

Effects of Simulated Nitrogen Deposition and Micro-Environment on the Functional Traits of Two Rare and Endangered Fern Species in a Subtropical Forest
 ,
,
Abstract
:1. Introduction
2. Results
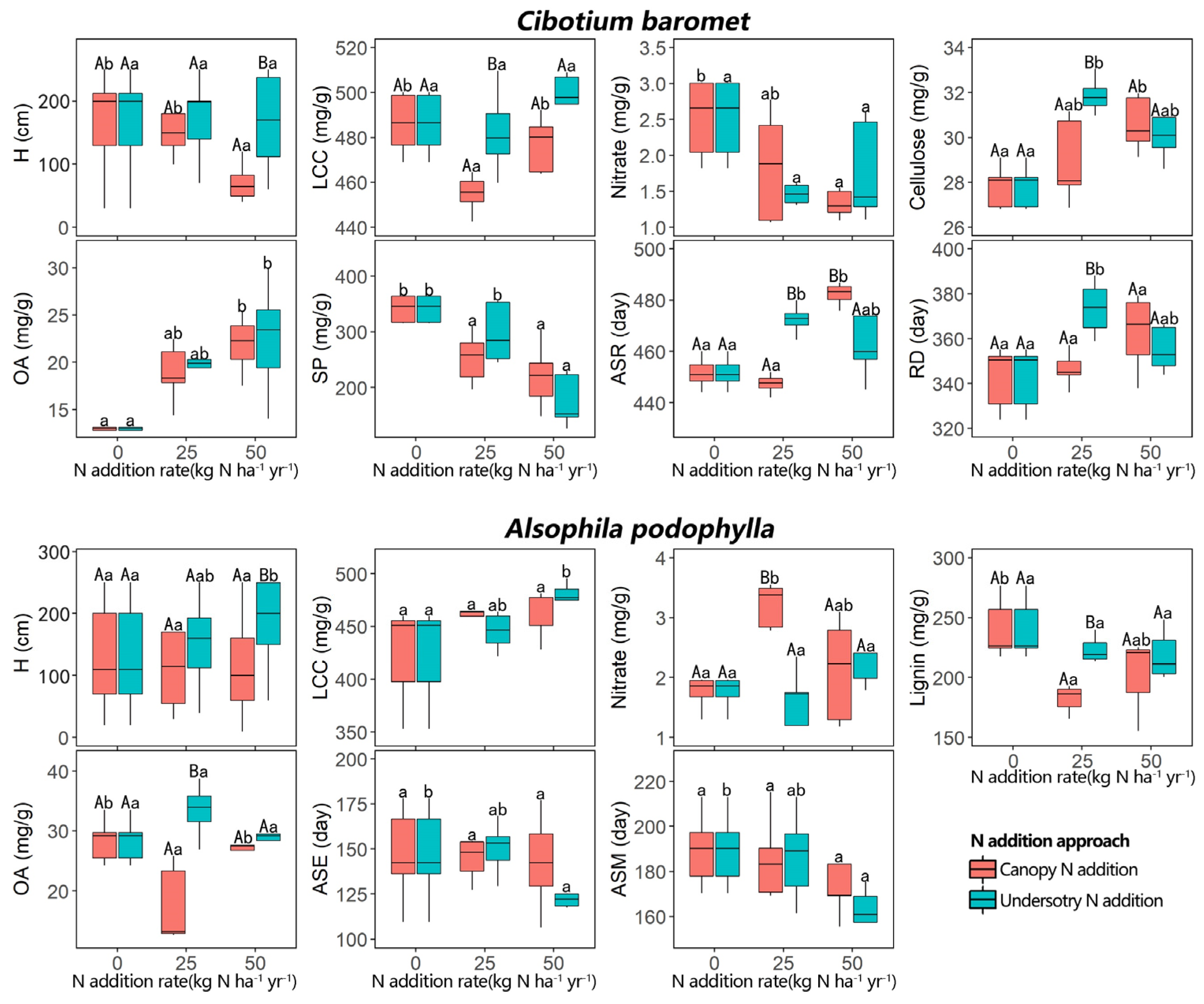
2.1. Effects of N Addition on Traits
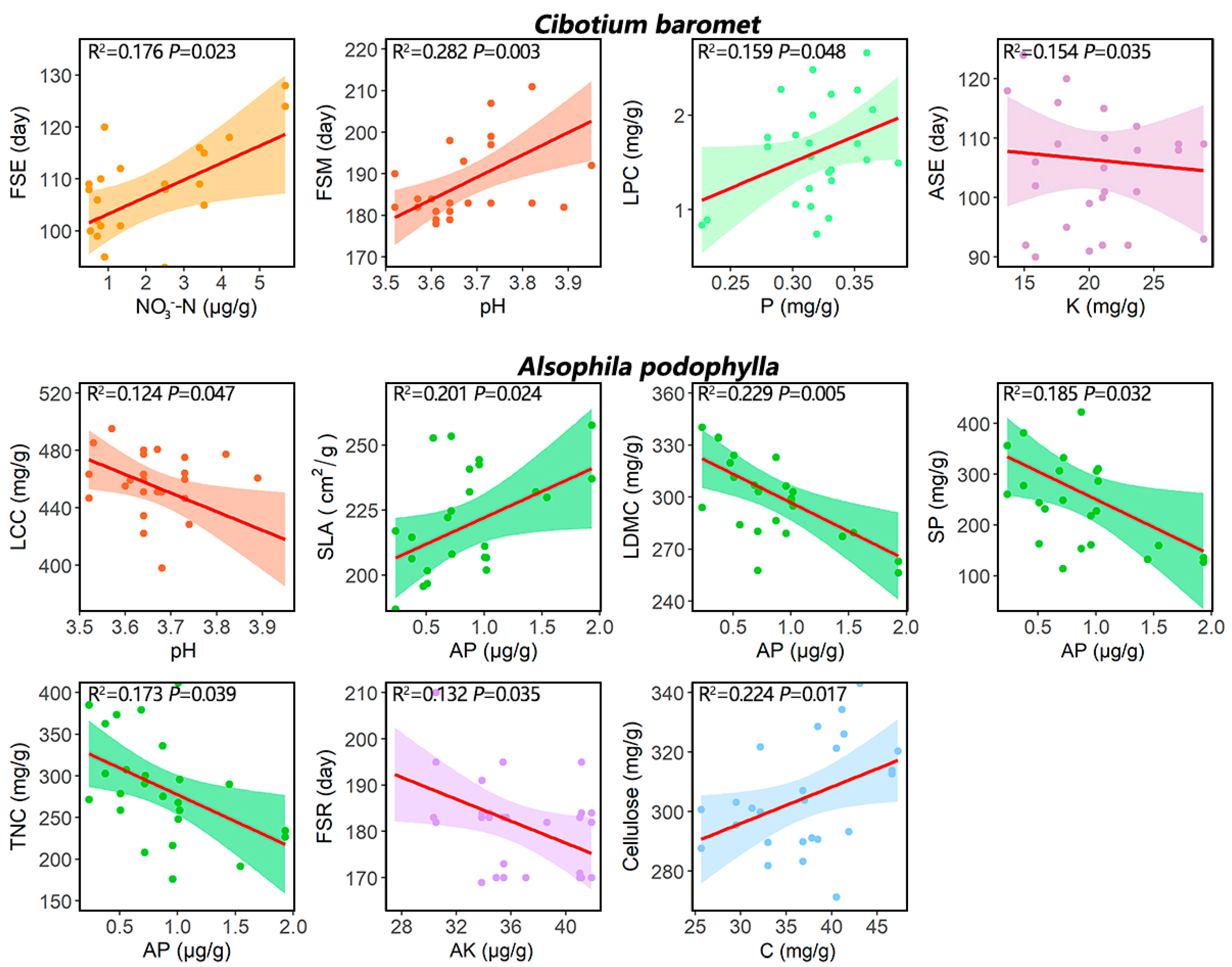
2.2. Micro-Environmental Variation and Its Effects on REF Traits
3. Discussion
3.1. Effects of N Addition on REF Traits
3.2. Micro-Environmental Variation and Its Effects on REF Traits
4. Materials and Methods
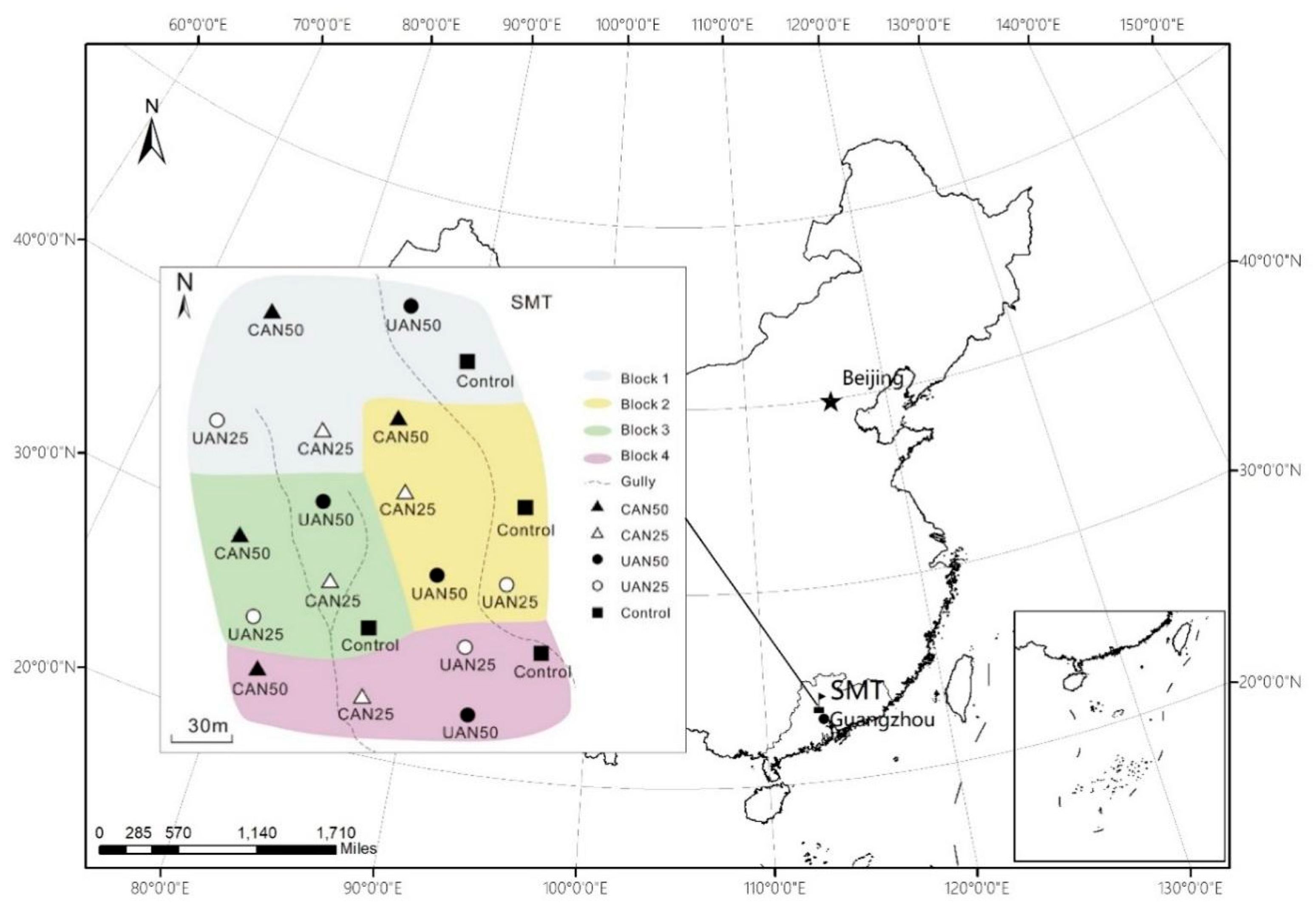
4.1. Study Site
4.2. Rare and Endangered Fern Species (REFs)
4.3. Experimental Design
4.4. Assessment of Plant and Soil Chemical Properties
4.5. Assessment of Functional Traits
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Battye, W.; Aneja, V.P.; Schlesinger, W.H. Is nitrogen the next carbon? Earth’s Future 2017, 5, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Steffen, W.; Richardson, K.; Rockstrom, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tang, Y.J. Responses of plant phenology to nitrogen addition: A meta-analysis. Oikos 2019, 128, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.K.; Mo, J.M.; Gilliam, F.S.; Zhou, G.Y.; Fang, Y.T. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob. Chang. Biol. 2010, 16, 2688–2700. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Zhang, S.K.; Huang, Y.; Cai, H.Y.; Zhu, X.Y. Understory and canopy additions of nitrogen differentially affect carbon and nitrogen metabolism of Psychotria rubra in an evergreen broad-leaved forest. Sci. Total Environ. 2020, 724, 138183. [Google Scholar] [CrossRef]
- Fang, Y.T.; Mo, J.M.; Zhou, G.Y.; Fang, Y.T. Response of diameter at breast height increment to N additions in forests of Dinghushan biophere reserve. J. Trop. Subtrop. Bot. 2005, 13, 198–204. [Google Scholar]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Romero-Pérez, I.E.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef]
- De Long, J.R.; Sundqvist, M.K.; Gundale, M.J.; Giesler, R.; Wardle, D.A.; Rasmann, S. Effects of elevation and nitrogen and phosphorus fertilization on plant defence compounds in subarctic tundra heath vegetation. Funct. Ecol. 2016, 30, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Wortman, E.; Tomaszewski, T.; Waldner, P.; Schleppi, P.; Thimonier, A.; Eugster, W.; Buchmann, N.; Sievering, H. Atmospheric nitrogen deposition and canopy retention influences on photosynthetic performance at two high nitrogen deposition Swiss forests. Tellus B Chem. Phys. Meteorol. 2012, 64, 17216. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Zhou, G.Y.; Zou, X.M.; Bai, E.; Scanlon, T.M.; Hou, E.Q.; et al. Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest. Proc. Natl. Acad. Sci. USA 2018, 115, 5187–5192. [Google Scholar] [CrossRef] [Green Version]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage–an empirical test in semi-arid forest ecosystems. J. Ecol. 2013, 101, 18–28. [Google Scholar] [CrossRef]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar] [CrossRef]
- Wu, Q.; Ding, J.; Yan, H.; Zhang, S.R.; Fang, T.; Ma, K.P. Effects of simulated precipitation and nitrogen addition on seedling growth and biomass in five tree species in Gutian Mountain, Zhejiang Province, China. Chin. J. Plant Ecol. 2011, 35, 256–267. [Google Scholar] [CrossRef]
- Teodoro, G.S.; Costa, P.D.B.; Brum, M.; Signori-Müller, C.; Alcantara, S.; Dawson, T.E.; West, A.G.; Lambers, H.; Oliveira, R.S. Desiccation tolerance implies costs to productivity but allows survival under extreme drought conditions in Velloziaceae species in campos rupestres. Environ. Exp. Bot. 2021, 189, 104556. [Google Scholar] [CrossRef]
- Liu, S.J.; Behm, J.E.; Wan, S.Q.; Yan, J.H.; Ye, Q.; Zhang, W.; Yang, X.D.; Fu, S.L. Effects of canopy nitrogen addition on soil fauna and litter decomposition rate in a temperate forest and a subtropical forest. Geoderma 2021, 382, 114703. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Tang, S.B.; Zhang, L.L.; Lambers, H.; Ren, W.D.; Lu, X.F.; Hou, E.Q.; Fu, S.L.; Kuang, Y.W.; Gilliam, F. Addition of nitrogen to canopy versus understorey has different effects on leaf traits of understorey plants in a subtropical evergreen broad-leaved forest. J. Ecol. 2020, 109, 692–702. [Google Scholar] [CrossRef]
- Jiang, X.Y.; Liu, N.; Lu, X.K.; Huang, J.G.; Cheng, J.; Guo, X.L.; Wu, S.H. Canopy and understory nitrogen addition increase the xylem tracheid size of dominant broadleaf species in a subtropical forest of China. Sci. Total Environ. 2018, 642, 733–741. [Google Scholar] [CrossRef]
- Magill, A.H.; Aber, J.D.; Berntson, G.M.; Mcdowell, W.H.; Nadelhoffer, K.J.; Melillo, J.M.; Steudler, P. Long-term nitrogen additions and nitrogen saturation in two temperate forests. Ecosystems 2000, 3, 238–253. [Google Scholar] [CrossRef]
- Rianey, S.M.; Nadelhoffer, K.J.; Silver, W.L.; Downs, M.R. Effects of chronic nitrogen additions on understory species in a red pine plantation. Ecol. Addit. 1999, 9, 949–957. [Google Scholar] [CrossRef]
- Lu, X.F.; Ren, W.D.; Hou, E.Q.; Tang, S.B.; Zhang, L.L.; Liu, Z.F.; Lin, Y.B.; Fu, S.L.; Wen, D.Z.; Kuang, Y.W. Different effects of canopy and understory nitrogen addition on soil organic carbon and its related processes in a subtropical forest. J. Soils Sediments 2020, 21, 235–244. [Google Scholar] [CrossRef]
- Shan, L.P.; Song, C.C.; Zhang, X.H.; Ren, J.S. Effects of long-term nitrogen and phosphorus addition on plant defence compounds in a freshwater wetland. Ecol. Indic. 2018, 94, 1–6. [Google Scholar] [CrossRef]
- Tian, D.; Du, E.Z.; Jiang, L.; Ma, S.H.; Zeng, W.J.; Zou, A.L.; Feng, C.Y.; Xu, L.C.; Xing, A.J.; Wang, W.; et al. Responses of forest ecosystems to increasing N deposition in China: A critical review. Environ. Pollut. 2018, 243, 75–86. [Google Scholar] [CrossRef]
- Liang, X.Y.; Zhang, T.; Lu, X.K.; Ellsworth, D.S.; BassiriRad, H.; You, C.M.; Wang, D.; He, P.C.; Deng, Q.; Liu, H.; et al. Global response patterns of plant photosynthesis to nitrogen addition: A meta-analysis. Glob. Chang. Biol. 2020, 26, 3585–3600. [Google Scholar] [CrossRef]
- Palmroth, S.; Bach, L.H.; Nordin, A.; Palmqvist, K. Nitrogen-addition effects on leaf traits and photosynthetic carbon gain of boreal forest understory shrubs. Oecologia 2014, 175, 457–470. [Google Scholar] [CrossRef]
- Keeler, B.L.; Hobbie, S.E.; Kellogg, L.E. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: Implications for litter and soil organic matter decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar] [CrossRef]
- Alvarez-Clare, S.; Mack, M.C.; Brooks, M. A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest. Ecology 2013, 94, 1540–1551. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.K.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Mo, J.M. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.K.; Perks, M.P.; Weatherall, A.; Baggs, E.M.; Mencuccini, M. Does canopy nitrogen uptake enhance carbon sequestration by trees? Glob. Chang. Biol. 2016, 22, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.L.; Zhang, H.Z.; Liu, T.; Zhang, W.; Shao, Y.H.; Ha, D.L.; Li, Y.Q.; Zhang, C.M.; Cai, X.A.; Rao, X.Q.; et al. Consistent effects of canopy vs. understory nitrogen addition on the soil exchangeable cations and microbial community in two contrasting forests. Sci. Total Environ. 2016, 553, 349–357. [Google Scholar] [CrossRef]
- Tian, Y.; Lu, H.F.; Wang, J.; Lin, Y.B.; Campbell, D.E.; Jian, S.G. Effects of canopy and understory nitrogen addition on the structure and eco-exergy of a subtropical forest community. Ecol. Indic. 2019, 106, 105459. [Google Scholar] [CrossRef]
- Tian, Y.; Tao, L.B.; Wang, J.; Lu, H.F.; Zhou, L.X.; Campbell, D.E.; Ren, H. Canopy and understory nitrogen additions did not significantly change the community structure of soil fauna under a mature subtropical forest. Sci. Total Environ. 2020, 718, 137438. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.T.; Zhao, P.; Zhu, L.W.; Zhao, X.H.; Ni, G.Y.; Ouyang, L.; Schafer, K.V.R.; Shen, W.J. Responses of sap flux and intrinsic water use efficiency to canopy and understory nitrogen addition in a temperate broadleaved deciduous forest. Sci. Total Environ. 2019, 648, 325–336. [Google Scholar] [CrossRef]
- Yu, B.Y.; Huang, J.G.; Ma, Q.Q.; Guo, X.L.; Liang, H.X.; Zhang, S.K.; Fu, S.L.; Wan, S.Q.; Yan, J.H.; Zhang, W. Comparison of the effects of canopy and understory nitrogen addition on xylem growth of two dominant species in a warm temperate forest, China. Dendrochronologia 2019, 56, 125604. [Google Scholar] [CrossRef]
- Zhang, S.K.; Rossi, S.; Huang, J.G.; Jiang, S.W.; Yu, B.Y.; Zhang, W.; Ye, Q. Intra-annual Dynamics of Xylem Formation in Liquidambar formosana Subjected to Canopy and Understory N Addition. Front. Plant Sci. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.W.; Hu, Y.T.; Zhao, X.H.; Zhao, P.; Ouyang, L.; Ni, G.Y.; Liu, N. Specific responses of sap flux and leaf functional traits to simulated canopy and understory nitrogen additions in a deciduous broadleaf forest. Funct. Plant Biol. 2019, 46, 986–993. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, S.K.; Huang, Y.; Wang, J.X.; Cai, H.Y. Canopy and understory additions of nitrogen change the chemical composition, construction cost, and payback time of dominant woody species in an evergreen broadleaved forest. Sci. Total Environ. 2020, 727, 138738. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Zhao, P.; Zhang, Z.Z.; Zhu, L.W.; Ouyang, L.; Ni, G.Y. Inconsistent Responses of Transpiration of Different Canopy Layers to Simulated Canopy and Understory N Depositions in a Low-Subtropical Evergreen Broadleaf Forest. J. Geophys. Res.-Biogeosci. 2020, 125, e2019JG005594. [Google Scholar] [CrossRef]
- Liu, N.; Wu, S.; Guo, Q.; Wang, J.; Cao, C.; Wang, J. Leaf nitrogen assimilation and partitioning differ among subtropical forest plants in response to canopy addition of nitrogen treatments. Sci. Total Environ. 2018, 637–638, 1026–1034. [Google Scholar] [CrossRef]
- Zhang, X.C.; Nishida, H. Cyatheaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2013; Volume 2–3, pp. 132–138. [Google Scholar]
- Xia, J.Y.; Wan, S.Q. Independent effects of warming and nitrogen addition on plant phenology in the Inner Mongolian steppe. Ann. Bot. 2013, 111, 1207–1217. [Google Scholar] [CrossRef]
- Levin, D.A. Flowering-time plasticity facilitates niche shifts in adjacent populations. New Phytol. 2009, 183, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Guo, J.J.; Li, Y.R.; Luo, G.W.; Li, L.; Yuan, H.Y.; Mur, L.A.J.; Guo, S.W. Negative effects of the simulated nitrogen deposition on plant phenolic metabolism: A meta-analysis. Sci. Total Environ. 2020, 719, 137442. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hui, D.; Ren, H.; Liu, N.; Sun, Z.; Yang, L.; Lu, H. Short-term canopy and understory nitrogen addition differ in their effects on seedlings of dominant woody species in a subtropical evergreen broadleaved forest. Glob. Ecol. Conserv. 2021, 31, e01855. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Wang, G.Y.; Zhuang, H.F.; Wang, L.H.; Innes, J.L.; Ma, K.P. Integrating hotspots for endemic, threatened and rare species supports the identification of priority areas for vascular plants in SW China. For. Ecol. Manag. 2021, 484, 118952. [Google Scholar] [CrossRef]
- Poorter, H.; Villar, R. The fate of acquired carbon in plants: Chemical composition and construction costs. In Plant Resource Allocation; Bazzaz, F.A., Grace, J., Eds.; Academic Press: New York, NY, USA, 1997; pp. 39–72. [Google Scholar]
- Li, H.; Li, M.; Luo, J.; Cao, X.; Qu, L.; Gai, Y.; Jiang, X.; Liu, T.; Bai, H.; Janz, D.; et al. N-fertilization has different effects on the growth, carbon and nitrogen physiology, and wood properties of slow- and fast-growing Populus species. J. Exp. Bot. 2012, 63, 6173–6185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Niu, K.C.; Liu, X.D.; Jia, P.; Du, G.Z. Linking flowering and reproductive allocation in response to nitrogen addition in an alpine meadow. J. Plant Ecol. 2013, 7, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Wangqiao, S.Y.; Xing, J.; Zeng, F.X.; Fang, F.Q.; Zheng, C.Y. Effects of nitrogen addition on the radial growth of Castanopsis carlesiiin Wuyi mountain. Acta Sci. Nat. Univ. Pekin. 2017, 53, 1143–1149. [Google Scholar]
- Liu, N.; Wang, J.; Guo, Q.; Wu, S.; Rao, X.; Cai, X.; Lin, Z. Alterations in leaf nitrogen metabolism indicated the structural changes of subtropical forest by canopy addition of nitrogen. Ecotoxicol. Environ. Saf. 2018, 160, 134–143. [Google Scholar] [CrossRef]
- Wang, J.X. Effects of Canopy and Understory Additions of Nitrogen on Carbon Metabolism of Representative Tree Species in Evergreen Broad-Leaved Forest; Chinese Academy of Sciences: Beijing, China, 2019. [Google Scholar]
- Mooney, H.A.; Fichtner, K.; Schulze, E.D. Growth, photosynthesis and storage of carbohydrates and nitrogen in Phaseolus lunatus in relation to resource availability. Oecologia 1995, 104, 17–23. [Google Scholar] [CrossRef]
- Yang, X.; Guo, R.; Knops, J.M.H.; Mei, L.; Kang, F.; Zhang, T.; Guo, J. Shifts in plant phenology induced by environmental changes are small relative to annual phenological variation. Agric. For. Meteorol. 2020, 294, 108144. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chiariello, N.R.; Loarie, S.R.; Mooney, H.A.; Field, C.B. Diverse responses of phenology to global changes in a grassland ecosystem. Proc. Natl. Acad. Sci. USA 2006, 103, 13740–13744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, I.W.; Miller, A.E.; Bowman, W.D.; Suding, K.N. Nitrogen preferences and plant-soil feedbacks as influenced by neighbors in the alpine tundra. Oecologia 2008, 156, 625–636. [Google Scholar] [CrossRef]
- Liu, L.; Monaco, T.A.; Sun, F.D.; Liu, W.; Gan, Y.M.; Sun, G. Altered precipitation patterns and simulated nitrogen deposition effects on phenology of common plant species in a Tibetan Plateau alpine meadow. Agric. For. Meteorol. 2017, 236, 36–47. [Google Scholar] [CrossRef]
- Hunt, M.A.; Davidson, N.J.; Unwin, G.L.; Close, D.C. Ecophysiology of the soft tree fern, Dicksonia antarctica Labill. Austral Ecol. 2002, 27, 360–368. [Google Scholar] [CrossRef]
- Brock, J.M.; Perry, G.L.W.; Lee, W.G.; Burns, B.R. Tree fern ecology in New Zealand: A model for southern temperate rainforests. For. Ecol. Manag. 2016, 375, 112–126. [Google Scholar] [CrossRef]
- Dearden, F.M.; Wardle, D.A. The potential for forest canopy litterfall interception by a dense fern understorey, and the consequences for litter decomposition. Oikos 2008, 117, 83–92. [Google Scholar] [CrossRef]
- Jiang, L.; Wei, C.S.; He, Z.S.; Zhu, J.; Xing, C.; Wang, X.L.; Liu, J.F.; Shen, C.X.; Shi, Y.W. Functional trait variation of plant communities in canopy gaps of Castanopsis kawakamii natural forest. Chin. J. Plant Ecol. 2022, 46, 267–279. [Google Scholar] [CrossRef]
- Wang, J.L.; Wang, H.M.; Fu, X.L.; Yang, F.T.; Chen, F.S. Effects of intraspecific competition and litter coverage on fine root morphological traits of Cunninghamia lanceolata and Loropetalum chinensis. Chin. J. Ecol. 2015, 34, 596–603. [Google Scholar]
- Tian, Y.; Wang, J.; Zhou, L.; Tao, L.B.; Lin, Y.B.; Hui, D.F.; Ren, H.; Lu, H.F. Nitrogen budgets of a lower subtropical forest as affected by 6 years of over-canopy and understory nitrogen additions. Sci. Total Environ. 2022, 852, 158546. [Google Scholar] [CrossRef]
- Yang, Y.; Li, P.; He, H.; Zhao, X.; Datta, A.; Ma, W.; Zhang, Y.; Liu, X.; Han, W.; Wilson, M.C.; et al. Long-term changes in soil pH across major forest ecosystems in China. Geophys. Res. Lett. 2015, 42, 933–940. [Google Scholar] [CrossRef]
- Bauer, G.A.; Bazzaz, F.A.; Minocha, R.; Long, S.; Magill, A.; Aber, J.; Berntson, G.M. Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (Pinus resinosa Ait.) stand in the NE United States. For. Ecol. Manag. 2004, 196, 173–186. [Google Scholar] [CrossRef]
- Wheeler, J.A.; Frey, S.D.; Stinson, K.A. Tree seedling responses to multiple environmental stresses: Interactive effects of soil warming, nitrogen fertilization, and plant invasion. For. Ecol. Manag. 2017, 403, 44–51. [Google Scholar] [CrossRef]
- Sigurdsson, B.D. Elevated CO2 and nutrient status modified leaf phenology and growth rhythm of young Populus trichocarpa trees in a 3-year field study. Trees-Struct. Funct. 2001, 15, 403–413. [Google Scholar] [CrossRef]
- Raich, J.W.; Russell, A.E.; Crews, T.E.; Farrington, H.; Vitousek, P. Both nitrogen and phosphorus limit plant production on young Hawaiian lava flows. Biogeochemistry 1996, 32, 1–14. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995. [Google Scholar]
- Long, W.X.; Yang, X.B.; Wu, Q.S.; Li, D.H.; Guo, T. Characteristics of Alsophila podophylla population and its community in the tropical rainforest in Wuzhi Mountains. Biodivers. Sci. 2008, 16, 83–91. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, W.J.; Zhu, S.D.; Wan, S.Q.; Luo, Y.Q.; Yan, J.H.; Wang, K.Y.; Liu, L.; Dai, H.T.; Li, P.X.; et al. CAN Canopy Addition of Nitrogen Better Illustrate the Effect of Atmospheric Nitrogen Deposition on Forest Ecosystem? Sci. Rep. 2015, 5, 11245. [Google Scholar] [CrossRef] [Green Version]
- Fenn, M.E.; Jovan, S.; Yuan, F.; Geiser, L.; Meixner, T.; Gimeno, B.S. Empirical and simulated critical loads for nitrogen deposition in California mixed conifer forests. Environ. Pollut. 2008, 155, 492–511. [Google Scholar] [CrossRef]
- Roth, T.; Kohli, L.; Rihm, B.; Meier, R.; Achermann, B. Using change-point models to estimate empirical critical loads for nitrogen in mountain ecosystems. Environ. Pollut. 2017, 220, 1480–1487. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.P.; Cheng, X.Y.; Zhang, G.F.; Li, H.; Gui, M. Observation on gametophyte development and apogamy of the endangered fern Cibotium baromet. Plant Physiol. J. 2016, 52, 1305–1311. [Google Scholar]
- Wang, S.; Xie, T. China Species Red List; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Botanical Journal of the Linnean Society. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Miatto, R.C.; Wright, I.J.; Batalha, M.A. Relationships between soil nutrient status and nutrient-related leaf traits in Brazilian cerrado and seasonal forest communities. Plant Soil 2016, 404, 13–33. [Google Scholar] [CrossRef]
- Han, T.T.; Lu, H.F.; Ren, H.; Wang, J.; Song, G.M.; Hui, D.F.; Guo, Q.F.; Zhu, S.D. Are reproductive traits of dominant species associated with specific resource allocation strategies during forest succession in southern China? Ecol. Indic. 2019, 102, 538–546. [Google Scholar] [CrossRef]
- Huang, D.H.; Zhou, L.L.; Fan, H.M.; Jia, Y.F.; Liu, M.B. Responses of aggregates and associated soil available phosphorus, and soil organic matter in different slope aspects, to seasonal freeze–thaw cycles in Northeast China. Geoderma 2021, 402, 115184. [Google Scholar] [CrossRef]
- Xu, Y.M.; Wang, X.X.; Bai, J.H.; Wang, D.W.; Wang, W.; Guan, Y.A. Estimating the spatial distribution of soil total nitrogen and available potassium in coastal wetland soils in the Yellow River Delta by incorporating multi-source data. Ecol. Indic. 2020, 111, 106002. [Google Scholar] [CrossRef]
- Lee, P.H.; Chen, S.W.; Chiou, W.L.; Huang, Y.M.; Liu, H.Y. Phenology of 13 fern species in a tropical monsoon forest of southern Taiwan. Int. J. Plant Reprod. Biol. 2016, 8, 87–97. [Google Scholar] [CrossRef]
- Fernández, H. Current Advances in Fern Research.; Springer International Publishing: Oviedo, Spain, 2018. [Google Scholar]
- Gough, L.; Gross, K.L.; Cleland, E.E.; Clark, C.M.; Collins, S.L.; Fargione, J.E.; Pennings, S.C.; Suding, K.N. Incorporating clonal growth form clarifies the role of plant height in response to nitrogen addition. Oecologia 2012, 169, 1053–1062. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, X.; Lin, H.; Lin, X.; Huang, L. Summary of changes in plant functional traits and environmental factors in different successional stages of island plants. Acta Ecol. Sin. 2019, 39, 3749–3758. [Google Scholar]
- Wen, B.B.; Xiao, W.; Mu, Q.; Li, D.M.; Chen, X.D.; Wu, H.Y.; Li, L.; Peng, F.T. How does nitrate regulate plant senescence? Plant Physiol. Biochem. 2020, 157, 60–69. [Google Scholar] [CrossRef]
- Guo, A.; Zuo, X.; Zhang, S.; Hu, Y.; Yue, P.; Lv, P.; Li, X.; Zhao, S.; Yu, Q. Contrasting effects of plant inter- and intraspecific variation on community trait responses to nitrogen addition and drought in typical and meadow steppes. BMC Plant Biol. 2022, 22, 90. [Google Scholar] [CrossRef]
- Liu, J.; Wu, N.; Wang, H.; Sun, J.; Peng, B.; Jiang, P.; Bai, E. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 2016, 97, 1796–1806. [Google Scholar] [CrossRef]
- Aho, K.; Derryberry, D.; Peterson, T. Model selection for ecologists: The worldviews of AIC and BIC. Ecology 2014, 95, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Symonds, M.R.E.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Ranc, N.; Cagnacci, F.; Moorcroft, P.R. Memory drives the formation of animal home ranges: Evidence from a reintroduction. Ecol. Lett. 2022, 25, 716–728. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.P.; Archaux, F.; Hulin, F.; Bilger, I.; Gosselin, F. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests. For. Ecol. Manag. 2020, 460, 117897. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Category | Trait | C. baromet | A. podophylla | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAN | UAN | 25 | 50 | CAN | UAN | 25 | 50 | ||||||||||
| 0 vs. 25 | 0 vs. 50 | 25 vs. 50 | 0 vs. 25 | 0 vs. 50 | 25 vs. 50 | CAN vs. UAN | CAN vs. UAN | 0 vs. 25 | 0 vs. 50 | 25 vs. 50 | 0 vs. 25 | 0 vs. 50 | 25 vs. 50 | CAN vs. UAN | CAN vs. UAN | ||
| Growth trait | H | 0.82 | <0.01 ** | 0.35 | 1.00 | 0.98 | 1.00 | 0.97 | 0.04 * | 0.90 | 0.76 | 1.00 | 0.99 | 0.05 * | 0.62 | 0.80 | <0.01 *** |
| SLA | 0.89 | 1.00 | 0.81 | 1.00 | 0.58 | 0.57 | 0.89 | 0.47 | 0.65 | 0.99 | 0.41 | 1.00 | 0.97 | 0.99 | 0.55 | 1.00 | |
| LCC | 0.01 * | 0.83 | 0.12 | 0.99 | 0.79 | 0.54 | 0.04 * | 0.23 | 0.11 | 0.26 | 0.99 | 0.66 | 0.03 * | 0.37 | 0.74 | 0.80 | |
| LNC | 0.99 | 0.92 | 0.70 | 0.93 | 0.63 | 0.97 | 0.72 | 0.98 | 1.00 | 0.33 | 0.34 | 0.99 | 0.48 | 0.76 | 0.99 | 1.00 | |
| LPC | 1.00 | 0.93 | 0.91 | 0.98 | 0.65 | 0.91 | 0.97 | 0.97 | 0.58 | 1.00 | 0.65 | 0.98 | 1.00 | 1.00 | 0.89 | 1.00 | |
| Nitrate | 0.45 | 0.04 * | 0.67 | 0.32 | 0.35 | 1.00 | 1.00 | 0.77 | 0.01 * | 0.86 | 0.06 | 1.00 | 0.33 | 0.22 | <0.01 ** | 0.87 | |
| Defense trait | LDMC | 1.00 | 0.99 | 1.00 | 1.00 | 0.62 | 0.44 | 0.98 | 0.85 | 0.47 | 0.98 | 0.20 | 0.83 | 1.00 | 0.81 | 0.09 | 0.97 |
| Lignin | 0.98 | 0.74 | 0.42 | 0.99 | 1.00 | 0.94 | 1.00 | 0.90 | <0.01 ** | 0.07 | 0.55 | 0.70 | 0.50 | 1.00 | 0.04 * | 0.73 | |
| Cellulose | 0.61 | 0.01 * | 0.25 | <0.01 *** | 0.07 | 0.15 | 0.01 * | 0.94 | 0.26 | 0.99 | 0.46 | 0.90 | 0.92 | 1.00 | 0.74 | 0.99 | |
| OA | 0.31 | 0.04 * | 0.82 | 0.24 | 0.03 * | 0.79 | 1.00 | 1.00 | 0.03 * | 1.00 | 0.03 * | 0.60 | 1.00 | 0.65 | <0.01 ** | 1.00 | |
| SP | 0.03 * | <0.01 ** | 0.86 | 0.38 | <0.01 *** | 0.01 * | 0.65 | 0.66 | 0.61 | 0.98 | 0.29 | 1.00 | 0.94 | 0.92 | 0.58 | 0.67 | |
| TNC | 0.90 | 0.29 | 0.73 | 0.59 | 0.73 | 0.09 | 0.20 | 0.93 | 1.00 | 0.42 | 0.65 | 0.34 | 0.16 | 0.99 | 0.55 | 0.97 | |
| Reproductive trait | FSE | 1.00 | 0.80 | 0.67 | 0.28 | 1.00 | 0.51 | 0.54 | 0.76 | 0.89 | 0.99 | 0.99 | 0.98 | 1.00 | 1.00 | 0.66 | 0.96 |
| ASE | 0.49 | 1.00 | 0.54 | 1.00 | 1.00 | 1.00 | 0.54 | 1.00 | 1.00 | 0.99 | 0.98 | 1.00 | 0.05 * | 0.05 | 1.00 | 0.26 | |
| FSM | 0.94 | 1.00 | 0.99 | 0.95 | 1.00 | 0.95 | 0.68 | 1.00 | 0.98 | 0.95 | 0.77 | 1.00 | 0.45 | 0.66 | 0.96 | 0.93 | |
| ASM | 0.54 | 1.00 | 0.51 | 0.99 | 0.97 | 0.89 | 0.84 | 1.00 | 0.98 | 0.67 | 0.93 | 1.00 | 0.04 * | 0.11 | 1.00 | 0.64 | |
| FSR | 1.00 | 1.00 | 1.00 | 1.00 | 0.83 | 0.70 | 0.99 | 0.91 | 0.83 | 0.72 | 1.00 | 1.00 | 0.76 | 0.89 | 0.94 | 1.00 | |
| ASR | 0.99 | <0.01 ** | <0.01 ** | 0.03 * | 0.47 | 0.86 | 0.04 * | 0.05 * | 0.93 | 1.00 | 1.00 | 0.99 | 0.73 | 0.56 | 0.79 | 0.94 | |
| RD | 0.99 | 0.12 | 0.35 | <0.01 ** | 0.46 | 0.18 | 0.02 * | 0.91 | 0.43 | 1.00 | 0.65 | 0.92 | 0.99 | 0.80 | 0.18 | 1.00 | |
| Category | Traits | C. baromet | A. podophylla | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N Treatment | Soil | Subcanopy Cover | Soil × N Treatment | Subcanopy Cover × N Treatment | N Treatment | Soil | Subcanopy Cover | Soil × N Treatment | Subcanopy Cover × N Treatment | ||
| Growth trait | H | 1.31 | 0.00 | 2.90 | 9.37 | 11.16 | 0.00 | 1.67 | 14.83 | 4.57 | 1.56 |
| SLA | 8.05 | 0.00 | 3.32 | 28.06 | 31.49 | 8.38 | 0.00 | 3.38 | 19.14 | 27.90 | |
| LCC | 0.00 | 3.48 | 6.94 | 11.33 | 12.95 | 3.15 | 0.00 | 4.66 | 3.80 | 25.43 | |
| LNC | 7.41 | 1.46 | 0.00 | 25.43 | 27.60 | 5.54 | 0.00 | 1.72 | 24.92 | 25.15 | |
| LPC | 10.08 | 0.00 | 2.33 | 19.45 | 28.46 | 9.86 | 0.00 | 3.58 | 19.6 | 24.5 | |
| Nitrate | 0.17 | 0.00 | 0.01 | 21.96 | 17.16 | 0.00 | 7.61 | 9.34 | 24.33 | 19.50 | |
| Defense trait | LDMC | 9.69 | 0.00 | 3.96 | 24.66 | 28.89 | 4.04 | 0.00 | 3.16 | 23.20 | 23.49 |
| Lignin | 5.26 | 0.00 | 0.33 | 25.42 | 28.79 | 0.00 | 6.56 | 8.02 | 19.28 | 20.56 | |
| Cellulose | 0.00 | 11.99 | 13.6 | 10.87 | 20.51 | 8.7 | 0.00 | 2.27 | 20.82 | 20.24 | |
| OA | 0.00 | 2.10 | 2.88 | 22.77 | 23.04 | 0.00 | 7.54 | 10.03 | 19.41 | 20.22 | |
| SP | 0.00 | 4.27 | 3.19 | 11.55 | 22.03 | 43.4 | 0.00 | 37.95 | 50.72 | 60.88 | |
| TNC | 0.00 | 0.93 | 1.31 | 22.35 | 24.17 | 6.11 | 0.00 | 4.97 | 20.99 | 25.04 | |
| Reproductive trait | FSE | 6.24 | 0.00 | 2.93 | 19.48 | 20.82 | 7.44 | 0.00 | 2.30 | 20.38 | 22.05 |
| ASE | 8.44 | 0.00 | 2.70 | 27.62 | 23.84 | 0.83 | 0.00 | 3.03 | 11.65 | 10.35 | |
| FSM | 17.03 | 0.00 | 9.53 | 17.25 | 33.82 | 0.00 | 6.78 | 3.77 | 16.51 | 12.89 | |
| ASM | 7.06 | 0.40 | 0.00 | 19.44 | 20.97 | 0.60 | 0.00 | 2.84 | 11.38 | 10.08 | |
| FSR | 10.07 | 0.00 | 2.93 | 23.91 | 26.33 | 9.69 | 0.00 | 4.61 | 16.76 | 15.60 | |
| ASR | 0.00 | 10.16 | 8.17 | 18.33 | 14.92 | 4.93 | 0.00 | 2.13 | 4.69 | 4.41 | |
| RD | 0.00 | 7.13 | 8.83 | 13.99 | 17.78 | 2.92 | 0.00 | 0.70 | 15.57 | 14.55 | |
| Category | Trait | Abbreviation | Specific Function |
|---|---|---|---|
| Growth traits | Plant height (cm) | H | Light capture and competition capacities [12,82] |
| Specific leaf area (cm2/g) | SLA | Light capture and gaseous exchange capacities [12,77] | |
| Leaf carbon concentrations (mg/g) | LCC | Carbon fixation capacity [17,83] | |
| Leaf nitrogen concentrations (mg/g) | LNC | Maximum photosynthetic rate [12,77] | |
| Leaf phosphorus concentrations (mg/g) | LPC | Maximum photosynthetic rate [12,77] | |
| Leaf nitrate concentrations (mg/g) | Nitrate | Chlorophyll synthesis [12,84] | |
| Defense traits | Leaf dry matter content (mg/g) | LDMC | Resistance to herbivory and drought [12,77,85] |
| Leaf cellulose concentrations (mg/g) | Cellulose | Resistance to herbivory [26,86] | |
| Leaf lignin concentrations (mg/g) | Lignin | Resistance to herbivory [8,26] | |
| Leaf organic acid concentrations (mg/g) | OA | Resistance to oxidation by air pollutants [17,37] | |
| Leaf soluble phenolics concentrations (mg/g) | SP | Resistance to microbial pathogens and herbivory [8,37] | |
| Leaf total non-structural carbohydrates (mg/g) | TNC | Resistance to drought [14,52] | |
| Reproductive traits | First spore emergence date (day) | FSE | Reproductive phenology, niche separation of phenology, and reproductive efficiency [42,56,80,81] |
| All-spore emergence date (day) | ASE | ||
| First spore maturation date (day) | FSM | ||
| All-spore maturation date (day) | ASM | ||
| First spore release date (day) | FSR | ||
| All-spore release date (day) | ASR | ||
| Reproductive duration (day) | RD |
| Category | Variable | Explanation |
|---|---|---|
| N treatment | N treatment | Canopy N addition at 25 and 50 kg N ha−1 yr−1 (CAN25 and CAN50), understory N addition at 25 and 50 kg N ha−1 yr−1 (UAN25 and UAN50), and a control |
| Micro-environment | Soil chemical property | Soil pH (pH), soil organic carbon content (C, mg/g), Soil total nitrogen content (N, mg/g), Soil total phosphorus content (P, mg/g), Soil total potassium content (P, mg/g), Soil nitrate nitrogen content (NH4+-N, μg/g), Soil ammonium nitrogen content (NO3−-N, μg/g), Soil available phosphorus content (AP, μg/g), Soil available potassium content (AK, μg/g) |
| Subcanopy cover | Subcanopy cover (%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, L.; Wei, L.; Zhang, L.; Li, Y.; Tian, Y.; Liu, K.; Ren, H. Effects of Simulated Nitrogen Deposition and Micro-Environment on the Functional Traits of Two Rare and Endangered Fern Species in a Subtropical Forest. Plants 2022, 11, 3320. https://doi.org/10.3390/plants11233320
Ji L, Wei L, Zhang L, Li Y, Tian Y, Liu K, Ren H. Effects of Simulated Nitrogen Deposition and Micro-Environment on the Functional Traits of Two Rare and Endangered Fern Species in a Subtropical Forest. Plants. 2022; 11(23):3320. https://doi.org/10.3390/plants11233320
Chicago/Turabian StyleJi, Lingbo, Liping Wei, Lingling Zhang, Yuanqiu Li, Yang Tian, Ke Liu, and Hai Ren. 2022. "Effects of Simulated Nitrogen Deposition and Micro-Environment on the Functional Traits of Two Rare and Endangered Fern Species in a Subtropical Forest" Plants 11, no. 23: 3320. https://doi.org/10.3390/plants11233320