
Energy vs. Nutritional Potential of Virginia Mallow (Sida hermaphrodita L.) and Cup Plant (Silphium perfoliatum L.)
 ,
,  , , , , ,
, , , , ,
Abstract
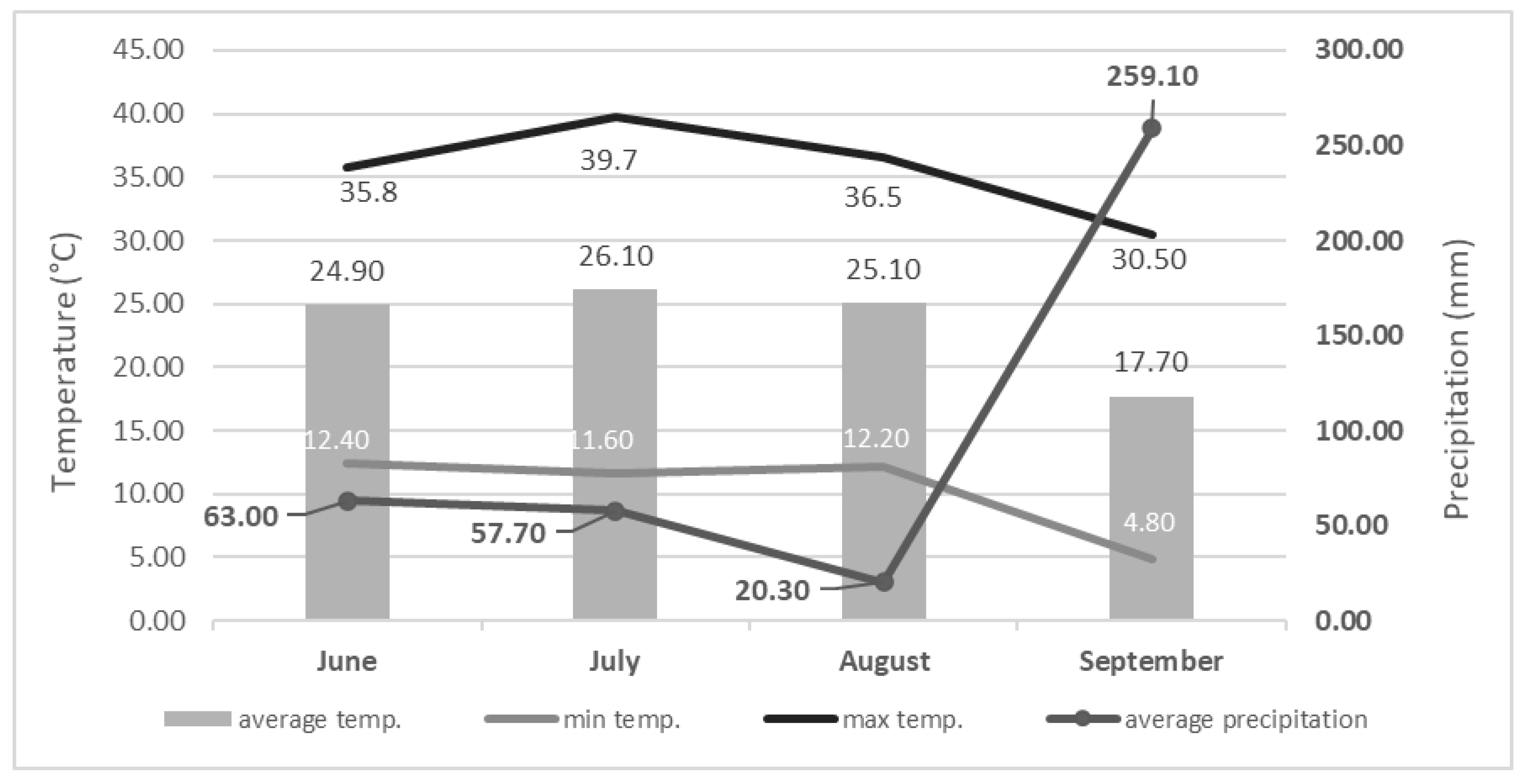
:1. Introduction
2. Results and Discussion
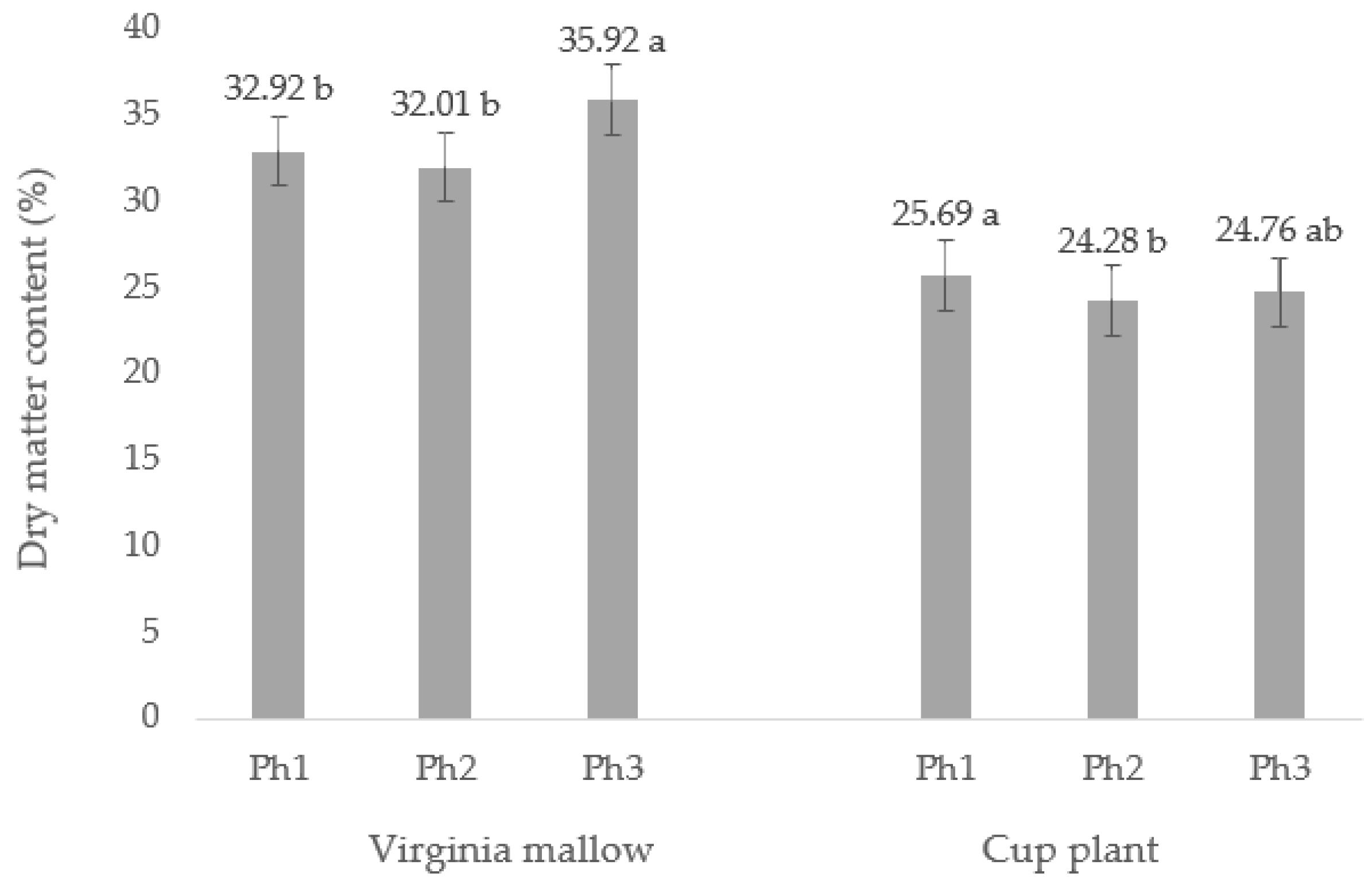
2.1. Dry Matter Content
2.2. Energetic Characteristics of Virginia Mallow and Cup Plant
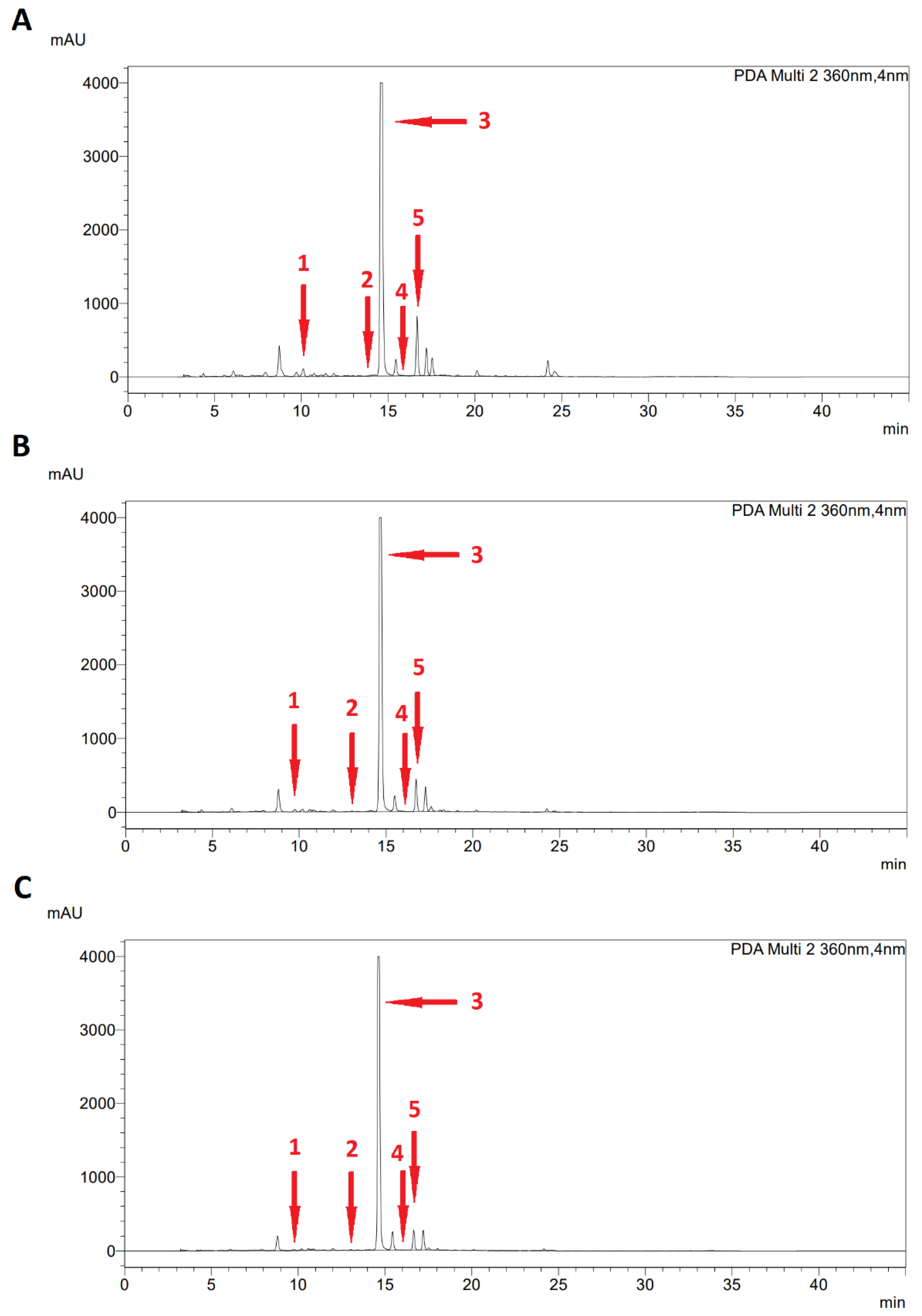
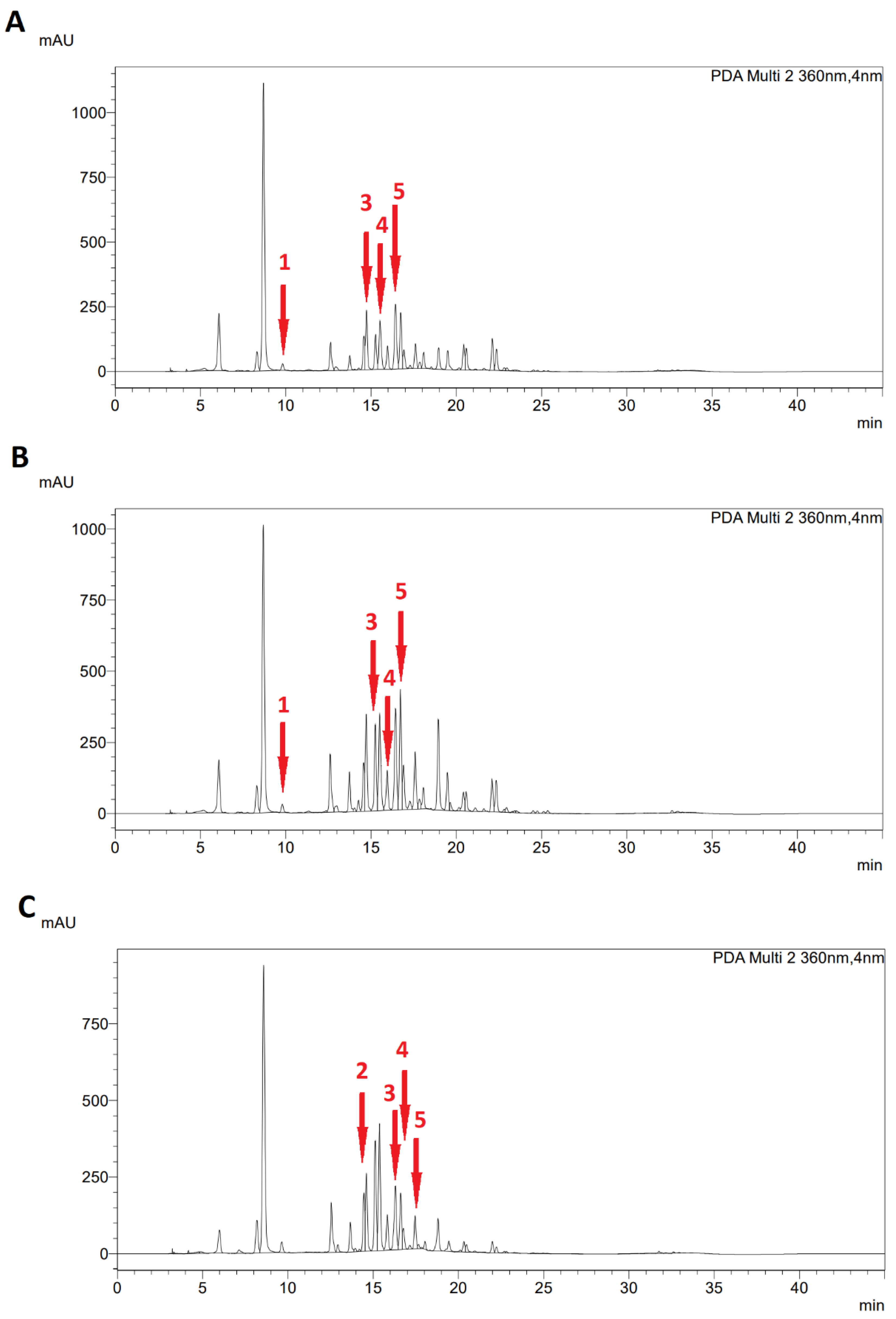
2.3. Specialized Metabolites of Fresh Leaves
2.4. Pigment Compounds Content
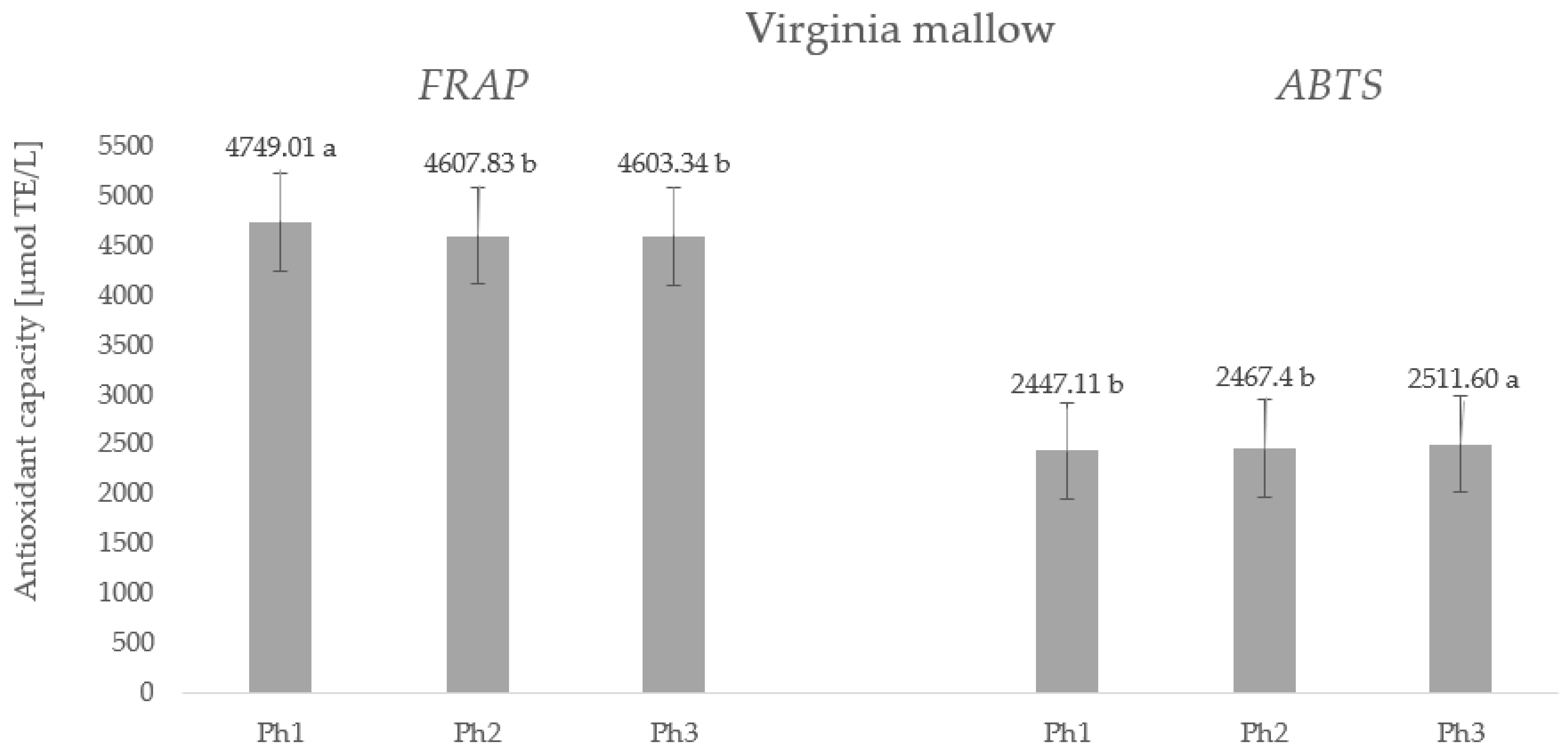
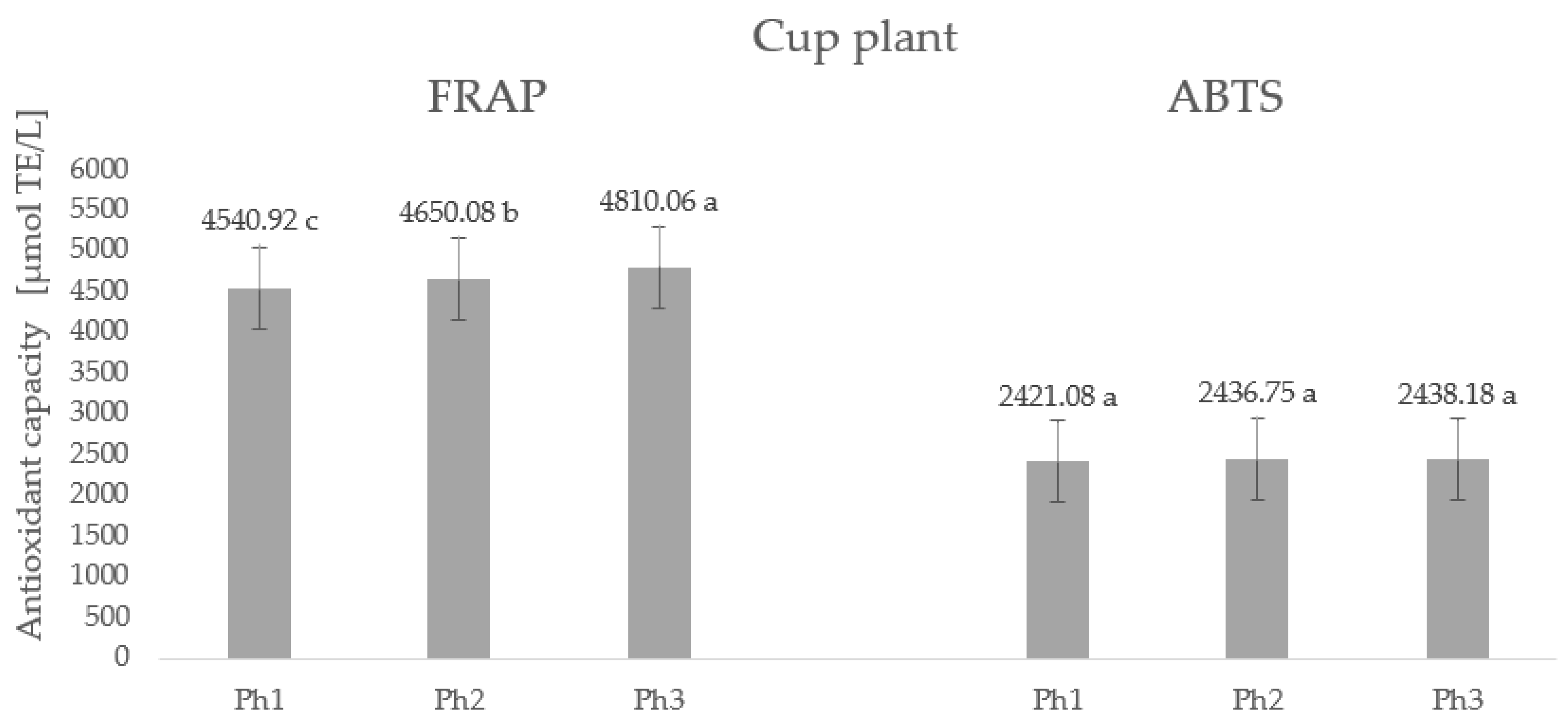
2.5. Antioxidant Capacity
3. Materials and Methods
3.1. Plant Material
3.2. Total Dry Matter Content
3.3. Energy Biomass Characterization
3.4. Determination of Specialized Metabolites
3.5. High-Performance Liquid Chromatography (HPLC)
3.6. Determination of Antioxidant Capacity (FRAP and ABTS)
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Greenpeace International Annual Report. Greenpeace. 2019. Available online: https://www.greenpeace.org/static/planet4-international-stateless/2020/06/544c2eca-greenpeace-international-annual-report-2019.pdf (accessed on 14 September 2022).
- Berners-Lee, M.; Kennelly, C.; Watson, R.; Hewitt, C.N. Current global food production is sufficient to meet human nutritional needs in 2050 provided there is radical societal adaptation. Elem. Sci. Anthr. 2018, 6, 52. [Google Scholar] [CrossRef]
- Janiszewska, D.; Ossowska, L. The Role of Agricultural Biomass as a Renewable Energy Source in European Union Countries. Energies 2022, 15, 6756. [Google Scholar] [CrossRef]
- Ekins, P.; Domenech, T.; Drummond, P.; Bleischwitz, R.; Hughes, N.; Lotti, L. The Circular Economy: What, Why, How and Where. In Proceedings of the OECD/EC Workshop, Managing Environmental and Energy Transitions for Regions and Cities, Paris, France, 5 July 2019; Available online: https://www.oecd.org/cfe/regionaldevelopment/Ekins-2019-Circular-Economy-What-Why-How-Where.pdf (accessed on 6 October 2022).
- European Comission. A new circular economy action plan for a cleaner and more competitive Europe. In Communication from the Commission to the European Parliament; The Council; The European Economic and Social Committee; Committee of the Regions: Brussels, Belgium, 2019; Available online: https://eur-lex.europa.eu/legal-content/HR/TXT/HTML/?uri=CELEX:52020DC0098&from=DA (accessed on 6 October 2022).
- Knápek, J.; Králík, T.; Vávrová, K.; Valentová, M.; Horák, M.; Outrata, D. Policy implications of competition between conventional and energy crops. Renew. Sust. Energ. Rev. 2021, 151, 111618. [Google Scholar] [CrossRef]
- Zieliński, M.; Rusanowska, P.; Zielińska, M.; Dudek, M.; Nowicka, A.; Purwin, C.; Fijalowska, M.; Dębowski, M. Influence of preparation of Sida hermaphrodita silages on its conversion to methane. Renew. Energy. 2021, 163, 437–444. [Google Scholar] [CrossRef]
- Cumplido-Marin, L.; Graves, A.R.; Burgess, P.J.; Morhart, C.; Paris, P.; Jablonowski, N.D.; Facciotto, G.; Bury, M.; Martens, R.; Nahm, M. Two Novel Energy Crops: Sida hermaphrodita (L.) Rusby and Silphium perfoliatum L.—State of Knowledge. Agronomy 2020, 10, 928. [Google Scholar] [CrossRef]
- Popp, J.; Lakner, Z.; Harangi-Rákos, M.; Fári, M. The effect of bioenergy expansion: Food, energy, and environment. Renew. Sust. Energ. Rev. 2014, 32, 559–578. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, G.; Pankiewicz, U.; Kowalski, R. Evaluation of Chemical Composition of Some Silphium L. Species as Alternative Raw Materials. Agriculture 2020, 10, 132. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, N.H.; Wojcinska, M.; Drost-Karbowska, K.; Matławska, I.; Williams, J.; Mabry, T.J. Kaempferol triosides from Silphium perfoliatum. Phytochemistry 2002, 60, 835–838. [Google Scholar] [CrossRef]
- Kowalski, R.; Wolski, T. TLC and HPLC Analysis of the phenolic acids in Silphium perfoliatum L. leaves, inflorescences and rhizomes. JPC-J Planar Chromat. 2003, 16, 230–236. [Google Scholar] [CrossRef]
- Kowalski, R. Selected secondary metabolites in leaves, inflorescences, and rhizomes of Silphium perfoliatum L.—Alternative herbal plant. Folia Hortic. 2003, 15, 203–209. [Google Scholar]
- Subramanian, S.; Blanton, L.V.; Frese, S.A.; Charbonneau, M.; Mills, D.A.; Gordon, J.I. Cultivating Healthy Growth and Nutrition through the Gut Microbiota. Cell. 2015, 161, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Aminah, N.S.; Laili, E.R.; Rafi, M.; Rochman, A.; Insanu, M.; Tun, K.N.W. Secondary metabolite compounds from Sida genus and their bioactivity. Heliyon 2021, 7, e06682. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Guo, L.; Jiang, G.; Song, Y.; Muminov, M.A. Advances of organic products over conventional productions with respect to nutritional quality and food security. Acta Ecol. Sin. 2018, 38, 53–60. [Google Scholar] [CrossRef]
- Assefa, A.; Debella, A. Review on dry matter production and partitioning as affected by different environmental conditions. Int. J. Adv. Res. Biol. Sci. 2020, 7, 37–46. [Google Scholar]
- Gao, K.; Yu, Y.F.; Xia, Z.T.; Yang, G.; Xing, Z.L.; Qi, L.T.; Ling, L.Z. Response of height, dry matter accumulation and partitioning of oat (Avena sativa L.) to planting density and nitrogen in Horqin Sandy Land. Sci. Rep. 2019, 9, 7961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baafi, E.; Gracen, V.E.; Manu-Aduening, J.; Blay, E.T.; Ofori, K.; Carey, E.E. Genetic control of dry matter, starch and sugar content in sweetpotato. Acta Agric. Scand. B Soil Plant Sci. 2017, 67, 110–118. [Google Scholar] [CrossRef]
- Antoszkiewicz, Z.; Fijałkowska, M.; Mazur-Ku ́snirek, M.; Przemieniecki, S.; Purwin, C. Effect of a harvest date and cutting height on the concentrations of carotenoids and tocopherols in Virginia fanpetals (Sida hermaphrodita) Herbage and Silage. J. Elem. 2019, 24, 1195–1202. [Google Scholar] [CrossRef]
- Voća, N.; Leto, J.; Karažija, T.; Bilandžija, N.; Peter, A.; Kutnjak, H.; Šurić, J.; Poljak, M. Energy Properties and Biomass Yield of Miscanthus × Giganteus Fertilized by Municipal Sewage Sludge. Molecules 2021, 26, 4371. [Google Scholar] [CrossRef]
- Bilandžija, N.; Jurišić, V.; Voća, N.; Leto, J.; Matin, A.; Sito, S.; Krička, T. Combustion properties of Miscanthus × giganteus biomass—Optimization of harvest time. J. Energy Inst. 2017, 90, 528–533. [Google Scholar] [CrossRef]
- Bilandžija, N.; Voća, N.; Leto, J.; Jurišić, V.; Grubor, M.; Matin, A.; Geršić, A.; Krička, T. Yield and biomass composition of Miscanthus × giganteus in the mountain area of Croatia. Trans. Famena 2018, 42, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Jurišić, V.; Bilandžija, N.; Krička, T.; Leto, J.; Matin, A.; Kuže, I. Fuel properties comparision of allochtonous Miscanthus × giganteus and autochtonous Arundo donax L.: A case stady in Croatia. Agric. Conspec. Sci. 2014, 79, 7–11. [Google Scholar]
- Rusanowska, P.; Zielinski, M.; Dudek, M.; Debowski, M. Mechanical pretreatment of lignocellulosic biomass for methane fermentation in innovative reactor with cage mixing system. J. Ecol. Eng. 2018, 19, 219–224. [Google Scholar] [CrossRef]
- Krička, T.; Bilandžija, N.; Jurišić, V.; Voća, N.; Matin, A. Energy analysis of main residual biomass in Croatia. Afr. J. Agric. Res. 2012, 7, 6383–6388. [Google Scholar]
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G. An overview of the chemical composition of biomass. Fuel 2010, 89, 913–933. [Google Scholar] [CrossRef]
- Barth, C.; De Tullio, M.; Conklin, P.L. The role of ascorbic acid in the control of flowering time and the onset of senescence. J. Exp. Bot. 2006, 57, 1657–1665. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Singh Tomar, N.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef]
- Bhattarai, S.; Biswas, D.; Fu, Y.B.; Biligetu, B. Morphological, physiological, and genetic responses to salt stress in Alfalfa: A review. Agronomy 2020, 10, 577–593. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Bhardwaj, R.; Thukral, A.K.; Al-Huqail, A.A.; Siddiqui, M.H.; Ahmad, P. Oxidative stress mitigation and initiation of antioxidant and osmoprotectant responses mediated by ascorbic acid in Brassica juncea L. subjected to copper (II) stress. Ecotoxicol. Environ. Saf. 2019, 182, 109436–109445. [Google Scholar] [CrossRef]
- Zhou, X.; Gu, Z.; Xu, H.; Chen, L.; Tao, G.; Yu, Y.; Li, K. The Effects of Exogenous Ascorbic Acid on the Mechanism of Physio-logical and Biochemical Responses to Nitrate Uptake in Two Rice Cultivars (Oryza sativa L.) Under Aluminum Stress. J. Plant Growth Regul. 2016, 35, 1013–1024. [Google Scholar] [CrossRef]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voca, S.; Šic Žlabur, J.; Fabek Uher, S.; Peša, M.; Opačić, N.; Radman, S. Neglected Potential of Wild Garlic (Allium ursinum L.)—Specialized Metabolites Content and Antioxidant Capacity of Wild Populations in Relation to Location and Plant Phenophase. Horticulturae 2022, 8, 24. [Google Scholar]
- Čvorović, J.; Ziberna, L.; Fornasaro, S.; Tramer, F.; Passamonti, S. Bioavailability of Flavonoids: The Role of Cell Membrane Transporters. In Polyphenols: Mechanisms of Action in Human Health and Disease; Zibadi, S., Watson, R., Preedy, V., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 295–320. [Google Scholar] [CrossRef]
- Vattem, D.A.; Shetty, K. Biological functionality of ellagic acid: A review. J. Food Biochem. 2005, 29, 234–266. [Google Scholar] [CrossRef]
- Taofiq, O.; González-Paramás, A.; Barreiro, M.; Ferreira, I. Hydroxycinnamic Acids and Their Derivatives: Cosmeceutical Significance, Challenges and Future Perspectives, a Review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef]
- Espíndola, K.M.M.; Ferreira, R.G.; Narvaez, L.E.M.; Silva Rosario, A.C.R.; da Silva, A.H.M.; Silva, A.G.B.; Vieira, P.O.; Monteiro, M.C. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Vaňková, K.; Marková, I.; Jašprová, J.; Dvořák, A.; Subhanová, I.; Zelenka, J.; Novosádová, I.; Rasl, J.; Vomastek, T.; Sobotka, R.; et al. Chlorophyll-Mediated Changes in the Redox Status of Pancreatic Cancer Cells Are Associated with Its Anticancer Effects. Oxi. Med. Cell. Longev. 2018, 2018, 4069167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and Chlorophylls as Antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Croatian Meteorological and Hydrological Service. Available online: https://meteo.hr/index_en.php (accessed on 6 October 2022).
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- CEN/TS 14774-2:2009; Solid Biofuels—Determination of Moisture Content. CEN: Brussels, Belgium, 2009.
- CEN/TS 15148:2009; Solid Biofuels—Determination of the Content of Volatile Matter. CEN: Brussels, Belgium, 2009.
- EN ISO 18122:2015; Solid Biofuels. Determination of Ash Content. ISO: Geneva, Switzerland, 2015.
- EN 15104:2011; Solid Biofuels. Determination of Total Content of Carbon, Hydrogen and Nitrogen—Instrumental Methods. SIS: Stockholm, Sweden, 2011.
- EN 15289:2011; Solid Biofuels. Determination of Total Content of Sulfur and Chlorine. ISO: Geneva, Switzerland, 2011.
- CEN/TS 14918:2005; Solid Biofuels—Method for the Determination of Calorific Value. CEN: Brussels, Belgium, 2005.
- Ough, C.S.; Amerine, M.A. Methods for Analysis of Musts and Wines, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1988. [Google Scholar]
- Holm, G. Chlorophyll mutations in barley. Acta Agric. Scand. 1954, 4, 457–471. [Google Scholar] [CrossRef]
- Wettstein, D. Chlorophyll-letale und der submikroskopische Formwechsel der Plastiden. Exp. Cell Res. 1957, 12, 427–434. [Google Scholar] [CrossRef]
- Otles, S.; Yalcin, B. Phenolic compounds analysis of root, stalk, and leaves of nettle. Sci. World J. 2012, 2012, 564367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repajić, M.; Cegledi, E.; Kruk, V.; Pedisić, S.; Çınar, F.; Kovačević, D.B.; Žutić, I.; Dragović-Uzelac, V. Accelerated Solvent Extraction as a Green Tool for the Recovery of Polyphenols and Pigments from Wild Nettle Leaves. Processes 2020, 8, 803. [Google Scholar] [CrossRef]
- Miller, N.J.; Diplock, A.T.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS® Proprietary Software 9,4 (TS1M7)/STAT 9.3; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virginia Mallow | Cup Plant | |
|---|---|---|
| Ash (%) | 2.92 | 5.86 |
| Coke (%) | 10.45 | 15.49 |
| Fixed Carbon (%) | 7.53 | 9.28 |
| Volatile Matters (%) | 80.31 | 74.45 |
| HHV * (MJ/kg) | 17.36 | 15.46 |
| LHV * (MJ/kg) | 16.02 | 14.54 |
| Virginia Mallow | Cup Plant | |
|---|---|---|
| Carbon (C) | 49.57% | 42.94% |
| Hydrogen (H) | 6.15% | 5.30% |
| Nitrogen (N) | 0.60% | 0.37% |
| Sulfur (S) | 0.06% | 0.02% |
| Oxygen (O) | 43.63% | 50.92% |
| Phenophases | AsA (mg/100 g fw) | TPC (mg GAE/100 g fw) | TFC (mg GAE/100 g fw) | TNFC (mg GAE/100 g fw) |
|---|---|---|---|---|
| Virginia Mallow | ||||
| Ph1 | 223.07 ± 10.17 a | 1079.59 ± 0.60 a | 434.00 ± 0.83 b | 645.59 ± 1.16 a |
| Ph2 | 229.79 ± 11.09 a | 835.58 ± 1.33 c | 393.44 ± 2.21 c | 442.15 ± 1.82 b |
| Ph3 | 170.30 ± 2.36 b | 1072.45 ± 0.92 b | 493.13 ± 1.90 a | 579.32 ± 2.81 c |
| LSD | 26.624 | 3.021 | 5.305 | 6.197 |
| ANOVA | p ≤ 0.0003 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 |
| Cup plant | ||||
| Ph1 | 24.42 ± 1.95 b | 674.94 ± 0.15 c | 399.09 ± 2.12 c | 399.09 ± 2.21 c |
| Ph2 | 38.10 ± 5.19 b | 1115.21 ± 0.59 a | 557.25 ± 1.70 a | 557.25 ± 2.00 a |
| Ph3 | 122.57 ± 9.36 a | 917.88 ± 2.45 b | 492.67 ± 4.25 b | 492.67 ± 1.91 b |
| LSD | 19.007 | 4.422 | 6.190 | 6.190 |
| ANOVA | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 |
| Phenophases | Caffeic Acid | Coumaric Acid | Ellagic Acid | Ferulic Acid | Naringin |
|---|---|---|---|---|---|
| Virginia Mallow | |||||
| Ph1 | 32.90 ± 6.46 a | 15.25 ± 2.48 b | 1421.95 ± 75.43 a | 4.27 ± 0.02 | 1404.98 ± 120.94 a |
| Ph2 | 26.05 ± 0.64 a | 19.90 ± 1.35 b | 1398.43 ± 13.23 a | 4.74 ± 0.57 | 760.79 ± 22.84 b |
| Ph3 | 13.07 ± 0.58 b | 30.55 ± 0.45 a | 1213.63 ± 16.58 b | 4.10 ± 0.07 | 530.35 ± 10.81c |
| LSD | 11.384 | 5.0047 | 136.95 | 0.978 | 215.93 |
| ANOVA | p ≤ 0.0018 | p ≤ 0.0001 | p ≤ 0.0025 | NS | p ≤ 0.0001 |
| Cup plant | |||||
| Ph1 | 16.22 ± 0.07 b | 0.00 b | 46.63 ± 3.94 c | 28.43 ± 0.71 c | 157.07 ± 2.98 b |
| Ph2 | 18.62 ± 0.24 a | 0.00 b | 66.71 ± 5.36 b | 50.77 ± 3.28 a | 302.57 ± 15.42 a |
| Ph3 | 0.00c | 287.43 ± 0.01 a | 81.95 ± 1.45 a | 37.45 ± 0.06 b | 134.68 ± 4.12 b |
| LSD | 0.4479 | 0.0267 | 11.904 | 5.8643 | 28.381 |
| ANOVA | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 |
| Phenophases | Chl a (mg/g) | Chl b (mg/g) | TCh (mg/g) | TCa (mg/g) |
|---|---|---|---|---|
| Virginia Mallow | ||||
| Ph1 | 0.96 a | 0.47 ± 0.02 b | 1.43 ± 0.02 b | 0.37 a |
| Ph2 | 0.99 ± 0.01 a | 0.65 ± 0.03 a | 1.64 ± 0.05 a | 0.26 b |
| Ph3 | 0.75 ± 0.03 b | 0.56 ± 0.05 ab | 1.31 ± 0.07 b | 1.31 ± 0.02 b |
| LSD | 0.056 | 0.110 | 0.155 | 0.038 |
| ANOVA | p ≤ 0.0001 | p ≤ 0.0030 | p ≤ 0.0005 | p ≤ 0.0001 |
| Cup plant | ||||
| Ph1 | 0.59 a | 0.48 a | 1.07 a | 0.16 a |
| Ph2 | 0.58 ± 0.02 a | 0.53 ± 0.05 a | 1.11 ± 0.07 a | 0.15 a |
| Ph3 | 0.27 ± 0.02 b | 0.21 ± 0.02 b | 0.47 ± 0.02 b | 0.09 b |
| LSD | 0.048 | 0.086 | 0.134 | 0.010 |
| ANOVA | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 |
| Phenophase ID | Virginia Mallow | Cup Plant |
|---|---|---|
| Ph1—pre-flowering | 2 June | 17 June |
| Ph2—full flowering | 20 June | 27 June |
| Ph3—end of flowering | 13 July | 5 September |
| Standard | Calibration Curve Equation | R2 Value |
|---|---|---|
| Caffeic acid | y = 0.2137x − 0.0318 | 0.9997 |
| Coumaric acid | y = 0.2244x + 2.3792 | 0.9947 |
| Ellagic acid | y = 0.0338x + 7.0339 | 0.9309 |
| Ferulic acid | y = 0.3104x + 1.0316 | 0.9946 |
| Naringin | y = 0.1633x − 0.0833 | 0.9998 |
| Antioxidant Assay | Calibration Curve Equation | R2 Value |
|---|---|---|
| ABTS | y = −0.0002x + 0.614 | 0.8747 |
| FRAP | y = 0.0007x + 0.0116 | 0.9958 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šurić, J.; Šic Žlabur, J.; Peter, A.; Brandić, I.; Voća, S.; Dujmović, M.; Leto, J.; Voća, N. Energy vs. Nutritional Potential of Virginia Mallow (Sida hermaphrodita L.) and Cup Plant (Silphium perfoliatum L.). Plants 2022, 11, 2906. https://doi.org/10.3390/plants11212906
Šurić J, Šic Žlabur J, Peter A, Brandić I, Voća S, Dujmović M, Leto J, Voća N. Energy vs. Nutritional Potential of Virginia Mallow (Sida hermaphrodita L.) and Cup Plant (Silphium perfoliatum L.). Plants. 2022; 11(21):2906. https://doi.org/10.3390/plants11212906
Chicago/Turabian StyleŠurić, Jona, Jana Šic Žlabur, Anamarija Peter, Ivan Brandić, Sandra Voća, Mia Dujmović, Josip Leto, and Neven Voća. 2022. "Energy vs. Nutritional Potential of Virginia Mallow (Sida hermaphrodita L.) and Cup Plant (Silphium perfoliatum L.)" Plants 11, no. 21: 2906. https://doi.org/10.3390/plants11212906