlates with germination whereas total ATP concentration does not. Seed Sci. Res. 2002, 12, 231–238. [Google Scholar] [CrossRef]
- Eastmond, P.J.; Graham, I.A. Re-examining the role of the glyoxylate cycle in oilseeds. Trends Plant Sci. 2001, 6, 72–78. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- Onomo, P.E.; Niemenak, N.; Ndoumou, D.O.; Lieberei, R. Change in amino acids content during germination and seedling growth of Cola sp. Afr. J. Biotechnol. 2010, 9, 5632–5642. [Google Scholar]
- Fait, A.; Angelovici, R.; Less, H.; Ohad, I.; Urbanczyk-Wochniak, E.; Fernie, A.R.; Galili, G. Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 2006, 142, 839–854. [Google Scholar] [CrossRef] [Green Version]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Kirma, M.; Araújo, W.L.; Fernie, A.R.; Galili, G. The multifaceted role of aspartate-family amino acids in plant metabolism. J. Exp. Bot. 2012, 63, 4995–5001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Galili, G. Increased lysine synthesis coupled with a knockout of its catabolism synergistically boosts lysine content and also transregulates the metabolism of other amino acids in Arabidopsis seeds. Plant Cell 2003, 15, 845–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.K.; Ray, P.M.; Wade-Jardetzky, N.; Jardetzky, O. Estimation of cytoplasmic and vacuolar pH in higher plant cells by 31P NMR. Nature 1980, 283, 870–872. [Google Scholar] [CrossRef]
- Kime, M.; Ratcliffe, R.; Loughman, B. The Application of 31P Nuclear Magnetic Resonance to Higher Plant Tissue: II. Detection of intracellular changes. J. Exp. Bot. 1982, 33, 670–681. [Google Scholar] [CrossRef]
- Spickett, C.M.; Smirnoff, N.; Ratcliffe, R.G. Metabolic response of maize roots to hyperosmotic shock: An in vivo 31P nuclear magnetic resonance study. Plant Physiol. 1992, 99, 856–863. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.K.; Wade-Jardetzky, N.; Jardetsky, O. Intracellular pH measurements by phosphorus-31 nuclear magnetic resonance. Influence of factors other than pH on phosphorus-31 chemical shifts. Biochemistry 1981, 20, 5389–5394. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M. GMD@ CSB. DB: The Golm metabolome database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Sansone, S.-A.; Fan, T.; Goodacre, R.; Griffin, J.L.; Hardy, N.W.; Kaddurah-Daouk, R.; Kristal, B.S.; Lindon, J.; Mendes, P.; Morrison, N. The metabolomics standards initiative. Nat. Biotechnol. 2007, 25, 846–849. [Google Scholar]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Time (h) | Cytoplasmic pH | Vacuolar pH | ||
|---|---|---|---|---|
| Control | Coumarin | Control | Coumarin | |
| 24 | 7.74 ± 0.07 a | 7.77 ± 0.04 a | 5.92 ± 0.11 a | 6.08 ± 0.04 a |
| 48 | 7.63 ± 0.04 a | 7.57 ± 0.03 a | 5.73 ± 0.13 b | 5.97 ± 0.07 a |
| T1 | T2 | ||||||
|---|---|---|---|---|---|---|---|
| Total Cmpd | Hits | Raw p | FDR | Raw p | FDR | Impact | |
| Linoleic acid metabolism | 4 | 1 | 2.60 × 10−8 | 1.34 × 10−7 | 9.34 × 10−9 | 1.81 × 10−8 | 1 |
| Alanine aspartate and glutamate metabolism | 22 | 7 | 3.92 × 10−9 | 4.05 × 10−8 | 2.78 × 10−10 | 1.01 × 10−9 | 0.58274 |
| Isoquinoline alkaloid biosynthesis | 6 | 1 | 8.49 × 10−5 | 0.00011 | 4.46 × 10−9 | 1.04 × 10−8 | 0.5 |
| Phenylalanine metabolism | 11 | 1 | 3.24 × 10−5 | 4.28 × 10−5 | 1.99 × 10−6 | 2.75 × 10−6 | 0.47059 |
| Arginine biosynthesis | 18 | 6 | 2.95 × 10−6 | 5.22 × 10−6 | 6.90 × 10−9 | 1.53 × 10−8 | 0.39224 |
| Flavone and flavonol biosynthesis | 10 | 1 | 0.000517 | 0.000616 | 1.45 × 10−6 | 2.05 × 10−6 | 0.35 |
| beta-Alanine metabolism | 18 | 3 | 3.35 × 10−11 | 1.04 × 10−9 | 4.96 × 10−10 | 1.62 × 10−9 | 0.3254 |
| Glycine serine and threonine metabolism | 33 | 5 | 3.18 × 10−6 | 5.47 × 10−6 | 1.21 × 10−10 | 4.99 × 10−10 | 0.3242 |
| Arginine and proline metabolism | 34 | 6 | 2.15 × 10−6 | 3.92 × 10−6 | 1.80 × 10−10 | 6.98 × 10−10 | 0.31071 |
| Galactose metabolism | 27 | 7 | 2.05 × 10−5 | 2.77 × 10−5 | 1.06 × 10−10 | 4.69 × 10−10 | 0.28841 |
| Glyo×ylate and dicarbo×ylate metabolism | 29 | 5 | 1.97 × 10−9 | 2.44 × 10−8 | 1.82 × 10−12 | 2.26 × 10−11 | 0.21995 |
| Tyrosine metabolism | 16 | 3 | 3.22 × 10−8 | 1.43 × 10−7 | 1.81 × 10−8 | 3.12 × 10−8 | 0.21622 |
| Cysteine and methionine metabolism | 46 | 5 | 2.83 × 10−11 | 1.04 × 10−9 | 2.93 × 10−9 | 7.25 × 10−9 | 0.19893 |
| Starch and sucrose metabolism | 22 | 2 | 1.24 × 10−6 | 2.32 × 10−6 | 3.82 × 10−8 | 5.90 × 10−8 | 0.17067 |
| Sulphur metabolism | 15 | 5 | 1.86 × 10−5 | 2.56 × 10−5 | 3.90 × 10−8 | 5.90 × 10−8 | 0.16851 |
| Butanoate metabolism | 17 | 3 | 1.13 × 10−8 | 8.53 × 10−8 | 6.70 × 10−13 | 1.64 × 10−11 | 0.13636 |
| Stilbenoid diarylheptanoid and gingerol biosynthesis | 8 | 1 | // | // | 0.024158 | 0.024554 | 0.13235 |
| Phenylpropanoid biosynthesis | 46 | 4 | // | // | 0.00847 | 0.009054 | 0.13199 |
| Flavonoid biosynthesis | 47 | 3 | // | // | 0.004188 | 0.004556 | 0.12717 |
| Tryptophan metabolism | 28 | 2 | 0.001924 | 0.002168 | 2.57 × 10−9 | 6.64 × 10−9 | 0.12037 |
| Pantothenate and CoA biosynthesis | 23 | 3 | 3.06 × 10−8 | 1.43 × 10−7 | 4.96 × 10−11 | 2.56 × 10−10 | 0.11663 |
| Citrate cycle (TCA cycle) | 20 | 3 | 4.97 × 10−7 | 1.14 × 10−6 | 5.50 × 10−11 | 2.62 × 10−10 | 0.11468 |
| Aminoacyl-tRNA biosynthesis | 46 | 14 | 7.07 × 10−7 | 1.51 × 10−6 | 9.43 × 10−13 | 1.64 × 10−11 | 0.11111 |
| Amino sugar and nucleotide sugar metabolism | 50 | 2 | 1.24 × 10−8 | 8.53 × 10−8 | 9.03 × 10−10 | 2.64 × 10−9 | 0.10791 |
| Inositol phosphate metabolism | 28 | 1 | 2.12 × 10−7 | 6.26 × 10−7 | 3.09 × 10−11 | 1.74 × 10−10 | 0.10251 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araniti, F.; Prinsi, B.; Espen, L. The Delay of Raphanus raphanistrum subsp. sativus (L.) Domin Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism. Plants 2022, 11, 843. https://doi.org/10.3390/plants11070843
Araniti F, Prinsi B, Espen L. The Delay of Raphanus raphanistrum subsp. sativus (L.) Domin Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism. Plants. 2022; 11(7):843. https://doi.org/10.3390/plants11070843
Chicago/Turabian StyleAraniti, Fabrizio, Bhakti Prinsi, and Luca Espen. 2022. "The Delay of Raphanus raphanistrum subsp. sativus (L.) Domin Seed Germination Induced by Coumarin Is Mediated by a Lower Ability to Sustain the Energetic Metabolism" Plants 11, no. 7: 843. https://doi.org/10.3390/plants11070843