
Antioxidative and Immunomodulating Properties of Aronia melanocarpa Extract Rich in Anthocyanins
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition and Antioxidant Activity of Extracts
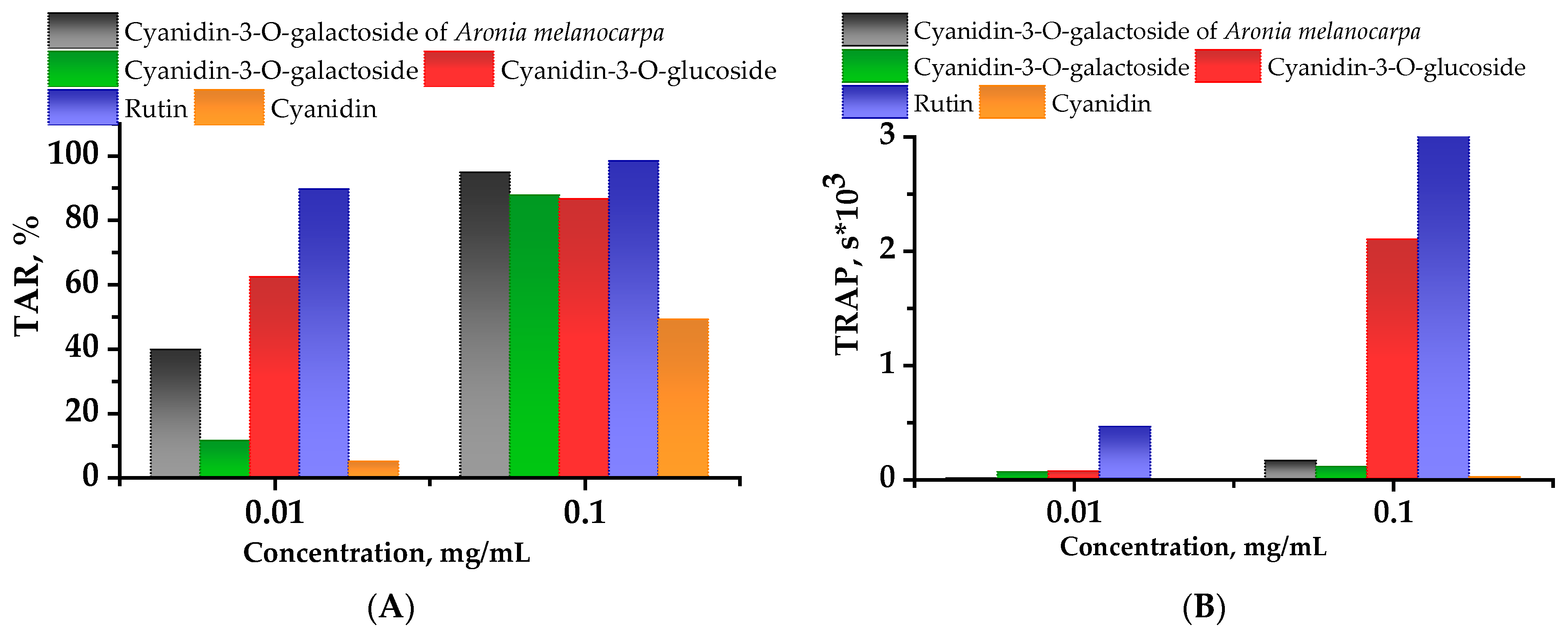
2.2. Chemiluminescent Activity of Investigated Compounds
2.3. Extracts’ Effect on DPPH-Scavenging Activity
2.4. In Vitro Tests Analyzing the Extracts’ Effect on Hemolysis of Rats
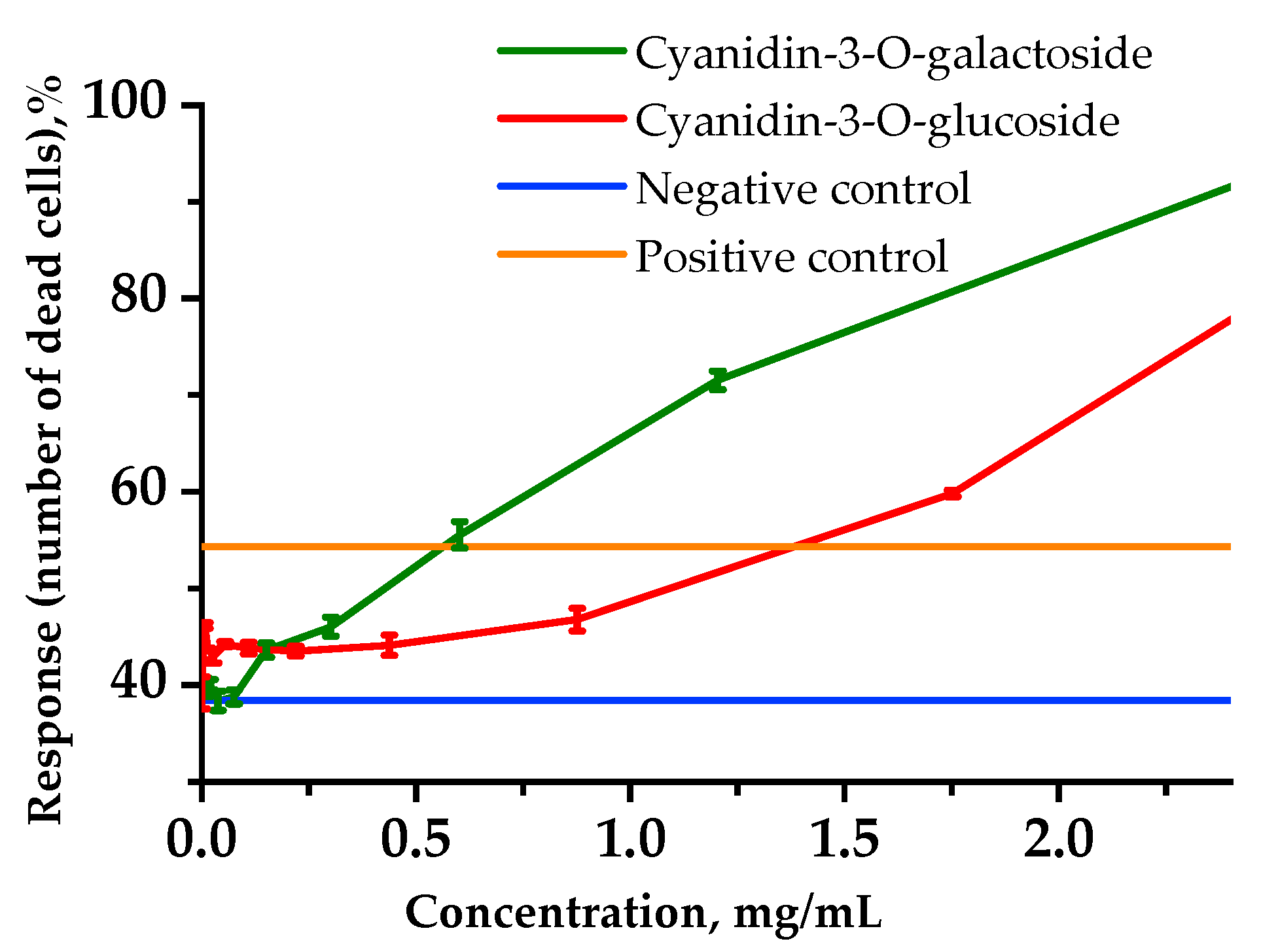
2.5. Cytoprotective Activity via In Vitro Experiments
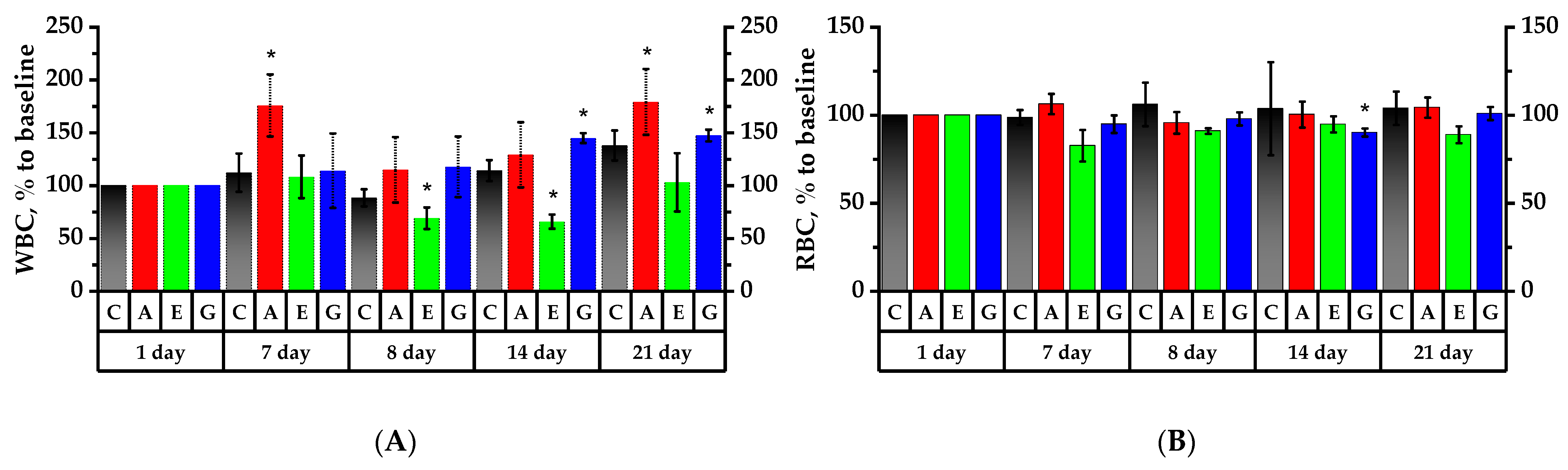
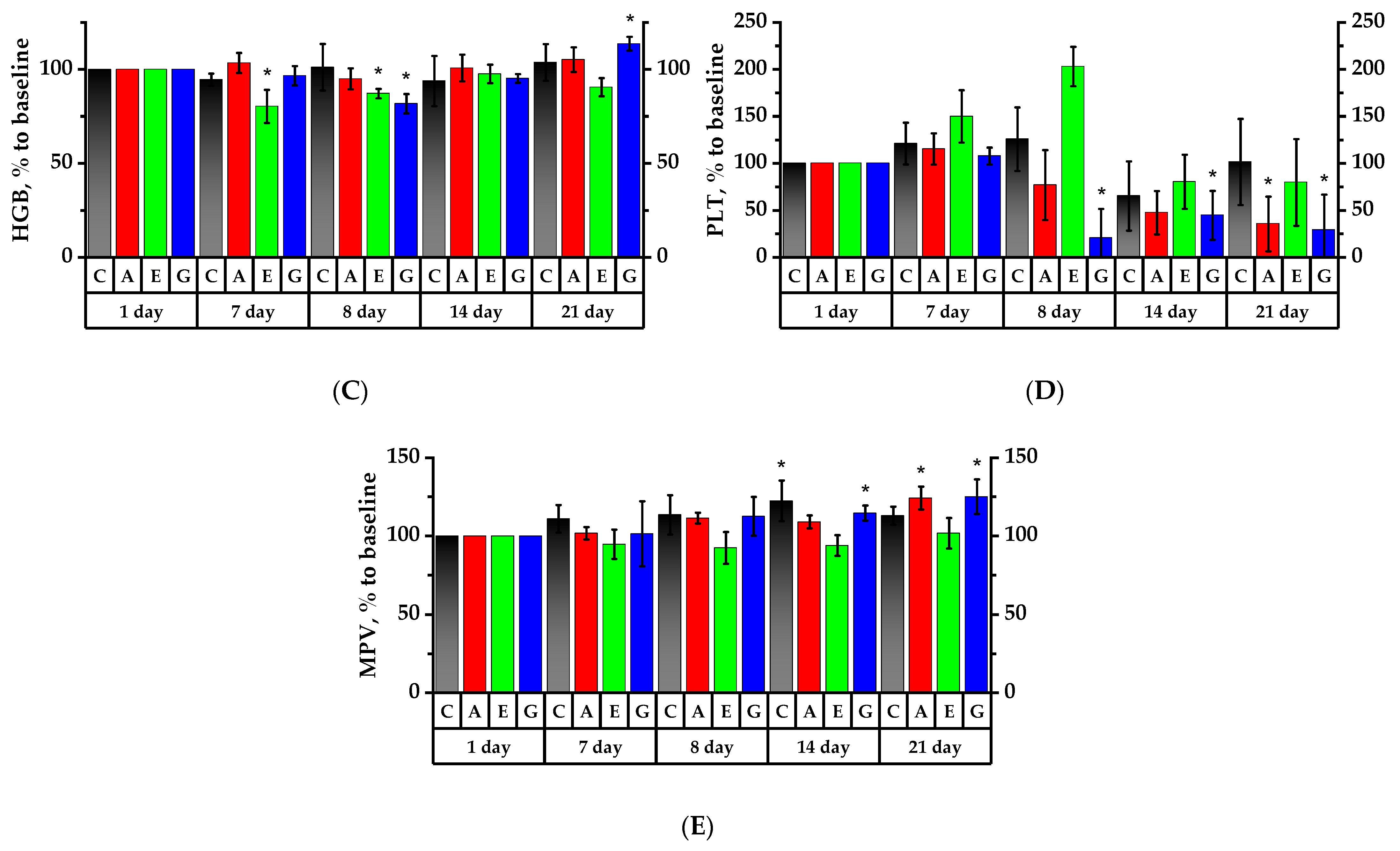
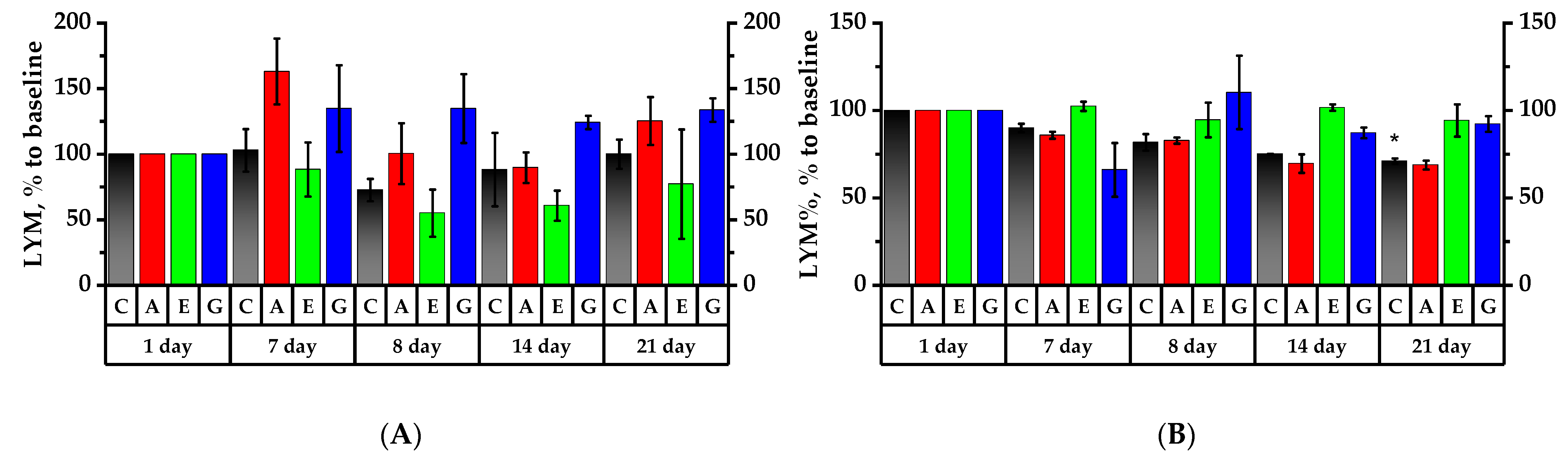
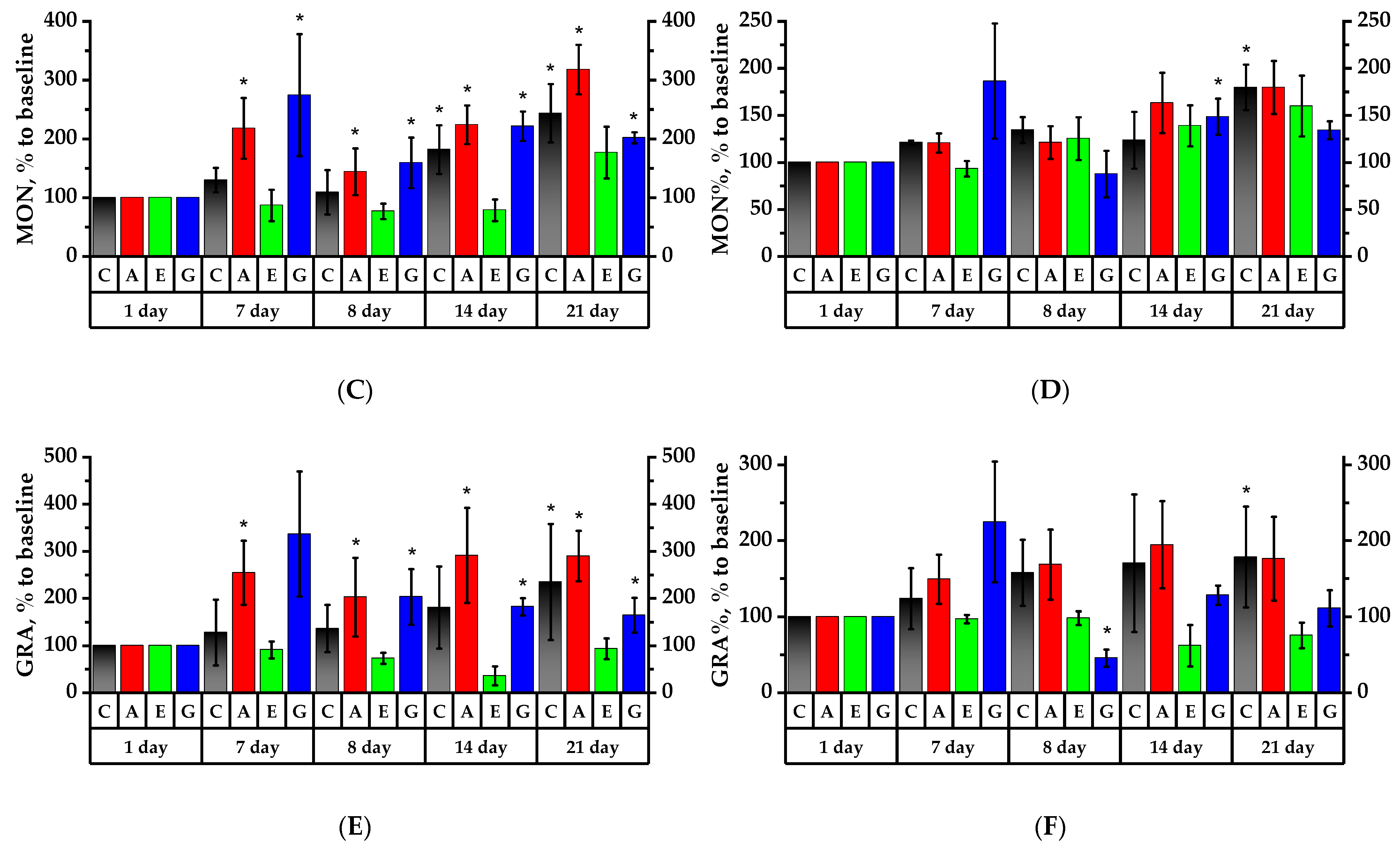
2.6. Hematology Analysis of Rats’ Peripheral Blood
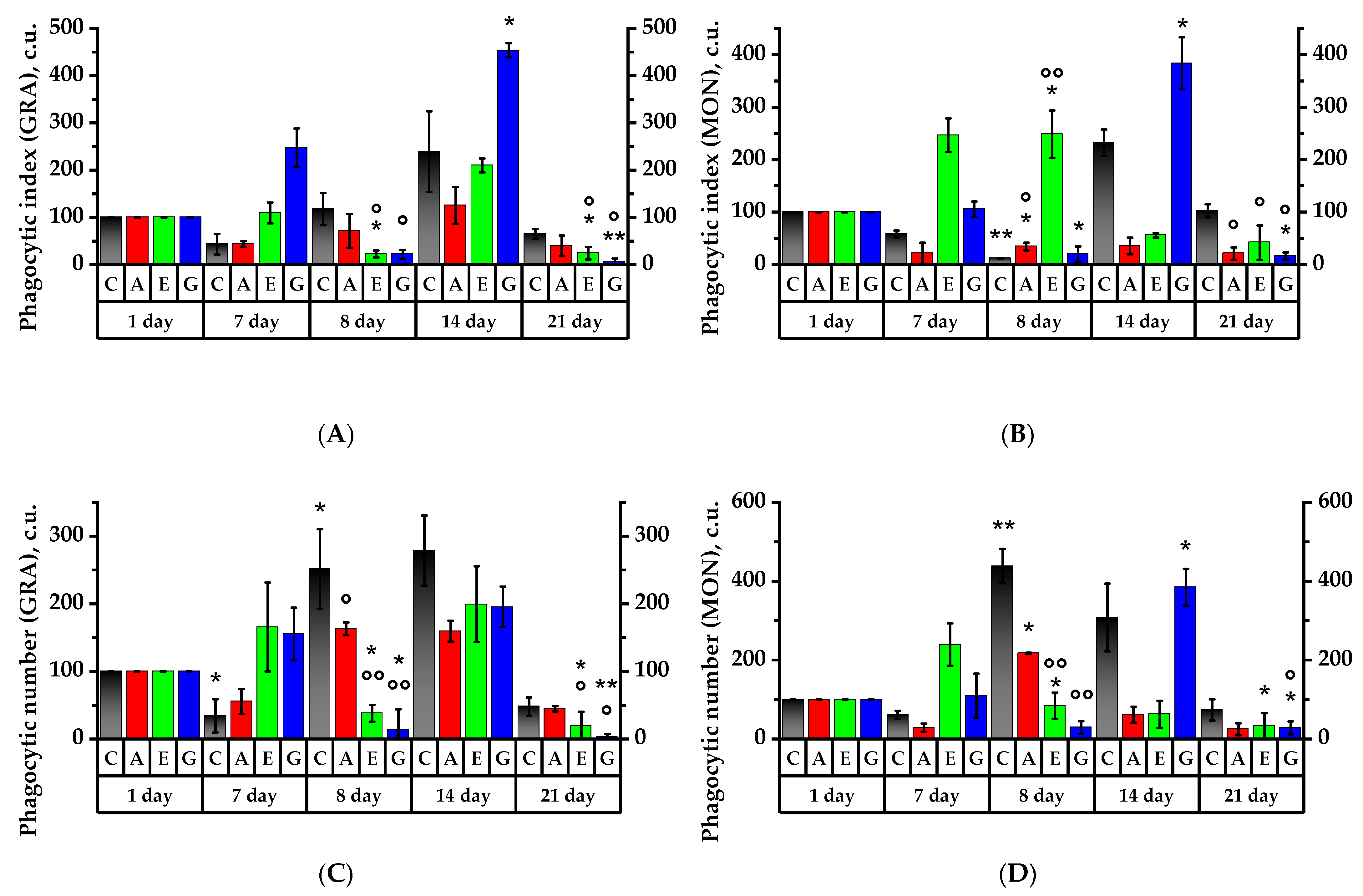
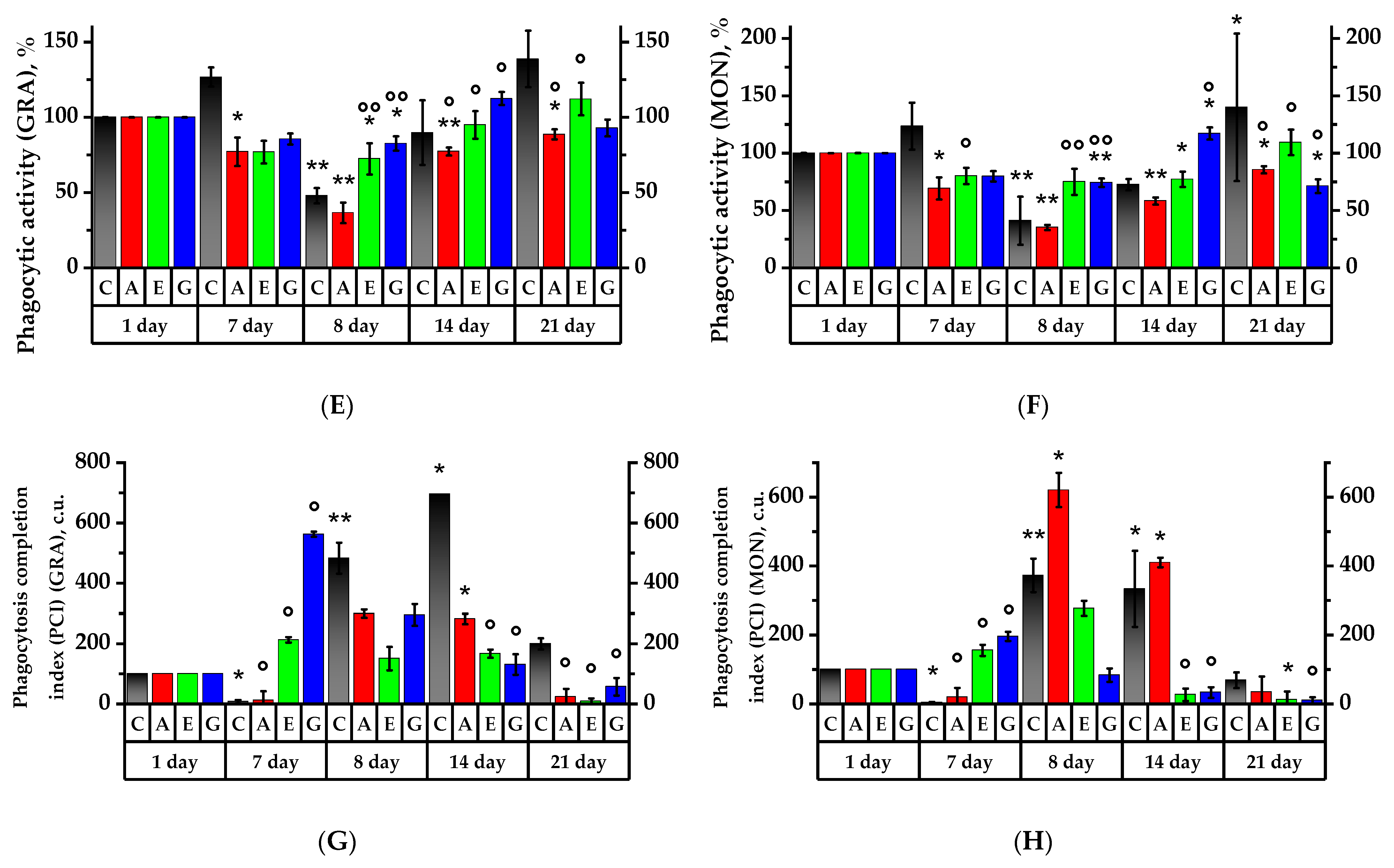
2.7. Leukocyte Phagocytic Activity Test
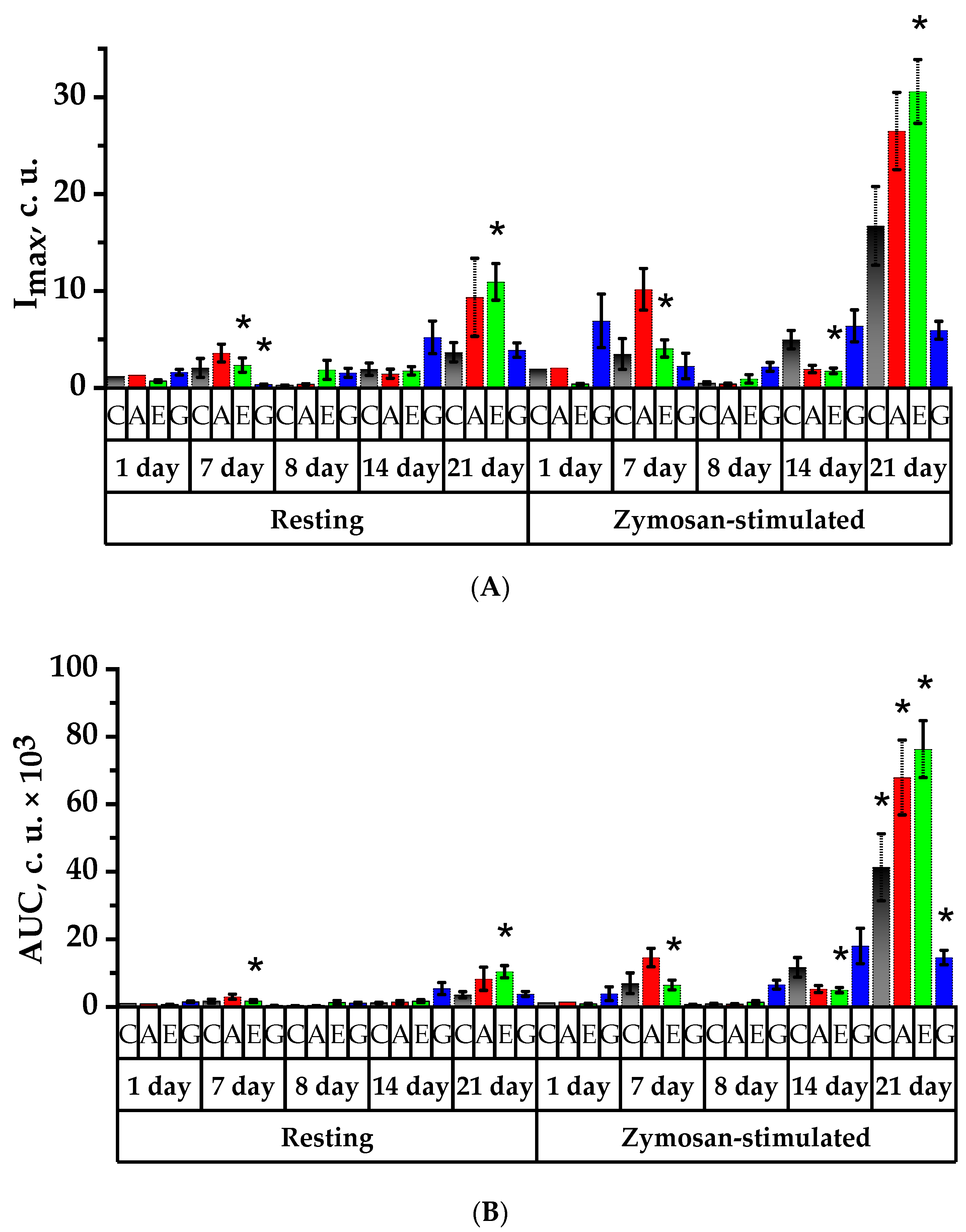
2.8. Study of Spontaneous and Activated Neutrophil Chemiluminescent Activity
2.9. Lipid Peroxidation Study
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Aronia Extract Preparation
4.3. Anthocyanin Extract Preparation
4.4. Chemical Composition Analysis
4.5. Chromatographic Analysis of Anthocyanin Extract
4.6. Chemicals and Reagents
4.7. Antioxidant Activity and Biologically Active Substance Contents in Aronia Extracts
4.8. Rat Erythrocyte In Vitro Tests for Extracts’ Protective Activity on Osmotic and Peroxide Hemolysis Models
4.9. In Vitro Tests on the Lymphocyte Cell Line RPMI-1788
4.9.1. Determination of Cytoprotective Properties In Vitro
4.9.2. Cell Color Staining and Counting
4.10. In Vivo Tests
4.10.1. Hematological Studies
4.10.2. Functional State of Neutrophilic Granulocytes in Rats’ Blood
4.10.3. Lipid Peroxidation in Rats’ Blood
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Oszmiański, J.; Wojdylo, A. Aronia melanocarpa phenolics and their antioxidant activity. Eur. Food Res. Technol. 2005, 221, 809–813. [Google Scholar] [CrossRef]
- Denev, P.; Číž, M.; Kratchanova, M.; Blazheva, D. Black chokeberry (Aronia melanocarpa) polyphenols reveal different antioxidant, antimicrobial and neutrophil-modulating activities. Food Chem. 2019, 284, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Oszmiański, J.; Lachowicz, S. Effect of the production of dried fruits and juice from chokeberry (Aronia melanocarpa L.) on the content and antioxidative activity of bioactive compounds. Molecules 2016, 21, 1098. [Google Scholar] [CrossRef] [PubMed]
- Sidor, A.; Drożdżyńska, A.; Gramza-Michałowska, A. Black chokeberry (Aronia melanocarpa) and its products as potential health-promoting factors-An overview. Trends Food Sci. Technol. 2019, 89, 45–60. [Google Scholar] [CrossRef]
- Veberic, R.; Slatnar, A.; Bizjak, J.; Stampar, F.; Mikulic-Petkovsek, M. Anthocyanin composition of different wild and cultivated berry species. LWT-Food Sci. Technol. 2015, 60, 509–517. [Google Scholar] [CrossRef]
- Strugała, P.; Gładkowski, W.; Kucharska, A.Z.; Sokół-Łętowska, A.; Gabrielska, J. Antioxidant activity and anti-inflammatory effect of fruit extracts from blackcurrant, chokeberry, hawthorn, and rosehip, and their mixture with linseed oil on a model lipid membrane. Eur. J. Lipid Sci. Technol. 2016, 118, 461–474. [Google Scholar] [CrossRef]
- Kulling, S.E.; Rawel, H.M. Chokeberry (Aronia melanocarpa)–A review on the characteristic components and potential health effects. Planta Med. 2008, 74, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Wang, S.Y. Oxygen radical absorbing capacity of phenolics in blueberries, cranberries, chokeberries, and lingonberries. J. Agric. Food Chem. 2003, 51, 502–509. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Giampieri, F.; Tulipani, S.; Casoli, T.; Di Stefano, G.; González-Paramás, A.M.; Santos-Buelga, C.; Busco, F.; Quiles, J.L.; Cordero, M.D. One-month strawberry-rich anthocyanin supplementation ameliorates cardiovascular risk, oxidative stress markers and platelet activation in humans. J. Nutr. Biochem. 2014, 25, 289–294. [Google Scholar] [CrossRef]
- Jiao, X.; Li, B.; Zhang, Q.; Gao, N.; Zhang, X.; Meng, X. Effect of in vitro-simulated gastrointestinal digestion on the stability and antioxidant activity of blueberry polyphenols and their cellular antioxidant activity towards HepG2 cells. Int. J. Food Sci. Technol. 2018, 53, 61–71. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Pan, P.; Echeveste, C.E.; Wang, H.T.; Oshima, K.; Lin, C.W.; Yearsley, M.; Xiao, J.; Chen, J.; Sun, C. Transplanting fecal material from wild-type mice fed black raspberries alters the immune system of recipient mice. Food Front. 2020, 1, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, A.; Dzierżanowski, T. Targeting inflammation by anthocyanins as the novel therapeutic potential for chronic diseases: An update. Molecules 2021, 26, 4380. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Tian, H.; Liu, C.; Zhang, X.; Peng, Y.; Yang, X.; Chen, F.; Li, J. Metformin and cyanidin 3-O-galactoside from Aronia melanocarpa synergistically alleviate cognitive impairment in SAMP8 mice. Food Funct. 2021, 12, 10994–11008. [Google Scholar] [CrossRef] [PubMed]
- Victoria-Campos, C.I.; de Jesús Ornelas-Paz, J.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Failla, M.L.; Pérez-Martínez, J.D.; Rios-Velasco, C.; Ibarra-Junquera, V. Gastrointestinal metabolism and bioaccessibility of selected anthocyanins isolated from commonly consumed fruits. Food Chem. 2022, 383, 132451. [Google Scholar] [CrossRef] [PubMed]
- Bushmeleva, K.; Vyshtakalyuk, A.; Terenzhev, D.; Belov, T.; Parfenov, A.; Sharonova, N.; Nikitin, E.; Zobov, V. Radical scavenging actions and immunomodulatory activity of Aronia melanocarpa propylene glycol extracts. Plants 2021, 10, 2458. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhao, L.; Cha, Q.; Sun, Y.; Ye, H.; Zeng, X. Structural characterization and immunostimulatory activity of a homogeneous polysaccharide from Sinonovacula constricta. J. Agric. Food Chem. 2015, 63, 7986–7994. [Google Scholar] [CrossRef]
- Brand, M.H.; Connolly, B.A.; Levine, L.H.; Richards, J.T.; Shine, S.M.; Spencer, L.E. Anthocyanins, total phenolics, ORAC and moisture content of wild and cultivated dark-fruited Aronia species. Sci. Hortic. 2017, 224, 332–342. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, J.; Meng, X. Aronia melanocarpa anthocyanin extracts are an effective regulator of suppressor of cytokine signaling 3-dependent insulin resistance in HepG2 and C2C12 cells. J. Funct. Foods 2020, 75, 104258. [Google Scholar] [CrossRef]
- Tsuda, T.; Horio, F.; Osawa, T. Dietary cyanidin 3-O-β-d-glucoside increases ex vivo oxidation resistance of serum in rats. Lipids 1998, 33, 583. [Google Scholar] [CrossRef]
- Matsumoto, H.; Nakamura, Y.; Hirayama, M.; Yoshiki, Y.; Okubo, K. Antioxidant activity of black currant anthocyanin aglycons and their glycosides measured by chemiluminescence in a neutral pH region and in human plasma. J. Agric. Food Chem. 2002, 50, 5034–5037. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Mi, Y.; Cao, H.; Chen, L. Enzymatic acylation of raspberry anthocyanin: Evaluations on its stability and oxidative stress prevention. Food Chem. 2022, 372, 130766. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Deng, Y.; Dai, R.; Yu, Y.; Saeed, M.K.; Li, L.; Meng, W.; Zhang, X. Chromatographic fingerprint analysis of Cephalotaxus sinensis from various sources by high-performance liquid chromatography–diodearray detection–electrospray ionization-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2007, 45, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Dudek, A.; Spiegel, M.; Strugała-Danak, P.; Gabrielska, J. Analytical and Theoretical Studies of Antioxidant Properties of Chosen Anthocyanins; A Structure-Dependent Relationships. Int. J. Mol. Sci. 2022, 23, 5432. [Google Scholar] [CrossRef]
- Youdim, K.A.; Martin, A.; Joseph, J.A. Incorporation of the elderberry anthocyanins by endothelial cells increases protection against oxidative stress. Free Radic. Biol. Med. 2000, 29, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Forester, S.C.; Choy, Y.Y.; Waterhouse, A.L.; Oteiza, P.I. The anthocyanin metabolites gallic acid, 3-O-methylgallic acid, and 2, 4, 6-trihydroxybenzaldehyde decrease human colon cancer cell viability by regulating pro-oncogenic signals. Mol. Carcinog. 2014, 53, 432–439. [Google Scholar] [CrossRef]
- Vadhan-Raj, S. Management of chemotherapy-induced thrombocytopenia: Current status of thrombopoietic agents. In Seminars in Hematology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 46, pp. S26–S32. [Google Scholar]
- Dygai, A.M.; Zhdanov, V.V. The theory of hemopoiesis regulation in health and disease. Bull. Sib. Med. 2004, 3, 5–14. [Google Scholar] [CrossRef]
- Chen, J.-R.; Yang, Z.-Q.; Hu, T.-J.; Yan, Z.-T.; Niu, T.-X.; Wang, L.; Cui, D.-A.; Wang, M. Immunomodulatory activity in vitro and in vivo of polysaccharide from Potentilla anserina. Fitoterapia 2010, 81, 1117–1124. [Google Scholar] [CrossRef]
- Shah, A.S.; Juvekar, A.R. In vitro and in vivo immunostimulatory activity of Woodfordia fruticosa flowers on non-specific immunity. Pharm. Biol. 2010, 48, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Mahmoud, A.; Sharoud, M. Potential immunological benefits of Echinacea. J. Radiat. Res. Appl. Sci. 2009, 2, 1078–1090. [Google Scholar]
- Goel, V.; Chang, C.; Slama, J.V.; Barton, R.; Bauer, R.; Gahler, R.; Basu, T.K. Echinacea stimulates macrophage function in the lung and spleen of normal rats. J. Nutr. Biochem. 2002, 13, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Murthy, H.N.; Park, S.-Y.; Lee, E.J.; Paek, K.Y. Biosafety and toxicological evaluation of tissue-cultured Echinacea purpurea adventitious roots. Hortic. Sci. Technol. 2015, 33, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Bucki, R.; Pastore, J.; Giraud, F.; Sulpice, J.C.; Janmey, P. Flavonoid inhibition of platelet procoagulant activity and phosphoinositide synthesis. J. Thromb. Haemost. 2003, 1, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Mayansky, D.N. Lectures on Clinical Pathology: A Guide for Doctors; GEOTAR-Media: Moscow, Russia, 2008; p. 462. [Google Scholar]
- Matsumoto, T.; Kaneko, A.; Koseki, J.; Matsubara, Y.; Aiba, S.; Yamasaki, K. Pharmacokinetic study of bioactive flavonoids in the traditional Japanese medicine Keigairengyoto exerting antibacterial effects against Staphylococcus aureus. Int. J. Mol. Sci. 2018, 19, 328. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Qin, P.; Ren, G. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. Japonica) on chronically alcohol-induced liver damage in rats. J. Agric. Food Chem. 2010, 58, 3191–3196. [Google Scholar] [CrossRef]
- Herawati, E.R.N.; Santosa, U.; Sentana, S.; Ariani, D. Protective effects of anthocyanin extract from purple sweet potato (Ipomoea batatas L.) on blood MDA levels, liver and renal activity, and blood pressure of hyperglycemic rats. Prev. Nutr. Food Sci. 2020, 25, 375. [Google Scholar] [CrossRef]
- Fallah, A.A.; Sarmast, E.; Jafari, T. Effect of dietary anthocyanins on biomarkers of oxidative stress and antioxidative capacity: A systematic review and meta-analysis of randomized controlled trials. J. Funct. Foods 2020, 68, 103912. [Google Scholar] [CrossRef]
- Li, W.-G.; Zhang, X.; Wu, Y.; Tian, X. Anti-inflammatory effect and mechanism of proanthocyanidins from grape seeds. Acta Pharmacol. Sin. 2001, 22, 1117–1120. [Google Scholar]
- Colombo, F.; Di Lorenzo, C.; Regazzoni, L.; Fumagalli, M.; Sangiovanni, E.; de Sousa, L.P.; Bavaresco, L.; Tomasi, D.; Bosso, A.; Aldini, G. Phenolic profiles and anti-inflammatory activities of sixteen table grape (Vitis vinifera L.) varieties. Food Funct. 2019, 10, 1797–1807. [Google Scholar] [CrossRef]
- Rodríguez-Morgado, B.; Candiracci, M.; Santa-María, C.; Revilla, E.; Gordillo, B.; Parrado, J.; Castaño, A. Obtaining from grape pomace an enzymatic extract with anti-inflammatory properties. Plant Foods Hum. Nutr. 2015, 70, 42–49. [Google Scholar] [CrossRef]
- Hariri, B.M.; Payne, S.J.; Chen, B.; Mansfield, C.; Doghramji, L.J.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Niv, M.Y.; Lee, R.J. In vitro effects of anthocyanidins on sinonasal epithelial nitric oxide production and bacterial physiology. Am. J. Rhinol. Allergy 2016, 30, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platonova, E.Y.; Shaposhnikov, M.V.; Lee, H.-Y.; Lee, J.-H.; Min, K.-J.; Moskalev, A. Black chokeberry (Aronia melanocarpa) extracts in terms of geroprotector criteria. Trends Food Sci. Technol. 2021, 114, 570–584. [Google Scholar] [CrossRef]
- Watrelot, A.A.; Bouska, L. Optimization of the ultrasound-assisted extraction of polyphenols from Aronia and grapes. Food Chem. 2022, 386, 132703. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Zhu, J.; Ma, Y.; Sun, X.; Li, D.; Li, L.; Bai, H.; Xin, G.; Meng, X. Composition and antioxidant activity of anthocyanins from Aronia melanocarpa cultivated in Haicheng, Liaoning, China. Food Biosci. 2019, 30, 100413. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Sandate-Flores, L.; Rodríguez-Rodríguez, J.; Velázquez, G.; Mayolo-Deloisa, K.; Rito-Palomares, M.; Torres, J.A.; Parra-Saldívar, R. Low-sugar content betaxanthins extracts from yellow pitaya (Stenocereus pruinosus). Food Bioprod. Process. 2020, 121, 178–185. [Google Scholar] [CrossRef]
- Stankovic, M.S. Total phenolic content, flavonoid concentration and antioxidant activity of Marrubium peregrinum L. extracts. Kragujevac J. Sci. 2011, 33, 63–72. [Google Scholar]
- Jafarova, E.E.; Mustafayeva, L.A.; Serkerov, S.V. Composition and content of anthocyanins of the Polygonum aviculare L. flowers. Khimija Rastit. Syr’ja 2016, 2, 103–107. [Google Scholar]
- Lubsandorzhieva, P.B.; Baldanova, N.B.; Popov, D.V. The quantitative analysis of flavonoids in vegetable remedy by HPLC method. Sib. Med. J. 2013, 116, 114–115. [Google Scholar]
- Tynyanaya, I.I. Separation, Concentration and Analysis of Anthocyanins and Betacyanins in Plant Extracts using Optical and Chromatographic Methods. Ph.D. Thesis, Belgorod State University, Belgorod, Russia, 2015; p. 147. [Google Scholar]
- Krasowska, A.; Rosiak, D.; Szkapiak, K.; Lukaszewicz, M. Chemiluminescence detection of peroxyl radicals and comparison of antioxidant activity of phenolic compounds. Curr. Top. Biophys. 2000, 24, 89–95. [Google Scholar]
- Vyshtakalyuk, A.B.; Semenov, V.E.; Sudakov, I.A.; Bushmeleva, K.N.; Gumarova, L.F.; Parfenov, A.A.; Nazarov, N.G.; Galyametdinova, I.V.; Zobov, V.V. Xymedon conjugate with biogenic acids. Antioxidant properties of a conjugate of Xymedon with L-ascorbic acid. Russ. Chem. Bull. 2018, 67, 705–711. [Google Scholar] [CrossRef]
- Lissi, E.; Salim-Hanna, M.; Pascual, C.; del Castillo, M.D. Evaluation of total antioxidant potential (TRAP) and total antioxidant reactivity from luminol-enhanced chemiluminescence measurements. Free Radic. Biol. Med. 1995, 18, 153–158. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sowemimo-Coker, S.O. Red blood cell hemolysis during processing. Transfus. Med. Rev. 2002, 16, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Bushmeleva, K.; Vyshtakalyuk, A.; Terenzhev, D.; Nikitin, E. Evaluation of seasonal differences in the antioxidant activity of needle juices of Picea abies L. and Pinus sylvestris L. with luminol-enhanced chemiluminescence. Indian J. Agric. Res. 2021, 55, 265–272. [Google Scholar] [CrossRef]
- Freshney, R.I. Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Mironov, A. A Guide to Preclinical Drug Research; Grif and K: Moscow, Russia, 2012. [Google Scholar]
- Oliferuk, N.S. Optimization of Methods for Evaluation of Phagocytic Activity of Peripheral Blood Leukocytes using Laser Flow Cytometry. Ph.D. Thesis, NRC Institute of Immunology FMBA of Russia, Moscow, Russia, 24 September 2008. [Google Scholar]
- Miliukene, V.; Bizulyavicienė, G.Y.; Khaustova, L.P.; Pilinkene, A.V.; Bizulyavichyus, G.A. Quantification of Escherichia coli phagocytosis by mouse peritoneal macrophages. Cytology 2007, 49, 853–857. [Google Scholar]
- Muniz-Junqueira, M.I.; Peçanha, L.M.F.; da Silva-Filho, V.L.; de Almeida Cardoso, M.C.; Tosta, C.E. Novel microtechnique for assessment of postnatal maturation of the phagocytic function of neutrophils and monocytes. Clin. Vaccine Immunol. 2003, 10, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Komine, K.i.; Komine, Y.; Kuroishi, T.; Kozutsumi, T.; Kobayashi, J.; Ohta, M.; Kitamura, H.; Kumagai, K. Lactoferrin stimulates a Staphylococcus aureus killing activity of bovine phagocytes in the mammary gland. Microbiol. Immunol. 2002, 46, 187–194. [Google Scholar] [CrossRef]
- Klimova, E.M.; Drozdova, L.A.; Lavinskaya, E.V.; Bychenko, E.A. Integral methodology of I.I. Mechnikov and modern targeted immunocorrection in myasthenia gravis. Ann. Mechnikov Inst. 2015, 2, 30–37. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1978; Volume 52, pp. 302–310. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | pH | Total Sugars 1, mg Xy/g Extract | Total Flavonoids 2, mg Rut/g Extract | Total Anthocyanins, mg/g Extract |
|---|---|---|---|---|
| Aronia ethanol extract | 7.27 | 63.65 | 37.50 | 93.60 |
| Cy-3-Gal Aronia | 3.93 | 0.125 | 15.35 | 917.31 |
| Sample | Cyanidin-3-O-galactoside | Cyanidin-3-O-glucoside | Cyanidin-3-O-arabinozide | Cyanidin-3-O-xyloside |
|---|---|---|---|---|
| mg/g ext. | ||||
| Aronia ethanol extract | 58.97 | 6.21 | 18.15 | 8.89 |
| Cy-3-Gal Aronia | 735.33 | 12.58 | 115.22 | 31.33 |
| Fraction | Output Relative to Anthocyanin Fraction | Major Molecular Mass Peak, [M-H] | Anthocyanins | ||||
|---|---|---|---|---|---|---|---|
| Cyanidin-3-O-galactoside | Cyanidin-3-O-glucoside | Cyanidin-3-O-arabinozide | Cyanidin-3-O-xyloside | ||||
| % | Anth. | Anth.HCl− | % | % | % | % | |
| 6–9 | 81.7 | 447 | 484 | 93.7 | 0.15 | 2.1 | 0.49 |
| 13–15 | 3.1 | 447 | 484 | 1.1 | 97.8 | 0.7 | 0.3 |
| 20–22 | 11.3 | 417 | 453 | 0.6 | 0.3 | 94.9 | 0.4 |
| 26–2 | 3.5 | 417 | 453 | 0.5 | 0.2 | 3.7 | 93.9 |
| Sample | Individual Form, mg/g Dry ext. | Glycosidic Form, mg/g Dry ext. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Quercetin | Kaempfe-rol | Rhamnetin | Isorhamnetin | Dihydroquercetin | Quercetin-3-O-rutinoside | Hesperetin 7-O-rutinoside | Quercetin-3-O-rhamnoside | Quercetin-3-O-galactoside | Quercetin-3-O-glucoside | Dihydroquercetin hexoside | |
| Aronia ethanol extract | 12.35 | 0.11 | 0.63 | - | 2.17 | 15.35 | 0.97 | 0.93 | 0.18 | 0.49 | 0.27 |
| Cy-3-Gal Aronia | 3.235 | 0.029 | 0.371 | 0.023 | 1.153 | 7.973 | 0.69 | 0.067 | 0.036 | 0.075 | 0.115 |
| Sample | EC50 1, mg/mL | ARP 2, mg/mL−1 |
|---|---|---|
| Cyanidin-3-O-galactoside | 0.221 | 4.534 |
| Cyanidin-3-O-glucoside | 0.169 | 5.909 |
| Cy-3-Gal Aronia | 0.154 | 6.484 |
| Quercetin-3-O-rutinoside (Rutin) | 0.089 | 11.170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bushmeleva, K.; Vyshtakalyuk, A.; Terenzhev, D.; Belov, T.; Nikitin, E.; Zobov, V. Antioxidative and Immunomodulating Properties of Aronia melanocarpa Extract Rich in Anthocyanins. Plants 2022, 11, 3333. https://doi.org/10.3390/plants11233333
Bushmeleva K, Vyshtakalyuk A, Terenzhev D, Belov T, Nikitin E, Zobov V. Antioxidative and Immunomodulating Properties of Aronia melanocarpa Extract Rich in Anthocyanins. Plants. 2022; 11(23):3333. https://doi.org/10.3390/plants11233333
Chicago/Turabian StyleBushmeleva, Kseniya, Alexandra Vyshtakalyuk, Dmitriy Terenzhev, Timur Belov, Evgeny Nikitin, and Vladimir Zobov. 2022. "Antioxidative and Immunomodulating Properties of Aronia melanocarpa Extract Rich in Anthocyanins" Plants 11, no. 23: 3333. https://doi.org/10.3390/plants11233333