
PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
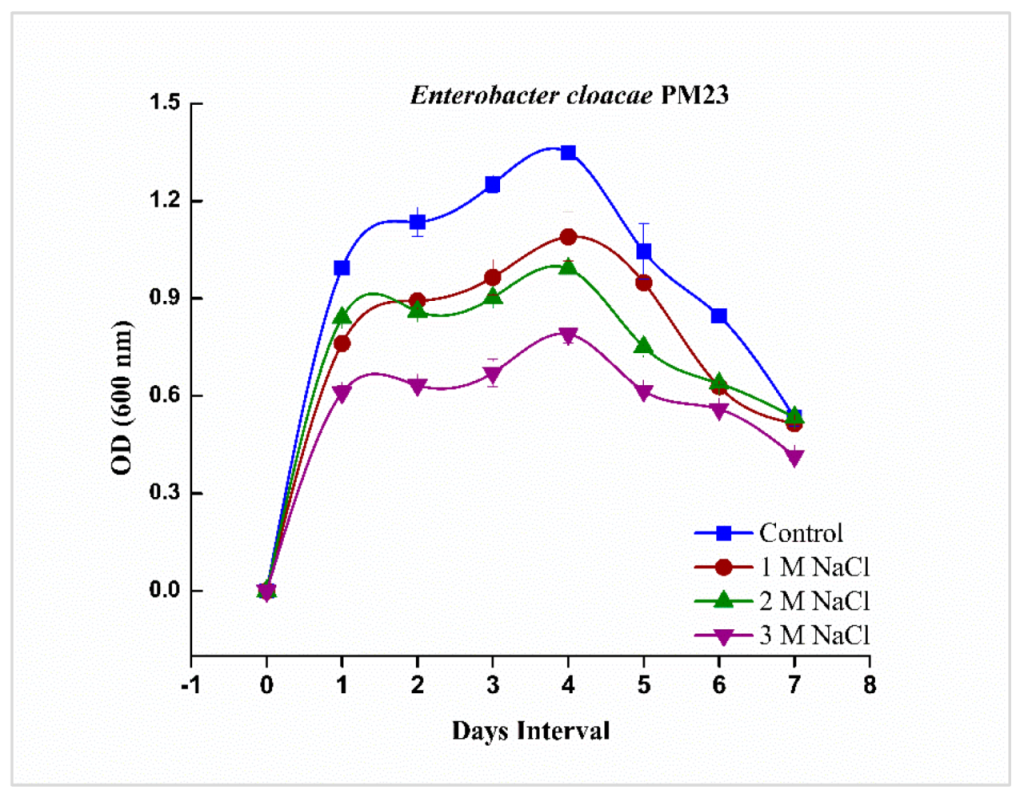
2.1. Growth Curve Analysis of Enterobacter cloacae PM23
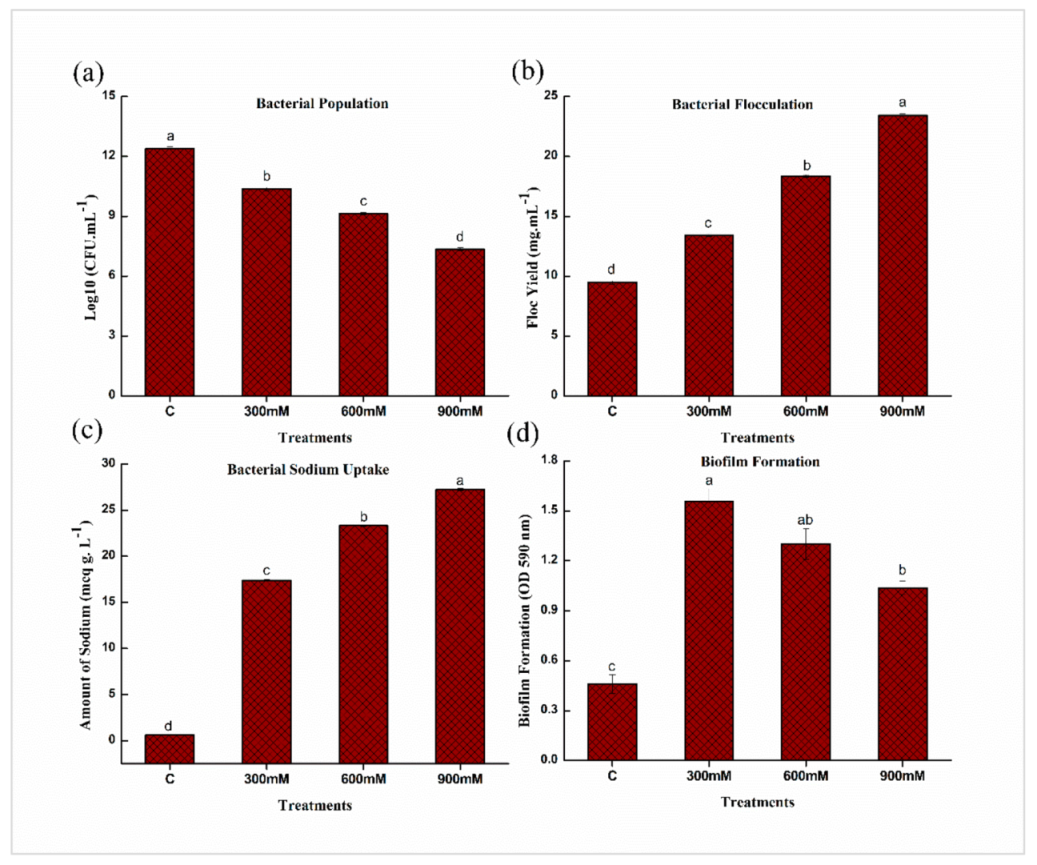
2.2. Salinity Tolerance Traits of E. cloacae PM23 under Salinity Stress
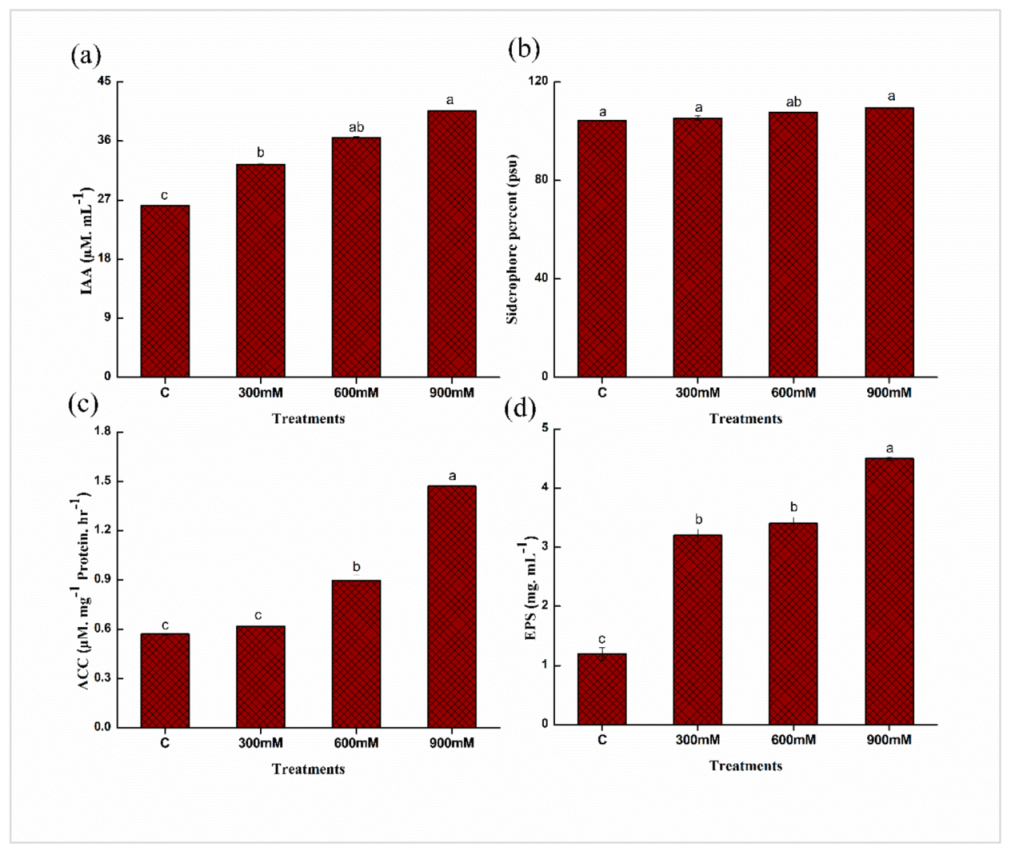
2.3. Quantitative Estimation of Plant Growth Promoting (PGP) Traits of E. cloacae PM23
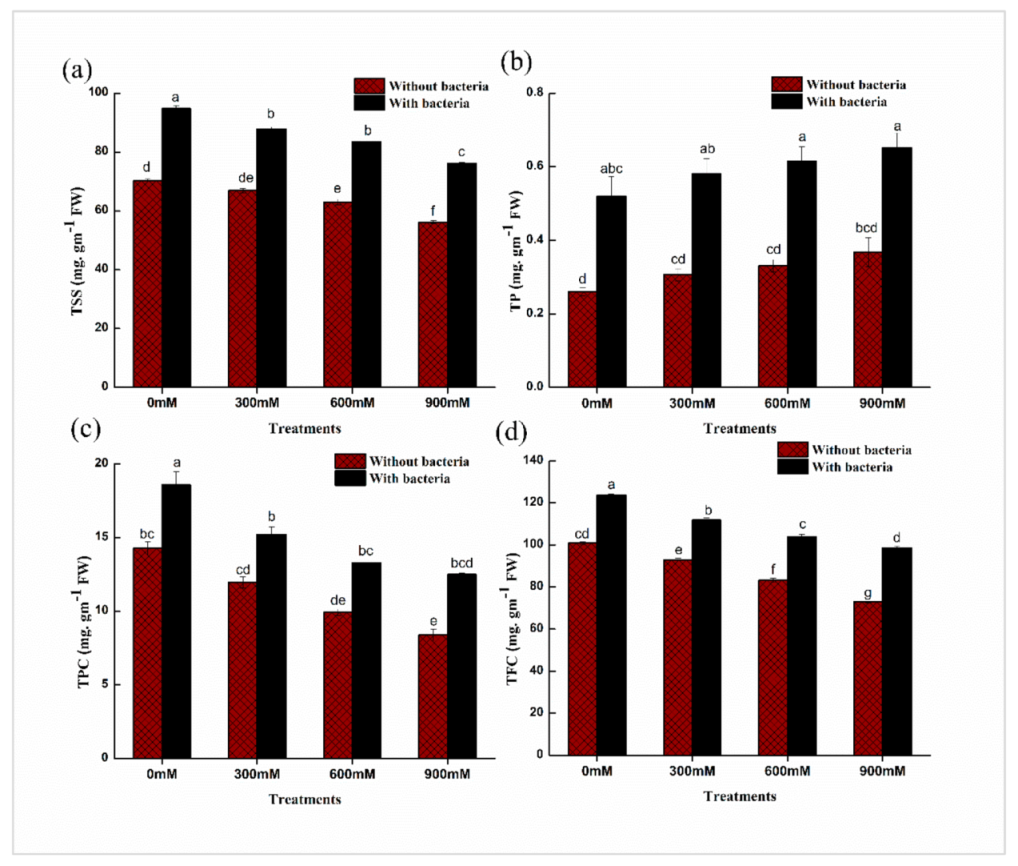
2.4. Physio-Chemical Properties of Soil
2.5. Effects of E. cloacae PM23 on Biomass and Growth of Zea mays L.
2.6. Effects of E. cloacae PM23 on Pigmented Content, Carotenoids, and Relative Water Content of Maize Plants
2.7. Effects of E. cloacae PM23 on Level of Antioxidant Enzymes
2.8. Effects of E. cloacae PM23 on Total Soluble Sugars, Proteins, Flavonoids and Phenolic Content of Leaves
2.9. Assessment of Potential Markers of Oxidative Stress and Compatible Solutes
2.10. PCR Gene Amplification of Biotic and Abiotic Stress-Related Genes
2.11. Gene Expression Analysis
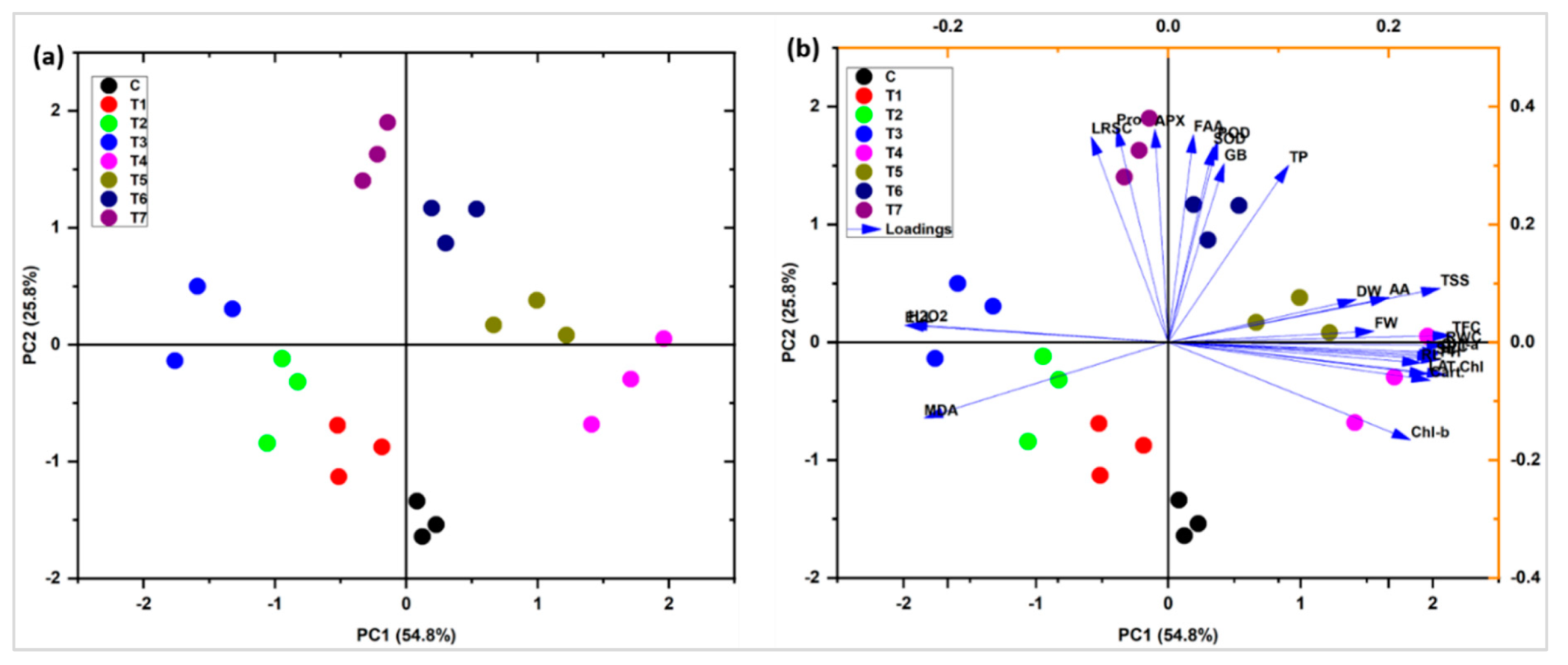
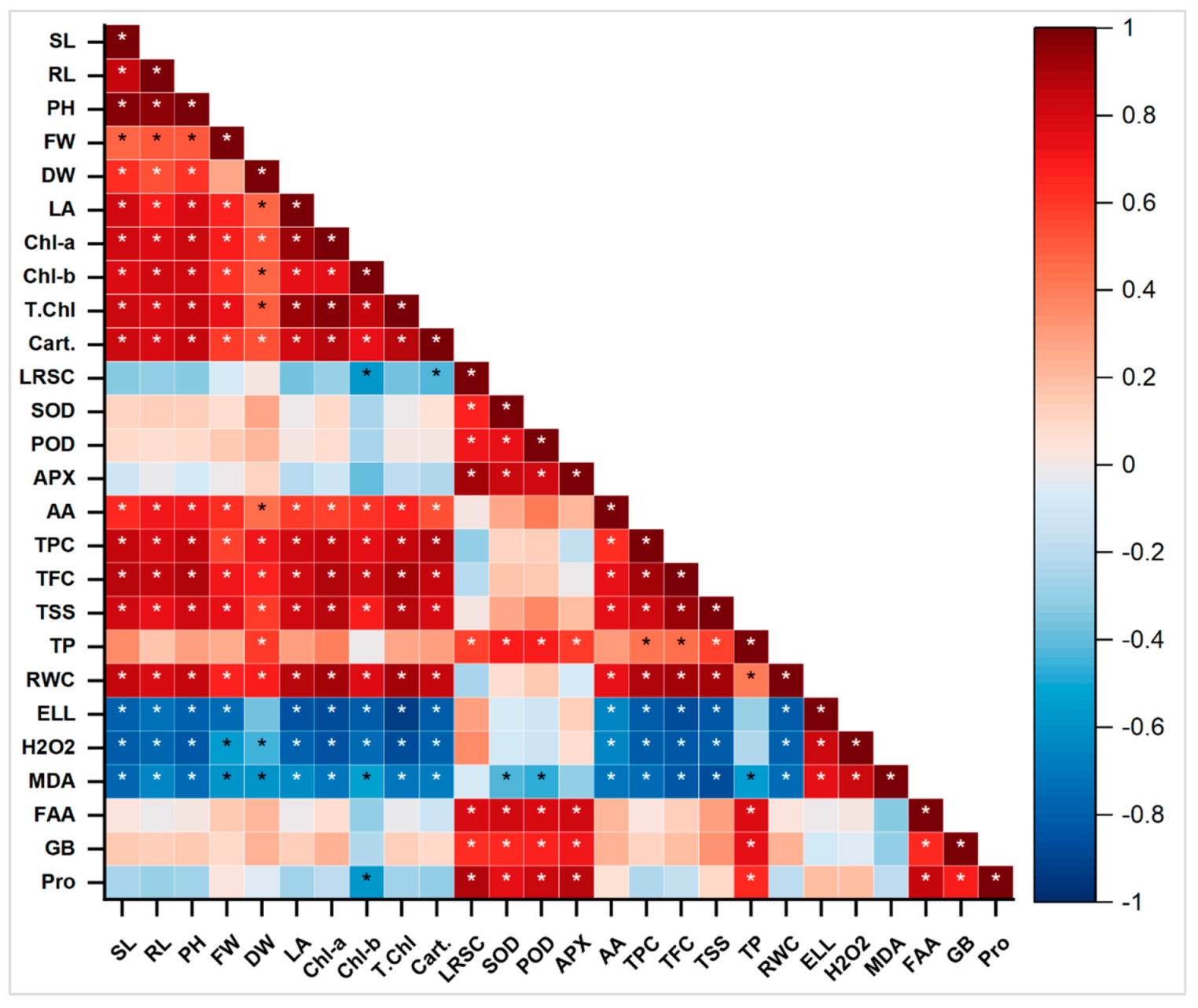
2.12. Principal Component and Pearson Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Procurement of Bacterial Strain
4.2. Salinity Tolerance Characteristics of E. cloacae PM23
4.2.1. Bacterial Survivability
4.2.2. Bacterial Flocculation
4.2.3. Bacterial Sodium Absorption
4.2.4. Biofilm Formation
4.3. Quantitative Assay of Plant Growth-Promoting (PGP) Traits under Salinity Stress
4.3.1. Estimation of Indole Acetic Acid (IAA)
4.3.2. Siderophore Production
4.3.3. Quantitative Estimation of ACC Deaminase
4.3.4. Exopolysaccharide Production (EPS)
4.4. Soil Collection, Analysis, and Seed Inoculation
4.5. Pot Experiment under Controlled Conditions
4.6. Estimation of Agro-Morphological Parameters of Zea Mays L.
4.7. Estimation of Photosynthetic Pigments of Plants
4.8. Relative Water Content (RWC) and Radical Scavenging Capacity of Leaves
4.9. Antioxidant Enzymatic Assays
4.10. Total Soluble Sugars (TSS) and Protein Content of Leaves
4.11. Total Flavonoids and Phenolic Content
4.12. Valuation of Oxidative Stress Markers and Osmolytes
4.13. Amplification of Iturin C (ItuC) and Bio-Surfactant Producing Genes
4.14. Gene Expression Analysis of Antioxidant (APX and SOD) Genes in E. cloacae PM23
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Fan, C.; Wang, Y.; Xia, Y.; Xiao, W.; Cui, X. Salt-tolerant and plant-growth-promoting bacteria isolated from high-yield paddy soil. Can. J. Microbiol. 2018, 64, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Hamayun, M.; Lee, S.-K.; Lee, I.-J. Methyl jasmonate alleviated salinity stress in soybean. J. Crop Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Hamayun, M.; Khan, S.A.; Khan, A.L.; Shin, J.-H.; Ahmad, B.; Shin, D.-H.; Lee, I.-J. Exogenous gibberellic acid reprograms soybean to higher growth and salt stress tolerance. J. Agric. Food Chem. 2010, 58, 7226–7232. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl– ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.-M.; Lee, I.-J. Halotolerant rhizobacterial strains mitigate the adverse effects of NaCl stress in soybean seedlings. BioMed Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Ali, S.; Ali, B.; Hameed, S.; Zhou, W. Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul. 2016, 80, 23–36. [Google Scholar] [CrossRef]
- Zerrouk, I.Z.; Benchabane, M.; Khelifi, L.; Yokawa, K.; Ludwig-Müller, J.; Baluska, F. A Pseudomonas strain isolated from date-palm rhizospheres improves root growth and promotes root formation in maize exposed to salt and aluminum stress. J. Plant Physiol. 2016, 191, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Carpici, E.B.; Celik, N.; Bayram, G. The effects of salt stress on the growth, biochemical parameter and mineral element content of some maize (Zea mays L.) cultivars. Afr. J. Biotechnol. 2010, 9, 6937–6942. [Google Scholar]
- Afridi, M.S.; Van Hamme, J.D.; Bundschuh, J.; Khan, M.N.; Salam, A.; Waqar, M.; Munis, M.F.H.; Chaudhary, H.J. Biotechnological approaches in agriculture and environmental management-bacterium Kocuria rhizophila 14ASP as heavy metal and salt-tolerant plant growth-promoting strain. Biologia 2021, 76, 3091–3105. [Google Scholar] [CrossRef]
- Ullah, S.; Bano, A. Isolation of plant-growth-promoting rhizobacteria from rhizospheric soil of halophytes and their impact on maize (Zea mays L.) under induced soil salinity. Can. J. Microbiol. 2015, 61, 307–313. [Google Scholar] [CrossRef]
- Mehmood, S.; Muneer, M.A.; Tahir, M.; Javed, M.T.; Mahmood, T.; Afridi, M.S.; Pakar, N.P.; Abbasi, H.A.; Munis, M.F.H.; Chaudhary, H.J. Deciphering distinct biological control and growth promoting potential of multi-stress tolerant Bacillus subtilis PM32 for potato stem canker. Physiol. Mol. Biol. Plants 2021, 27, 2101–2114. [Google Scholar] [CrossRef]
- Li, X.; Sun, P.; Zhang, Y.; Jin, C.; Guan, C. A novel PGPR strain Kocuria rhizophila Y1 enhances salt stress tolerance in maize by regulating phytohormone levels, nutrient acquisition, redox potential, ion homeostasis, photosynthetic capacity and stress-responsive genes expression. Environ. Exp. Bot. 2020, 174, 104023. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Han, Q.-Q.; Lü, X.-P.; Bai, J.-P.; Qiao, Y.; Paré, P.W.; Wang, S.-M.; Zhang, J.-L.; Wu, Y.-N.; Pang, X.-P.; Xu, W.-B.; et al. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front. Plant Sci. 2014, 5, 525. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, J.S.; Singh, D.P. Exopolysaccharide-producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 2011, 21, 214–222. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of soil salinity on the plant-growth—promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Kasim, W.A.; Gaafar, R.M.; Abou-Ali, R.M.; Omar, M.N.; Hewait, H.M. Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann. Agric. Sci. 2016, 61, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S. Regulation of antioxidant production, ion uptake and productivity in potato (Solanum tuberosum L.) plant inoculated with growth promoting salt tolerant Bacillus strains. Ecotoxicol. Environ. Saf. 2019, 178, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Egamberdiyeva, D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-Tolerant Plant Growth-Promoting Rhizobacteria Associated with Foxtail Millet in a Semi-arid Agroecosystem and Their Potential in Alleviating Drought Stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef]
- Zhao, X.; Tan, H.J.; Liu, Y.B.; Li, X.R.; Chen, G.X. Effect of salt stress on growth and osmotic regulation in Thellungiella and Arabidopsis callus. Plant Cell Tissue Organ Cult. 2009, 98, 97–103. [Google Scholar] [CrossRef]
- Watanabe, M.; Sasaki, K.; Nakashimada, Y.; Kakizono, T.; Noparatnaraporn, N.; Nishio, N. Growth and flocculation of a marine photosynthetic bacterium Rhodovulum sp. Appl. Microbiol. Biotechnol. 1998, 50, 682–691. [Google Scholar] [CrossRef]
- Fujishige, N.A.; Kapadia, N.N.; De Hoff, P.L.; Hirsch, A.M. Investigations of Rhizobium biofilm formation. FEMS Microbiol. Ecol. 2006, 56, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Fujishige, N.A.; Kapadia, N.N.; Hirsch, A.M. A feeling for the micro-organism: Structure on a small scale. Biofilms on plant roots. Bot. J. Linn. Soc. 2006, 150, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Vimal, S.R.; Patel, V.K.; Singh, J.S. Plant growth promoting Curtobacterium albidum strain SRV4: An agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol. Indic. 2019, 105, 553–562. [Google Scholar] [CrossRef]
- Abbas, H.; Patel, R.M.; Parekh, V.B. Culturable endophytic bacteria from halotolerant Salicornia brachata L.: Isolation and plant growth promoting traits. Indian J. Appl. Microbiol. 2018, 21, 10–21. [Google Scholar]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-producing rhizobacteria as a promising tool for empowering plants to cope with iron limitation in saline soils: A review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Wang, Q.; Dodd, I.C.; Belimov, A.A.; Jiang, F. Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase growth and photosynthesis of pea plants under salt stress by limiting Na+ accumulation. Funct. Plant Biol. 2016, 43, 161–172. [Google Scholar] [CrossRef]
- Mohammed, A.F. Effectiveness of exopolysaccharides and biofilm forming plant growth promoting rhizobacteria on salinity tolerance of faba bean (Vicia faba L.). Afr. J. Microbiol. Res. 2018, 12, 399–404. [Google Scholar]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-tolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [Green Version]
- Desoky, E.-S.M.; El-maghraby, L.M.M.; Awad, A.E.; Abdo, A.I.; Rady, M.M.; Semida, W.M. Fennel and ammi seed extracts modulate antioxidant defence system and alleviate salinity stress in cowpea (Vigna unguiculata). Sci. Hortic. 2020, 272, 109576. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Asghar, H.N.; Arshad, M. Rhizobacteria capable of producing ACC-deaminase may mitigate salt stress in wheat. Soil Sci. Soc. Am. J. 2010, 74, 533–542. [Google Scholar] [CrossRef]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin improves the photosynthetic apparatus in pea leaves stressed by paraquat via chlorophyll breakdown regulation and its accelerated de novo synthesis. Front. Plant Sci. 2017, 8, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, K.P.; Sharma, S.; Singh, N.K.; Singh, V.; Tiwari, K.; Singh, S. Nature and role of root exudates: Efficacy in bioremediation. Afr. J. Biotechnol. 2011, 10, 9717–9724. [Google Scholar]
- Li, H.; Lei, P.; Pang, X.; Li, S.; Xu, H.; Xu, Z.; Feng, X. Enhanced tolerance to salt stress in canola (Brassica napus L.) seedlings inoculated with the halotolerant Enterobacter cloacae HSNJ4. Appl. Soil Ecol. 2017, 119, 26–34. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A. Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ. Exp. Bot. 2019, 159, 55–65. [Google Scholar] [CrossRef]
- Gupta, P.; Kumar, V.; Usmani, Z.; Rani, R.; Chandra, A.; Gupta, V.K. A comparative evaluation towards the potential of Klebsiella sp. and Enterobacter sp. in plant growth promotion, oxidative stress tolerance and chromium uptake in Helianthus annuus (L.). J. Hazard. Mater. 2019, 377, 391–398. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Abraham, J.; Valentine, A. Simultaneous mitigation of aluminum, salinity and drought stress in Lactuca sativa growth via formulated plant growth promoting Rhodotorula mucilaginosa CAM4. Ecotoxicol. Environ. Saf. 2019, 180, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Akhtar, S.S.; Iqbal, S.; Amjad, M.; Naveed, M.; Zahir, Z.A.; Jacobsen, S.-E. Enhancing salt tolerance in quinoa by halotolerant bacterial inoculation. Funct. Plant Biol. 2016, 43, 632–642. [Google Scholar] [CrossRef]
- Aguilera, A.; Tereucan, G.; Ercoli, S.; Cornejo, P.; Gomez, M.R.; Uhlmann, L.; Guigas, C.; Esatbeyoglu, T.; Ruiz, A. Influence of organic and chemical fertilisation on antioxidant compounds profiles and activities in fruits of Fragaria ananassa var. Camarosa. J. Soil Sci. Plant Nutr. 2020, 20, 715–724. [Google Scholar] [CrossRef]
- Patel, D.; Saraf, M. Influence of soil ameliorants and microflora on induction of antioxidant enzymes and growth promotion of Jatropha curcas L. under saline condition. Eur. J. Soil Biol. 2013, 55, 47–54. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Saad, A.M.; El-Saadony, M.T.; Merwad, A.-R.M.; Rady, M.M. Plant growth-promoting rhizobacteria: Potential improvement in antioxidant defense system and suppression of oxidative stress for alleviating salinity stress in Triticum aestivum (L.) plants. Biocatal. Agric. Biotechnol. 2020, 30, 101878. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium protects wheat seedlings against salt stress-mediated oxidative damage by up-regulating antioxidants and osmolytes metabolism. Plant Physiol. Biochem. 2019, 137, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid—A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Veronican Njeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Gusain, Y.S.; Singh, U.S.; Sharma, A.K. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar]
- Morcillo, R.J.L.; Manzanera, M. The Effects of Plant-Associated Bacterial Exopolysaccharides on Plant Abiotic Stress Tolerance. Metabolites 2021, 11, 337. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Mishra, J.; Fatima, T.; Arora, N.K. Role of secondary metabolites from plant growth-promoting rhizobacteria in combating salinity stress. In Plant Microbiome: Stress Response; Springer: Berlin/Heidelberg, Germany, 2018; pp. 127–163. [Google Scholar]
- Bremer, E.; Krämer, R. Responses of microorganisms to osmotic stress. Annu. Rev. Microbiol. 2019, 73, 313–334. [Google Scholar] [CrossRef]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef]
- Rodríguez-Salazar, J.; Suárez, R.; Caballero-Mellado, J.; Iturriaga, G. Trehalose accumulation in Azospirillum brasilense improves drought tolerance and biomass in maize plants. FEMS Microbiol. Lett. 2009, 296, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Glick, B.R.; Rathore, D. Biosurfactants as a biological tool to increase micronutrient availability in soil: A review. Pedosphere 2018, 28, 170–189. [Google Scholar] [CrossRef]
- Płaza, G.; Achal, V. Biosurfactants: Eco-friendly and innovative biocides against biocorrosion. Int. J. Mol. Sci. 2020, 21, 2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Yuan, D.; Jin, C.; Wang, G.; Li, X.; Guan, C. Enhancement of growth and salt tolerance of rice seedlings (Oryza sativa L.) by regulating ethylene production with a novel halotolerant PGPR strain Glutamicibacter sp. YD01 containing ACC deaminase activity. Acta Physiol. Plant. 2020, 42, 1–17. [Google Scholar] [CrossRef]
- Afridi, M.S.; Amna; Sumaira; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; et al. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; López, G.; Coniglio, A.; Furlan, A.; Mora, V.; Rosas, S.; Cassán, F. Day and blue light modify growth, cell physiology and indole-3-acetic acid production of Azospirillum brasilense Az39 under planktonic growth conditions. J. Appl. Microbiol. 2021, 130, 1671–1683. [Google Scholar] [CrossRef]
- Shultana, R.; Kee, Z.A.T.; Yusop, M.R.; Saud, H.M. Characterization of salt-tolerant plant growth-promoting rhizobacteria and the effect on growth and yield of saline-affected rice. PLoS ONE 2020, 15, e0238537. [Google Scholar] [CrossRef]
- Prouty, A.M.; Schwesinger, W.H.; Gunn, J.S. Biofilm formation and interaction with the surfaces of gallstones by Salmonella spp. Infect. Immun. 2002, 70, 2640–2649. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Loper, J.E.; Schroth, M.N. Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology 1986, 76, 386–389. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Mehmood, S.; Khatoon, Z.; Amna; Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F.H. Bacillus sp. PM31 harboring various plant growth-promoting activities regulates Fusarium dry rot and wilt tolerance in potato. Arch. Agron. Soil Sci. 2021, 1–15. [Google Scholar] [CrossRef]
- Zainab, N.; Amna; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; et al. PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania sesban L. in Industrially Contaminated Soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]
- Zainab, N.; Din, B.U.; Javed, M.T.; Afridi, M.S.; Mukhtar, T.; Kamran, M.A.; Khan, A.A.; Ali, J.; Jatoi, W.N.; Munis, M.F.H. Deciphering metal toxicity responses of flax (Linum usitatissimum L.) with exopolysaccharide and ACC-deaminase producing bacteria in industrially contaminated soils. Plant Physiol. Biochem. 2020, 152, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric acid assisted phytoremediation of cadmium by Brassica napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef]
- Ali, J.; Ali, F.; Ahmad, I.; Rafique, M.; Munis, M.F.H.; Hassan, S.W.; Sultan, T.; Iftikhar, M.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of Sesbania sesban seedlings with Bacillus anthracis PM21 under heavy metals stress: An in vitro study. Ecotoxicol. Environ. Saf. 2021, 208, 111769. [Google Scholar] [CrossRef]
- Shi, P.J.; Li, Y.R.; Niinemets, Ü.; Olson, E.; Schrader, J. Influence of leaf shape on the scaling of leaf surface area and length in bamboo plants. Trees 2021, 35, 709–715. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 improves salt stress tolerance in maize by regulating redox potential, ion homeostasis, leaf gas exchange and stress-related gene expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef] [Green Version]
- Asgari, H.T.; Es-haghi, A.; Karimi, E. Anti-angiogenic, antibacterial, and antioxidant activities of nanoemulsions synthesized by Cuminum cyminum L. tinctures. J. Food Meas. Charact. 2021, 15, 3649–3659. [Google Scholar] [CrossRef]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar]
- El-Saadony, M.T.; Desoky, E.-S.M.; Saad, A.M.; Eid, R.S.M.; Selem, E.; Elrys, A.S. Biological silicon nanoparticles improve Phaseolus vulgaris L. yield and minimize its contaminant contents on a heavy metals-contaminated saline soil. J. Environ. Sci. 2021, 106, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Grad, W.E.; Kandil, S.H.; Kenawy, E.; Massoud, M.I. The potential of sugarcane bagasse polymer composite for sustainable of Stevia rebaudiana productivity under deficit irrigation. SVU-Int. J. Agric. Sci. 2021, 3, 22–36. [Google Scholar] [CrossRef]
- Mendez, R.L.; Kwon, J.Y. Effect of extraction condition on protein recovery and phenolic interference in Pacific dulse (Devaleraea mollis). J. Appl. Phycol. 2021, 33, 2497–2509. [Google Scholar] [CrossRef]
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apic. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Seke, F.; Manhivi, V.E.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Effect of Freeze Drying and Simulated Gastrointestinal Digestion on Phenolic Metabolites and Antioxidant Property of the Natal Plum (Carissa macrocarpa). Foods 2021, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Caser, M.; D’Angiolillo, F.; Chitarra, W.; Lovisolo, C.; Ruffoni, B.; Pistelli, L.; Pistelli, L.; Scariot, V. Water deficit regimes trigger changes in valuable physiological and phytochemical parameters in Helichrysum petiolare Hilliard & BL Burtt. Ind. Crops Prod. 2016, 83, 680–692. [Google Scholar]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2007, 104, 1372–1378. [Google Scholar] [CrossRef]
- Nisar, S.; Dar, R.A.; Bhat, A.A.; Farooq, Z.; Tahir, I. Some important biochemical changes orchestrating flower development and senescence in Nicotiana plumbaginifolia Viv. and Petunia hybrida Vilm. flowers. J. Hortic. Sci. Biotechnol. 2021, 96, 759–769. [Google Scholar] [CrossRef]
- Kapoor, R.T.; Alyemeni, M.N.; Ahmad, P. Exogenously applied spermidine confers protection against cinnamic acid-mediated oxidative stress in Pisum sativum. Saudi J. Biol. Sci. 2021, 28, 2619–2625. [Google Scholar] [CrossRef]
- Tulkova, E.; Kabashnikova, L. Malondialdehyde content in the leaves of small-leaved linden tilia cordata and Norway maple acer platanoides under the influence of volatile organic compounds. Plant Biosyst. 2021, 1–9. [Google Scholar] [CrossRef]
- Shafiq, F.; Iqbal, M.; Ali, M.; Ashraf, M.A. Fullerenol regulates oxidative stress and tissue ionic homeostasis in spring wheat to improve net-primary productivity under salt-stress. Ecotoxicol. Environ. Saf. 2021, 211, 111901. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Bustos, M.G.; Aguado-Santacruz, G.A.; Tiessen-Favier, A.; Robledo-Paz, A.; Munoz-Orozco, A.; Rasco’n-Cruz, Q.; Santacruz-Varela, A. A reliable method for spectrophotometric determination of glycine betaine in cell suspension and other systems. Anal. Biochem. 2016, 498, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Parveen, A.; Siddiqui, Z.A. Zinc oxide nanoparticles affect growth, photosynthetic pigments, proline content and bacterial and fungal diseases of tomato. Arch. Phytopathol. Plant Prot. 2021, 54, 1–20. [Google Scholar] [CrossRef]
- Ahmad, S.; Chaudhary, H.J.; Damalas, C.A. Microbial detoxification of dimethoate through mediated hydrolysis by Brucella sp. PS4: Molecular profiling and plant growth-promoting traits. Environ. Sci. Poll. Res. 2021, 29, 2420–2431. [Google Scholar] [CrossRef]
- Swaathy, S.; Kavitha, V.; Sahaya Pravin, A.; Sekaran, G.; Mandal, A.B.; Gnanamani, A. Phylogenetic framework and biosurfactant gene expression analysis of marine Bacillus spp. of Eastern Coastal Plain of Tamil Nadu. Int. J. Bacteriol. 2014, 2014, 860491. [Google Scholar] [CrossRef]
- Chung, S.; Kong, H.; Buyer, J.S.; Lakshman, D.K.; Lydon, J.; Kim, S.-D.; Roberts, D.P. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soilborne pathogens of cucumber and pepper. Appl. Microbiol. Biotechnol. 2008, 80, 115–123. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Parameters | Soil 1 (Pre-Sowing) | Soil 2 (Post-Harvesting) |
|---|---|---|
| Soil texture | Loamy | Loamy |
| pH | 7.94 | 7.87 |
| Electrical conductivity (dS/m) | 1.53 | 4.49 |
| Organic matter (%) | 3.49 | 1.88 |
| Available Phosphorus (mg/kg) | 45.62 | 33.85 |
| Available Potassium (mg/kg) | 601 | 124 |
| Saturation (%) | 44 | 43 |
| NaCl (mM) | E. cloacae PM23 | SL (cm) | RL (cm) | PH (cm) | FW (g) | DW (g) | LA (cm2) |
|---|---|---|---|---|---|---|---|
| 0 mM | −PM23 | 30.4 ± 1.22 bc | 13.55 ± 0.5 b | 43.95 ± 1.01 bc | 1.30 ± 0.19 abc | 0.5 ± 0.10 ab | 15.01 ± 0.58 bc |
| +PM23 | 38.5 ± 0.88 a | 18.9 ± 1.10 a | 57.4 ± 1.71 a | 2.15 ± 0.11 a | 0.93 ± 0.08 a | 20.75 ± 1.24 a | |
| 300 mM | −PM23 | 26 ± 1.06 cd | 10.83 ± 0.42 bc | 36.83 ± 1.47 cd | 0.93 ± 0.16 c | 0.42 ± 0.06 ab | 13.22 ± 0.42 c |
| +PM23 | 34.2 ± 1.13 ab | 15 ± 1.21 ab | 49.2 ± 2.35 ab | 1.83 ± 0.15 ab | 0.86 ± 0.13 a | 18.51 ± 0.88 ab | |
| 600 mM | −PM23 | 22 ± 0.79 de | 7.87 ± 0.41 cd | 29.87 ± 1.09 de | 0.79 ± 0.14 c | 0.37 ± 0.07 ab | 11.46 ± 0.27 cd |
| +PM23 | 30.45 ± 1.09 bc | 13.25 ± 0.80 b | 43.7 ± 1.05 bc | 1.44 ± 0.12 abc | 0.75 ± 0.11 ab | 14.54 ± 0.56 bc | |
| 900 mM | −PM23 | 16.33 ± 1.19 e | 6 ± 0.43 d | 22.33 ± 1.63 e | 0.62 ± 0.06 c | 0.26 ± 0.03 b | 8.36 ± 0.31 d |
| +PM23 | 25.38 ± 1.17 cd | 10.9 ± 0.22 bc | 36.28 ± 1.34 cd | 1.26 ± 0.06 bc | 0.62 ± 0.06 ab | 11.34 ± 0.17 cd |
| NaCl (mM) | E. cloacae PM23 | Chl a (mg/g FW) | Chl b (mg/g FW) | Total Chl (mg/g FW) | Carotenoids (mg/g FW) | RWC (%) |
|---|---|---|---|---|---|---|
| 0 mM | −PM23 | 17.1 ± 0.90 cd | 5.44 ± 0.10 ab | 22.54 ± 0.79 b | 6.01 ± 0.20 bc | 55.43 ± 0.74 cd |
| +PM23 | 25.16 ± 0.80 a | 6.1 ± 0.16 a | 31.26 ± 0.67 a | 8.58 ± 0.15 a | 72.34 ± 0.95 a | |
| 300 mM | −PM23 | 14.05 ± 0.32 cde | 4.50 ± 0.12 c | 18.55 ± 0.19 bcd | 5.43 ± 0.12 bcd | 50.73 ± 1.15 de |
| +PM23 | 21.96 ± 0.83 ab | 5.31 ± 0.1 b | 25.2 ± 0.58 b | 6.5 ± 0.17 b | 66.27 ± 1.07 b | |
| 600 mM | −PM23 | 12.36 ± 0.55 ef | 3.51 ± 0.11 de | 15.87 ± 0.52 d | 4.77 ± 0.13 de | 47.21 ± 0.81 ef |
| +PM23 | 17.84 ± 0.48 bc | 4.29 ± 0.11 cd | 22.14 ± 0.43 bc | 5.57 ± 0.14 bcd | 59.41 ± 1.09 c | |
| 900 mM | −PM23 | 9.33 ± 0.39 f | 2.39 ± 0.13 e | 11.72 ± 0.49 e | 3.91 ± 0.21 e | 44.22 ± 0.79 f |
| +PM23 | 13.48 ± 0.57 def | 3.46 ± 0.08 f | 16.95 ± 0.52 cd | 4.96 ± 0.21 cde | 50.94 ± 0.67 de |
| NaCl (mM) | E. cloacae PM23 | DPPH (IC50) % | EL (%) | H2O2 (µmol/g FW) | MDA (nmol/g FW) |
|---|---|---|---|---|---|
| 0 mM | −PM23 | 30.80 ± 0.64 e | 45.4 ± 1.09 cd | 28.50 ± 0.60 cd | 9.73 ± 0.30 bc |
| +PM23 | 35.81 ± 0.42 d | 39.1 ± 0.81 e | 21.36 ± 0.66 e | 4.32 ± 0.22 d | |
| 300 mM | −PM23 | 35.15 ± 1.10 d | 51.33 ± 0.66 bc | 31.24 ± 0.92 bc | 10.54 ± 0.34 b |
| +PM23 | 40.42 ± 0.18 c | 43.53 ± 1.17 de | 25.53 ± 0.22 de | 5.28 ± 0.55 d | |
| 600 mM | −PM23 | 42.97 ± 0.80 c | 56.6 ± 1.34 b | 35.72 ± 1.09 ab | 12.02 ± 0.47 ab |
| +PM23 | 58.27 ± 0.19 a | 45.8 ± 0.81 cd | 29.38 ± 1.06 cd | 5.72 ± 0.49 d | |
| 900 mM | −PM23 | 48.74 ± 0.87 b | 63.43 ± 0.41 a | 38.63 ± 0.89 a | 14.24 ± 0.96 a |
| +PM23 | 60.31 ± 0.17 a | 51.96 ± 0.82 b | 31.08 ± 1.05 bcd | 6.49 ± 0.11 cd |
| NaCl (mM) | E. cloacae PM23 | FAA (mg/g DW) | GB (µg/g DW) | Proline (µmol/g FW) |
|---|---|---|---|---|
| 0 mM | −PM23 | 13.39 ± 1.25 d | 8.01 ± 0.43 d | 60.48 ± 1.06 f |
| +PM23 | 18.26 ± 1.35 cd | 11.63 ± 0.16 abc | 68.96 ± 0.80 de | |
| 300 mM | −PM23 | 15.97 ± 0.93 d | 8.82 ± 0.40 cd | 65.88 ± 1.06 ef |
| +PM23 | 22.67 ± 0.66 bc | 12.09 ± 0.39 ab | 73.8 ± 1.48 cd | |
| 600 mM | −PM23 | 17.25 ± 0.79 cd | 9.43 ± 0.53 bcd | 70.39 ± 1.05 de |
| +PM23 | 26.78 ± 0.94 ab | 12.64 ± 0.52 a | 80.47 ± 0.40 ab | |
| 900 mM | −PM23 | 19.42 ± 0.78 cd | 11.78 ± 0.52 abc | 78.53 ± 1.05 bc |
| +PM23 | 30.86 ± 0.79 a | 13.56 ± 0.66 a | 86.55 ± 0.69 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Zaib-Un-Nisa; Ullah, I.; Amaral Júnior, A.T.d.; et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. https://doi.org/10.3390/plants11030345
Ali B, Wang X, Saleem MH, Sumaira, Hafeez A, Afridi MS, Khan S, Zaib-Un-Nisa, Ullah I, Amaral Júnior ATd, et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants. 2022; 11(3):345. https://doi.org/10.3390/plants11030345
Chicago/Turabian StyleAli, Baber, Xiukang Wang, Muhammad Hamzah Saleem, Sumaira, Aqsa Hafeez, Muhammad Siddique Afridi, Shahid Khan, Zaib-Un-Nisa, Izhar Ullah, Antônio Teixeira do Amaral Júnior, and et al. 2022. "PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes" Plants 11, no. 3: 345. https://doi.org/10.3390/plants11030345