
Contrasting Responses of Alien and Ancient Forest Indicator Plant Species to Fragmentation Process in the Temperate Lowland Forests

Abstract
:1. Introduction
2. Results
2.1. Floristic Richness and Composition
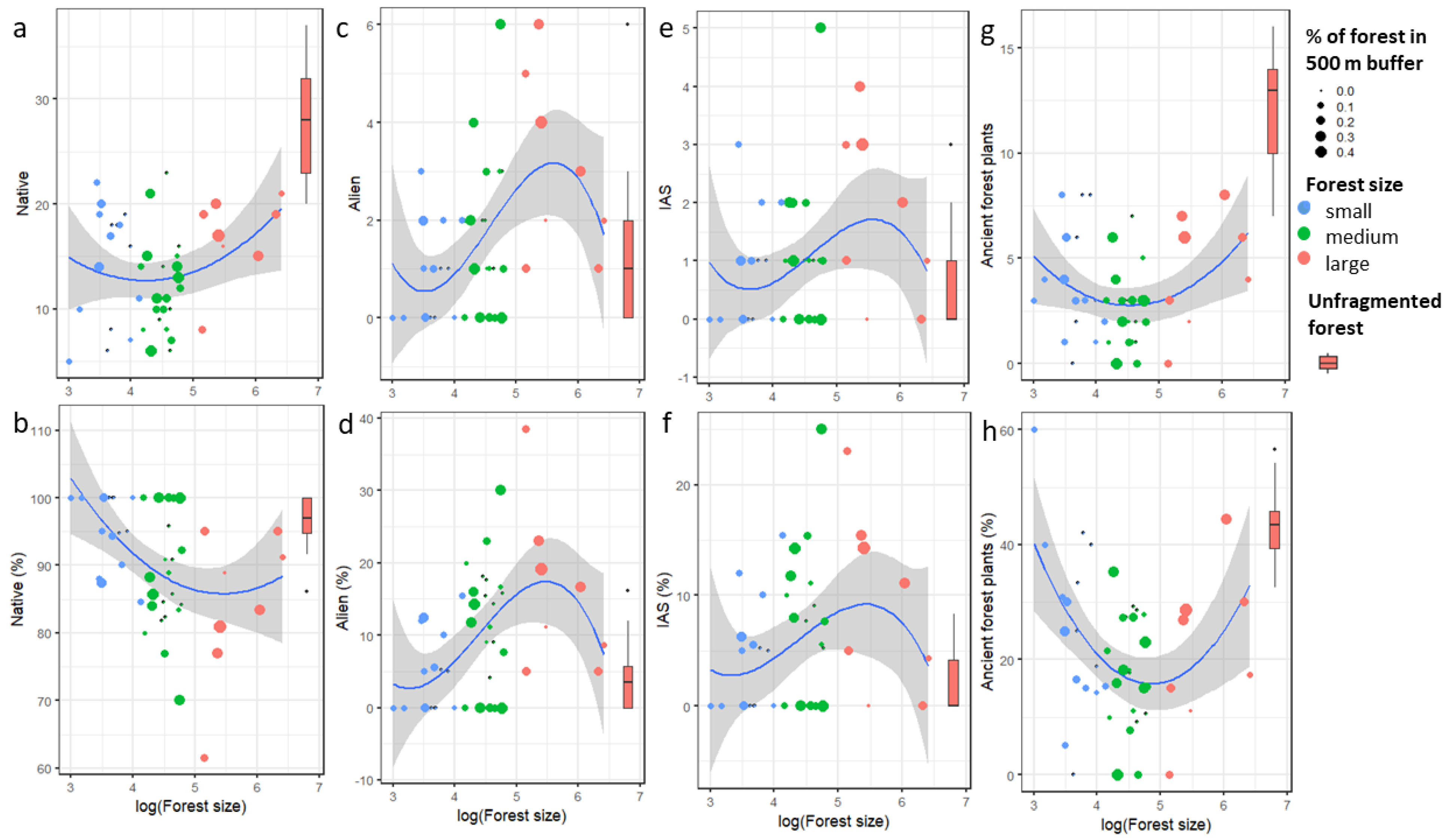
2.2. Effect of Fragment Size on Plant Species Richness
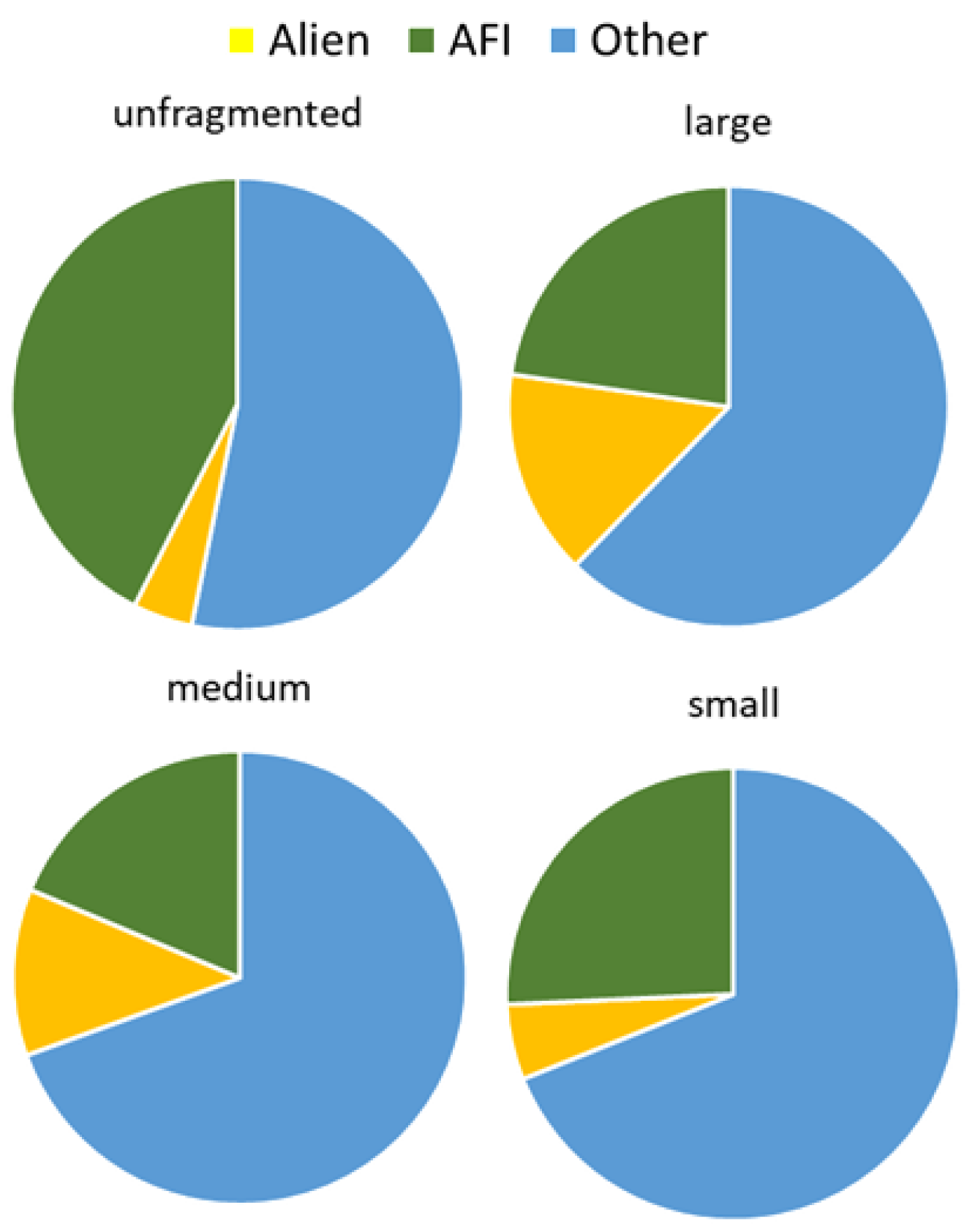
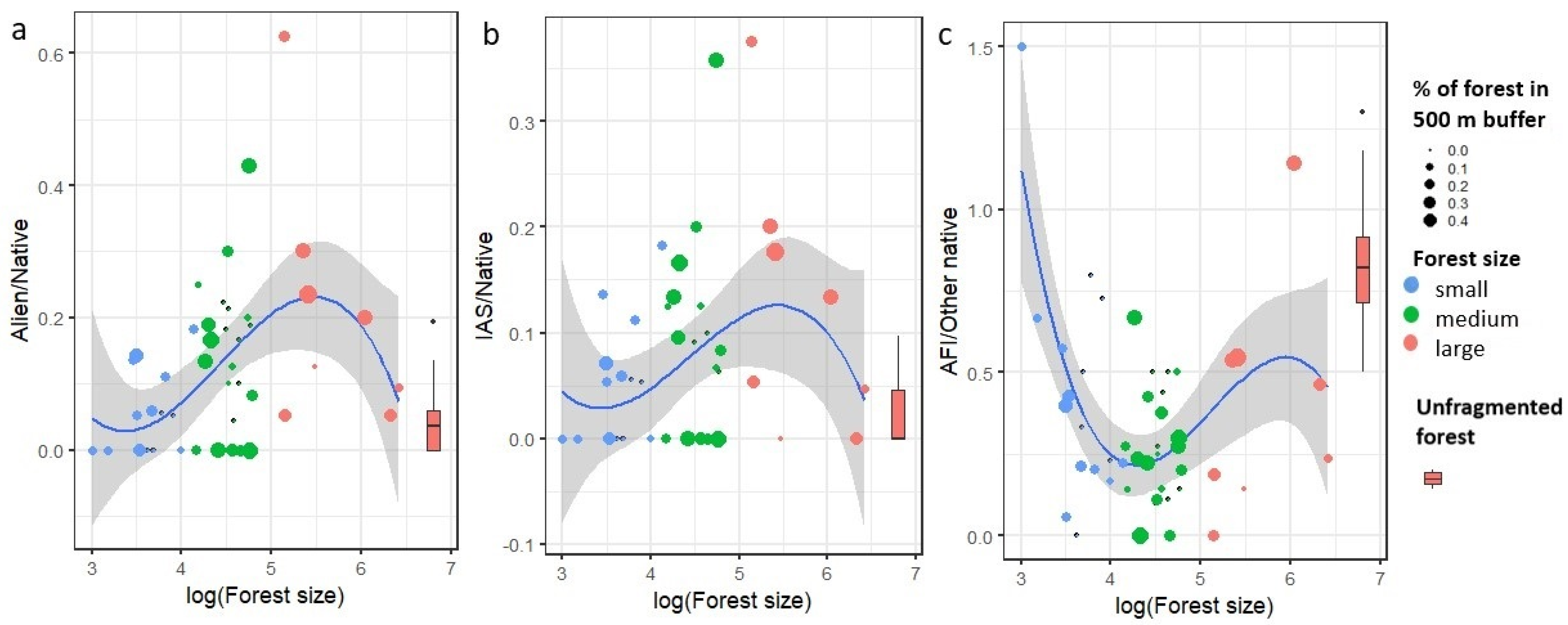
2.3. Effect of Forest Fragment Size on Plant Species Composition Quality
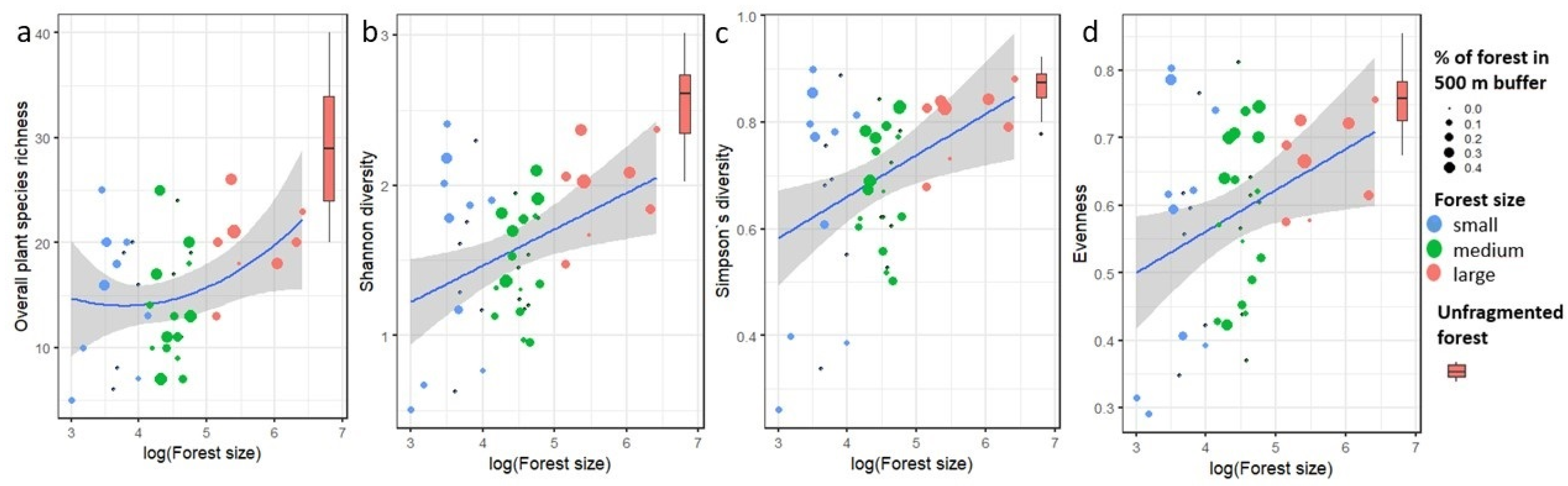
2.4. Effect of Forest Fragment Size on Diversity
3. Discussion
4. Materials and Methods
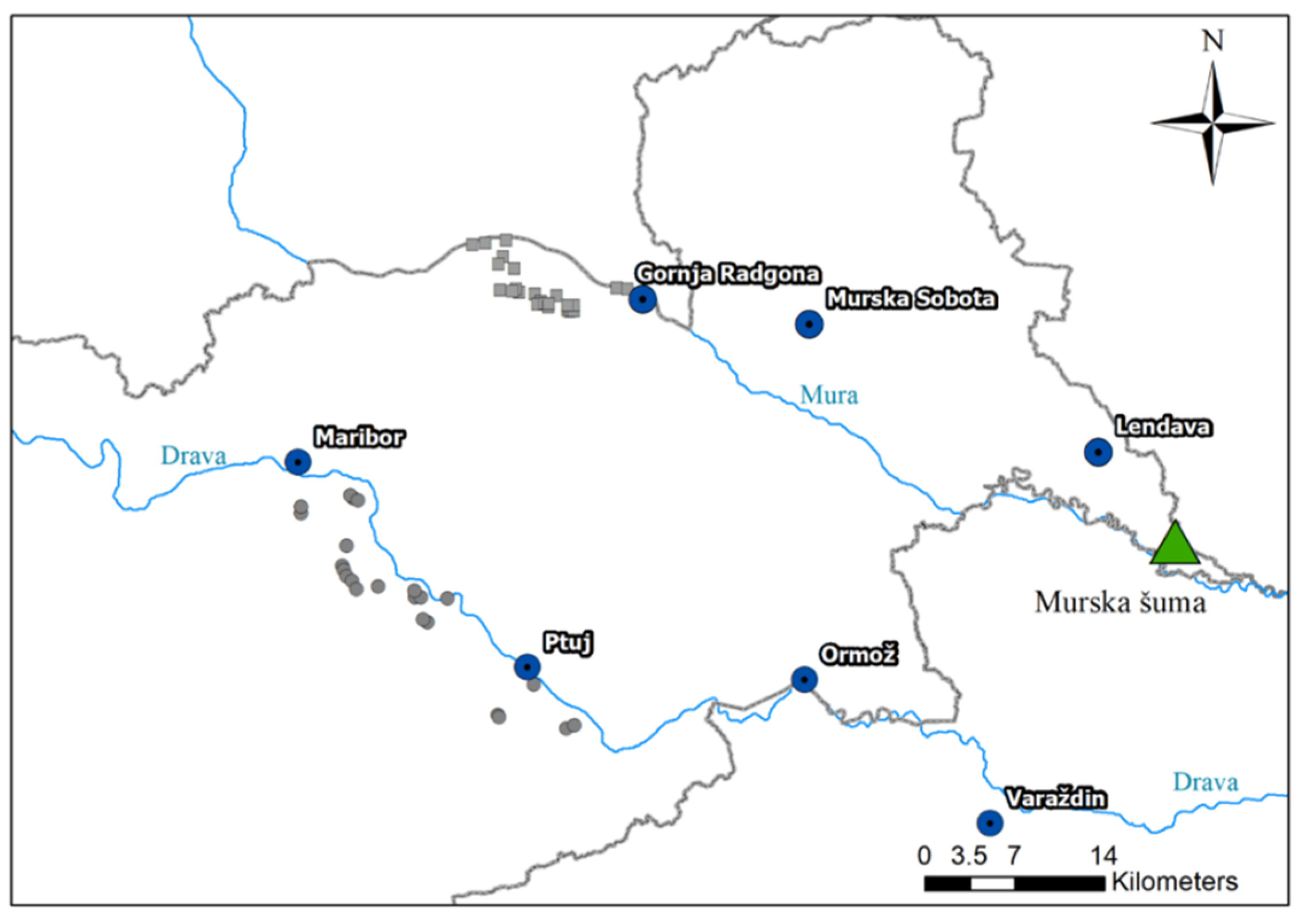
4.1. Study Area
4.2. Data Collection
4.3. Plant Species Classification
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debinski, D.M.; Holt, R.D. A survey and overview of habitat fragmentation experiments. Conserv. Biol. 2022, 14, 342–355. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Laurance, W.F. Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 2008, 141, 1731–1744. [Google Scholar] [CrossRef]
- Matthews, T.J.; Cottee-Jones, H.E.; Whittaker, R.J. Habitat Fragmentation and the Species–Area Relationship: A Focus on Total Species Richness Obscures the Impact of Habitat Loss on Habitat Specialists. Divers. Distrib. 2014, 20, 1136–1146. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Kelemen, A.; Bede, Á.; Csathó, Á.I.; Tóthmérész, B. Landscape and habitat and filters jointly drive richness and abundance of grassland specialist plants in terrestrial habitat islands. Landsc. Ecol. 2018, 33, 1117–1132. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; ISBN 9780691088365. [Google Scholar]
- Matthews, T. On the Biogeography of Habitat Islands: The Importance of Matrix Effects, Noncore Species, and Source-Sink Dynamics. Q. Rev. Biol. 2021, 96, 73–104. [Google Scholar] [CrossRef]
- Nascimento, H.; Andrade, A.; Camargo, J.; Laurance, W.F.; Laurance, S.G.; Ribeiro, J. Effects of the surrounding matrix on tree recruitment in Amazonian forest fragments. Conserv. Biol. 2006, 20, 853–860. [Google Scholar] [CrossRef]
- Zulka, K.P.; Abensperg-Traun, M.; Milasowszky, N.; Bieringer, G.; Gereben-Krenn, B.-A.; Holzinger, W.; Hölzler, G.; Rabitsch, W.; Reischütz, A.; Querner, P.; et al. Species Richness in Dry Grassland Patches of Eastern Austria: A Multi-Taxon Study on the Role of Local, Landscape and Habitat Quality Variables. Agric. Ecosyst. Environ. 2014, 182, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, C.E.; Simonetti, J.A.; Henriquez, C.A. Depressed pollination of Lapageria rosea Ruiz et Pav. (Philesiaceae) in the fragmented temperate rainforest of southern South America. Biodivers. Conserv. 2005, 15, 1845–1856. [Google Scholar] [CrossRef]
- Hauser, T.P.; Damgaard, C.; Loeschcke, V. Effects of inbreeding in small plant populations: Expectations and implications for conservation. In Conservation Genetics; EXS, Loeschcke, V., Jain, S.K., Tomiuk, J., Eds.; Birkhäuser: Basel, Switzerland, 1994; p. 68. ISBN 978-3-0348-9657-3. [Google Scholar]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, 17. [Google Scholar] [CrossRef]
- Naaf, T.; Feigs, J.T.; Huang, S.; Brunet, J.; Cousins, S.A.O.; Decocq, G.; De Frenne, P.; Diekmann, M.; Govaert, S.; Hedwall, P.-O.; et al. Sensitivity to habitat fragmentation across European landscapes in three temperate forest herbs. Landsc. Ecol. 2021, 36, 2831–2848. [Google Scholar] [CrossRef]
- Hermy, M.; Honnay, O.; Firbank, L.; Grashof-Bokdam, C.; Lawesson, J.E. An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol. Conserv. 1999, 91, 9–22. [Google Scholar] [CrossRef]
- Ohlemüller, R.; Walker, S.; Wilson, J.B. Local vs. Regional factors as determinants of the invasibility of indigenous forest fragments by alien plant species. Oikos 2006, 112, 493–501. [Google Scholar] [CrossRef]
- Vilà, M.; Ibáñez, I. Plant invasions in the landscape. Landsc. Ecol. 2011, 26, 461–472. [Google Scholar] [CrossRef]
- Mavimbela, L.Z.; Sieben, E.J.J.; Procheş, Ş. Invasive alien plant species, fragmentation and scale effects on urban forest community composition in Durban, South Africa. N. Z. J. For. 2018, 48, 19. [Google Scholar] [CrossRef] [Green Version]
- Gignac, L.D.; Dale, M.R.T. Effects of size, shape, and edge on vegetation in remnants of the upland boreal mixed-wood forest in agro-environments of Alberta, Canada. Can. J. Bot. 2007, 85, 273–284. [Google Scholar] [CrossRef]
- Šipek, M.; Šajna, N. Alien plant diversity in urban forest «Miklavški gozd». Hladnikia 2020, 46, 53–65. [Google Scholar]
- Decocq, G.; Andrieu, E.; Brunet, J.; Chabrerie, O.; De Frenne, P.; De Smedt, P.; Deconchat, M.; Diekmann, M.; Ehrmann, S.; Giffard, B.; et al. Ecosystem Services from Small Forest Patches in Agricultural Landscapes. Curr. For. 2016, 2, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Zahawi, R.A.; Oviedo-Brenes, F.; Peterson, C.J. A degradation debt? Large-scale shifts in community composition and loss of biomass in a tropical forest fragment after 40 years of isolation. PLoS ONE 2017, 12, e0183133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possley, J.; Woodmansee, S.W.; Maschinski, J. Patterns of Plant Composition in Fragments of Globally Imperiled Pine Rockland Forest: Effects of Soil Type, Recent Fire Frequency, and Fragment Size. Nat. Areas J. 2008, 28, 379–394. [Google Scholar] [CrossRef]
- Gallé, R.; Tölgyesi, C.; Császár, P.; Bátori, Z.; Gallé-Szpisjak, N.; Kaur, H.; Maák, I.; Torma, A.; Batáry, P. Landscape Structure is a Major Driver of Plant and Arthropod Diversity in Natural European Forest Fragments. Ecosphere 2022, 13, e3905. [Google Scholar] [CrossRef]
- Pisek, R. Forest Area. Available online: http://kazalci.arso.gov.si/sl/content/clone-povrsina-gozda-0 (accessed on 1 September 2019).
- Ceccherini, G.; Duveiller, G.; Grassi, G.; Lemoine, G.; Avitabile, V.; Pilli, R.; Cescatti, A. Abrupt increase in harvested forest area over Europe after 2015. Nature 2020, 583, 72–77. [Google Scholar] [CrossRef]
- Marinšek, A.; Kutnar, L. Occurrence of invasive alien plant species in the floodplain forests along the Mura River in Slovenia. Period. Biol. 2017, 119, 251–260. [Google Scholar] [CrossRef]
- Kohn, D.D.; Walsh, D.M. Plant species richness-the effect of island size and habitat diversity. J. Ecol. 1994, 82, 367–377. [Google Scholar] [CrossRef]
- Jogan, N.; Bačič, M.; Strgulc Krajšek, S. Neobiota of Slovenia: Invasive Alien Species in SLOVENIA and the Impact on the Conservation of Biodiversity and Sustainable Use of Resources. (In Slovenian: Neobiota Slovenije: Invazivne Tujerodne Vrste v Sloveniji ter Vpliv na Ohranjanje Biotske Raznovrstnosti in Trajnostno Rabo Virov). In Final Report, CRP «Konkurenčnost Slovenije 2006–2013»; University of Ljubljana (Biotechnical faculty): Ljubljana, Slovenia, 2012. [Google Scholar]
- Kutnar, L.; Kermavnar, J.; Pintar, A.M. Climate change and disturbances will shape future temperate forests in the transition zone between Central and SE Europe. Ann. For. Res. 2021, 64, 67–87. [Google Scholar] [CrossRef]
- Herrera, L.P.; Laterra, P. Relative influence of size, connectivity and disturbance history on plant species richness and assemblages in fragmented grasslands. Appl. Veg. Sci. 2011, 14, 181–188. [Google Scholar] [CrossRef]
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Gaublomme, E.; Eggermont, H.; Hendrickx, F. Local extinction processes rather than edge effects affect ground beetle assemblages from fragmented and urbanised old beech forests. Insect. Conserv. Divers. 2013, 7, 82–90. [Google Scholar] [CrossRef]
- McIntyre, N. Effects of forest patch size on avian diversity. Landsc. Ecol. 1995, 10, 85–99. [Google Scholar] [CrossRef]
- dos Anjos, L.; Bochio, G.M.; Medeiros, H.R.; de Arruda Almeida, B.; Lindsey, B.R.A.; Calsavara, L.C.; Ribeiro, M.C.; Domingues, T.; José, M. Insights on the functional composition of specialist and generalist birds throughout continuous and fragmented forests. Ecol. Evol. 2019, 9, 6318–6328. [Google Scholar] [CrossRef] [Green Version]
- Pardini, R.; de Souza, S.M.; Braga-Neto, R.; Metzger, J.P. The role of forest structure, fragment size and corridors in maintaining small mammal abundance and diversity in an Atlantic forest landscape. Biol. Conserv. 2005, 124, 253–266. [Google Scholar] [CrossRef]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.M.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Daru, B.H.; Davies, T.J.; Willis, C.G.; Meineke, E.K.; Ronk, A.; Zobel, M.; Pärtel, M.; Antonelli, A.; Davis, C.C. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 2021, 12, 698. [Google Scholar] [CrossRef]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Loinaz, G.; Amezaga, I.; Onaindia, M. Does forest fragmentation affect the same way all growth-forms? J. Environ. Manag. 2012, 94, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Honnay, O.; Jacquemyn, H.; Bossuyt, B.; Hermy, M. Forest fragmentation effects on patch occupancy and population viability of herbaceous plant species. New Phytol. 2005, 166, 723–736. [Google Scholar] [CrossRef]
- Davies, C.E.; Moss, D.; Hill, M.O. EUNIS Habitat Classification Revised 2004; European Topic Centre on Nature Protection and Biodiversity: Paris, France, 2004. [Google Scholar]
- ARSO. ARCHIVE—Observed and Measured Meteorological Data in Slovenia (In Slovenian: ARHIV—Opazovani in Merjeni meteorološki Podatki po Sloveniji). Available online: https://meteo.arso.gov.si/met/sl/archive/ (accessed on 29 January 2022).
- Ščap, D. Apaško Polje on the Path of Tolerance through History (In Slovenian: Apaško Polje po Poti Strpnosti Skozi Zgodovino; Občina Apače: Apače, Slovenia, 2018. [Google Scholar]
- Vodeb, V.; Mladenovič, L.; Nikšič, M.; Cotič, B.; Šuklje Erjavec, I.; Praper, S.; Gulič, A.; Sendi, R.; Urban Planning Institute of the Republic of Slovenia; Miklavčič, T.; et al. National Report on Urban Development—Habitat III; Ministry of the Environment and Spatial Planning, Spatial Planning, Construction and Housing Directorate: Ljubljana, Slovenia, 2016. Available online: https://www.gov.si/assets/ministrstva/MOP/Publikacije/9d3d8bd710/porocilo_urbani_razvoj_HabitatIII_en.pdf (accessed on 10 January 2022).
- Google Earth Pro. Available online: https://www.google.com/intl/sl/earth/versions/#earth-pro (accessed on 20 September 2020).
- ESRI. ArcGIS 10.2.2 for Desktop; Environmental Systems Research Institute: Redlands, CA, USA, 2014. [Google Scholar]
- EEA. Data and Maps: Copernicus Land Monitoring Service—Corine Land Cover. Available online: https://www.eea.europa.eu/data-and-maps/data/copernicus-land-monitoring-service-corine (accessed on 5 September 2018).
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer: Wien, Austria, 1964. [Google Scholar]
- Martinčič, A.; Wraber, T.; Jogan, N.; Podobnik, A.; Turk, B.; Vreš, B.; Ravnik, V.; Frajman, B.; Strgulc Krajšek, S.; Trčak, B.; et al. Small flora of Slovenia. Identification Key of Vascular Plants, 4th ed., Supplemented and Amended Edition (In Slovenian: Mala Flora Slovenije. Ključ za Določanje Praprotnic in Semenk, Četrta, Dopolnjena in Spremenjena Izdaja); Tehniška založba Slovenije: Ljubljana, Slovenia, 2007; ISBN 978-961-251-026-8. [Google Scholar]
- Tichý, L. JUICE, software for vegetation classification. J. Veg. Sci. 2002, 13, 451–453. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Wien, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 5 September 2018).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alien Plant | % in Forest Fragments | % in Unfragmented Forest | AFI | % in Forest Fragments | % in Unfragmented Forest |
|---|---|---|---|---|---|
| Acer negundo L. | 2.13 | 3.45 | Acer campestre L. | 17.02 | 100 |
| Ailanthus altissima (Mill.) Swingle | 4.26 | 0 | Asarum europaeum L. | 12.77 | 37.93 |
| Allium sp. | 2.13 | 0 | Athyrium filix-femina (L.) Roth | 0 | 37.93 |
| Ambrosia artemisiifolia L. | 0 | 3.45 | Berberis vulgaris L. | 2.13 | 0 |
| Duchesnea indica (Andrews) Focke | 4.26 | 0 | Brachypodium sylvaticum (Huds.) Beauv. | 19.15 | 51.72 |
| Erigeron annuus (L.) Pers. | 4.26 | 6.9 | Carex pallescens L. | 2.13 | 0 |
| Fraxinus americana L. | 0.00 | 6.9 | Carex pendula Huds. | 0 | 44.83 |
| Impatiens glandulifera Royle | 8.51 | 20.69 | Carex remota L. | 2.13 | 44.83 |
| Impatiens parviflora DC. | 36.17 | 3.44 | Carex sylvatica Huds. | 12.77 | 89.66 |
| Juglans nigra L. | 0 | 13.79 | Circaea lutetiana L. | 4.26 | 86.21 |
| Oxalis fontana Bunge | 4.26 | 0 | Clematis vitalba L. | 2.13 | 0 |
| Parthenocissus quinquefolia (L.) Planch. | 6.38 | 0 | Convallaria majalis L. | 2.13 | 0 |
| Phytolacca americana L. | 17.02 | 0 | Cornus mas L. | 0 | 3.45 |
| Populus x canadensis Moench | 0 | 6.9 | Cornus sanguinea L. | 19.15 | 65.52 |
| Prunus laurocerasus L. | 4.26 | 0 | Corylus avellana L. | 19.15 | 24.14 |
| Prunus serotine Ehrh. | 12.77 | 0 | Crataegus sp. | 14.89 | 44.83 |
| Quercus rubra L. | 2.13 | 0 | Dactylis glomerata subsp. lobata (Drejer) Lindb. F. | 2.13 | 0 |
| Robinia pseudoacacia L. | 34.04 | 0 | Dentaria bulbifera L. | 0 | 17.24 |
| Rudbeckia laciniata L. | 4.26 | 0 | Dryopteris carthusiana (Vill.) H.P. Fuchs | 14.89 | 0 |
| Solidago gigantea Aiton | 25.53 | 34.48 | Dryopteris filix-mas (L.) Schott | 4.26 | 6.90 |
| Taxus baccata L. | 2.13 | 0 | Euonymus europaeus L. | 31.91 | 31.03 |
| Festuca gigantea (L.) Vill. | 2.13 | 0 | |||
| Festuca heterophylla Lam. | 6.38 | 0 | |||
| Galeobdolon montanum (Pers.) Rchb. | 14.89 | 89.66 | |||
| Luzula luzuloides (Lam.) Dandy and Wilmott | 12.77 | 0 | |||
| Luzula pilosa (L.) Willd. | 4.26 | 0 | |||
| Maianthemum bifolium (L.) F.W.Schmidt | 0 | 3.45 | |||
| Melampyrum sp. | 2.13 | 0 | |||
| Milium effusum L. | 0 | 44.83 | |||
| Oxalis acetosella L. | 6.38 | 31.03 | |||
| Paris quadrifolia L. | 0 | 58.62 | |||
| Polygonatum multiflorum (L.) All. | 2.13 | 34.48 | |||
| Pteridium aquilinum (L.) Kuhn | 17.02 | 0 | |||
| Pulmonaria officinalis L. | 6.38 | 100 | |||
| Ranunculus auricomus L. | 2.13 | 0 | |||
| Rhamnus catharticus L. | 4.26 | 0 | |||
| Sanicula europaea L. | 0 | 6.90 | |||
| Scrophularia nodosa L. | 2.13 | 10.34 | |||
| Stachys sylvatica L. | 0 | 79.31 | |||
| Stellaria holostea L. | 0 | 6.90 | |||
| Stellaria nemorum L. | 0 | 13.79 | |||
| Tilia cordata Mill. | 19.15 | 0 | |||
| Tilia platyphyllos Scop. | 2.13 | 0 | |||
| Ulmus laevis Pall. | 0 | 79.31 | |||
| Ulmus minor Mill. | 14.89 | 0 | |||
| Vaccinium myrtillus L. | 8.51 | 0 | |||
| Viburnum opulus L. | 14.89 | 10.34 | |||
| Vinca minor L. | 2.13 | 0 | |||
| Viola reichenbachiana Jord. ex Boreau | 4.26 | 55.17 |
| Adjusted R2 | F-Statistic | Coefficient Estimates | Standard Error | |||
|---|---|---|---|---|---|---|
| a | b | c | ||||
| Plant Species Richness | ||||||
| Overall | 0.07 | 2.63 + | 10.66 + | 6.52 | 5.45 | |
| Native | 0.05 | 2.21 | 6.34 | 8.02 | 4.87 | |
| Alien | 0.19 | 4.58 ** | 4.32 ** | −1.50 | −2.65 + | 1.43 |
| IAS | 0.03 | 1.43 | 1.73 | −0.67 | −1.53 | 1.16 |
| AFI | 0.08 | 3.09 + | 1.27 | 5.43 * | 2.24 | |
| % Native | 0.17 | 5.79 ** | −22.87 ** | 15.79 + | 8.17 | |
| % Alien | 0.20 | 4.79 ** | 22.87 ** | −15.79 + | −12.52 | 8.04 |
| % IAS | 0.03 | 1.50 | 9.14 | −7.91 | −6.44 | 6.47 |
| % AFI | 0.19 | 6.45 ** | −13.80 | 38.92 ** | 11.49 | |
| Composition | ||||||
| Alien/Native | 0.19 | 4.63 ** | 0.30 * | −0.22 + | −0.20 + | 0.11 |
| IAS/Native | 0.05 | 1.87 | 0.13 | −0.11 | −0.11 | 0.09 |
| AFI/Other native | 0.31 | 8.02 ** | −0.24 | 0.95 ** | −0.67 ** | 0.24 |
| Diversity | ||||||
| Shannon | 0.13 | 7.64 ** | 1.24 ** | 0.45 | ||
| Simpson’s | 0.13 | 7.95 ** | 0.40 ** | 0.14 | ||
| Evenness | 0.09 | 5.67 * | 0.31 * | 0.13 | ||
| Species Richness | Native | Alien | IAS | AFI | ||||
|---|---|---|---|---|---|---|---|---|
| Average | SD | Average | SD | Average | SD | Average | SD | |
| Unfragmented | 27.90 A | 5.27 | 1.24 a | 1.41 | 0.72 a | 0.92 | 12.38 A | 2.31 |
| Large | 16.88 B | 4.12 | 3.00 b | 1.85 | 1.75 a | 1.49 | 4.50 B | 2.73 |
| Medium | 11.91 B | 4.29 | 1.61 a,b | 1.53 | 0.83 a | 1.15 | 2.52 B | 1.75 |
| Small | 14.25 B | 5.57 | 0.81 a | 0.98 | 0.75 a | 0.93 | 3.88 B | 2.60 |
| Proportion (%) | ||||||||
| Unfragmented | 96.99 a | 3.43 | 3.76 a | 4.04 | 2.10 a | 2.60 | 43.03 a | 5.99 |
| Large | 84.12 b | 11.23 | 15.88 b | 11.23 | 9.15 a | 8.20 | 21.68 b | 13.66 |
| Medium | 88.95 b | 8.52 | 11.05 b | 8.52 | 5.69 a | 6.76 | 17.55 b | 9.65 |
| Small | 95.58 a | 5.37 | 4.42 a | 5.37 | 4.03 a | 4.96 | 25.71 b | 15.34 |
| Cover (%) | ||||||||
| Unfragmented | 93.15 a | 5.40 | 4.23 a | 8.27 | 1.89 a | 2.86 | 59.37 A | 17.02 |
| Large | 65.84 b | 15.91 | 11.95 b | 7.94 | 7.34 a | 8.15 | 18.88 B | 19.37 |
| Medium | 51.00 b | 21.33 | 5.74 a | 13.70 | 2.38 a | 4.46 | 8.33 B | 15.01 |
| Small | 55.49 b | 18.78 | 6.36 a,c | 17.03 | 5.88 a | 15.86 | 16.41 B | 24.43 |
| Species Richness | Shannon | Simpson’s | Evenness | |||||
|---|---|---|---|---|---|---|---|---|
| Average | SD | Average | SD | Average | SD | Average | SD | |
| Unfragmented | 29.14 A | 6.15 | 2.54 a | 0.26 | 0.87 a | 0.03 | 0.76 a | 0.04 |
| Large | 19.88 B | 3.59 | 1.99 b | 0.29 | 0.8 b | 0.06 | 0.67 b | 0.07 |
| Medium | 13.52 C | 5.06 | 1.47 c | 0.32 | 0.68 c | 0.11 | 0.58 b | 0.12 |
| Small | 15.06 B,C | 6.1 | 1.5 b,c | 0.63 | 0.65 c,b | 0.21 | 0.55 b | 0.17 |
| Classification | Expected Forest Fragment Size Effect | Expected Effect of Forest Cover in Buffer |
|---|---|---|
| Origin (native vs. alien) plant species | Higher alien plant invasion potential in smaller fragments | The higher richness of alien plant species at lower forest cover |
| Invasive alien species (IAS) | Higher richness and proportion of IAS in smaller fragments | Higher IAS richness with decreasing forest cover |
| Ancient forest indicator plant species (AFI) | Higher richness and proportion in unfragmented forest | The increasing richness and proportion with increasing forest cover |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šipek, M.; Kutnar, L.; Marinšek, A.; Šajna, N. Contrasting Responses of Alien and Ancient Forest Indicator Plant Species to Fragmentation Process in the Temperate Lowland Forests. Plants 2022, 11, 3392. https://doi.org/10.3390/plants11233392
Šipek M, Kutnar L, Marinšek A, Šajna N. Contrasting Responses of Alien and Ancient Forest Indicator Plant Species to Fragmentation Process in the Temperate Lowland Forests. Plants. 2022; 11(23):3392. https://doi.org/10.3390/plants11233392
Chicago/Turabian StyleŠipek, Mirjana, Lado Kutnar, Aleksander Marinšek, and Nina Šajna. 2022. "Contrasting Responses of Alien and Ancient Forest Indicator Plant Species to Fragmentation Process in the Temperate Lowland Forests" Plants 11, no. 23: 3392. https://doi.org/10.3390/plants11233392