
Phenology and Floral Biology of Diospyros sericea A. DC. (Ebenaceae): Inconstant Males May Be behind an Enigma of Dioecy

Abstract
:1. Introduction
2. Results
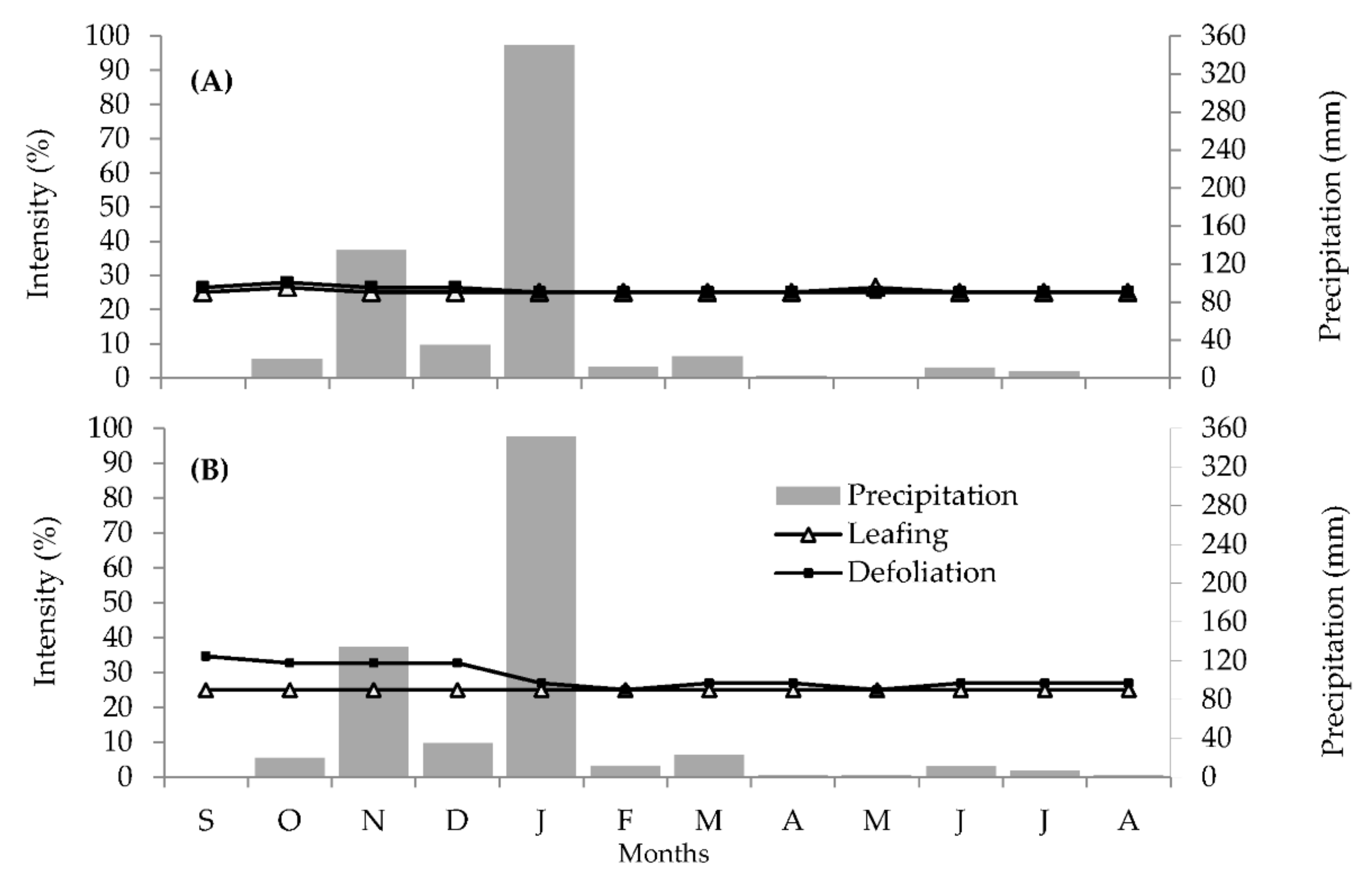
2.1. Phenological Analyses
2.1.1. Vegetative Phenology
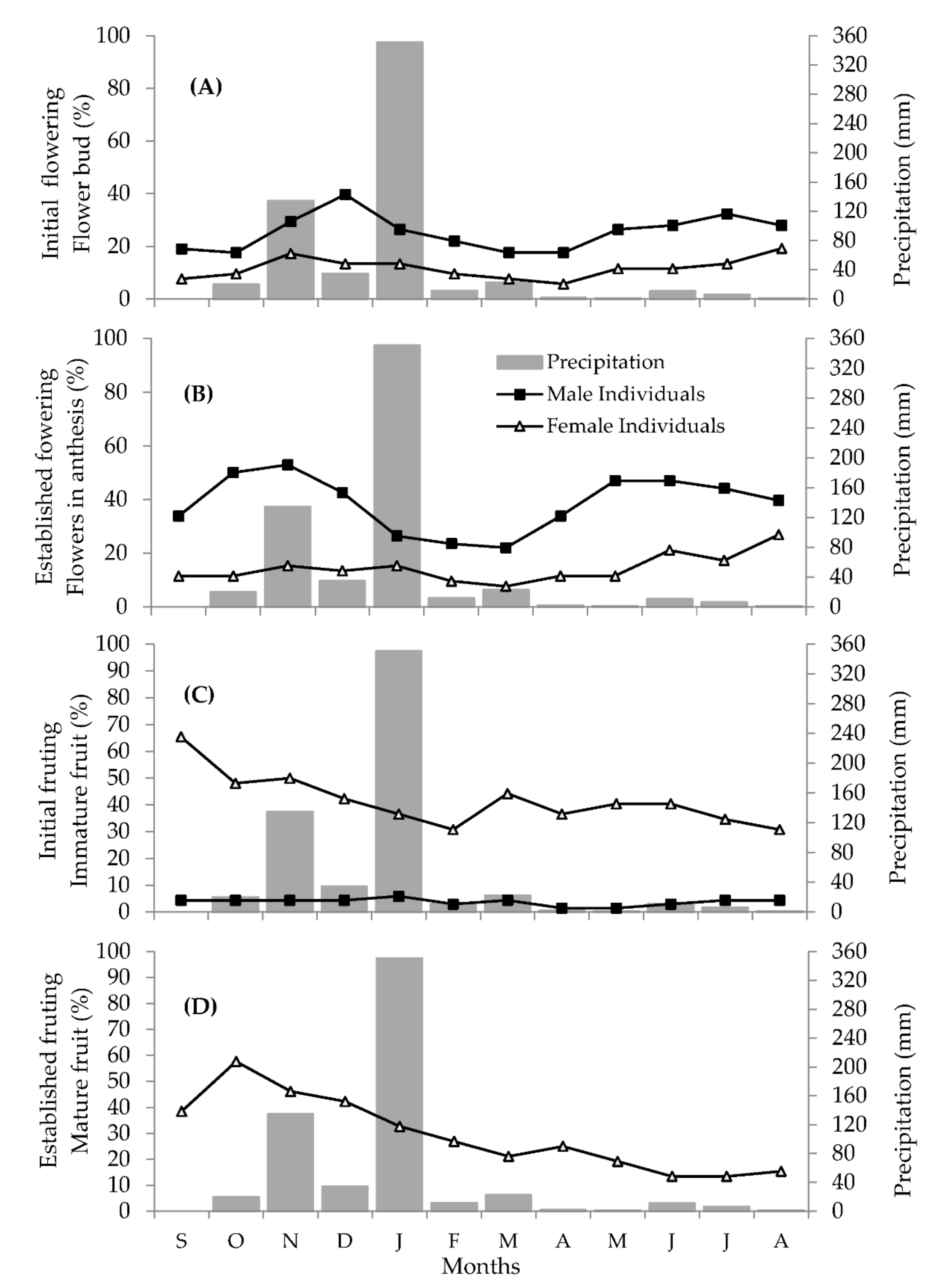
2.1.2. Reproductive Phenology
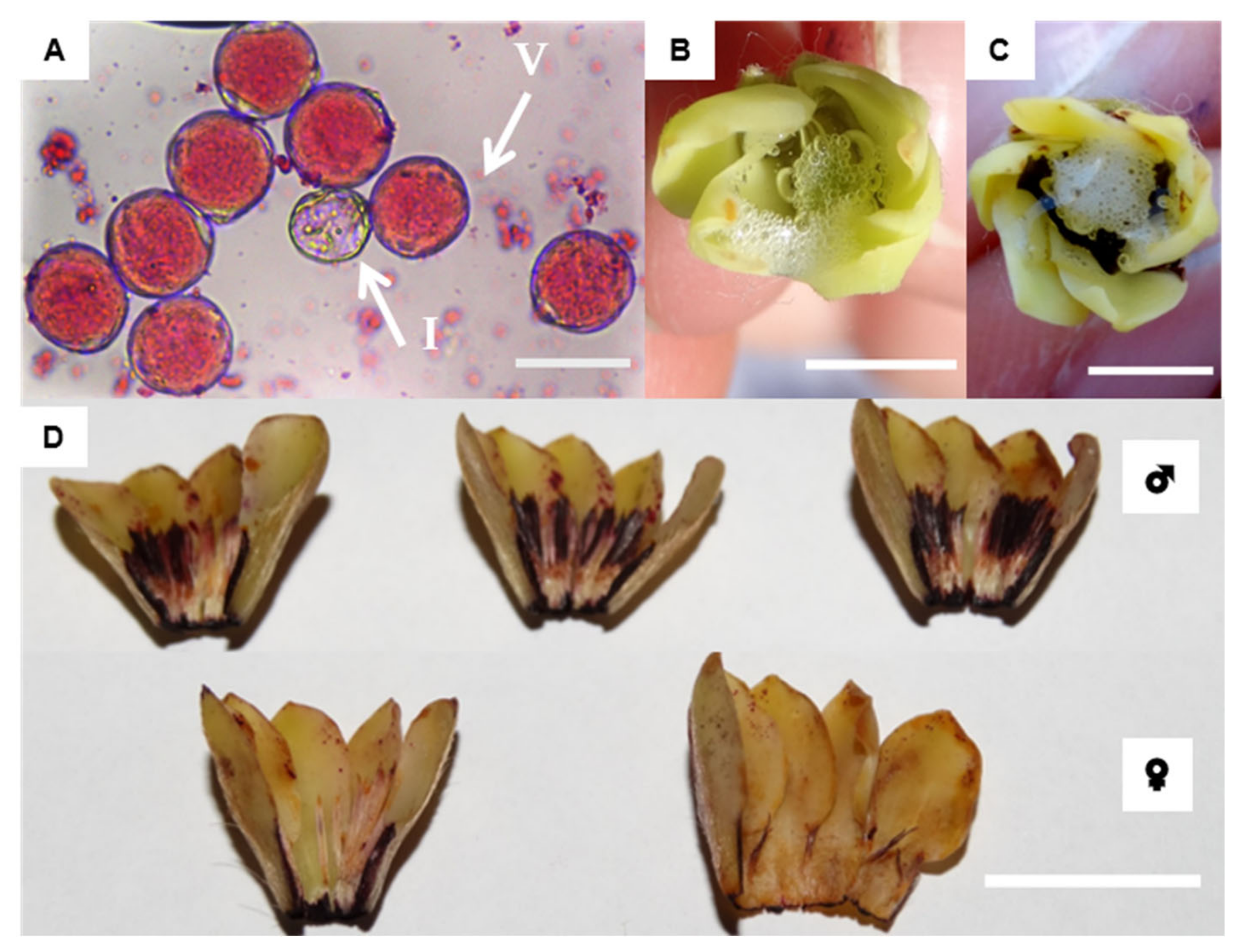
2.2. Floral Biology
2.2.1. Morphological Traits and Sexual Expression
2.2.2. Morphometrics
2.2.3. Floral Development
2.2.4. Pollen Integrity and Stigma Receptivity
2.2.5. Floral Attractants
3. Discussion
4. Materials and Methods
4.1. Study Site and Model Plant Species
4.2. Phenology
4.2.1. Data Collection
4.2.2. Data Analysis
4.3. Floral Biology
4.3.1. Morphometric Analyses
4.3.2. Pollen Integrity and Stigma Receptivity
4.3.3. Scent
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Crepet, W.L. The Abominable Mystery. Science 1998, 282, 1653–1654. [Google Scholar] [CrossRef]
- Barrett, S.C.H. The evolution of mating strategies in flowering plants. Trends Plant Sci. 1998, 3, 335–341. [Google Scholar] [CrossRef]
- Lloyd, D.G.; Barrett, S.C.H. (Eds.) Floral Biology: Studies on Floral Evolution in Animal-Pollinated Plants; Chapman & Hall: New York, NY, USA, 1996. [Google Scholar]
- Crepet, W.L. Progress in understanding angiosperm history, success, and relationships: Darwin’s abominably “perplexing phenomenon”. Proc. Natl. Acad. Sci. USA 2000, 97, 12939–12941. [Google Scholar] [CrossRef] [PubMed]
- Harder, L.D.; Barret, S.C.H. Pollen dispersal and mating patterns in animal-pollinated plants. In Floral Biology, Studies on Floral Evolution in Animal-Pollinated Plants; Chapman & Hall: New York, NY, USA, 1996; pp. 140–190. [Google Scholar]
- Cohen, D.; Shmida, A. The evolution of flower display and reward. Evol. Biol. 1993, 27, 197–243. [Google Scholar]
- Akagi, T.; Henry, I.M.; Tao, R.; Comai, L. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 2014, 346, 646–650. [Google Scholar] [CrossRef]
- Knight, T.A. XII. An account of some experiments on the fecundation of vegetables. In a letter from Thomas Andrew Knight, Esq. to the Right Hon. Sir Joseph Banks, K. B. P. R. S. Philos. Trans. R. Soc. Lond. 1799, 89, 195–204. [Google Scholar] [CrossRef]
- Ralls, K.; Frankham, R.; Ballou, J.D. Inbreeding and Outbreeding. Encycl. Biodivers. 2013, 2, 245–252. [Google Scholar] [CrossRef]
- Barrett, S.C.; Hough, J. Sexual dimorphism in flowering plants. J. Exp. Bot. 2012, 64, 67–82. [Google Scholar] [CrossRef]
- Barrett, S.C. Plant sex: Best to be bisexual when mates are scarce. Curr. Biol. 2021, 31, R298–R300. [Google Scholar] [CrossRef]
- Sakai, A.K.; Weller, S.G. Gender and sexual dimorphism in flowering plants: A review of terminology, biogeographic patterns, ecological correlates, and phylogenetic approaches. In Sexual and Gender Dimorphism in Flowering Plants; Geber, M.A., Dawson, T.E., Delph, L.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 1–31. [Google Scholar]
- Ainsworth, C. Boys and Girls Come Out to Play: The Molecular Biology of Dioecious Plants. Ann. Bot. 2000, 86, 211–221. [Google Scholar] [CrossRef]
- Akagi, T.; Charlesworth, D. Pleiotropic effects of sex-determining genes in the evolution of dioecy in two plant species. Proc. R. Soc. Boil. Sci. 2019, 286, 20191805. [Google Scholar] [CrossRef]
- Bawa, K.S. Evolution of dioecy in flowering plants. Annu. Rev. Ecol. Syst. 1980, 11, 15–39. [Google Scholar] [CrossRef]
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlates in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef]
- Renner, S.S. The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 2014, 101, 1588–1596. [Google Scholar] [CrossRef]
- Charlesworth, D. Plant sex determination and sex chromosomes. Heredity 2002, 88, 94–101. [Google Scholar] [CrossRef]
- Barrett, S.C.H. The evolution of plant reproductive systems: How often are transitions irreversible? Proc. R. Soc. Boil. Sci. 2013, 280, 20130913. [Google Scholar] [CrossRef]
- Charlesworth, B.; Charlesworth, D. A Model for the Evolution of Dioecy and Gynodioecy. Am. Nat. 1978, 112, 975–997. [Google Scholar] [CrossRef]
- Charlesworth, D. Theories of the Evolution of Dioecy. In Gender and Sexual Dimorphism in Flowering Plants; Geber, M.A., Dawson, T.E., Delph, L.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar] [CrossRef]
- Barrett, S.C.H. The evolution of plant sexual diversity. Nat. Rev. Genet. 2002, 3, 274–284. [Google Scholar] [CrossRef]
- Charlesworth, D. Does sexual dimorphism in plants promote sex chromosome evolution? Environ. Exp. Bot. 2018, 146, 5–12. [Google Scholar] [CrossRef]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex Chromosomes in Land Plants. Annu. Rev. Plant Biol. 2011, 62, 485–514. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D. Plant sex chromosome evolution. J. Exp. Bot. 2013, 64, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C.R. The Different Forms of Flowers on Plants of the Same Species; John Murray: London, UK, 1877. [Google Scholar]
- Munné-Bosch, S. Sex ratios in dioecious plants in the framework of global change. Environ. Exp. Bot. 2015, 109, 99–102. [Google Scholar] [CrossRef]
- Grant, V. Sexual selection in plants: Pros and cons. Proc. Natl. Acad. Sci. USA 1995, 92, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.K.; Wagner, W.L.; Ferguson, D.M.; Herbst, D.R. Biogeographical and Ecological Correlates of Dioecy in the Hawaiian Flora. Ecology 1995, 76, 2530–2543. [Google Scholar] [CrossRef]
- Oliveira, P.E. Dioecy in the Cerrado vegetation of Central Brazil. Flora 1996, 191, 235–243. [Google Scholar] [CrossRef]
- Webb, C.J.; Lloyd, D.G.; Delph, L.F. Gender dimorphism in indigenous New Zealand seed plants. N. Z. J. Bot. 1999, 37, 119–130. [Google Scholar] [CrossRef]
- Bawa, K.S.; Opler, P.A. Dioecism in tropical forest trees. Evolution 1975, 29, 167–179. [Google Scholar] [CrossRef]
- Givnish, T.J. Ecological constraints on the evolution of breeding systems in seed plants: Dioecy and dispersal in gymnosperms. Evolution 1980, 34, 959–972. [Google Scholar] [CrossRef]
- Weller, S.G.; Sakai, A.K. Using Phylogenetic Approaches for the Analysis of Plant Breeding System Evolution. Annu. Rev. Ecol. Syst. 1999, 30, 167–199. [Google Scholar] [CrossRef]
- Vamosi, J.C.; Otto, S.; Barrett, S.C.H. Phylogenetic analysis of the ecological correlates of dioecy in angiosperms. J. Evol. Biol. 2003, 16, 1006–1018. [Google Scholar] [CrossRef]
- Wallnöfer, B. The Biology and Systematics of Ebenaceae: A Review. Nat. Mus. Wien 2001, 103, 485–512. [Google Scholar]
- Santos, M.F.; Sano, P.T. Flora de Grão-Mogol, Minas Gerais: Ebenaceae. Bol. Bot. Univ. São Paulo 2004, 22, 93–95. [Google Scholar] [CrossRef]
- Santos, M.F.; Sano, P.T. Flora da Serra do Cipó, Minas Gerais: Ebenaceae. Bol. Bot. Univ. São Paulo 2018, 36, 23–28. [Google Scholar] [CrossRef]
- Duangjai, S.; Wallnöfer, B.; Samuel, R.; Munzinger, J.; Chase, M.W. Generic delimitation and relationships in Ebenaceae sensu lato: Evidence from six plastid DNA regions. Am. J. Bot. 2006, 93, 1808–1827. [Google Scholar] [CrossRef]
- Samuel, R.; Turner, B.; Duangjai, S.; Munzinger, J.; Paun, O.; Barfuss, M.H.J.; Chase, M.W. Systematics and evolution of the Old World Ebenaceae, a review with emphasis on the large genusDiospyrosand its radiation in New Caledonia. Bot. J. Linn. Soc. 2019, 189, 99–114. [Google Scholar] [CrossRef]
- Flora do Brasil. Jardim Botânico do Rio de Janeiro. 2020. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB7429 (accessed on 15 April 2021).
- Wallnöfer, B. A revision of neotropical Diospyros (Ebenaceae): Part 10. Nat. Mus. Wien 2017, 119, 183–226. [Google Scholar]
- Rebouças, N.C.; Cordeiro, L.S.; Araújo, R.S.; Ribeiro, R.T.M.; Loiola, M.I.B. Flora do Ceará, Brasil: Ebenaceae. Rodriguésia 2020, 71, e02122018. [Google Scholar] [CrossRef]
- Sothers, C. Diospyros cavalcantei (Ebenaceae): A New Species from Amazonia. Kew Bull. 2000, 55, 471. [Google Scholar] [CrossRef]
- De Campos, S.S.; Wittmann, M.T.S.; Veit, P.A.; Schwarz, S.F. Biologia Floral e viabilidade de pólen em cultivares de caquizeiro (Diospyros kaki L.) e Diospyros virginiania L. Rev. Bras. Frutic. 2015, 37, 685–691. [Google Scholar] [CrossRef]
- Lopes, R.C. Ebenaceae Vent. do Estado do Rio de Janeiro. Rev. Rodriguésia 1999, 50, 85–107. [Google Scholar] [CrossRef]
- Bawa, K.S. Patterns of flowering in tropical plants. In Handbook of Experimental Pollination Biology; Jones, C.E., Little, R.J., Eds.; Scientific and Academic Editions: New York, NY, USA, 1983; pp. 394–410. [Google Scholar]
- Lloyd, D.G.; Webb, C.J. Secondary sex characters in plants. Bot. Rev. 1977, 43, 177–216. [Google Scholar] [CrossRef]
- Forero-Montaña, J.; Zimmerman, J.K. Sexual dimorphism in the timing of flowering in two dioecious trees in a subtropical wet forest, Puerto Rico. Caribb. J. Sci. 2010, 46, 88–95. [Google Scholar] [CrossRef]
- Bullock, S.H.; Bawa, K.S. Sexual dimorphism and the annual flowering pattern in Jacaratia dolichaula (D. Smith) Woodson (Caricaceae) in a Costa Rican rain forest. Ecology 1981, 62, 1494–1504. [Google Scholar] [CrossRef]
- Rocca, M.A.; Sazima, M. The dioecious, sphingophilous species Citharexylum myrianthum (Verbenaceae): Pollination and visitor diversity. Flora 2016, 201, 440–450. [Google Scholar] [CrossRef]
- Otárola, M.F.; Sazima, M.; Solferini, V.N. Tree size and its relationship with flowering phenology and reproductive output in Wild Nutmeg trees. Ecol. Evol. 2013, 3, 3536–3544. [Google Scholar] [CrossRef] [PubMed]
- Delph, L.F.; Galloway, L.F.; Stanton, M.L. Sexual dimorphism in flower size. Am. Nat. 1996, 148, 299–320. [Google Scholar] [CrossRef]
- Murphy, C.G. Interaction-independent sexual selection and the mechanisms of sexual selection. Evolution 1998, 52, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Foresto, E.B. Levantamento Florístico dos Estratos Arbustivos e Arbóreo de Uma Mata de Galeria em Meio a Compôs Rupestres no Parque Estadual do Rio Preto, São Gonçalo do Rio Preto, MG. Master’s Thesis, Universidade de São Paulo, Instituto de Biociências, Departamento de Botânica, São Paulo, Brazil, 2018. [Google Scholar]
- Stanton, M.L. Male-male competition during pollination in plant populations. Am. Nat. 1994, 144, S40–S68. [Google Scholar] [CrossRef]
- Kudo, G. Flowering phenologies of animal-pollinated plants: Reproductive strategies and agents of selection. p. 139–158. In Ecology and Evolution of Flowers; Harder, L.D., Barrett, S.C.H., Eds.; Oxford University Press: New York, NY, USA, 1996; p. 370. [Google Scholar]
- Frankie, G.W.; Baker, H.G.; Opler, P.A. Comparative phonological studies of trees in tropical lowland wet and dry forest sites of Costa Rica. J. Ecol. 1947, 62, 881–913. [Google Scholar] [CrossRef]
- Grison-Pigé, L.; Bessière, J.-M.; Turlings, T.C.J.; Kjellberg, F.; Roy, J.; Hossaert-Mckey, M.M. Limited intersexmimicry of floral odour. Ficus Carica. Funct. Ecol. 2001, 15, 551–558. [Google Scholar] [CrossRef]
- Bawa, K.S. Plant-pollinator interactions in tropical rain forests. Annu. Rev. Ecol. Syst. 1990, 21, 399–422. [Google Scholar] [CrossRef]
- Renner, S.S.; Feil, J.P. Pollinators of Tropical Dioecious Angiosperms. Am. J. Bot. 1993, 80, 1100. [Google Scholar] [CrossRef] [Green Version]
- Baker, H.G. ‘‘Mistake’’ pollination as a reproductive system with special reference to the Caricaceae. Linn. Soc. Symp. Ser. 1976, 2, 161–169. [Google Scholar]
- Willson, M.F.; Ågren, J. Differential Floral Rewards and Pollination by Deceit in Unisexual Flowers. Oikos 1989, 55, 23. [Google Scholar] [CrossRef]
- Otárola, M.F.; Rocca, M.A. Flores no tempo: A floração como uma fase da fenologia reprodutiva. In Biologia da Polinização, 1st ed.; Rech, A.R., Agostini, K., Oliveira, P.E.., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; ISBN 978-85-68126-01-1. [Google Scholar]
- Barth, F.G. Insects and Flowers: The Biology of a Partnership; Princeton University Press: Princeton, NJ, USA, 1985. [Google Scholar]
- Schiestl, F.P. On the success of a swindle: Pollination by deception in orchids. Naturwissenschaften 2015, 92, 255–264. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L. Floral scent and intrafloral scent differentiation in Moneses and Pyrola (Pyrolaceae). Pl. Syst. Evol. 1991, 177, 81–91. [Google Scholar] [CrossRef]
- Proctor, M.; Yeo, P.; Lack, A. The Natural History of Pollination; Timber Press Inc.: Portland, OR, USA, 1996. [Google Scholar]
- Raguso, R.A. Why are some floral nectars scented? Ecology 2014, 85, 1486–1494. [Google Scholar] [CrossRef]
- Piratelli, A.J.; Piña-Rodrigues, F.C.M.; Gandara, F.B.; Santos, E.M.G.; Costa, L.G.S. Biologia da polinização de Jacaratia spinosa (AUBL) adc. (caricaceae) em mata residual do sudeste brasileiro. Rev. Bras. Biol. 1998, 58, 671–679. [Google Scholar] [CrossRef]
- Lenza, E.; Oliveira, P.E. Reprodução de Virola sebifera em mata mesofítica de Uberlândia, MG, Brasil. Rev. Bras. Bot. 2006, 29, 443–451. [Google Scholar] [CrossRef]
- Ashman, T.-L. Sniffing out patterns of sexual dimorphism in floral Scent. Funct. Ecol. 2009, 23, 852–862. [Google Scholar] [CrossRef]
- Aguiar, B.I.; Sebbenn, A.M.; Tarazi, R.; Vogado, N.O.; Morellato, L.P.C.; Tambarussi, E.V.; Moreno, M.A.; Pereira, L.C.S.M.; Montibeller, C.; Ferraz, E.M.; et al. Phenology, Seed Germination, and Genetics Explains the Reproductive Strategies of Diospyros lasiocalyx (Mart.) B. Wall. Trop. Plant Biol. 2020, 13, 23–35. [Google Scholar] [CrossRef]
- Reekie, E.; Bazzaz, F.A. Reproductive Allocation in Plants; Elsevier Academic Press: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Chapin, F.S., III; Schulze, E.-D.; Mooney, H.A. The storage and economics of storage in plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Escobar, D.E.; Silveira, F.A.O.; Morellato, L.P.C. Timing of seed dispersal and seed dormancy in Brazilian savanna: Two solutions to seasonality. Ann. Bot. 2018, 121, 1197–1209. [Google Scholar] [CrossRef]
- Batalha, M.A.; Martins, F.R. Reproductive phenology of the cerrado plant community in Emas National Park (Central Brazil). Aust. J. Bot. 2004, 52, 149–161. [Google Scholar] [CrossRef]
- Koltunow, A.M.; Grossniklaus, U. Apomixis: A developmental perspective. Annu. Rev. Plant Biol. 2003, 54, 547–574. [Google Scholar] [CrossRef]
- Bicknell, R.A.; Koltunow, A.M. Understanding apomixis: Recent advances and remaining conundrums. Plant Cell 2004, 16, S228–S245. [Google Scholar] [CrossRef]
- Tomlinson, P. Family: Ebenaceae. In The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016; pp. 242–243. [Google Scholar] [CrossRef]
- Ikeda, K.; Sugiura, A. Apomictic seed formation from inter- and intra- specific crosses of diospyros lotus. Acta Hortic. 2003, 601, 209–211. [Google Scholar] [CrossRef]
- Ashman, T.-L. The evolution of separate sexes: A focus on the ecological context. In Ecology and Evolution of Flowers; Harder, L.D., Barrett, S.H., Eds.; Oxford University Press: Oxford, UK, 2003; pp. 204–222. 390p. [Google Scholar]
- Lloyd, D.G. The distributions of gender in four angiosperm species illustrating two evolutionary pathways to dioecy. Evolution 1980, 34, 123–134. [Google Scholar] [CrossRef]
- Chagas, F.P.; Cabral, K.S.; Araújo, M.; Rocha, S.S. Proposta de ampliação do Parque Estadual Serra Nova e do Talhado; Instituto Estadual de Florestas: Belo Horizonte, Brazil, 1978. [Google Scholar]
- Wallnöfer, B.; Ebenaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. 2015. Available online: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB17314 (accessed on 15 April 2021).
- Marimon, B.S.; Felfili, J.M. Ethnobotanical comparison of “Pau Brasil” (Brosimum rubescens Taub.) forests in a xavante Indian and a non-xavante community in eastern Mato Grosso state, Brazil. Econ. Bot. 2001, 55, 555–569. [Google Scholar] [CrossRef]
- Funch, L.S.; Harley, R.; Funch, R.; Giulietti, A.M.; Melo, E. Plantas Úteis—Chapada Diamantina—Useful Plants; RiMa Editora: Sao Paulo, Brazil, 2004; p. 187. [Google Scholar]
- Forti, G.; Tambarussi, E.; Kageyama, P.; Moreno, M.; Ferraz, E.; Ibañes, B.; Vencovsky, R.; Mori, G.; Sebbenn, A. Low genetic diversity and intrapopulation spatial genetic structure of the Atlantic Forest tree, Esenbeckia leiocarpa Engl. (Rutaceae). Ann. For. Res. 2014, 57, 165–174. [Google Scholar] [CrossRef]
- Silvério, D.V.; Lenza, E. Phenology of woody species in a typical cerrado in the Bacaba Municipal Park, Nova Xavantina, Mato Grosso, Brazil. Biol. Neotrop. 2010, 10, 205–218. [Google Scholar] [CrossRef]
- Fournier, L.A. Un método cuantitativo para la medición de características fenológicas en árboles. Turrialba 1974, 24, 422–423. [Google Scholar]
- Morellato, L.P.C.; Leitão-Filho, H.F.; Rodrigues, R.R.; Joly, C.A. Estratégias fenológicas de espécies arbóreas em floresta de altitude na Serra do Japi, Jundiaí, São Paulo. Rev. Bras. Biol. 1990, 50, 149–162. [Google Scholar]
- Sarmiento, G.; Monasterio, M. Life forms and phenology. In Ecosystems of the World: Tropical Savannas; Bouliere, F., Ed.; Elsevier: Amsterdam, The Netherlands, 1983; pp. 79–108. [Google Scholar]
- Cesário, L.F.; Gaglianone, M.C. Biologia floral e fenologia reprodutiva de Schinus terebinthifolius Raddi (Anacardiaceae) em Restinga do Norte Fluminense. Acta Bot. Bras. 2008, 22, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Kearns, C.A.; Inouye, D.W. Techniques for Pollination Biologists; University Press of Colorado: Niwot, CO, USA, 1993; p. 583. [Google Scholar]
- Instituto Nacional de Meteorologia—INMET. Available online: http://www.inmet.gov.br/portal/index.php?r=home2/index (accessed on 20 September 2016).
- Ohya, I.; Nanami, S.; Itoh, A. Dioecious plants are more precocious than cosexual plants: A comparative study of relative sizes at the onset of sexual reproduction in woody species. Ecol. Evol. 2017, 7, 5660–5668. [Google Scholar] [CrossRef] [PubMed]
- Queenborough, S.A.; Mazer, S.J.; Vamosi, S.M.; Garwood, N.C.; Valencia, R.; Freckleton, R.P. Seed mass, abundance and breeding system among tropical forest species: Do dioecious species exhibit compensatory reproduction or abundances? J. Ecol. 2009, 97, 555–566. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenology | Individuals | Phenophases | Average Temperature | Relative Humidity | Precipitation |
|---|---|---|---|---|---|
| Vegetative | Male | Leafing | - | - | −0.16 |
| Defoliation | 0.75 * | −0.81 * | 0.21 | ||
| Female | Leafing | - | - | - | |
| Defoliation | 0.56 * | −0.75 * | 0.16 | ||
| Reproductive | Male | Initial flowering | −0.27 | - | 0.002 |
| Established flowering | −0.08 | −0.02 | 0.18 | ||
| Initial fruiting | 0.31 | −0.15 | 0.50 | ||
| Female | Initial flowering | −0.40 | 0.04 | −0.02 | |
| Established flowering | −0.05 | 0.03 | 0.27 | ||
| Initial fruiting | 0.52 | −0.43 | 0.16 | ||
| Established fruiting | 0.91 * | −0.60 * | 0.47 |
| Structures Measured | Male | Female | t-Test or U Test | p | ||
|---|---|---|---|---|---|---|
| ± SD | Min-Max Range | ± SD | Min-Max Range | |||
| Calyx | ||||||
| Length | 0.77 ± 0.077 | 0.59–0.94 | 0.80 ± 0.053 | 0.71–0.90 | t = −1.96 | 0.0549 |
| Diameter | 0.45 ± 0.046 | 0.32–0.53 | 0.53 ± 0.038 | 0.44–0.59 | t = −6.42 | <0.0001 * |
| Corolla | ||||||
| Length | 1.13 ± 0.107 | 0.98–1.35 | 1.09 ± 0.075 | 0.95–1.21 | t = 1.88 | 0.0655 |
| Diameter | 0.43 ± 0.054 | 0.30–0.55 | 0.48 ± 0.050 | 0.34–0.59 | t = −3.30 | 0.0018 * |
| Pistil/pistillodes | ||||||
| Length | 0.12 ± 0.087 | 0.06–0.52 | 0.68 ± 0.083 | 0.39–0.79 | t = −23.62 | <0.0001 * |
| Diameter | 0.13 ± 0.043 | 0.08–0.23 | 0.33 ± 0.022 | 0.30–0.41 | U = 0.00 | <0.0001 * |
| Stamens/staminodes | ||||||
| Length | 0.62 ± 0.056 | 0.49–0.72 | 0.43 ± 0.042 | 0.35–0.50 | t = 13.47 | <0.0001 * |
| Number | 43.07 ± 4.811 | 34–54 | 9.35 ± 3.908 | 4–15 | U = 0.00 | <0.0001 * |
| Number of buds per axil | 2.18 ± 0.566 | 1–7 | 1 | 1 | - | - |
| Floral Development Stage | Min–Max Range | ± SD | Pollen Integrity (%) |
|---|---|---|---|
| Flower buds | 163–197 | 182.4 ± 12.72 | 91.2% |
| Flowers in pre-anthesis | 167–197 | 186.0 ± 9.043 | 93% |
| Open flowers | 177–197 | 187.7 ± 6.667 | 94% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramaldes, B.; Santos, R.; Rech, A.R.; Soares, M. Phenology and Floral Biology of Diospyros sericea A. DC. (Ebenaceae): Inconstant Males May Be behind an Enigma of Dioecy. Plants 2022, 11, 2535. https://doi.org/10.3390/plants11192535
Ramaldes B, Santos R, Rech AR, Soares M. Phenology and Floral Biology of Diospyros sericea A. DC. (Ebenaceae): Inconstant Males May Be behind an Enigma of Dioecy. Plants. 2022; 11(19):2535. https://doi.org/10.3390/plants11192535
Chicago/Turabian StyleRamaldes, Bárbara, Renata Santos, André Rodrigo Rech, and Michellia Soares. 2022. "Phenology and Floral Biology of Diospyros sericea A. DC. (Ebenaceae): Inconstant Males May Be behind an Enigma of Dioecy" Plants 11, no. 19: 2535. https://doi.org/10.3390/plants11192535