2.1. Effects of Essential Oils (EOs) on Longest Sprout Length
After 90 days of storage, a statistically significant two-way interaction between treatment and time was observed on sprout length (
Table 1). This suggests that the impact of treatment on sprout length depends on the amount of time that has passed. Furthermore, the main effects of both treatment and time were significant (
Table 1).
The EOs of
Myrtus communis,
Melaleuca quinquenervia, and
Cymbopogon citratus resulted in significant differences in sprout length relative to the control (
Table 2). Treatment with
C. citratus EO resulted in significant differences in sprout length from the control at all time points, whereas the effects of
M. communis and
M. quinquenervia EO treatments were significant between 45–90 days and 45–75 days of storage, respectively (
Table 2).
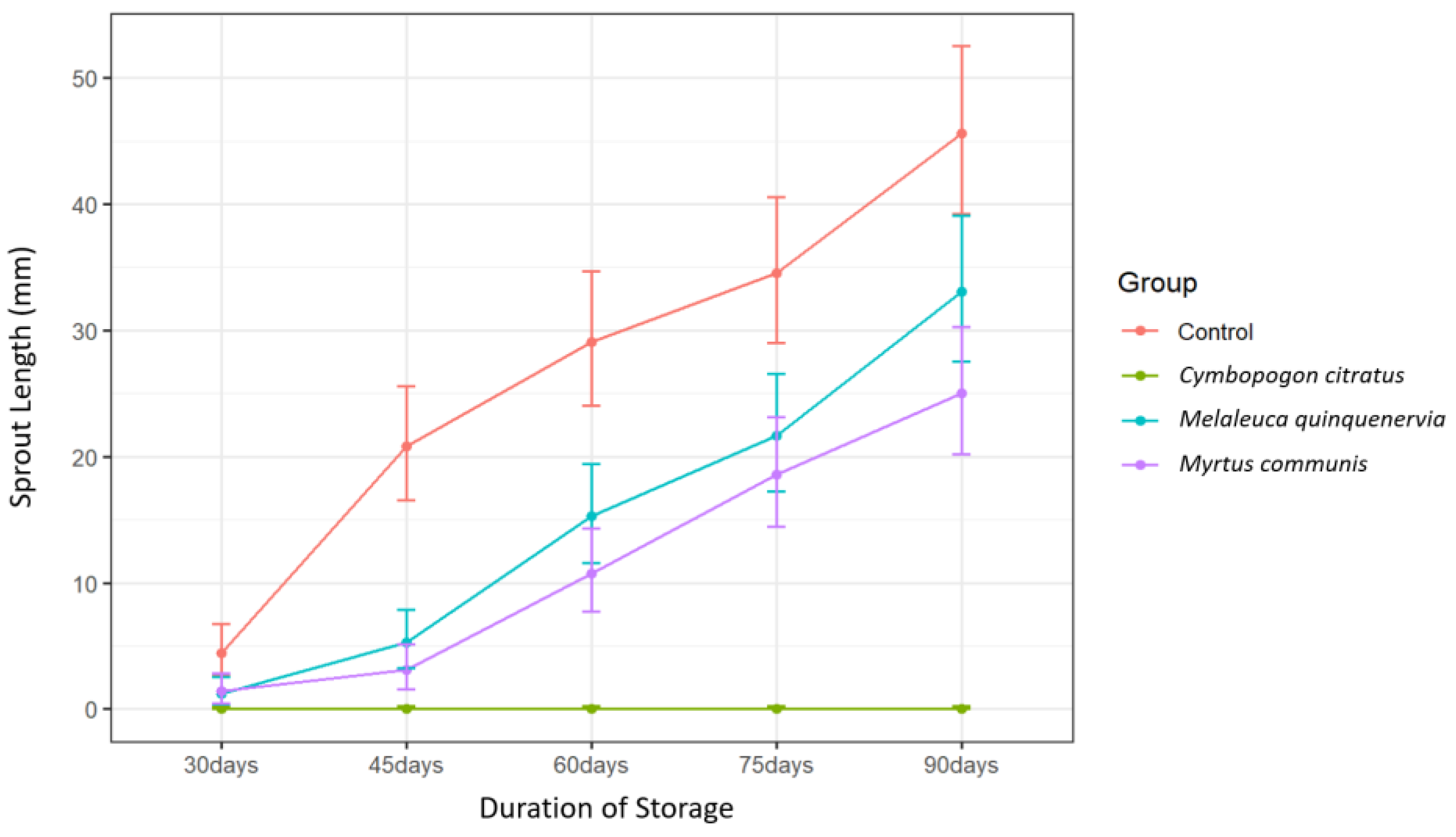
Complete suppression of sprouting was obtained over the entire 90-day storage period with
C. citratus EO treatment (
Figure 1). Sprout length due to
C. citratus EO treatment differed significantly from that due to either
M. communis or
M. quinquenervia EO treatment from 45 days until the end of the storage period (
p < 0.001, Tukey’s test) (
Table 3). Sprout length due to
M. communis EO treatment did not differ from that of
M. quinquenervia EO treatment at any time point (
p > 0.05, Tukey’s test) (
Table 3). Repeated applications of EOs are often required to maintain adequate sprout suppression over longer storage periods [
16,
22,
23]. Perhaps more effective sprout suppression could be achieved with
M. communis and
M. quinquenervia EOs if repeated applications were used.
Myrtus communis is a medicinal plant used in the food, pharmaceutical, and cosmetic industries [
24]. GC analysis of
M. communis EO reveals α-pinene, eucalyptol, linalool, p-cymene, geranyl acetate, and α-terpineol as major constituents (
Table 4). Geranyl acetate and α-pinene have previously been reported as an effective and somewhat effective sprout suppressant, respectively [
1]. Furthermore, these monoterpenes have also been shown to inhibit germination and growth in other species, perhaps through their induction of oxidative stress [
25,
26,
27]. It is possible that these compounds are responsible for this EO’s sprout suppressive properties.
M. communis EO has been associated with decreased weight and fruit firmness losses in strawberries [
28]. Furthermore, sunflower oil fortified with
M. communis EO can enhance the physiochemical properties of potato chips and could significantly increase their shelf-life [
29]. These reports and the findings of the current study suggest that
M. communis EO could be an effective component in EO sprout suppressant formulations.
Melaleuca quinquenervia is a medicinal plant with applications in aromatherapy, cosmetics, and pharmaceuticals, although its use in the food industry has been suggested [
34]. GC analysis reports eucalyptol, α-terpineol, α-pinene, viridiflorol, β-pinene, caryophyllene, viridiflorene, α-terpinyl acetate, and p-cymene as major constituents (
Table 5). Of these compounds, only α-pinene has been reported as a somewhat effective sprout suppressant [
1]. It is possible that the other major compounds, perhaps in combination with α-pinene, are responsible for the sprout suppressive capabilities of this EO. The findings of the current study encourage further investigation of both
M. quinquenervia and
M. communis EOs as sprout suppressants in other cultivars and using different application schemes, such as repeated or continuous application, to achieve more effective sprout control.
Cymbopogon citratus is another medicinal plant commonly used in the pharmaceutical and cosmetic industries with potential applications in the food industry [
37].
C. citratus EO is previously reported to be high in citral, a compound associated with effective sprout suppression in potato [
1,
37]. GC analysis of
C. citratus EO used in this study confirms the presence of citral, an aldehyde mixture of neral and geranial. Major compounds of
C. citratus EO include geranial, neral, geraniol, geranyl acetate, 6-methyl-5-heptene-2-one, camphene, linalool, 4-nonanone, γ-cadinene, caryophyllene, and limonene (
Table 6). In addition to geranial and neral, geranyl acetate has also been reported as an effective sprout suppressant [
1]. Given their high proportions within
C. citratus EO, it is likely that these compounds are responsible for the complete suppression of sprouting observed in this study.
Owolabi et al. [
38] demonstrated the potential of
C. citratus EO as a sprout suppressant in Russet Burbank potatoes, corroborating the findings in the present study. Furthermore, Belay et al. [
39] reported lower weight loss over a 14-week storage period in tubers treated with
C. citratus EO, however, no difference in sprout length relative to a control was observed in either cultivar tested. EOs are known to show differences in sprout suppression depending on potato cultivar [
16,
23,
39]. Therefore, it is possible that
C. citratus EO is more effective in cultivars Ranger Russet, used in the present study, and Russet Burbank than it is in Gudene or Jalene [
38,
39].
C. citratus EO application has also been associated with reductions in potato tuber moth (
Phthorimaea opperculella) infestation, suggesting that its use could provide additional benefits besides sprout suppression [
40].
2.2. Effects of Essential Oils (EOs) on Number of Germinated Eyes
After 90 days of storage, a statistically significant two-way interaction between treatment and time was observed on sprout number (
Table 1). This suggests that the impact of treatment on sprout number depends on the amount of time that has passed. Furthermore, the main effects of both treatment and time were significant (
Table 1).
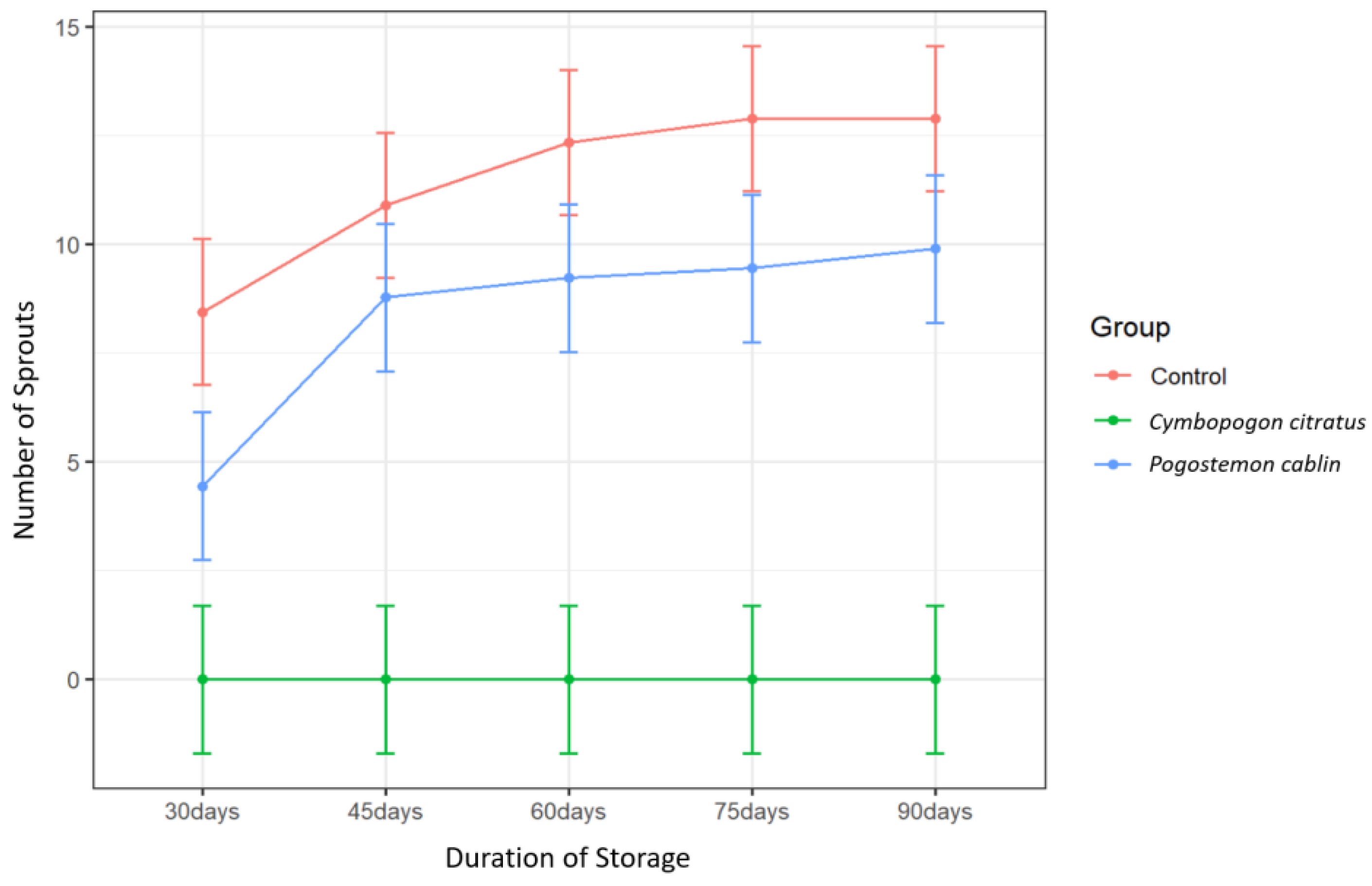
Only treatment with
C. citratus EO resulted in a significant difference in sprout number relative to the control (
Table 7). Though not significant, treatment with
Pogostemon cablin EO resulted in slightly fewer sprouts relative to the control from 60–90 days (
Figure 2). Complete suppression of sprouting was obtained over the entire 90-day storage period with
C. citratus EO treatment, resulting in zero sprouting throughout the study (
Figure 2). This contradicts findings by Belay et al. [
39] reporting no effect of
C. citratus EO on sprout numbers over the course of 16 weeks in cultivars Jalene and Gudene. However, like effects on sprout length, EO treatment effects on sprout number vary with cultivar [
39]. The results of the present study suggest that
C. citratus is a particularly effective sprout suppressant in Ranger Russet potatoes at room temperature.
Essential oils (EOs) are generally believed to achieve sprout suppression by damaging the meristematic tissue of developing sprouts [
22]. Indeed, 1,8-cineole application has been associated with complete necrosis of potato sprout tissue, whereas α-pinene and citral, an aldehyde mixture of geranial and neral, have been associated with necrosis of just the sprout tips [
41]. However, sprouting was completely inhibited with
C. citratus EO treatment, and no visible sprouts were observed throughout the 90-day storage period (
Figure 3). SEM of tuber eyes from
C. citratus treated potatoes reveals healthy, undeveloped bud tissue (
Figure 4). Due to lack of discernable damage to the tuber bud tissue, it is therefore possible that
C. citratus treatment may achieve sprout suppression through a mechanism that is different from the physical damage typically associated with EO sprout suppressants. For example, it is possible that
C. citratus EO treatment may suppress sprouting by interfering with the plant hormone balance within the tubers or through another mechanism entirely. Several plant hormones including auxins, gibberellins, cytokinins, and abscisic acid are known to regulate dormancy release and sprouting in potato tubers [
42]. Previous studies suggest that compounds such as citral may play a role in gibberellin and indole-3-acetic acid suppression in potato [
43], whereas pinene isomers have been associated with fluctuations in abscisic acid concentrations [
25]. Furthermore, 1,8-cineole and α-pinene have been shown to alter mitochondrial metabolism in corn, inhibiting root growth and interfering with germination [
44] Similar mechanisms may be present in potato. Additionally, garlic EO application on potato tubers has been shown to alter the abundance of specific proteins associated with seed germination [
45]. It is therefore possible that EOs achieve sprout suppression through a variety of mechanisms. Further studies performing proteomic analysis of EO-treated potatoes or investigating the effect of
C. citratus EO and its pure components on the levels of various plant hormones in treated potatoes could help to determine the active ingredient(s) and provide important insights into their mode of action. This could hasten the identification of additional EOs with sprout suppressive capabilities and identify new target genes in tuber breeding programs.
Recent studies suggest that various
Cymbopogon spp. EOs may be promising potato sprout manipulators.
Cymbopogon martini EO has been associated with sprout suppression at temperatures above 20 °C [
12,
39]. Similarly,
Cymbopogon nardus EO can completely suppress sprouting for up to 30 days after dormancy break [
46]. Conversely,
Cymbopogon schoenanthus EO has been associated with sprout enhancement and increased yields [
12]. The present study suggests that
C. citratus EO is also an effective sprout suppressant at room temperature, corroborating previous findings by Owolabi et al. [
38]. Greater focus and investigation into other species in this genus as potato sprout modulators in a wider range of cultivars is thus warranted.
The EO composition can vary widely depending on the plant parts used for extraction [
35]. For this reason, it is possible that EOs not observed to be effective sprout suppressants in the present study could possess sprout suppressive properties if different plant parts are used for extraction. Similarly, different extractions of EOs shown to be effective in the present study from one or more plant parts may display variable effects on potato sprouting due to varying compositions. Indeed, studies comparing the effects of EOs extracted from the bark, leaves, or fruit of these species on potato sprouting could more fully illustrate their potential as sprout suppressants while expediting the identification of active ingredients and the best sources of EOs for use in this industry. Nevertheless, the EO of
C. citratus, the most potent treatment, is commonly extracted from the whole aboveground plant parts in vegetative stage, that is stems and leaves.
Potatoes may be stored for many months before use, requiring multiple applications of sprout suppressants [
13]. As only a single application of each EO was used, and typically, EO-based products are applied every 2 or 4 weeks, the storage period in the present study was set to 90 days. Future studies could investigate longer storage lengths or may test different concentrations of the most effective EOs. Importantly, even if an EO possesses significant sprout suppressive or inhibitory properties, it is possible that its application may alter the flavor, texture, or nutritional quality of treated potatoes. Therefore, additional studies are needed to investigate
C. citratus and other EOs for their effects on other aspects of potato quality before commercial products may be developed.
Furthermore, larger studies comparing the effectiveness of C. citratus EO to conventional methods such as CIPC in semi-commercial and commercial settings are also needed. These types of studies are necessary to evaluate the efficacy of C. citratus EO in industrial settings and determine the feasibility of scaling up its use to a commercial scale.








{kind=link}
{kind=link}
{kind=link}
{kind=link}