
Effects of a Whole Plant Extract of Scutellaria rubropunctata var. rubropunctata on Bone Metabolism with Estrogen Receptor Activation

 , , and
, , and 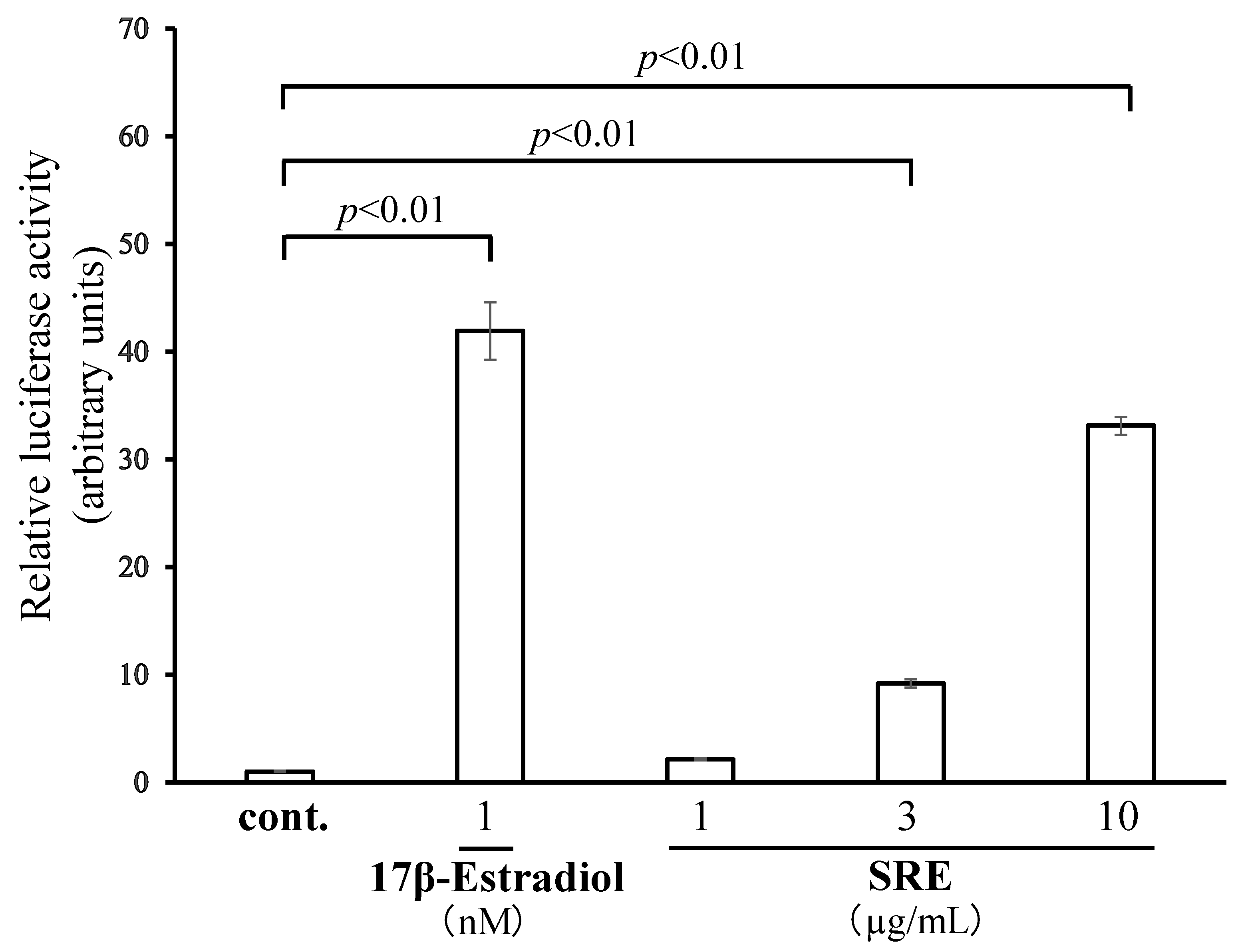{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SRE Has ER Transcription-Promoting Activity
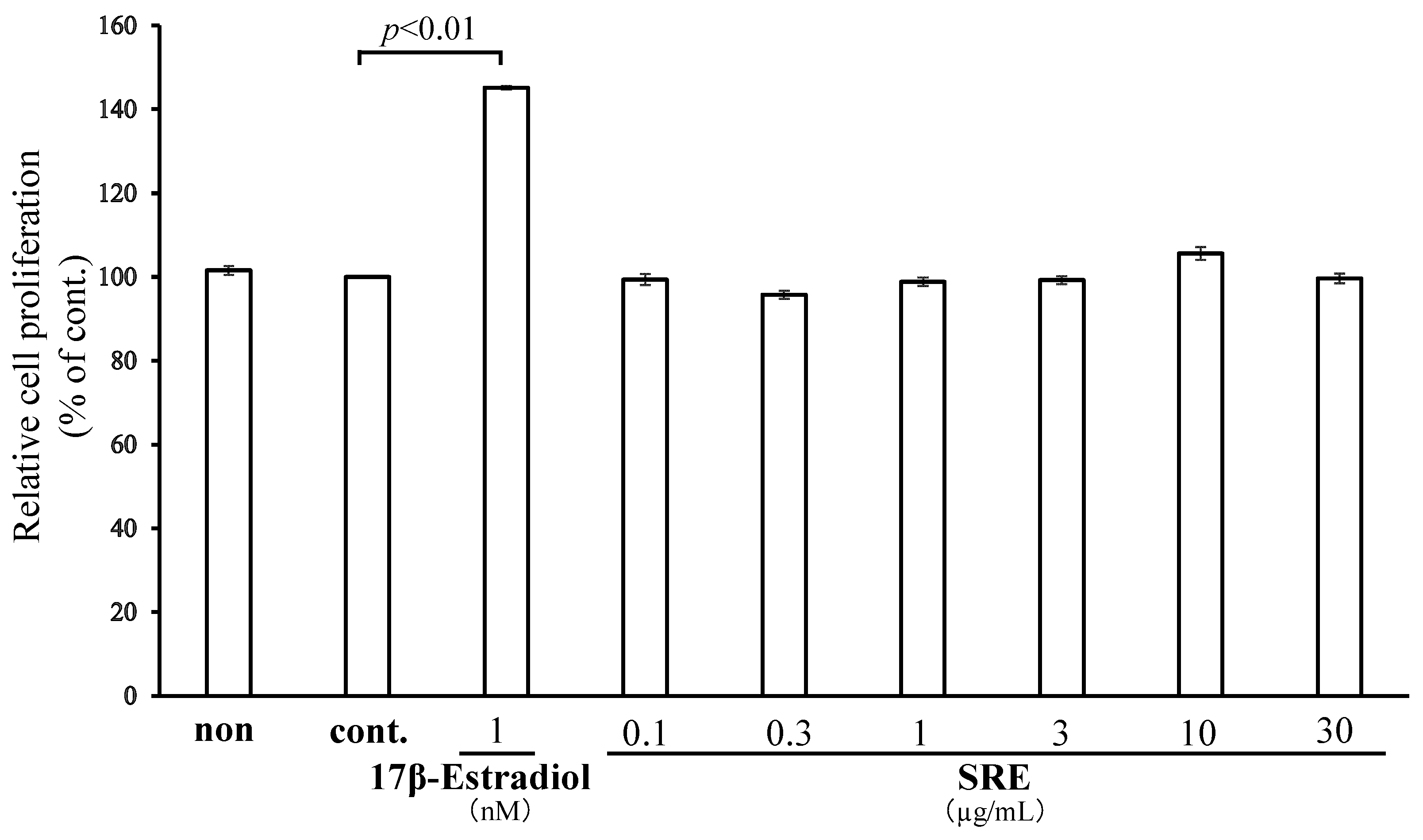
2.2. SRE Does Not Affect MCF7 Cell Proliferation
2.3. Effect of SRE on Osteoblastic Differentiation
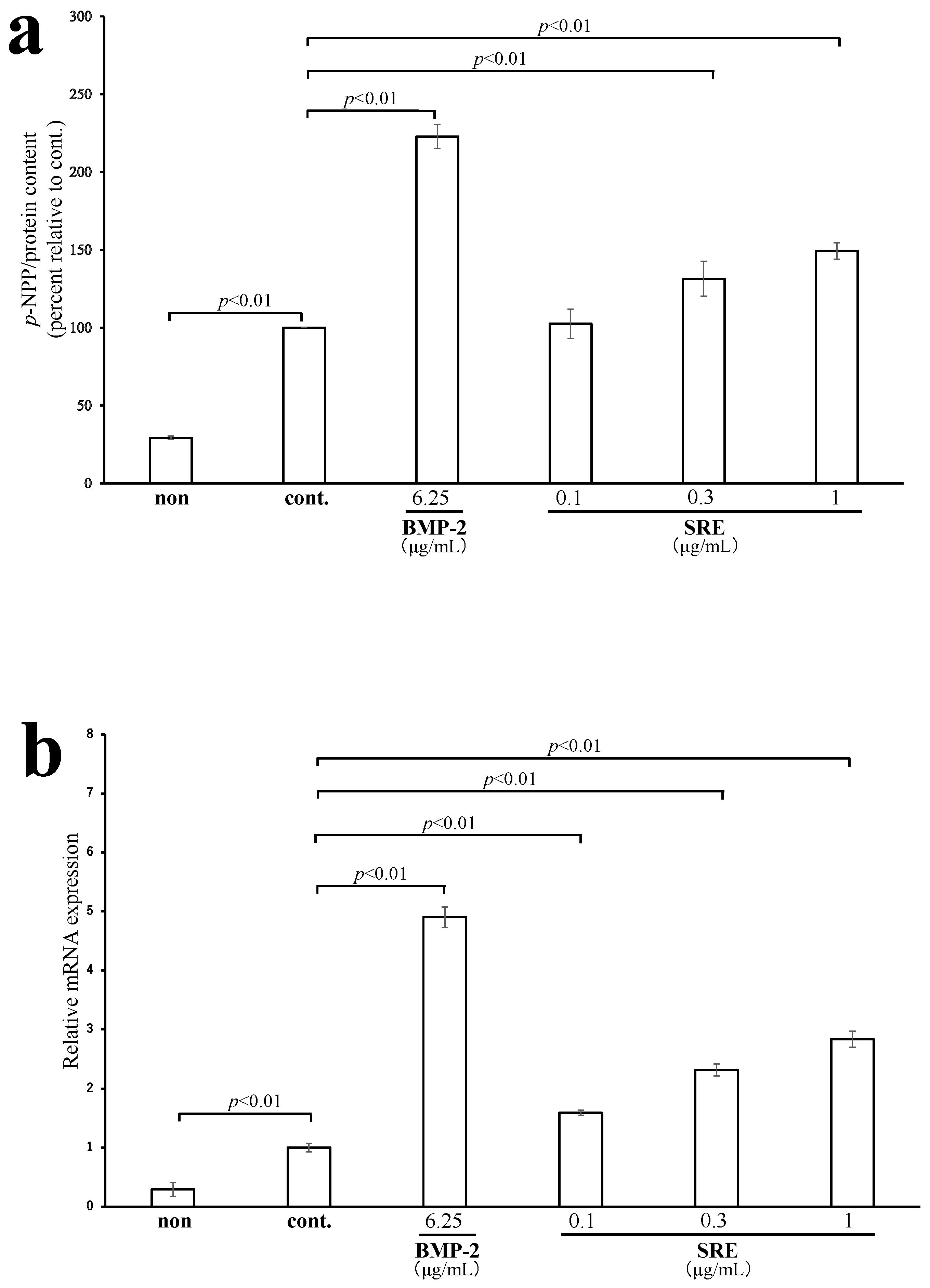
2.3.1. SRE Increased ALP Activity and the mRNA Expression of Alpl
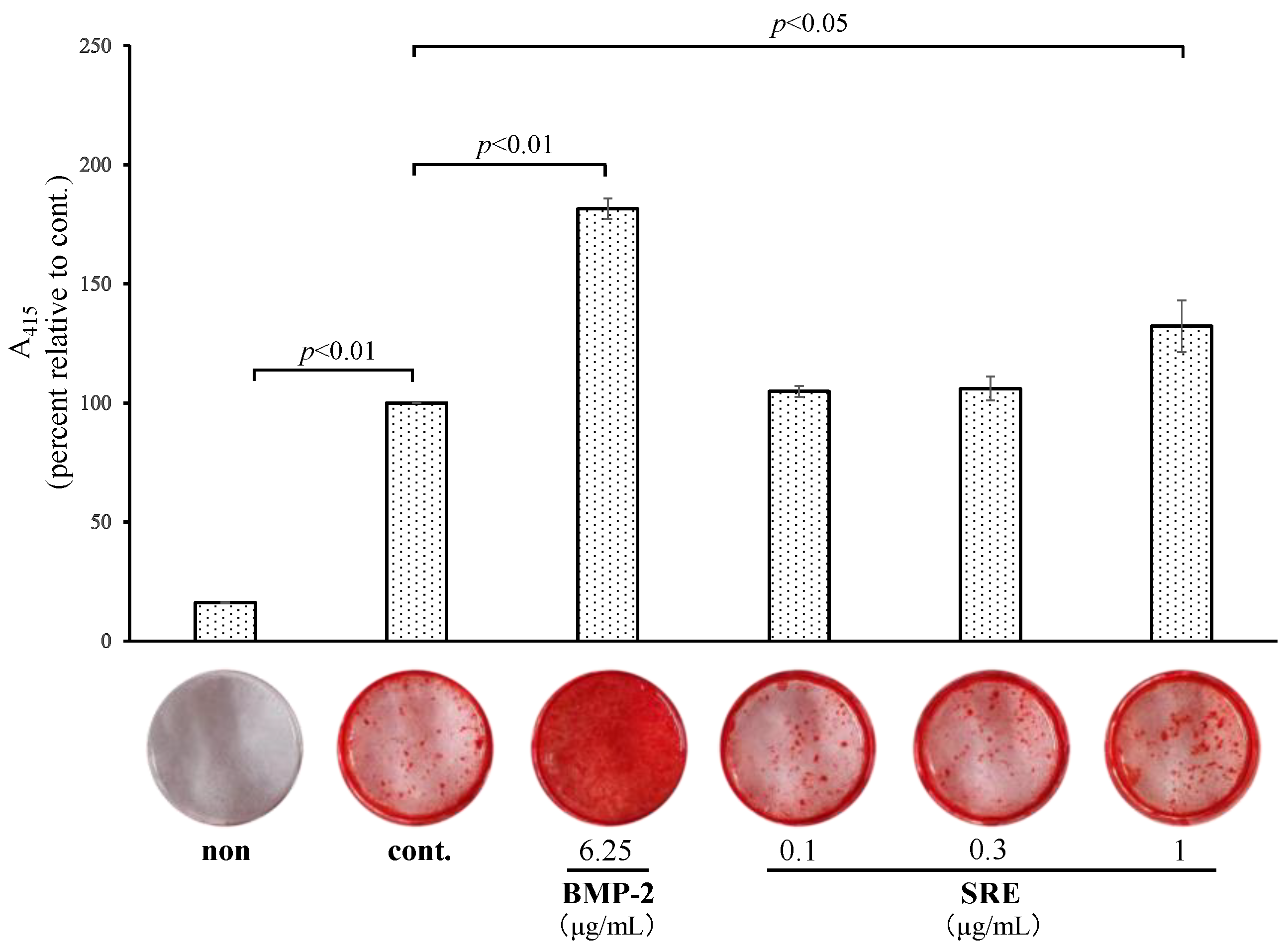
2.3.2. SRE Increased Mineralization
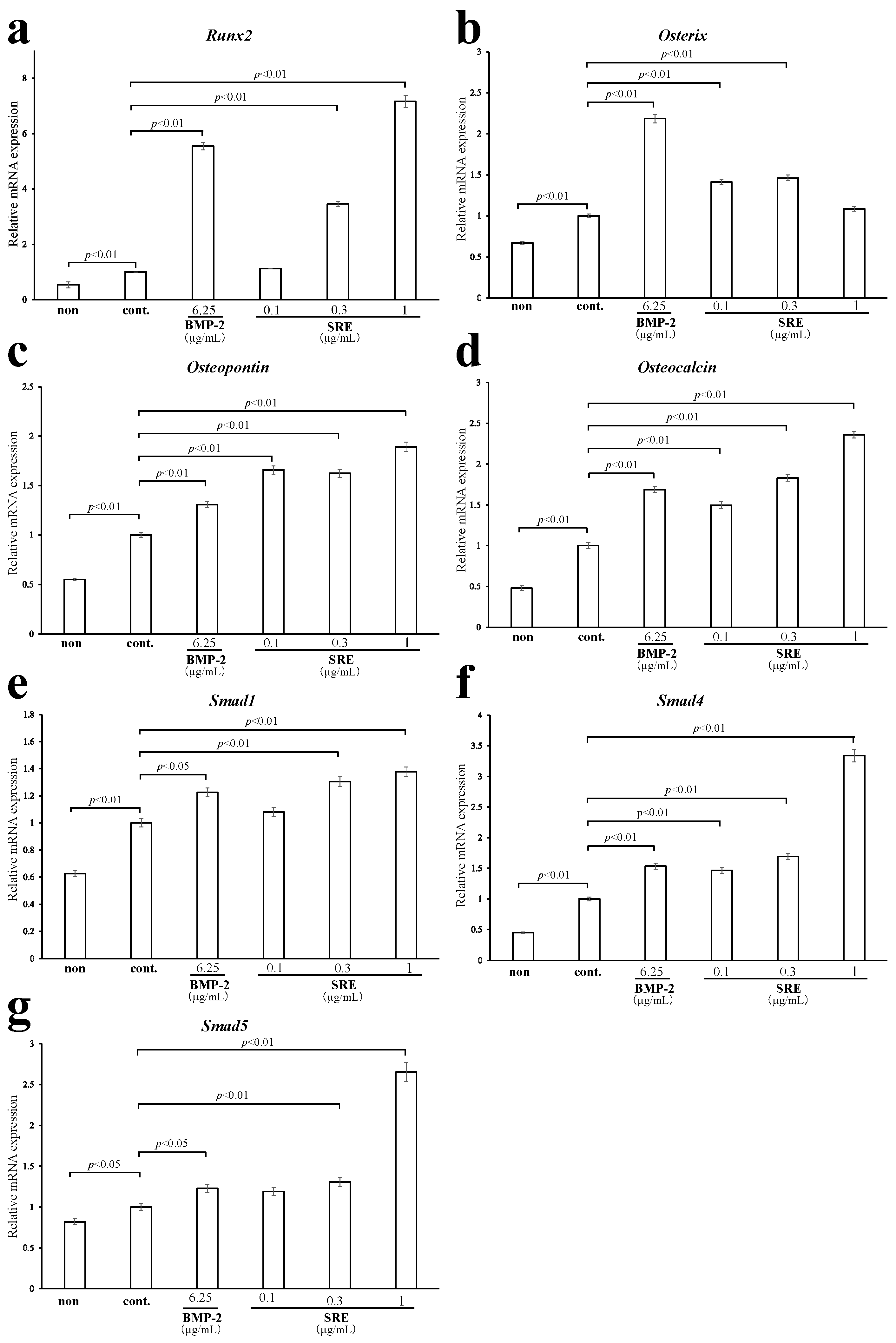
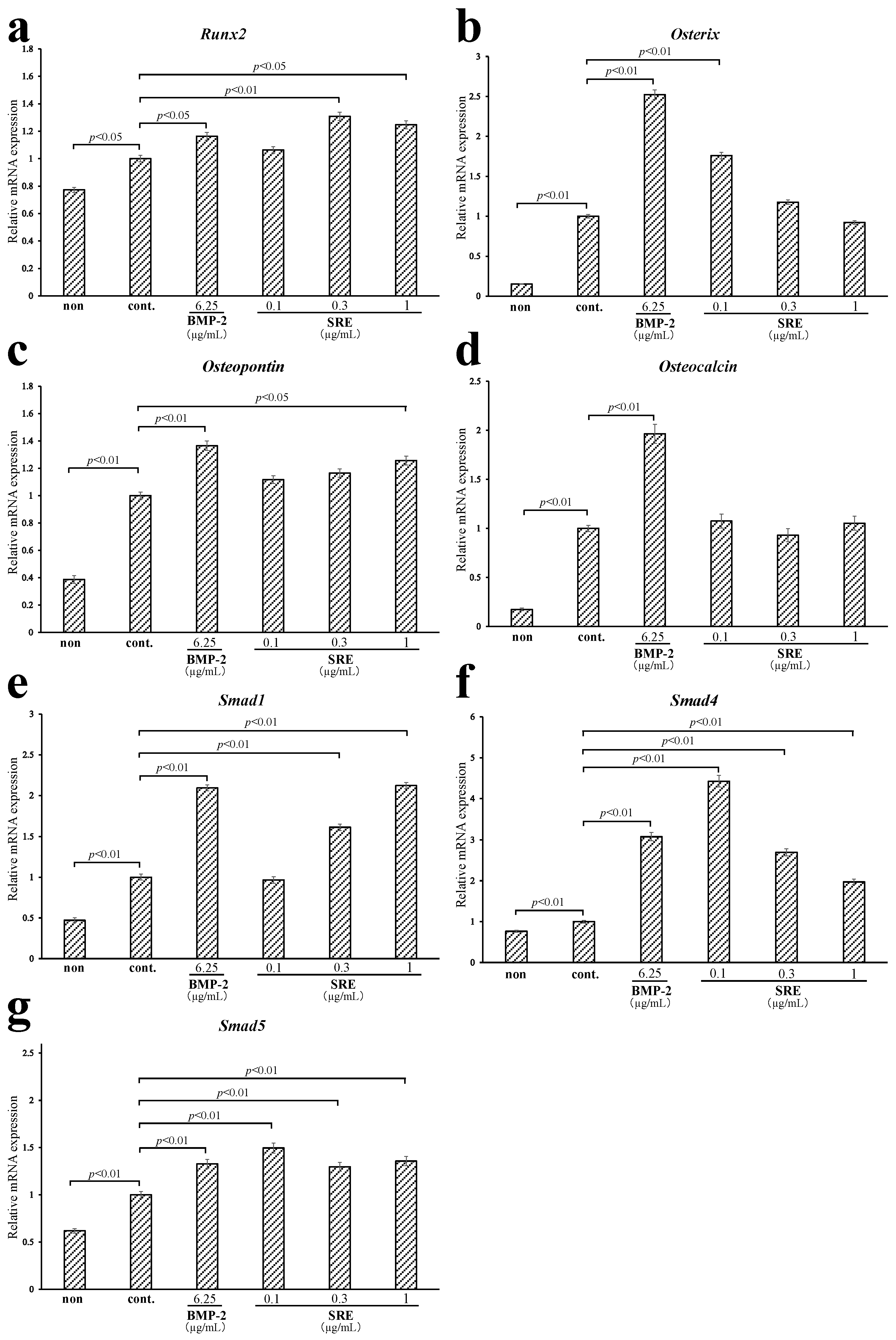
2.3.3. Effect of SRE on the Expression of Osteoblast-Differentiation-Related Genes
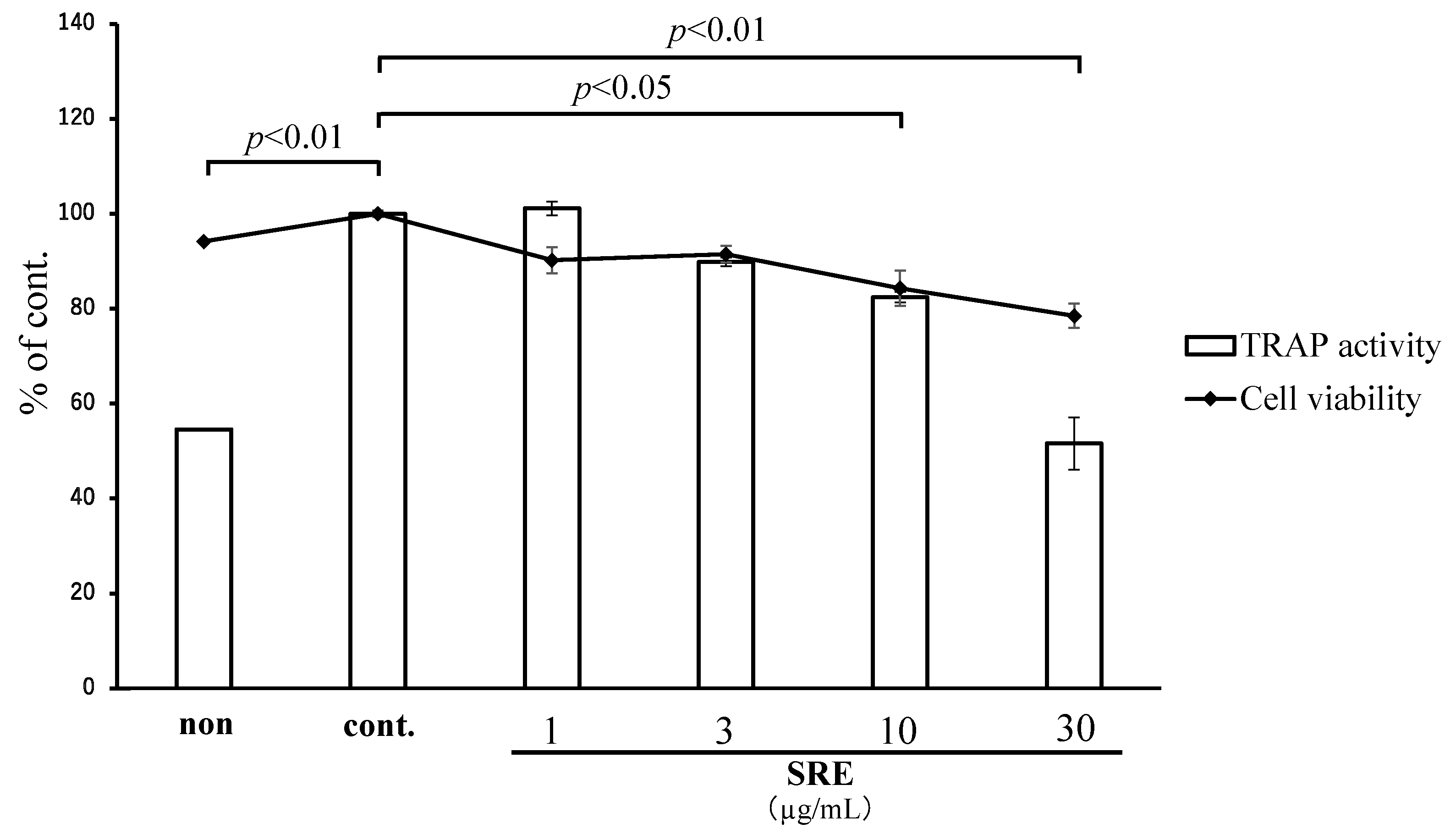
2.4. Effect of SRE on Osteoclastic Differentiation
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extract Preparation
4.2. Cell Lines and Cell Culture
4.3. Luciferase Reporter Assays of ER Transcription Activity
4.4. ALP Activity Assay
4.5. Mineralization Assay
4.6. TRAP Activity Assay
4.7. Measurement of Cell Proliferation by the MTT Assay
4.8. Quantitative Real-Time PCR
4.9. HPLC Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burger, H.G.; Dudley, E.C.; Robertson, D.M.; Dennerstein, L. Hormonal changes in the menopause transition. Recent Prog. Horm. Res. 2002, 57, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Krege, J.H.; Wan, X. Teriparatide and the risk of nonvertebral fractures in women with postmenopausal osteoporosis. Bone 2012, 50, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, F.; Cadogan, M.P. Recognition and treatment of postmenopausal osteoporosis. J. Gerontol. Nurs. 2014, 40, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.E.; Dai, A.; Tiffee, J.C.; Li, H.H.; Mundy, G.R.; Boyce, B.F. Estrogen promotes apoptosis of murine osteoclasts mediated by TGF-beta. Nat. Med. 1996, 2, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Heino, T.J.; Hentunen, T.A.; Väänänen, H.K. Osteocytes inhibit osteoclastic bone resorption through transforming growth factor-beta: Enhancement by estrogen. J. Cell. Biochem. 2002, 85, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Imai, Y.; Matsumoto, T.; Sato, S.; Takeuchi, K.; Igarashi, K.; Harada, Y.; Azuma, Y.; Krust, A.; Yamamoto, Y.; et al. Estrogen prevents bone loss via estrogen receptor alpha and induction of Fas ligand in osteoclasts. Cell 2007, 130, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Oursler, M.J.; Cortese, C.; Keeting, P.; Anderson, M.A.; Bonde, S.K.; Riggs, B.L.; Spelsberg, T.C. Modulation of transforming growth factor-beta production in normal human osteoblast-like cells by 17 beta-estradiol and parathyroid hormone. Endocrinology 1991, 129, 3313–3320. [Google Scholar] [CrossRef]
- Bord, S.; Beavan, S.; Ireland, D.; Horner, A.; Compston, J.E. Mechanisms by which high-dose estrogen therapy produces anabolic skeletal effects in postmenopausal women: Role of locally produced growth factors. Bone 2001, 29, 216–222. [Google Scholar] [CrossRef]
- Finkelman, R.D.; Bell, N.H.; Strong, D.D.; Demers, L.M.; Baylink, D.J. Ovariectomy selectively reduces the concentration of transforming growth factor beta in rat bone: Implications for estrogen deficiency-associated bone loss. Proc. Natl. Acad. Sci. USA 1992, 89, 12190–12193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, D.; Rubin, S.M.; Petitti, D.B.; Fox, C.S.; Black, D.; Ettinger, B.; Ernster, V.L.; Cummings, S.R. Hormone therapy to prevent disease and prolong life in postmenopausal women. Ann. Intern. Med. 1992, 117, 1016–1037. [Google Scholar] [CrossRef] [PubMed]
- Beral, V.; Million Women Study Collaborators. Breast cancer and hormone-replacement therapy in the Million Women Study. Lancet 2003, 362, 419–427. [Google Scholar] [CrossRef]
- Cos, P.; De Bruyne, T.; Apers, S.; Berghe, D.V.; Pieters, L.; Vlietinck, A.J. Phytoestrogens: Recent developments. Planta Med. 2003, 69, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picherit, C.; Coxam, V.; Bennetau-Pelissero, C.; Kati-Coulibaly, S.; Davicco, M.J.; Lebecque, P.; Barlet, J.P. Daidzein is more efficient than genistein in preventing ovariectomy-induced bone loss in rats. J. Nutr. 2000, 130, 1675–1681. [Google Scholar] [CrossRef] [PubMed]
- Bitto, A.; Burnett, B.P.; Polito, F.; Marini, H.; Levy, R.M.; Armbruster, M.A.; Minutoli, L.; Di Stefano, V.; Irrera, N.; Antoci, S.; et al. Effects of genistein aglycone in osteoporotic, ovariectomized rats: A comparison with alendronate, raloxifene and oestradiol. Br. J. Pharmacol. 2008, 155, 896–905. [Google Scholar] [CrossRef]
- Wong, R.; Rabie, B. Effect of puerarin on bone formation. Osteoarthr. Cartil. 2007, 15, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, H.; Kadota, Y.; Murata, J.; Yonekura, K.; Kihara, H. Wild Flowers of Japan Volume 5 Convolvulaceae~Caprifoliaceae; Heibonsha Limited, Publishers: Tokyo, Japan, 2017; pp. 115–119. ISBN 978-4-582-53535-8. [Google Scholar]
- Liu, W.; Toyosawa, S.; Furuichi, T.; Kanatani, N.; Yoshida, C.; Liu, Y.; Himeno, M.; Narai, S.; Yamaguchi, A.; Komori, T. Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures. J. Cell. Biol. 2001, 155, 157–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, T.A.; Aronow, M.; Shalhoub, V.; Barone, L.M.; Wilming, L.; Tassinari, M.S.; Kennedy, M.B.; Pockwinse, S.; Lian, J.B.; Stein, G.S. Progressive development of the rat osteoblast phenotype in vitro: Reciprocal relationships in expression of genes associated with osteoblast proliferation and differentiation during formation of the bone extracellular matrix. J. Cell. Physiol. 1990, 143, 420–430. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhry, M.; Hamade, E.; Badran, B.; Buchet, R.; Magne, D. Molecular mechanisms of mesenchymal stem cell differentiation towards osteoblasts. World J. Stem Cells 2013, 5, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon Med. Sch. 2010, 77, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodine, P.V.; Henderson, R.A.; Green, J.; Aronow, M.; Owen, T.; Stein, G.S.; Lian, J.B.; Komm, B.S. Estrogen receptor-alpha is developmentally regulated during osteoblast differentiation and contributes to selective responsiveness of gene expression. Endocrinology 1998, 139, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A transcriptional activator of osteoblast differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Chen, G.; Li, Y.P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Souchelnytskyi, S.; Heldin, C.H. Smad regulation in TGF-beta signal transduction. J. Cell Sci. 2001, 114 Pt 24, 4359–4369. [Google Scholar] [CrossRef] [PubMed]
- Majeska, R.J.; Ryaby, J.T.; Einhorn, T.A. Direct modulation of osteoblastic activity with estrogen. J. Bone Jt. Surg. Am. 1994, 76, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Kousteni, S.; Almeida, M.; Han, L.; Bellido, T.; Jilka, R.L.; Manolagas, S.C. Induction of osteoblast differentiation by selective activation of kinase-mediated actions of the estrogen receptor. Mol. Cell. Biol. 2007, 27, 1516–1530. [Google Scholar] [CrossRef] [Green Version]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Halleen, J.M.; Alatalo, S.L.; Suominen, H.; Cheng, S.; Janckila, A.J.; Väänänen, H.K. Tartrate-resistant acid phosphatase 5b: A novel serum marker of bone resorption. J. Bone Miner. Res. 2000, 15, 1337–1345. [Google Scholar] [CrossRef]
- Shane, E.; Burr, D.; Ebeling, P.R.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; et al. Atypical subtrochanteric and diaphyseal femoral fractures: Report of a task force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2010, 25, 2267–2294. [Google Scholar] [CrossRef]
- Shane, E.; Burr, D.; Abrahamsen, B.; Adler, R.A.; Brown, T.D.; Cheung, A.M.; Cosman, F.; Curtis, J.R.; Dell, R.; Dempster, D.W.; et al. Atypical subtrochanteric and diaphyseal femoral fractures: Second report of a task force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2014, 29, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Rotem, C.; Kaplan, B. Phyto-Female Complex for the relief of hot flushes, night sweats and quality of sleep: Randomized, controlled, double-blind pilot study. Gynecol. Endocrinol. 2007, 23, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Van Die, M.D.; Burger, H.G.; Teede, H.J.; Bone, K.M. Vitex agnus-castus (Chaste-Tree/Berry) in the treatment of menopause-related complaints. J. Altern. Complement. Med. 2009, 15, 853–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stovall, D.W.; Utian, W.H.; Gass, M.L.; Qu, Y.; Muram, D.; Wong, M.; Plouffe, L., Jr. The effects of combined raloxifene and oral estrogen on vasomotor symptoms and endometrial safety. Menopause 2007, 14 Pt 1, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, R.; Gallagher, J.C.; Kagan, R.; Pickar, J.H.; Constantine, G. Efficacy of tissue-selective estrogen complex of bazedoxifene/conjugated estrogens for osteoporosis prevention in at-risk postmenopausal women. Fertil. Steril. 2009, 92, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Pickar, J.H.; Yeh, I.T.; Bachmann, G.; Speroff, L. Endometrial effects of a tissue selective estrogen complex containing bazedoxifene/conjugated estrogens as a menopausal therapy. Fertil. Steril. 2009, 92, 1018–1024. [Google Scholar] [CrossRef]
- Pinkerton, J.V.; Utian, W.H.; Constantine, G.D.; Olivier, S.; Pickar, J.H. Relief of vasomotor symptoms with the tissue-selective estrogen complex containing bazedoxifene/conjugated estrogens: A randomized, controlled trial. Menopause 2009, 16, 1116–1124. [Google Scholar] [CrossRef]
- Warren, M.P. A tissue selective estrogen complex: A new horizon for menopausal symptoms. Menopause 2009, 16, 1083–1084. [Google Scholar] [CrossRef]
- Lobo, R.A.; Pinkerton, J.V.; Gass, M.L.S.; Dorin, M.H.; Ronkin, S.; Pickar, J.H.; Constantine, G. Evaluation of bazedoxifene/conjugated estrogens for the treatment of menopausal symptoms and effects on metabolic parameters and overall safety profile. Fertil. Steril. 2009, 92, 1025–1038. [Google Scholar] [CrossRef]
- Wang, Z.L.; Wang, S.; Kuang, Y.; Hu, Z.M.; Qiao, X.; Ye, M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, M.; Yahagi, T.; Shirayama, T.; Miyake, K.; Kotani, H.; Ogawa, T.; Matsuzaki, K. Effects of a Whole Plant Extract of Scutellaria rubropunctata var. rubropunctata on Bone Metabolism with Estrogen Receptor Activation. Plants 2022, 11, 2075. https://doi.org/10.3390/plants11162075
Watanabe M, Yahagi T, Shirayama T, Miyake K, Kotani H, Ogawa T, Matsuzaki K. Effects of a Whole Plant Extract of Scutellaria rubropunctata var. rubropunctata on Bone Metabolism with Estrogen Receptor Activation. Plants. 2022; 11(16):2075. https://doi.org/10.3390/plants11162075
Chicago/Turabian StyleWatanabe, Misaki, Tadahiro Yahagi, Takahiro Shirayama, Katsunori Miyake, Hitoshi Kotani, Takuya Ogawa, and Keiichi Matsuzaki. 2022. "Effects of a Whole Plant Extract of Scutellaria rubropunctata var. rubropunctata on Bone Metabolism with Estrogen Receptor Activation" Plants 11, no. 16: 2075. https://doi.org/10.3390/plants11162075