
Effects of Soil Water Shortage on Seedling Shoot and Root Growth of Saragolle Lucana Tetraploid Wheat (Triticum durum Desf.) Landrace

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
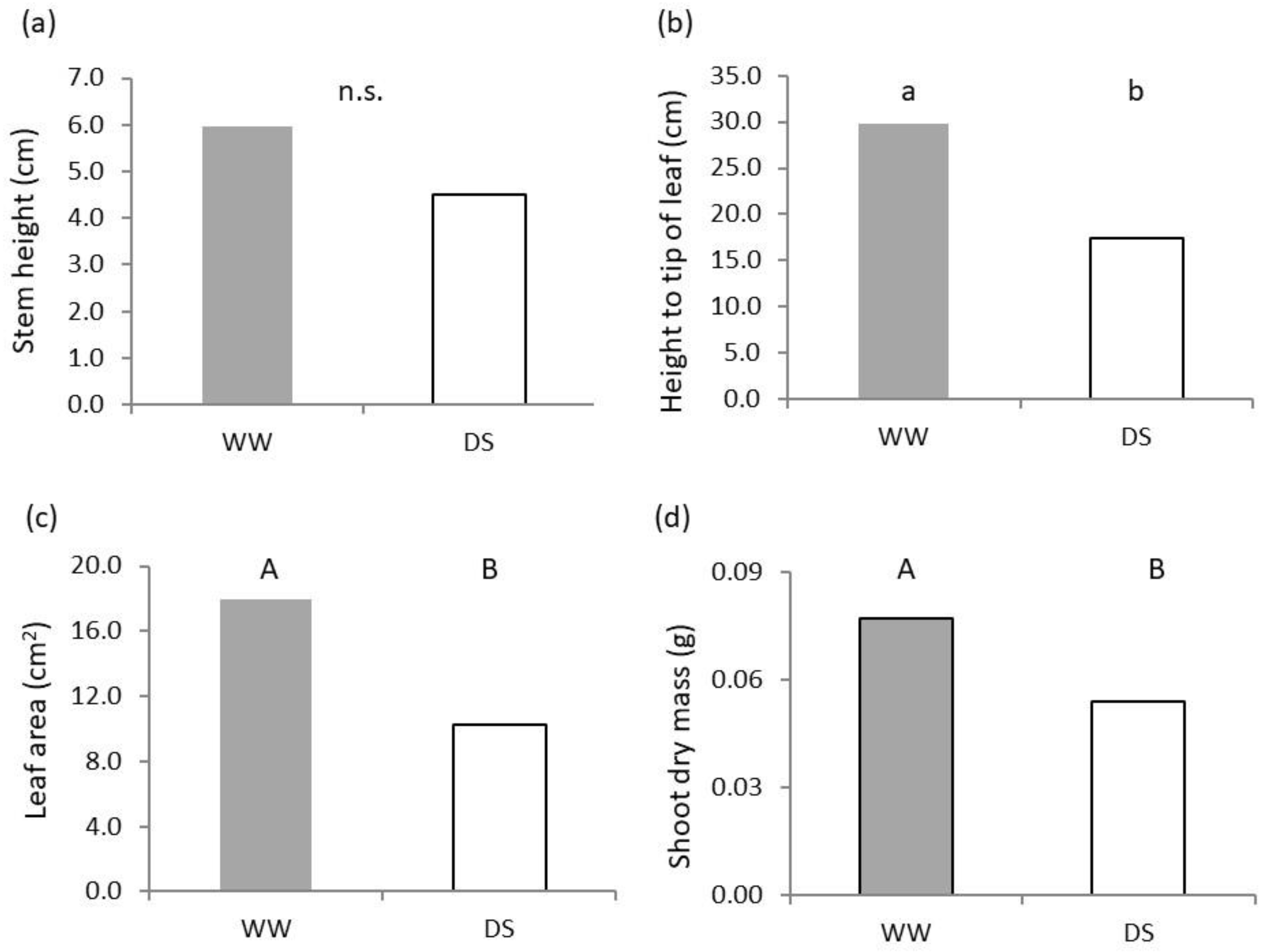
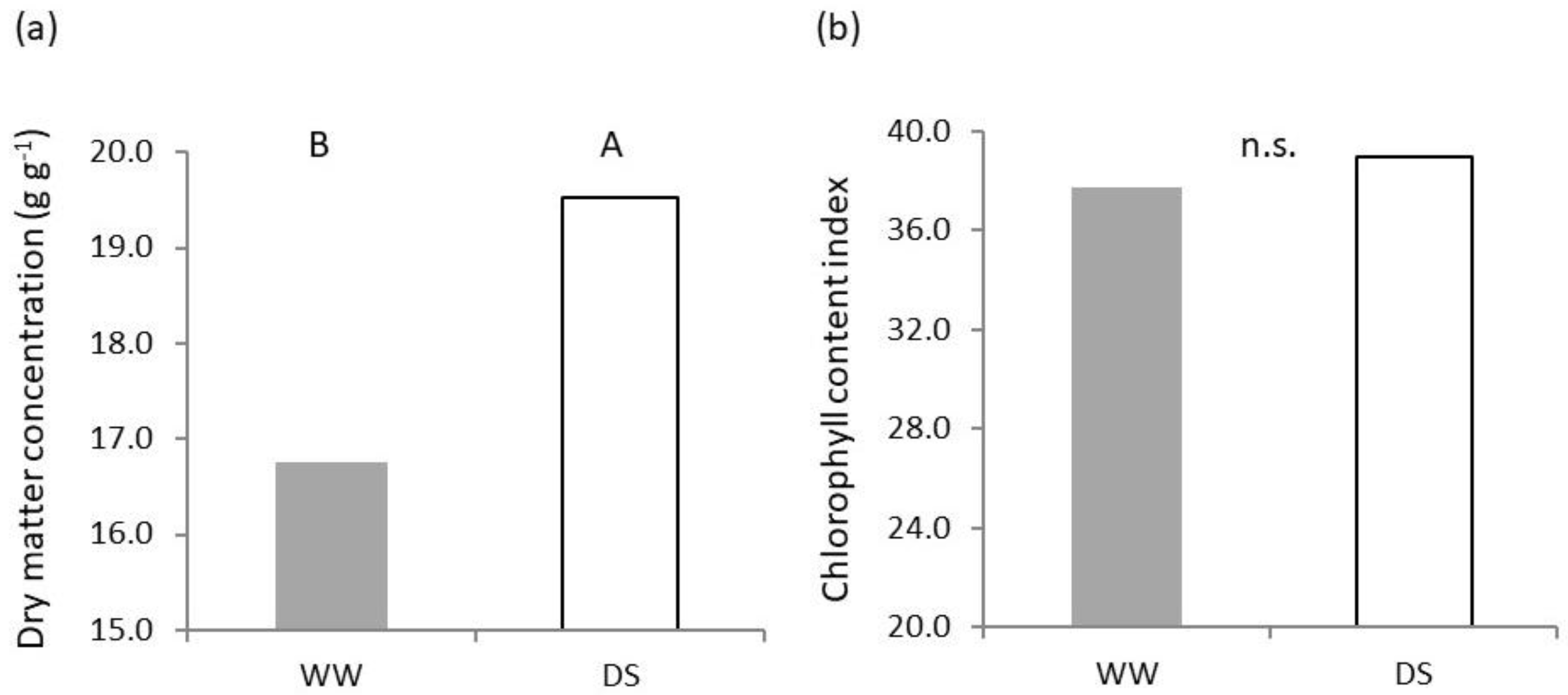
2.1. Shoot Growth
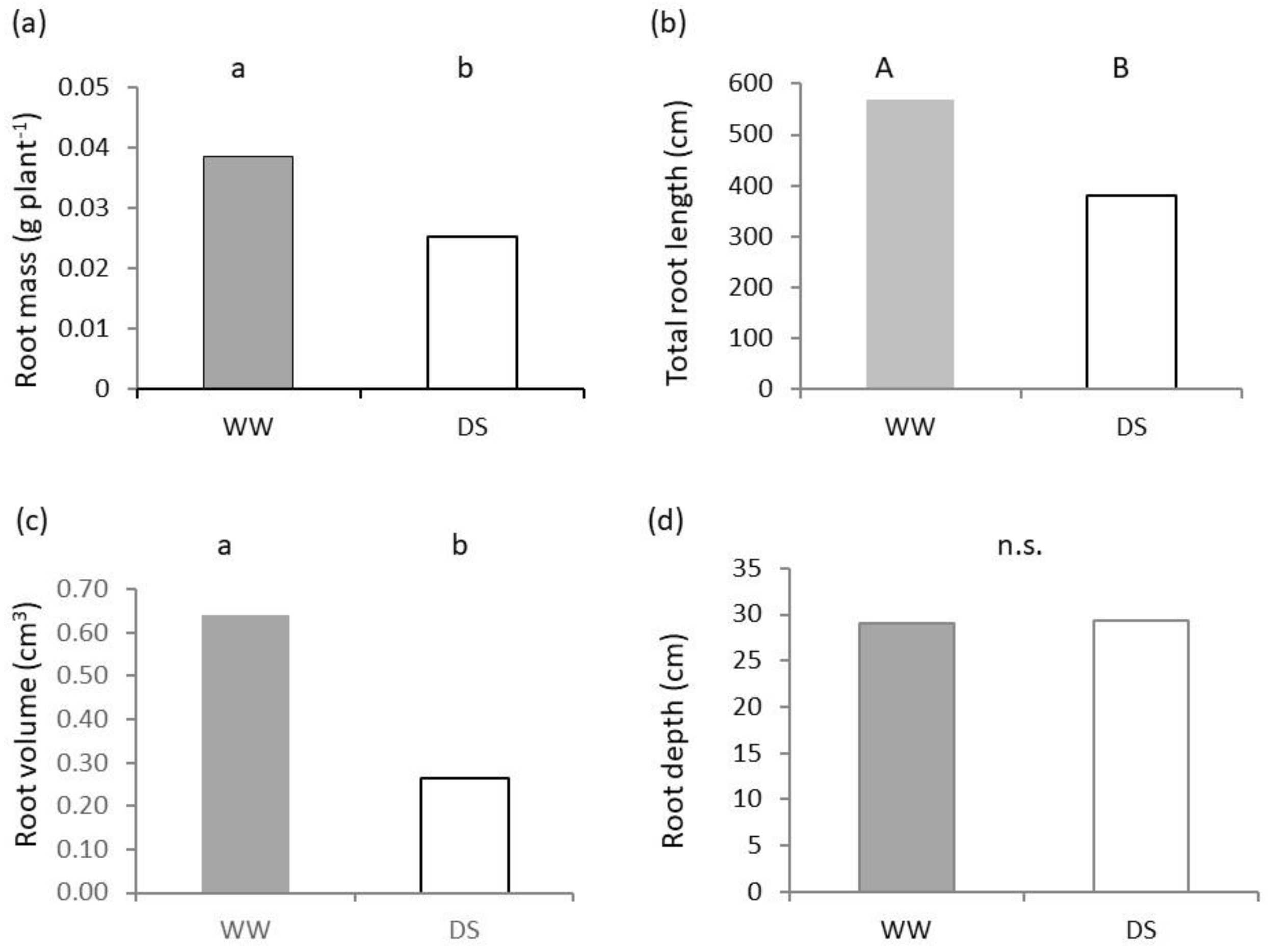
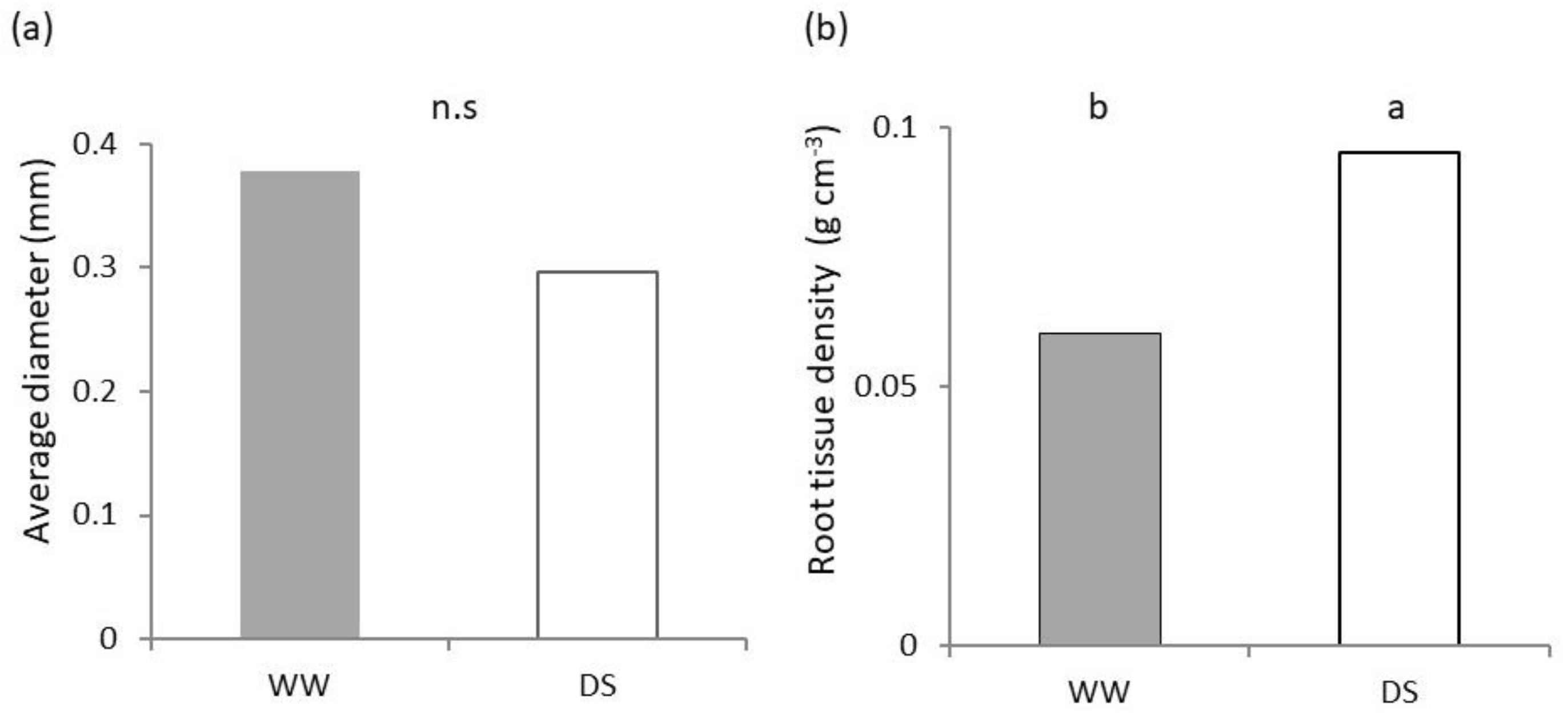
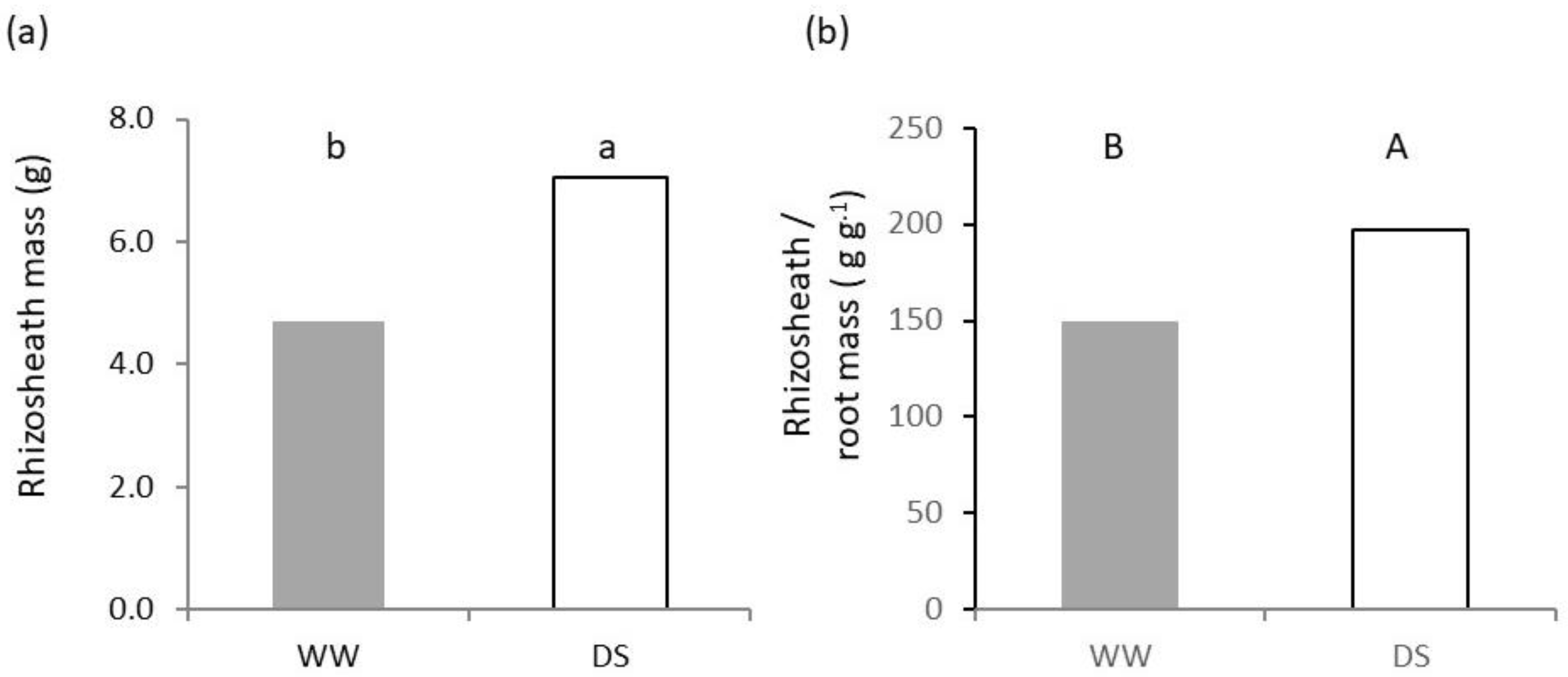
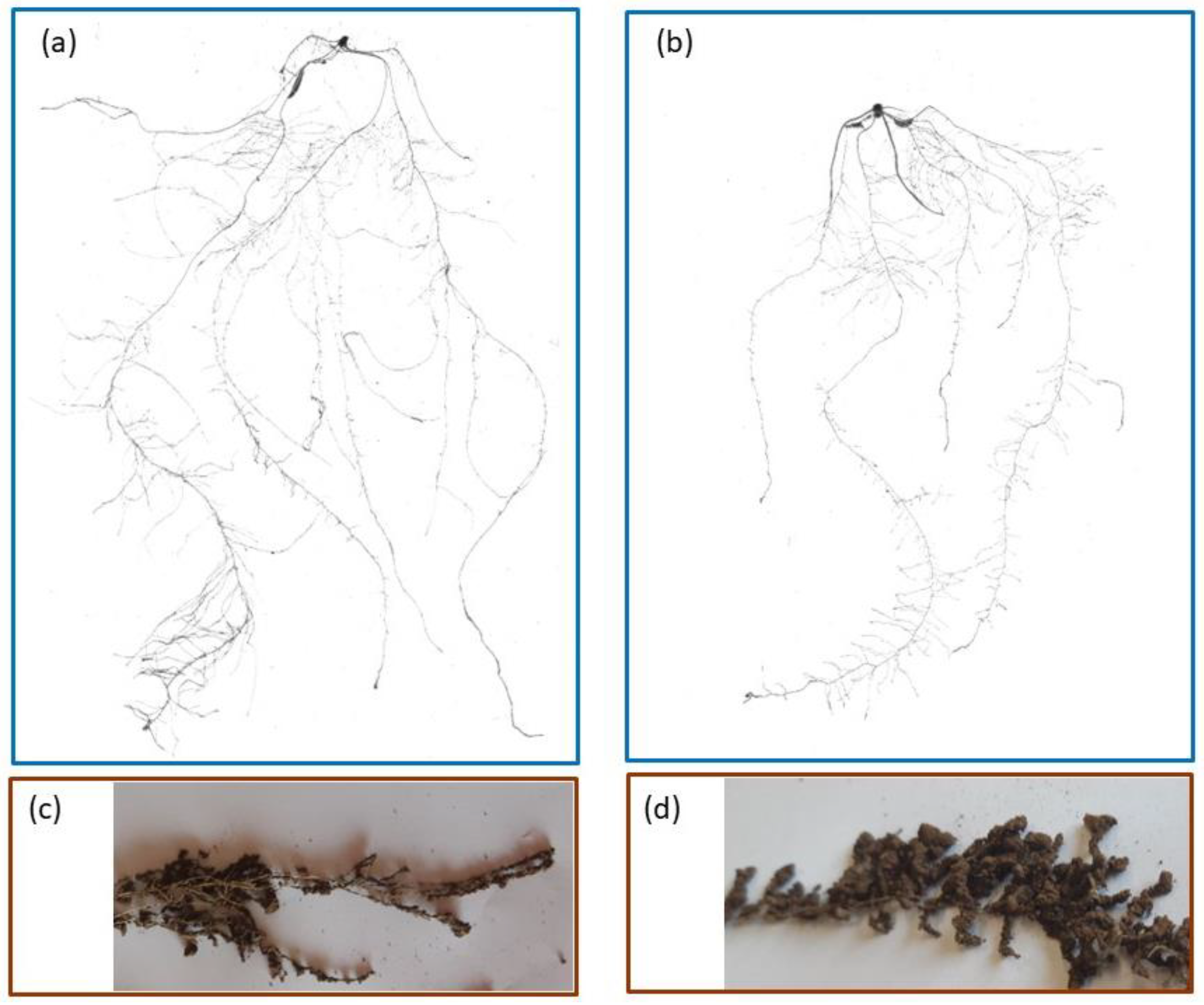
2.2. Belowground Plant Growth and Architecture
2.3. Allometrics
3. Discussion
4. Materials and Methods
4.1. Available Water Treatments
- (i)
- Well-watered control (WW) where potting soil was mixed with water in order to bring it to field capacity. This corresponds to filling pots to 100% of AWC corresponding to 47.8% of total porosity.
- (ii)
- Low available water for water deficit stress (DS) where potting soil was mixed with water in order to bring it to 50% of AWC corresponding to 23.9 % of total porosity.
4.2. Seedlings Growth Conditions
4.3. Measurements
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zampieri, M.; Ceglar, A.; Dentener, F.; Toreti, A. Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. 2017, 12, 064008. [Google Scholar] [CrossRef]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobell, D.B.; Tebaldi, C. Getting caught with our plants down: The risks of a global crop yield slowdown from climate trends in the next two decades. Environ. Res. Lett. 2014, 9, 074003. [Google Scholar] [CrossRef]
- Kimurto, P.K.; Kinyua, M.G.; Birechl, R.; Korir, P.C.; Njoka, E.M.; Njau, P.N. Root and shoot characteristics as selection criteria for drought tolerance in bread wheat (Triticum aestivum L.) at seedling stage under tropical environment. Discov. Innov. 2005, 17, 74–84. [Google Scholar] [CrossRef]
- Sanguineti, M.C.; Li, S.; Maccaferri, M.; Corneti, S.; Rotondo, F.; Chiari, T.; Tuberosa, R. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann. Appl. Biol. 2007, 151, 291–305. [Google Scholar] [CrossRef]
- Jacques, C.; Girodet, S.; Kreplak, J.; Salon, C.; Prudent, M. Do pea nodulated roots have a memory like a sieve or like an elephant when faced with recurrent water deficits? In Proceedings of the 11th Symposium of the International Society of Root Research ISRR-11-Rooting 2021 Joint Conference, Virtuel, France, 24–28 May 2021. [Google Scholar]
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Rabbi, S.M.F.; Warren, C.R.; Macdonald, C.; Trethowan, R.M.; Young, I.M. Soil-root interaction in the rhizosheath regulates the water uptake of wheat. Rhizosphere 2022, 21, 100462. [Google Scholar] [CrossRef]
- Becker, S.P.; Byrne, P.F.; Reid, S.D.; Bauerle, W.L.; McKay, J.K.; Haley, S.D. Root traits contributing to drought tolerance of synthetic hexaploid wheat in a greenhouse study. Euphytica 2016, 207, 213–224. [Google Scholar] [CrossRef]
- Ekanayake, I.J.; O’toole, J.C.; Garrity, D.P.; Masajo, T.M. Inheritance of root characters and their relations to drought resistance in rice 1. Crop Sci. 1985, 25, 927–933. [Google Scholar] [CrossRef]
- Salarpour, M.; Pakniyat, H.; Abdolshahi, R.; Heidari, B.; Razi, H.; Afzali, R. Mapping QTL for agronomic and root traits in the Kukri/RAC875 wheat (Triticum aestivum L.) population under drought stress conditions. Euphytica 2020, 216, 105. [Google Scholar] [CrossRef]
- Brown, L.K.; George, T.S.; Neugebauer, K.; White, P.J. The rhizosheath—A potential trait for future agricultural sustainability occurs in orders throughout the angiosperms. Plant Soil. 2017, 418, 115–128. [Google Scholar] [CrossRef]
- Ahmadi, K.; Zarebanadkouki, M.; Ahmed, M.A.; Ferrarini, A.; Kuzyakov, Y.; Kostka, S.I.; Carminati, A. Rhizosphere engineering: Innovative improvement of root environment. Rhizosphere 2017, 3, 176–184. [Google Scholar] [CrossRef]
- Basirat, M.; Mousavi, S.M.; Abbaszadeh, S.A.; Ebrahimi, M.; Zarebanadkouki, M. The rhizosheath: A potential root trait helping plants to tolerate drought stress. Plant Soil 2019, 445, 565–575. [Google Scholar] [CrossRef]
- Amato, M.; Bochicchio, R.; Mele, G.; Labella, R.; Rossi, R. Soil structure and stability in the spermosphere of myxosdiaspore chia (Salvia hispanica L.). Soil Res. 2018, 57, 546–558. [Google Scholar] [CrossRef]
- Watt, M.; Mc Cully, M.E.; Canny, M.J. Formation and stabilization of rhizosheaths of Zea mays L. Plant Physiol. 1994, 106, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giunta, F.; Bassu, S.; Mefleh, M.; Motzo, R. Is the technological quality of old durum wheat cultivars superior to that of modern ones when exposed to moderately high temperatures during grain filling? Foods 2020, 9, 778. [Google Scholar] [CrossRef]
- Mefleh, M.; Conte, P.; Fadda, C.; Giunta, F.; Piga, A.; Hassoun, G.; Motzo, R. From ancient to old and modern durum wheat varieties: Interaction among cultivar traits, management, and technological quality. J. Sci. Food Agric. 2019, 99, 2059–2067. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Motzo, R. Grain yield and grain protein of old and modern durum wheat cultivars grown under different cropping systems. Field. Crop. Res. 2019, 230, 107–120. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Cadeddu, F.; Motzo, R. Old tall durum wheat cultivars are suited for dual-purpose utilization. Eur. J. Agron. 2017, 90, 67–77. [Google Scholar] [CrossRef]
- Cadeddu, F.; Motzo, R.; Mureddu, F.; Giunta, F. Effects of clipping on the nitrogen economy of four Triticum species grown in a Mediterranean environment. Field Crop. Res. 2021, 267, 108162. [Google Scholar] [CrossRef]
- Bochicchio, R.; Labella, R.; Vitti, A.; Nuzzaci, M.; Logozzo, G.; Amato, M. Root Morphology, Allometric Relations and Rhizosheath of Ancient and Modern Tetraploid Wheats (Triticum durum Desf.) in Response to Inoculation with Trichoderma harzianum T-22. Plants 2022, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Nakhforoosh, A.; Nagel, K.A.; Fiorani, F.; Bodner, G. Deep soil exploration vs. topsoil exploitation: Distinctive rooting strategies between wheat landraces and wild relatives. Plant Soil 2021, 459, 397–421. [Google Scholar] [CrossRef] [PubMed]
- Akman, H.; Akgun, N.; Tamkoc, A. Screening for root and shoot traits in different wheat species and wild wheat relatives. Bot. Sci. 2017, 95, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Salvi, S.; Royo, C.; Soriano, J.M. Exploring the genetic architecture of root-related traits in mediterranean bread wheat landraces by genome-wide association analysis. Agronomy 2020, 10, 613. [Google Scholar] [CrossRef]
- Bektas, H.; Hohn, C.E.; Waines, J.G. Root and shoot traits of bread wheat (Triticum aestivum L.) landraces and cultivars. Euphytica 2016, 212, 297–311. [Google Scholar] [CrossRef]
- Roselló, M.; Royo, C.; Sanchez-Garcia, M.; Soriano, J.M. Genetic dissection of the seminal root system architecture in mediterranean durum wheat landraces by genome-wide association study. Agronomy 2019, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- HongBo, S.; ZongSuo, L.; MingAn, S. Osmotic regulation of 10 wheat (Triticum aestivum L.) genotypes at soil water deficits. Colloids Surf. B Biointerfaces 2019, 47, 132–139. [Google Scholar] [CrossRef]
- Passioura, J. The drought environment: Physical, biological and agricultural perspectives. J. Exp. Bot. 2007, 58, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Chouard, P. Vernalization and its relations to dormancy. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1960, 11, 191–238. [Google Scholar] [CrossRef]
- Hessini, K.; Martínez, J.P.; Gandour, M.; Albouchi, A.; Soltani, A.; Abdelly, C. Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water-use efficiency in Spartina alterniflora. Environ. Exp. Bot. 2009, 67, 312–319. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.M.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef]
- Hessini, K.; Ghandour, M.; Albouchi, A.; Soltani, A.; Werner, K.H.; Abdelly, C. Biomass production, photosynthesis, and leaf water relations of Spartina alterniflora under moderate water stress. J. Plant Res. 2008, 121, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, M.; Toorchi, M.; Valizadeh, M.; Shakiba, M.R. Morpho-physiological screening of sunflower inbred lines under drought stress condition. Turk. J. Field Crop. 2012, 17, 185–190. [Google Scholar]
- Ali, M.L.; Luetchens, J.; Nascimento, J.; Shaver, T.M.; Kruger, G.R.; Lorenz, A.J. Genetic variation in seminal and nodal root angle and their association with grain yield of maize under water-stressed field conditions. Plant Soil 2015, 397, 213–225. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, R.E.; Poroyko, V.; Hejlek, L.G.; Spollen, W.G.; Springer, G.K.; Bohnert, H.J. Root growth maintenance during water deficits: Physiology to functional genomics. J. Exp. Bot. 2004, 55, 2343–2351. [Google Scholar] [CrossRef] [Green Version]
- Rauf, M.; Munir, M.; ul Hassan, M.; Ahmad, M.; Afzal, M. Performance of wheat genotypes under osmotic stress at germination and early seedling growth stage. Afr. J. Biotechnol. 2007, 6, 971–975. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Violle, C.; Bourget, M.Y.; Scherer-Lorenzen, M.; Fort, F. Allocation, Morphology, Physiology, Architecture: The multiple facets of plant above- and below-ground responses to resource stress. New Phytol. 2018, 219, 1338–1352. [Google Scholar] [CrossRef]
- Brouwer, R. Nutritive influences on the distribution of dry matter in the plant. Neth. J. Agric. Sci. 1962, 10, 399–408. [Google Scholar] [CrossRef]
- Wang, T.; Huang, L.; Zhang, X.; Wang, M.; Tan, D. Root morphology and biomass allocation of 50 annual ephemerals in cold desert. Plants 2022, 11, 2495. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Palta, J.; Prasad, P.V.V.; Siddique, K.H.M. Phenotypic variability in bread wheat root systems at the early vegetative stage. BMC Plant Biol. 2020, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Westoby, M. Differences in seedling growth behaviour among species: Trait correlations across species, and trait shifts 350 along nutrient compared to rainfall gradients. J. Ecol. 2010, 87, 5–97. [Google Scholar] [CrossRef]
- Delhaize, E.; Rathyen, T.M.; Cavanagh, C.R. The genetics of rhizosheath size in a multiparent mapping population of wheat. J. Exp. Bot. 2015, 66, 4527–4536. [Google Scholar] [CrossRef] [Green Version]
- Di Marsico, A.; Scrano, L.; Amato, M.; Gàmiz, B.; Real, M.; Cox, L. Influence of mucilage from seeds of chia (Salvia hispanica L.) as an amendment, on the sorption-desorption of herbicides in agricultural soils. Sci. Tot. Environ. 2018, 625, 531–538. [Google Scholar] [CrossRef]
- Rabbi, S.M.; Tighe, M.K.; Warren, C.R.; Zhou, Y.; Denton, M.D.; Barbour, M.M.; Young, I.M. High water availability in drought tolerant crops is driven by root engineering of the soil micro-habitat. Geoderma 2021, 383, 114738. [Google Scholar] [CrossRef]
- Van Veelen, A.; Koebernick, N.; Scotson, C.S.; McKay-Fletcher, D.; Huthwelker, T.; Borca, C.N.; Mosselmans, J.F.; Roose, T. Root-induced soil deformation infuences Fe, S and P: Rhizosphere chemistry investigated using synchrotron XRF and XANES. New Phytol. 2020, 225, 1476–1490. [Google Scholar] [CrossRef]
- Marin, M.; Feeney, D.S.; Brown, L.K.; Naveed, M.; Riuz, S.; Koebernick, N.; Bengough, A.G.; Hallett, P.D.; Roose, T.; Puertolas, J. Significance of root hairs for plant performance under contrasting field conditions and water deficit. Ann. Bot. 2020, 128, 1–16. [Google Scholar] [CrossRef]
- Dodig, D.; Zorić, M.; Kandić, V.; Perović, D.; Šurlan-Momirović, G. Comparison of responses to drought stress of 100 wheat accessions and landraces to identify opportunities for improving wheat drought resistance. Plant Breed. 2012, 131, 369–379. [Google Scholar] [CrossRef]
- Friedli, C.N.; Abiven, S.; Fossati, D.; Hund, A. Modern wheat semi-dwarfs root deep on demand: Response of rooting depth to drought in a set of Swiss era wheats covering 100 years of breeding. Euphytica 2019, 215, 85. [Google Scholar] [CrossRef] [Green Version]
- Mejri, M.; Siddique, K.H.; Saif, T.; Abdelly, C.; Hessini, K. Comparative effect of drought duration on growth, photosynthesis, water relations, and solute accumulation in wild and cultivated barley species. J. Plant Nutr. Soil Sci. 2016, 179, 327–335. [Google Scholar] [CrossRef]
- Jacques, C.; Salon, C.; Barnard, R.L.; Vernoud, V.; Prudent, M. Drought stress memory at the plant cycle level: A review. Plants 2021, 10, 1873. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. Decimal code for growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bochicchio, R.; Labella, R.; Rossi, R.; Perniola, M.; Amato, M. Effects of Soil Water Shortage on Seedling Shoot and Root Growth of Saragolle Lucana Tetraploid Wheat (Triticum durum Desf.) Landrace. Plants 2022, 11, 3492. https://doi.org/10.3390/plants11243492
Bochicchio R, Labella R, Rossi R, Perniola M, Amato M. Effects of Soil Water Shortage on Seedling Shoot and Root Growth of Saragolle Lucana Tetraploid Wheat (Triticum durum Desf.) Landrace. Plants. 2022; 11(24):3492. https://doi.org/10.3390/plants11243492
Chicago/Turabian StyleBochicchio, Rocco, Rosanna Labella, Roberta Rossi, Michele Perniola, and Mariana Amato. 2022. "Effects of Soil Water Shortage on Seedling Shoot and Root Growth of Saragolle Lucana Tetraploid Wheat (Triticum durum Desf.) Landrace" Plants 11, no. 24: 3492. https://doi.org/10.3390/plants11243492