
Genetic Variation and Genotype by Environment Interaction for Agronomic Traits in Maize (Zea mays L.) Hybrids

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Genetic Materials
2.2. Experimental Site and Design
2.3. Experimental Details
2.4. Evaluation of Agronomic Traits
2.5. Statistical Analysis
2.5.1. Analysis of Variance
2.5.2. Variability Estimates
Phenotypic and Genotypic Variance
Estimation of Heritability
Estimation of Genetic Advance
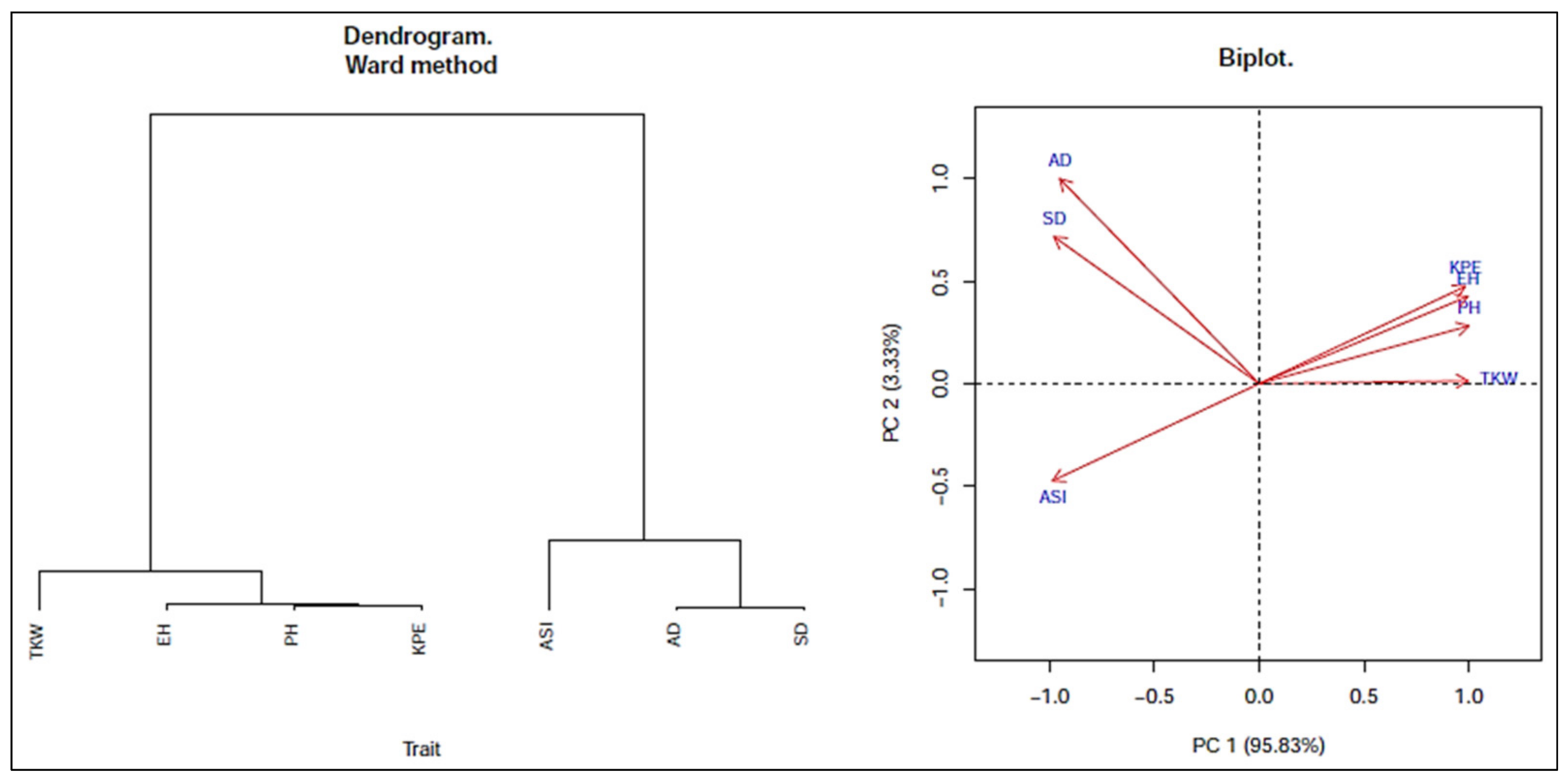
Association Analysis
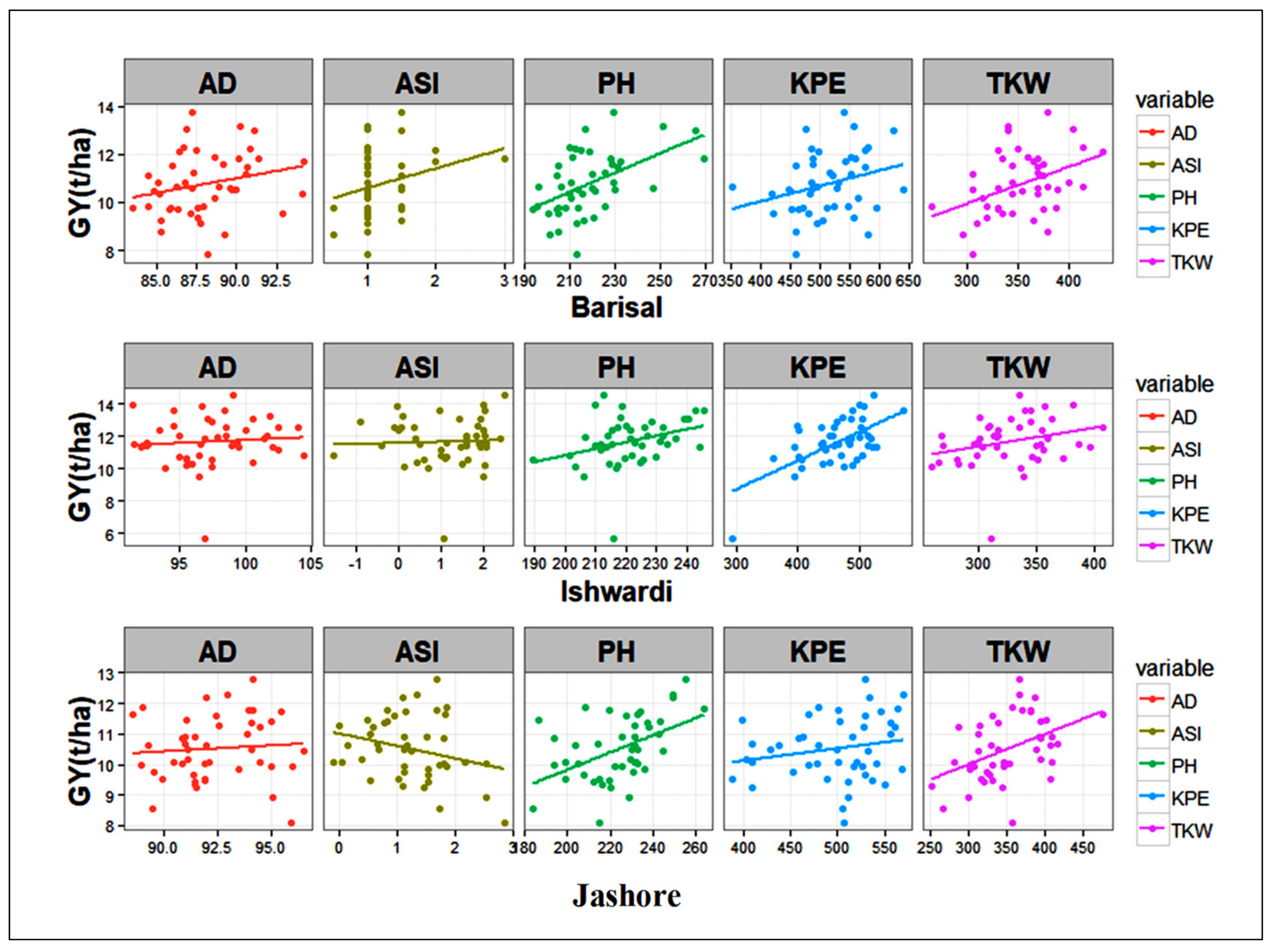
Regression Analysis
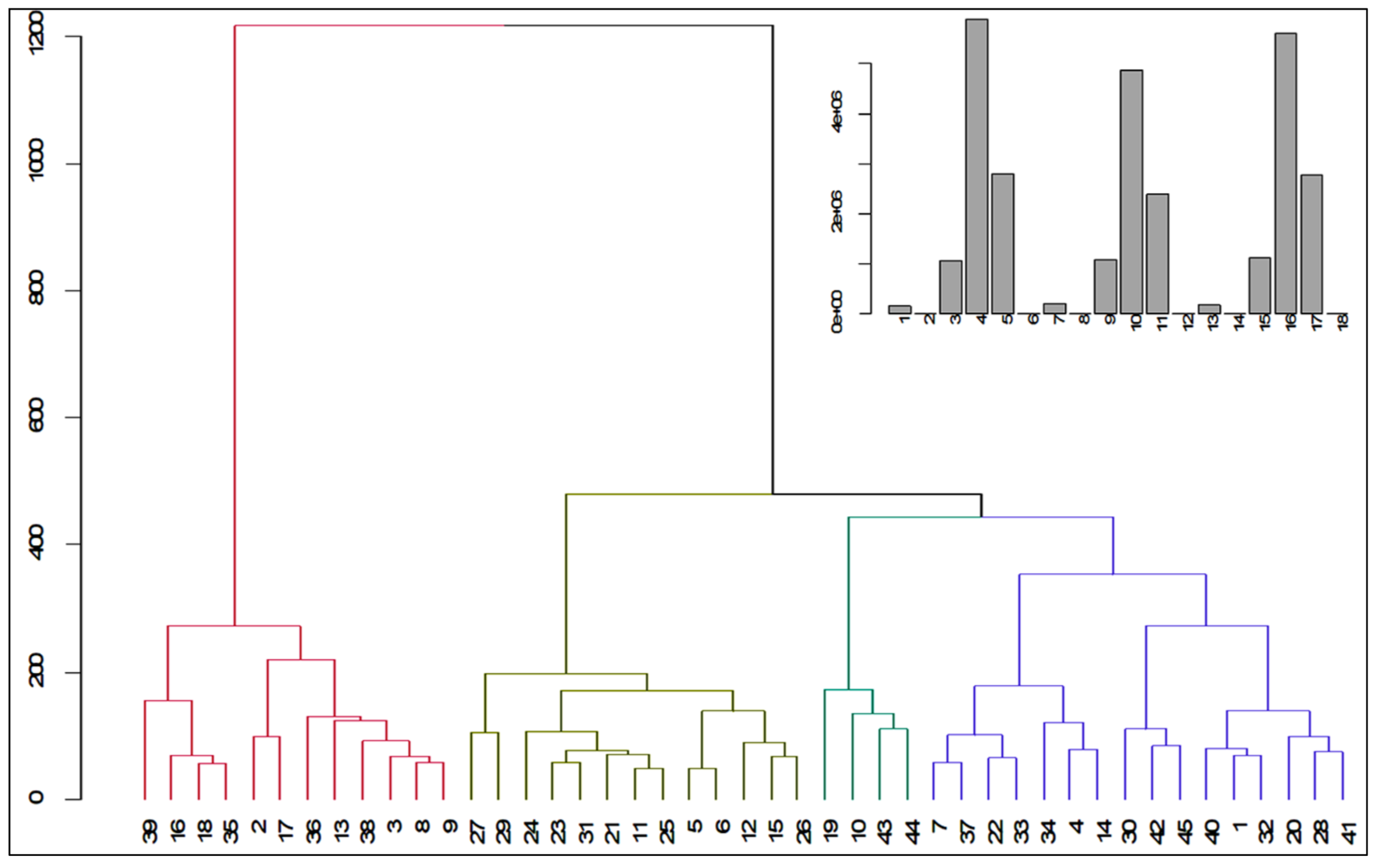
Grouping or Clustering
3. Results and Discussion
3.1. Genetic Variability among Genotypes
3.2. Heritability and Genetic Advance
3.3. Association of Traits among Genotypes
3.4. Genetic Diversity
3.5. Genotype × Location Interaction Analysis
3.6. Top Yielder at Locations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogunniyan, D.J.; Olakojo, S.A. Genetic variation, heritability, genetic advance and agronomic character association of yellow elite inbred lines of maize (Zea mays L.). Niger. J. Genet. 2014, 28, 24–28. [Google Scholar] [CrossRef]
- Nzuve, F.; Githiri, S.; Mukunya, D.M.; Gethi, J. Genetic variability and correlation studies of grain yield and related agronomic traits in maize. J. Agric. Sci. 2014, 6, 166–176. [Google Scholar] [CrossRef]
- Kumar, G.P.; Reddy, V.N.; Kumar, S.S.; Rao, P.V. Genetic variability, heritability and genetic advance studies in newly developed maize genotypes (Zea mays L.). Int. J. Pure Appl. Biosci. 2014, 2, 272–275. [Google Scholar]
- Ali Abdikadir, H.; Abdikadir, M.; Hasan, M.; Kalam Azad, M.A.; Hasanuzzaman, M. Genetic variability and diversity studies in maize (Zea mays L.) Inbred lines. IOSR J. Agric. Vet. Sci. 2018, 11, 69–76. [Google Scholar] [CrossRef]
- Islam, S.; Ferdausi, A.; Sweety, A.Y.; Das, A.; Ferdoush, A.; Haque, M.A. Morphological characterization and genetic diversity analyses of plant traits contributed to grain yield in maize (Zea mays L.). J. Biosci. Agric. Res. 2020, 25, 2047–2059. [Google Scholar] [CrossRef]
- Maruthi, R.T.; Rani, K.J. Genetic variability, heritability and genetic advance estimates in maize (Zea mays L.) Inbred lines. J. Appl. Natl. Sci. 2015, 7, 149–154. [Google Scholar] [CrossRef]
- Sesay, S.; Ojo, D.K.; Ariyo, O.J.; Meseka, S. Genetic variability, heritability and genetic advance studies in top-cross and three-way cross maize (Zea mays L.) hybrids. Maydica 2016, 61, 1–7. [Google Scholar]
- Kinfe, H.; Tsehaye, Y. Studies of heritability, genetic parameters, correlation and path coefficient in elite maize hybrids. Acad. Res. J. Agric. Sci. Res. 2015, 3, 296–303. [Google Scholar] [CrossRef]
- Olakojo, S.A.; Olaoye, G. Correlation and heritability estimates of maize agronomic traits for yield improvement and striga asiatica (L.) kuntze tolerance. Plant Sci. 2011, 5, 365–369. [Google Scholar]
- Roy, P.; Haque, M.; Ferdausi, A.; Bari, M. Field performance based in-depth genetic analysis of selected maize (Zea mays L.) genotypes. Fundam. Appl. Agric. 2018, 3, 1. [Google Scholar] [CrossRef]
- Syafii, M.; Cartika, I.; Ruswandi, D. Multivariate analysis of genetic diversity among some maize genotypes under maize-albizia cropping system in Indonesia. Asian J. Crop. Sci. 2015, 7, 244–255. [Google Scholar] [CrossRef]
- Ashraf, M.; Qureshi, A.G.; Khan, N.A. Genotype-environment interaction in wheat. Online J. Biol. Sci. 2001, 50, 356–367. [Google Scholar] [CrossRef]
- Tarakanovas, P.; Ruzgas, V. Additive main effect and multiplicative interaction analysis of grain yield of wheat varieties in Lithuania. Agron. Res. 2006, 491, 91–98. [Google Scholar]
- Annicchiarico, P. Additive main effects and multiplicative interaction (AMMI) analysis of genotype-location interaction in variety trials repeated over years. Theor. Appl. Genet. 1997, 4, 1072–1077. [Google Scholar] [CrossRef]
- International Board for Plant Genetic Resources (IBPGR); International Maize and Wheat Improvement Center (CIMMYT). Descriptors for Maize/Descriptores Para Maiz/Descripteurs Pour Le Maïs; International Board for Plant Genetic Resources: Wageningen, The Netherlands, 1991; p. 100. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 1 December 2017).
- Rosyara, U.R. Plant Breeding: Analysis and Visualization of Data from Plant Breeding and Genetics Experiments. R Package Version 1.1.1/r37. 2014. Available online: https://R-Forge.R-project.org/projects/plantbreeding/ (accessed on 1 December 2014).
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of genetic and environmental variability in soybeans. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Hanson, C.H.; Robinson, H.F.; Comstock, R.E. Biometrical studies of yield in segregating populations of korean lespedeza. Agron. J. 1956, 48, 268–272. [Google Scholar] [CrossRef]
- Burton, G.W.; DeVane, E.H. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J. 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Lush, J.L. Heritability of quantitative characters in farm animals. In Heritability of Quantitative Characters in Farm Animals; CABI: Wallingford, UK, 1949; pp. 356–375. [Google Scholar]
- Burton, G.W. Quantitative inheritance in grasses. In Proceedings of the 6th International Grassland Congress, State College, PA, USA, 17–23 August 1952; pp. 277–283. [Google Scholar]
- Robinson, H.F.; Cornstock, R.E.; Harvey, P.H. Estimates of heritability and degree of dominance in corn. Agron. J. 1949, 41, 353–359. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2017. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 12 January 2017).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Mahalanobis, P.C. On the generalized distance in statistics. Proc. Natl. Inst. Sci. India 1936, 2, 49–55. [Google Scholar]
- Rao, C.R. Advance Statistical Method in Biometrical Research; John Willey and Sons: New York, NY, USA, 1952. [Google Scholar]
- Maurya, A.K.; Kumar, R.; Kumar, V.; Verma, R.K.; Shweta, S.; Yadav, B.K.; Kumar, S. Studies on heritability and genetic advance estimates in maize genotypes. Plant Arch. 2014, 14, 1155–1157. [Google Scholar]
- Zaman, M.A.; Alam, M.A. Genetic diversity in exotic maize. Bangladesh J. Agric. Res. 2013, 38, 335–341. [Google Scholar] [CrossRef]
- Vishnuvardhan, K.M.; Kumar, K.P.; Murthy, K.D.; Sreevalli, M. Genetic variability, heritability and association analysis of morphological and yield components in maize hybrids. Ann. Plant Soil Res. 2021, 23, 424–431. [Google Scholar] [CrossRef]
- Gazal, A.; Nehvi, F.A.; Lone, A.A.; Dar, Z.A. Assessment of genetic variability of a set of maize inbred lines for drought tolerance under temperate conditions. Int. J. Curr. Microb. Appl. Sci. 2017, 6, 2380–2389. [Google Scholar] [CrossRef]
- Gazal, A.; Nehvi, F.; Lone, A.A. Morphological and molecular characterization of maize inbred lines showing variability for drought tolerance. Genet. Mol. Res. 2018, 17, 2. [Google Scholar] [CrossRef]
- Nagalakshmi, R.M.; Ravikesavan, R.; Paranidharan, V.; Manivannan, N.; Firoz, H.; Vignesh, M.; Senthil, N. Genetic variability, heritability and genetic advance studies in backcross populations of maize (Zea mays L.). Electron. J. Plant Breed. 2018, 9, 1137–1145. [Google Scholar] [CrossRef]
- Ganesan, K.N.; Nallathambi, G.; Senthil, N.; Tamilarasi, P.M. Genetic divergence (Zea mays L.) Analysis in indigenous maize. Electron. J. Plant Breed. 2010, 1, 1241–1243. [Google Scholar]
- Patil, S.M.; Kumar, K.; Jakhar, D.S.; Rai, A.; Borle, U.M.; Singh, P. Studies on variability, heritability, genetic advance and correlation in maize (Zea mays L.). Int. J. Agric. Environ. Biotech. 2016, 9, 1103. [Google Scholar] [CrossRef]
- Aminu, D.; Izge, A.U. Heritability and correlation estimates in maize (Zea mays L.) Under drought conditions in northern guinea and sudan savannas of Nigeria. World J. Agric. Sci. 2012, 8, 598–602. [Google Scholar] [CrossRef]
- Azam, M.G.; Sarker, U.; Maniruzzaman, M.; Banik, B.R. Genetic variability of yield and its contributing characters on CIMMYT maize inbreds under drought stress. Bangladesh J. Agric. Res. 2015, 39, 419–426. [Google Scholar] [CrossRef]
- Prakash, R.; Ravikesavan, R.; Vinodhana, N.K.; Senthil, A. Genetic variability, character association and path analysis for yield and yield component traits in maize (Zea mays L.). Electron. J. Plant Breed 2019, 10, 518–524. [Google Scholar] [CrossRef]
- Akeel, A.; Kameel Azzam, H.; Al-Ahmad, S.A. Breeding and potentials of high—Biomass rice varieties for bioenergy production in japan. Agric. Biol. J. N. Am. 2008, 4, 11–13. [Google Scholar]
- Mustafa, H.S.B.; Ahsan, M.; Aslam, M.; Ali, Q.; Hasan, E.; Bibi, T.; Mehmood, T. Genetic variability and traits association in maize (Zea mays L.). Agric. Res. 2013, 51, 231–238. [Google Scholar]
- Gokenapally, S.; Jabeen, F. Heritability, Character Association and Path Analysis for Grain Yield and Yield Contributing Characters in Maiz; Department of Genetics and Plant Breeding College of Agriculture, Central Agricultural University: Bihar, India, 2013. [Google Scholar]
- Mousavi, S.M.N.; Nagy, J. Evaluation of plant characteristics related to grain yield of FAO410 and FAO340 hybrids using regression models. Cereal Res. Commun. 2020, 49, 161–169. [Google Scholar] [CrossRef]
- Mirzamasoumzadeh, B.; Ahadzadeh, B.; Mollasadeghi, V. Effect of drought stress on yield of maize hybrids. Adv. Environ. Biol. 2014, 8, 120–122. [Google Scholar] [CrossRef]
- Chen, F.-B.; Yang, K.; Rong, T.; Pan, G. Analysis of genetic diversity of maize hybrids in the regional tests in sichuan and southwest China. Frontiers Agric. China 2008, 2, 162. [Google Scholar] [CrossRef]
- Biswas, A.; Sarker, U.; Banik, B.R.; Rohman, M.M.; Mian, M.A.K. Genetic divergence study in salinity stress tolerant maize (Zea mays L.). Bangladesh J. Agric. Res. 2015, 39, 621–630. [Google Scholar] [CrossRef]
- Rohman, M.M.; Banik, B.R.; Biswas, A.; Rahman, M.S. Genetic diversity of maize (Zea mays L.) inbreds under salinity stress. Bangladesh J. Agric. Res. 2016, 40, 529–536. [Google Scholar] [CrossRef]
- Kamara, A.Y.; Kling, J.G.; Menkir, A.; Ibikunle, O. Agronomic performance of maize (Zea mays L.) Breeding lines derived from a low nitrogen maize population. J. Agric. Sci. 2003, 141, 221–230. [Google Scholar] [CrossRef]
- Kandus, M.; Almora, D.; Ronceros, R.B.; Salenro, J.C. Statistical models for evaluating the genotype environment interaction in maize (Zea mays L.). Phyton 2010, 79, 39–46. [Google Scholar]
- Abakemal, D.; Shimelis, H.; Derera, J. Genotype-by-environment interaction and yield stability of quality protein maize hybrids developed from tropical highland adapted inbred lines. Euphytica 2016, 209, 757–769. [Google Scholar] [CrossRef]
- Amare, K.B.; Adisu, G. Adaptability and stability analysis of groundnut genotypes using AMMI model and GGE-biplot. J. Crop. Sci. Biotechnol. 2017, 20, 343–349. [Google Scholar]
- Parag-Agarwal, S.S.; Verma, S.N.; Mishra Agrarwal, P. Phenotypic stability for different quantitative traits in maize hybrids. Ind. J. Agric. Res. 2000, 34, 107–111. [Google Scholar]
- Malla, S.; Ibrahim, A.M.H.; Little, R.; Kalsbeck, S.; Glover, K.D.; Ren, C. Comparison of shifted multiplicative model, rank correlation, and biplot analysis for clustering winter wheat production environments. Euphytica 2010, 174, 357–370. [Google Scholar] [CrossRef]
- Makumbi, D.; Diallo, A.; Kanampiu, K.; Mugo, S.; Karaya, H. Agronomic performance and genotype x environment interaction of herbicide-resistant maize varieties in Eastern Africa. Crop. Sci. 2015, 55, 540–555. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M.; Obeng-Antwi, K.; Osuman, A.; Ado, S.; Coulibaly, N.; Yallou, C.; Abdulai, M.; Boakyewaa, G.; Didjeira, A. Performance of extra-early maize cultivars based on GGE biplot and AMMI analysis. J. Agric. Sci Camb. 2012, 150, 473–483. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Traits | X | LSD | CV% | σ2g | σ2p | σ2e | GCV | PCV | ECV | h2 | GA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Barishal location | |||||||||||
| AD | 88.03 | 4.40 | 2.37 | 4.10 ** | 8.44 | 4.34 | 2.30 | 3.30 | 24.69 | 0.65 | 11.37 |
| SD | 89.12 | 4.78 | 2.60 | 3.47 * | 8.85 | 5.38 | 2.09 | 3.34 | 26.22 | 0.56 | 10.26 |
| ASI | 1.20 | 1.34 | 52.07 | 0.00 ns | 0.39 | 0.39 | 0.00 | 52.07 | 160.05 | 0.00 | 0.00 |
| PH | 218.06 | 36.99 | 8.08 | 111.73 ns | 422.02 | 310.29 | 4.85 | 9.42 | 4.17 | 0.42 | 363.96 |
| EH | 119.18 | 33.00 | 13.56 | 129.77 * | 390.86 | 261.09 | 9.56 | 16.59 | 4.13 | 0.50 | 401.39 |
| KPE | 510.99 | 123.35 | 11.18 | 1288.16 ns | 4551.01 | 3262.85 | 7.02 | 13.20 | 1.26 | 0.44 | 4136.42 |
| TKW | 352.83 | 74.45 | 10.33 | 578.54 * | 1907.60 | 1329.07 | 6.82 | 12.38 | 1.91 | 0.47 | 1828.90 |
| GY | 10.75 | 2.61 | 11.87 | 0.90 * | 2.53 | 1.63 | 8.83 | 14.79 | 50.46 | 0.53 | 2.74 |
| Ishwardi location | |||||||||||
| AD | 97.40 | 1.64 | 0.82 | 11.25 ** | 11.89 | 0.64 | 3.44 | 3.54 | 6.75 | 0.97 | 23.82 |
| SD | 98.51 | 1.69 | 0.83 | 9.47 ** | 10.13 | 0.67 | 3.12 | 3.23 | 8.05 | 0.97 | 20.16 |
| ASI | 1.12 | 0.99 | 39.18 | 0.67 ** | 0.86 | 0.19 | 73.22 | 83.04 | 50.77 | 0.87 | 1.56 |
| PH | 220.95 | 16.42 | 3.34 | 103.48 ** | 157.90 | 54.42 | 4.60 | 5.69 | 4.67 | 0.79 | 257.54 |
| EH | 115.04 | 15.02 | 5.90 | 78.14 ** | 124.15 | 46.01 | 7.68 | 9.69 | 5.46 | 0.77 | 197.59 |
| KPE | 465.02 | 50.52 | 5.12 | 2258.45 ** | 2826.25 | 567.79 | 10.22 | 11.43 | 0.84 | 0.89 | 5171.93 |
| TKW | 326.60 | 23.13 | 3.47 | 1170.97 ** | 1299.27 | 128.30 | 10.48 | 11.04 | 0.87 | 0.95 | 2537.49 |
| GY | 11.65 | 1.64 | 6.79 | 1.84 ** | 2.47 | 0.63 | 11.65 | 13.48 | 32.04 | 0.85 | 4.35 |
| Jashore location | |||||||||||
| AD | 92.33 | 1.38 | 0.72 | 4.13 ** | 4.57 | 0.44 | 2.20 | 2.32 | 14.58 | 0.95 | 8.94 |
| SD | 93.55 | 1.90 | 0.92 | 4.41 ** | 5.16 | 0.75 | 2.24 | 2.43 | 16.78 | 0.92 | 9.79 |
| ASI | 1.22 | 1.48 | 53.54 | 0.18 ns | 0.61 | 0.43 | 34.37 | 63.62 | 108.06 | 0.45 | 0.56 |
| PH | 224.18 | 28.95 | 5.85 | 222.41 ** | 394.22 | 171.81 | 6.65 | 8.86 | 3.32 | 0.72 | 585.82 |
| EH | 115.23 | 12.45 | 4.57 | 215.42 ** | 243.16 | 27.74 | 12.74 | 13.53 | 2.17 | 0.94 | 470.62 |
| KPE | 497.40 | 81.36 | 8.01 | 1776.91 ** | 3363.96 | 1587.05 | 8.47 | 11.66 | 1.18 | 0.69 | 4790.46 |
| TKW | 351.33 | 55.03 | 7.62 | 1758.71 ** | 2476.29 | 717.57 | 11.94 | 14.16 | 1.08 | 0.83 | 4236.82 |
| GY | 10.51 | 2.05 | 9.16 | 0.60 * | 1.53 | 0.93 | 7.38 | 11.76 | 62.97 | 0.57 | 1.78 |
| Traits | AD | SD | ASI | PH | EH | KPE | TKW | |
|---|---|---|---|---|---|---|---|---|
| SD | rg | 0.99 ** | ||||||
| rp | 0.92 ** | |||||||
| ASI | rg | - | - | |||||
| rp | 0.06 | 0.02 | ||||||
| PH | rg | 0.80 ** | 0.74 ** | - | ||||
| rp | 0.43 ** | 0.39 * | 0.07 | |||||
| EH | rg | 0.36 * | 0.27 | - | 0.99 ** | |||
| rp | 0.32 * | 0.30 * | 0.17 | 0.70 ** | ||||
| KPE | rg | 0.17 | 0.22 | - | 0.62 ** | 0.35 * | ||
| rp | 0.23 | 0.27 | 0.21 | 0.29 | 0.18 | |||
| TKW | rg | −0.33 * | −0.19 | - | 0.23 | 24 | −0.77 ** | |
| rp | −0.19 | −0.11 | −0.04 | 0.13 | 0.15 | −0.31 * | ||
| GY | rg | 0.43 ** | 0.40 * | - | 0.99 ** | 0.73 ** | 0.61 ** | 0.48 ** |
| rp | 0.24 | 0.23 | 0.26 | 0.51 ** | 0.42 ** | 0.28 | 0.41 * |
| Traits | AD | SD | ASI | PH | EH | KPE | TKW | |
|---|---|---|---|---|---|---|---|---|
| SD | rg | 0.97 ** | ||||||
| rp | 0.96 ** | |||||||
| ASI | rg | −0.48 ** | −0.29 | |||||
| rp | −0.44 ** | −0.18 | ||||||
| PH | rg | 0.2 | 0.21 | −0.06 | ||||
| rp | 0.15 | 0.18 | 0.04 | |||||
| EH | rg | 0.43 ** | 0.53 ** | 0.2 | 0.78 ** | |||
| rp | 0.25 | 0.35 * | 0.24 | 0.75 ** | ||||
| KPE | rg | 0.32 * | 0.37 * | 0.13 | 0.23 | 0.46 ** | ||
| rp | 0.28 | 0.32 * | 0.04 | 0.26 | 0.34 * | |||
| TKW | rg | −0.16 | −0.16 | 0.04 | 0.04 | 0.2 | −0.19 | |
| rp | −0.15 | −0.16 | 0.03 | 0.08 | 0.14 | −0.17 | ||
| GY | rg | 0.09 | 0.13 | 0.02 | 0.36 * | 0.52 ** | 0.50 ** | 0.27 |
| rp | 0.08 | 0.11 | 0.05 | 0.33 * | 0.37 * | 0.62 | 0.28 |
| Traits | AD | SD | ASI | PH | EH | KPE | TKW | |
|---|---|---|---|---|---|---|---|---|
| SD | rg | 0.99 ** | ||||||
| rp | 0.95 ** | |||||||
| ASI | rg | 0.04 | 0.24 | |||||
| rp | −0.03 | 0.27 | ||||||
| PH | rg | 0.54 ** | 0.57 ** | 0.30 * | ||||
| rp | 0.42 ** | 0.41 * | 0.00 | |||||
| EH | rg | 0.61 ** | 0.59 ** | 0.15 | 0.84 ** | |||
| rp | 0.53 ** | 0.54 ** | 0.07 | 0.81 ** | ||||
| KPE | rg | 0.69 ** | 0.71 ** | 0.27 | 0.63 ** | 0.58 ** | ||
| rp | 0.56 ** | 0.55 ** | 0.05 | 0.42 ** | 0.43 ** | |||
| TKW | rg | −0.32 * | −0.29 | 0.15 | 0.26 | 0.10 | −0.81 ** | |
| rp | −0.27 | −0.25 | 0.04 | 0.12 | −0.01 | −0.54 ** | ||
| GY | rg | 0.10 | 0.03 | −0.38 * | 0.85 ** | 0.78 ** | −0.05 | 0.54 ** |
| rp | 0.08 | 0.00 | −0.26 | 0.49 ** | 0.42 ** | 0.20 | 0.44 ** |
| Traits | AD | SD | ASI | PH | EH | KPE | TKW | Multiple Regression | Stepwise Regression | |
|---|---|---|---|---|---|---|---|---|---|---|
| Barishal | b | 0.122 | 0.121 | 0.820 | 0.040 | 0.030 | 0.006 | 0.015 | ||
| r2 | 0.056 | 0.053 | 0.070 | 0.256 | 0.170 | 0.070 | 0.167 | 0.42 | 0.45 | |
| p-value | 0.116 | 0.120 | 0.082 | 0.000 | 0.004 | 0.060 | 0.005 | <0.000 | <0.000 | |
| Ishwardi | b | 0.036 | 0.050 | 0.070 | 0.040 | 0.046 | 0.017 | 0.012 | ||
| r2 | 0.006 | 0.011 | 0.001 | 0.106 | 0.133 | 0.380 | 0.070 | 0.47 | 0.51 | |
| p-value | 0.587 | 0.486 | 0.774 | 0.029 | 0.010 | <0.000 | 0.060 | <0.000 | <0.000 | |
| Jashore | b | 0.038 | 0.000 | −0.400 | 0.028 | 0.027 | 0.004 | 0.009 | ||
| r2 | 0.006 | 0.000 | 0.060 | 0.236 | 0.176 | 0.038 | 0.193 | 0.57 | 0.59 | |
| p-value | 0.608 | 0.990 | 0.080 | 0.001 | 0.004 | 0.190 | 0.002 | <0.000 | <0.000 |
| Cluster | Number of Genotypes | Percentage (%) | Accession Number |
|---|---|---|---|
| I | 12 | 26.66 | G39, G16, G18, G35, G2, G47, G36, G13, G33, G3, G8, G9 |
| II | 13 | 28.88 | G27, G29, G24, G23, G31, G21, G11,G25, G5, G6, G12, G15, G26 |
| III | 4 | 8.88 | G19, G10, G13,G44 |
| IV | 16 | 35.55 | G7,G37, G22, G33, G34, G4, G14, G30, G42, G45, G40, G1, G32, G20, G28, G41 |
| Source of Variation | Degrees of Freedom | Sum Squares | Mean Squares | % Total SS |
|---|---|---|---|---|
| Location | 2 | 53.901 | 26.951 | 11.286 |
| Genotype | 44 | 219.712 | 4.993 ** | 46.004 |
| Genotype × Location | 88 | 203.979 | 2.318 ** | 42.709 |
| Residuals | 132 | 139.081 | 1.0536 | - |
| Gen | Bar | Ish | Jas | Mean | Pi | Gen | Bar | Ish | Jas | Mean | Pi |
|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 11.12 | 11.35 | 11.43 | 11.30 | 0.28 | G24 | 10.54 | 10.17 | 10.61 | 10.44 | −0.58 |
| G2 | 10.82 | 12.53 | 11.00 | 11.45 | 0.43 | G25 | 10.20 | 11.88 | 9.57 | 10.55 | −0.48 |
| G3 | 9.78 | 12.05 | 10.10 | 10.64 | −0.38 | G26 | 11.86 | 13.19 | 9.94 | 11.66 | 0.64 |
| G4 | 10.38 | 11.54 | 8.53 | 10.15 | −0.87 | G27 | 12.18 | 13.62 * | 10.44 | 12.08 * | 1.06 |
| G5 | 11.68 | 13.58 * | 11.53 | 12.26 * | 1.24 | G28 | 10.58 | 11.92 | 12.07 * | 11.52 | 0.50 |
| G6 | 11.46 | 11.51 | 9.61 | 10.86 | −0.16 | G29 | 13.00 * | 11.84 | 12.89 * | 12.57 * | 1.55 |
| G7 | 11.51 | 13.91 * | 9.90 | 11.77 | 0.75 | G30 | 13.19 * | 10.37 | 12.15 * | 11.90 | 0.88 |
| G8 | 13.04 * | 12.51 | 11.39 | 12.31 * | 1.29 | G31 | 8.63 | 13.09 | 9.14 | 10.28 | −0.74 |
| G9 | 12.13 | 12.91 | 10.88 | 11.97 | 0.95 | G32 | 11.57 | 11.31 | 10.73 | 11.20 | 0.18 |
| G10 | 9.81 | 10.56 | 8.27 | 9.55 | −1.48 | G33 | 10.59 | 13.05 | 11.24 | 11.62 | 0.60 |
| G11 | 9.79 | 11.53 | 10.32 | 10.55 | −0.48 | G34 | 9.22 | 11.32 | 10.18 | 10.24 | −0.78 |
| G12 | 9.35 | 10.10 | 9.92 | 9.79 | −1.23 | G35 | 9.73 | 12.65 | 11.04 | 11.14 | 0.12 |
| G13 | 10.83 | 13.86 * | 10.12 | 11.60 | 0.58 | G36 | 10.48 | 11.99 | 11.40 | 11.29 | 0.27 |
| G14 | 9.80 | 11.38 | 10.88 | 10.68 | −0.34 | G37 | 9.70 | 14.51 * | 10.65 | 11.62 | 0.60 |
| G15 | 11.21 | 11.62 | 11.39 | 11.41 | 0.39 | G38 | 8.78 | 12.52 | 10.97 | 10.75 | −0.27 |
| G16 | 9.51 | 10.02 | 9.60 | 9.71 | −1.31 | G39 | 10.62 | 11.14 | 10.75 | 10.83 | −0.19 |
| G17 | 10.62 | 10.63 | 11.22 | 10.82 | −0.20 | G40 | 9.53 | 11.39 | 11.19 | 10.70 | −0.32 |
| G18 | 10.33 | 10.68 | 9.76 | 10.26 | −0.76 | G41 | 11.80 | 11.99 | 12.11 * | 11.97 | 0.95 |
| G19 | 9.75 | 5.68 | 9.97 | 8.46 | −2.56 | G42 | 13.79 * | 12.35 | 11.97 * | 12.70 * | 1.68 |
| G20 | 10.56 | 10.83 | 10.92 | 10.77 | −0.25 | G43 | 7.80 | 9.48 | 9.67 | 8.98 | −2.04 |
| G21 | 11.83 | 10.57 | 11.05 | 11.15 | 0.13 | G44 | 9.12 | 10.83 | 10.13 | 10.03 | −0.99 |
| G22 | 12.24 | 12.38 | 10.26 | 11.63 | 0.61 | G45 | 12.31 * | 10.32 | 11.79 | 11.47 | 0.45 |
| G23 | 11.17 | 11.71 | 11.10 | 11.32 | 0.30 | ||||||
| Mean | 10.75 | 11.65 | 10.66 | 11.02 | Mean | 10.75 | 11.65 | 10.66 | 11.02 | ||
| Li | −0.27 | 0.63 | −0.36 | Li | −0.27 | 0.63 | −0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.A.; Rahman, M.; Ahmed, S.; Jahan, N.; Khan, M.A.-A.; Islam, M.R.; Alsuhaibani, A.M.; Gaber, A.; Hossain, A. Genetic Variation and Genotype by Environment Interaction for Agronomic Traits in Maize (Zea mays L.) Hybrids. Plants 2022, 11, 1522. https://doi.org/10.3390/plants11111522
Alam MA, Rahman M, Ahmed S, Jahan N, Khan MA-A, Islam MR, Alsuhaibani AM, Gaber A, Hossain A. Genetic Variation and Genotype by Environment Interaction for Agronomic Traits in Maize (Zea mays L.) Hybrids. Plants. 2022; 11(11):1522. https://doi.org/10.3390/plants11111522
Chicago/Turabian StyleAlam, Mohammad Ashraful, Marufur Rahman, Salahuddin Ahmed, Nasrin Jahan, Mohammad Al-Amin Khan, Mohammad Rafiqul Islam, Amnah Mohammed Alsuhaibani, Ahmed Gaber, and Akbar Hossain. 2022. "Genetic Variation and Genotype by Environment Interaction for Agronomic Traits in Maize (Zea mays L.) Hybrids" Plants 11, no. 11: 1522. https://doi.org/10.3390/plants11111522