
Physiological and Metabolic Responses of Leymus chinensis Seedlings to Alkali Stress



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sand Cultures
2.2. Alkali Treatment
2.3. The Measurement of Photosynthetic Parameters
2.4. Sample Selection for Physiological Characteristics Determination and Growth Conditions Investigation
2.5. Measurement of Contents of Ions, Total C, Total N, and Total P
2.6. Determination of the Contents of Osmotic Metabolites, ATP, GS and GOGAT
2.7. Metabolite Extraction and Detection
2.8. Data Processing and Multivariate Data Analysis
2.9. Statistical Analysis
3. Results
3.1. Changes in Growth Parameters of L. chinensis under Alkali Stress
3.2. Changes in Photosynthetic Parameters of L. chinensis under Alkali Stress
3.3. Changes in Ions and Elements of L. chinensis under Alkali Stress
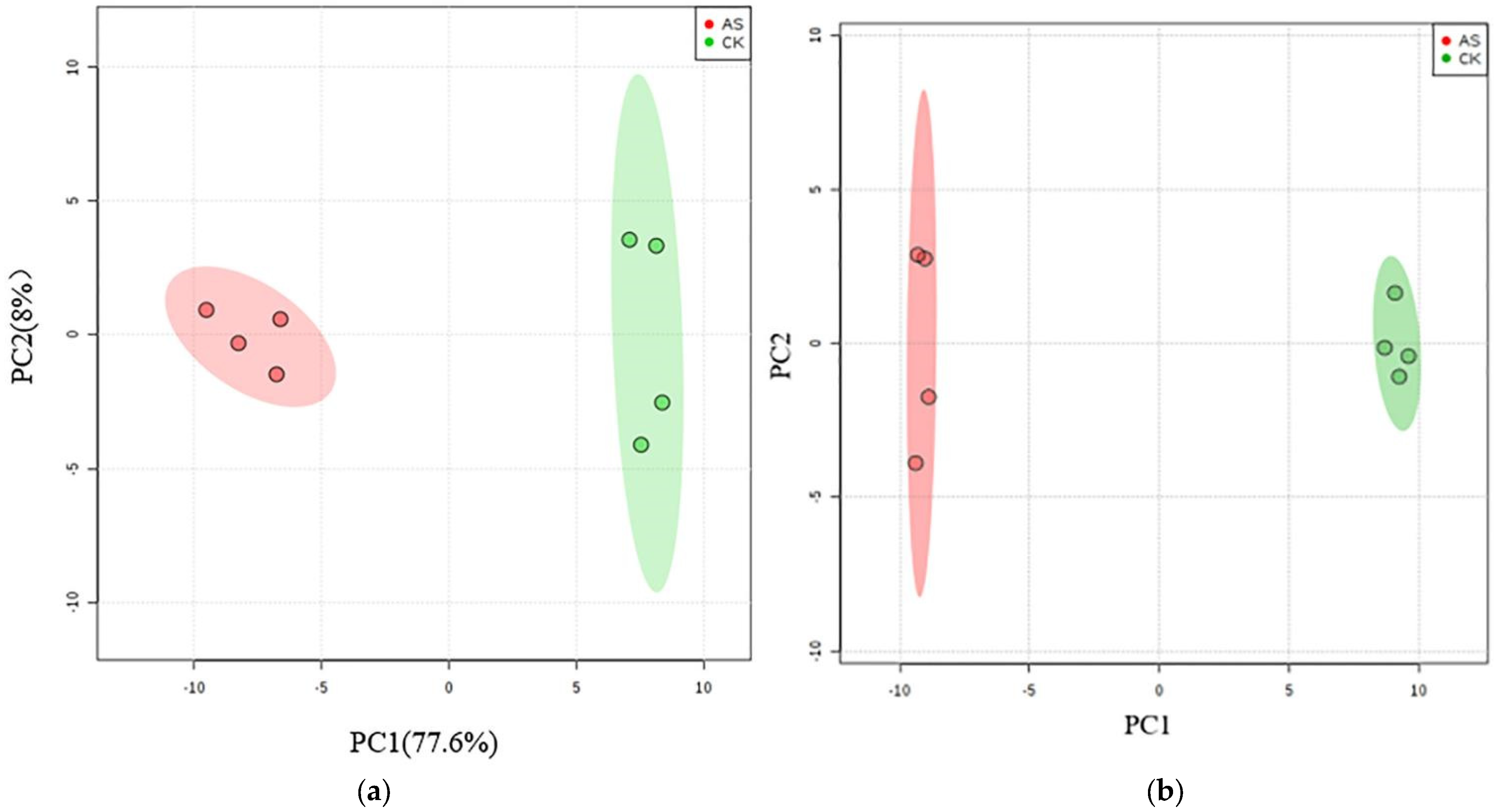
3.4. Changes in Metabolites in L. chinensis under Alkali Stress
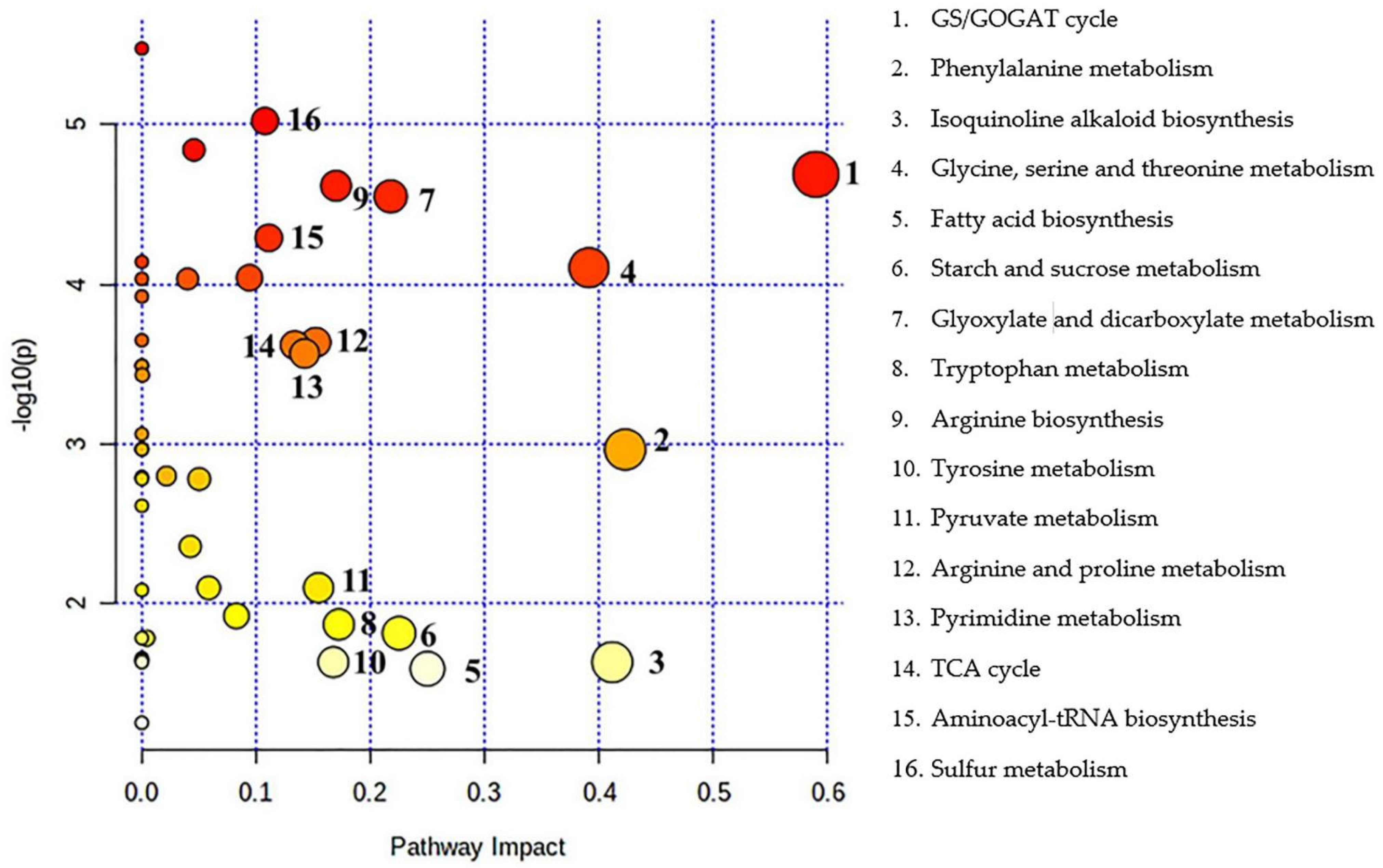
3.5. Changes to Metabolic Pathways in L. chinensis under Alkali Stress
3.6. Changes of Four Metabolites, Two Enzymes, and ATP in L. chinensis under Alkali Stress
4. Discussion
4.1. Physiological Response of L. chinensis Seedlings under Alkali Stress
4.2. Metabolites Responding to Alkali Stress
4.3. Metabolic Pathways Responding to Alkali Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sagervanshi, A.; Geilfus, C.M.; Kaiser, H.; Müehling, K.H. Alkali salt stress causes fast leaf apoplastic alkalinization together with shifts in ion and metabolite composition and transcription of key genes during the early adaptive response of Vicia faba L. Plant Sci. 2022, 319, 111253. [Google Scholar] [CrossRef] [PubMed]
- de Lacerda, C.F.; Cambraia, J.; Oliva, M.A.; Ruiz, H.A.; Prisco, J.T. Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ. Exp. Bot. 2003, 49, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Shi, L.X.; Yan, C.R.; Zhong, X.L.; Gu, F.X.; Liu, Q.; Xia, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Q.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.R.; Yang, Y.M.; Cheng, Y.X.; Zhou, T.; Duan, X.Y.; Gong, M.; Zou, Z.R. Generation of reactive oxygen species and their functions and deleterious effects in plants. Acta Bot. Boreal. -Occident. Sin. 2014, 34, 1916–1926. [Google Scholar]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Sharma, P.; Moulick, D.; Khairujjaman, M.M. Unrevealing metabolomics for abiotic stress adaptation and tolerance in plants. JCSB 2021, 24, 479–493. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Rathinasabapathi, B.; Babar, M.A. UPLC-HRMS based untargeted metabolic profiling reveals changes in Chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant Cell Environ. 2018, 42, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Chu, L.L.; Luo, C.K.; Tian, L.; Zhang, Y.X.; Yang, S.Q.; Li, P.F. Research advance in plants′adaptation to alkali stress. J. Plant Genet. Resour. 2019, 20, 836–844. [Google Scholar]
- Zou, C.L.; Wang, Y.B.; Wang, B.; Liu, D.; Liu, L.; Li, C.F. Efects of alkali stress on dry matter accumulation, root morphology, ion balance, free polyamines, and organic acids of sugar beet. Acta Physiol. Plant. 2021, 43, 13. [Google Scholar] [CrossRef]
- Yu, M.L.; Wu, Q.; Zheng, D.F.; Feng, N.J.; Liang, X.L.; Liu, M.L.; Li, Y.; Mou, B.M. Plant Growth Regulators Enhance Saline–Alkali Tolerance by Upregulating the Levels of Antioxidants and Osmolytes in Soybean Seedlings. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Jia, X.M.; Zhu, Y.F.; Hu, Y.; Zhang, R.; Cheng, L.; Zhu, Z.L.; Zhao, T.; Zhang, X.; Wang, Y.X. Integrated physiologic, proteomic, and metabolomic analyses of Malus halliana adaptation to saline–alkali stress. Hortic. Res. 2019, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.Q. Comparison of Organic Acid Accumulation in Puccinellia tenuiflora under Salt Stress and Alkali Stress. Master’s Thesis, Northeast Normal University, Changchun, China, 2005. [Google Scholar]
- Yang, C.W. Mechanisms of Alkali Tolerance in Chloris Virgata and Rice (Oryza sativa). Ph.D. Dissertation, Northeast Normal University, Changchun, China, 2010. [Google Scholar]
- Ma, Y. Physiological Mechanism of Salt-Alkaline Tolerance in Kochia sieversiana. Ph.D. Dissertation, Northeast Normal University, Changchun, China, 2011. [Google Scholar]
- Zhang, J.; Yang, D.S.; Li, M.X.; Shi, L.X. Metabolic profiles reveal changes in wild and cultivated soybean seedling leaves under salt stress. PLoS ONE 2016, 11, e0159622. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Zhou, J.; Yang, F.; Li, F. Effects of alkaline stress on metabonomic responses of wheat (Triticum aestivum Linn) leaves. Sci. Agric. Sin. 2017, 50, 250–259. [Google Scholar]
- Wang, H.; Xiang, Y.; Li, L.H.; Bhanbhro, N.; Yang, C.W.; Zhang, Z. Photosynthetic response and transcriptomic profiling provide insights into the alkali tolerance of clone halophyte Leymus chinensis. Photosynthetica 2020, 58, 780–789. [Google Scholar] [CrossRef]
- Lin, J.X.; Peng, X.Y.; Hua, X.Y.; Sun, S.N.; Wang, Y.N.; Yan, X.F. Effects of arbuscular mycorrhizal fungi on Leymus chinensis seedlings under salt-alkali stress and nitrogen deposition conditions: From osmotic adjustment and ion balance. RSC Adv. 2018, 8, 14500–14509. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Y.; Mu, C.S. Effects of salt and alkali stresses, and exogenous plant hormones on growth and development of wheat and L. chinensis. Acta Agrestia Sin. 2017, 25, 257–260. [Google Scholar]
- Wang, Y.N.; Tao, S.; Hua, X.Y.; Yu, X.Y.; Yan, X.F.; Lin, J.X. Effects of arbuscular mycorrhizal fungi on the growth and physiological metabolism of Leymus chinensis under salt-alkali stress. Acta Ecol. Sin. 2018, 38, 2187–2194. [Google Scholar]
- Liu, B.S.; Kang, C.L.; Wang, X.; Bao, G.Z. Tolerance mechanisms of Leymus chinensis to salt–alkaline stress. Acta Agric. Scand. B-Soil Plant Sci. 2015, 65, 723–734. [Google Scholar]
- Guo, R. The Study of Saline and Alkaline Tolerant Eco-Physiological Metabolism in Four Gramineae in the Songnen Grasslands. Ph.D. Dissertation, Northeast Normal University, Changchun, China, 2010. [Google Scholar]
- Liu, X.H.; Hou, G.L.; Zou, Y.C.; Wu, H.T.; Lv, X.G.; Jiang, M. Soil Carbon Storage and Climate Regulation by Natural Reserves in the Songnen Plain. J. Northeast. Agric. Univ. 2021, 49, 122–126. [Google Scholar]
- Guo, R.; Shi, L.X.; Yang, Y.F. Germination and growth, osmotic adjustment and ionic balance of wheat in response to saline and alkaline stresses. Soil Sci. Plant Nutr. 2009, 55, 667–679. [Google Scholar] [CrossRef]
- Cao, M. Effects of Saline-Alkali Stress on the Individual and Clonal Growth Traits of Leymus chinensis. Master’s Thesis, Northeast Normal University, Changchun, China, 2017. [Google Scholar]
- Xiang, G.; Yang, H.Y.; Yang, L.; Zhang, X.; Cao, Q.E.; Miao, M.M. Multivariate statistical analysis of tobacco of different origin, grade and variety according to polyphenols and organic acids. Microchem. J. 2010, 95, 198–206. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, G.S.; Hou, Y.Y.; Gao, Y.G.; Wang, S.J.; Cai, E.B.; Zhang, L.X. Determination of amino acids in seeds of Setaria viridis (L.) Beauv. by pre-column derivatization RP-HPLC. J. Northwest. Sci.-Tech. Univ. Agric. For. (Nat. Sci. Ed.) 2015, 43, 185–190. [Google Scholar]
- Chen, Q.; Zhang, Y.M.; Tao, M.M.; Li, M.S.; Wu, Y.; Qi, Q.; Yang, H.; Wan, X.C. Comparative metabolic responses and adaptive strategies of tea leaves (Camellia sinensis) to N2 and CO2 anaerobic treatment by a nontargeted metabolomics approach. J. Agric. Food Chem. 2018, 66, 9565–9572. [Google Scholar] [CrossRef]
- Guo, L.Q. The Study of Alkali-Tolerant Metabolism in Puccinellia tenuiflora. Ph.D. Dissertation, Northeast Normal University, Changchun, China, 2009. [Google Scholar]
- El Yamani, M.; Sakar, E.; Boussakouran, A.; Rharrabti, Y. Leaf water status, physiological behavior and biochemical mechanism involved in young olive plants under water deficit. Sci. Hortic. 2020, 261, 108906. [Google Scholar] [CrossRef]
- Salama, K.H.A.; Mansour, M.M.F. Choline priming-induced plasma membrane lipid alterations contributed to improved wheat salt tolerance. Acta Physiol. Plant. 2015, 37, 1–7. [Google Scholar] [CrossRef]
- Panda, A.; Rangani, J.; Parida, A.K. Unraveling salt responsive metabolites and metabolic pathways using non-targeted metabolomics approach and elucidation of salt tolerance mechanisms in the xero-halophyte Haloxylon salicornicum. Plant. Physiol. Bioch. 2021, 158, 284–296. [Google Scholar] [CrossRef]
- Yang, C.X.; Zhao, W.N.; Wang, Y.N.; Zhang, L.; Huang, S.C.; Lin, J.X. Metabolomics analysis reveals the alkali tolerance mechanism in Puccinellia tenuiflora plants inoculated with Arbuscular Mycorrhizal Fungi. Microorganisms 2020, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Manova, V.; Gruszka, D. DNA damage and repair in plants–from models to crops. Front. Plant Sci. 2015, 6, 885. [Google Scholar] [CrossRef] [Green Version]
- Stasolla, C.; Katahira, R.; Thorpe, T.A.; Ashihara, H. Purine and pyrimidine nucleotide metabolism in higher plants. J. Plant Physiol. 2003, 160, 1271–1295. [Google Scholar] [CrossRef]
- Zang, W.; Miao, R.Q.; Zhang, Y.; Yuan, Y.; Pang, Q.Y.; Zhou, Z.Q. Metabolic and molecular basis for the salt and alkali responses of Suaeda corniculata. Environ. Exp. Bot. 2021, 192, 104643. [Google Scholar] [CrossRef]
- Hossain, M.; Uddin, M.; Ismail, M.R.; Ashrafuzzaman, M. Responses of glutamine synthetase-glutamate synthase cycle enzymes in tomato leaves under salinity stress. Int. J. Agricul. Biol. 2012, 14, 509–515. [Google Scholar]
- Zhao, M.L.; Guo, R.; Li, M.X.; Liu, Y.; Wang, X.X.; Fu, H.; Wang, S.Y.; Liu, X.Y.; Shi, L.X. Physiological characteristics and metabolomics reveal the tolerance mechanism to low nitrogen in Glycine soja leaves. Physiol. Plantarum. 2020, 168, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Sun, M.; Lin, S.J.; Guo, Y.H.; Yang, Y.J.; Zhang, T.X.; Zhang, J.X. Transcriptome analysis of Crossostephium chinensis provides insight into the molecular basis of salinity stress responses. PLoS ONE 2017, 12, e0187124. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | log2(AS/CK) | p-Value |
|---|---|---|
| Germination rate (%) | −0.22 | <0.05 |
| Shoot height (cm) | −0.13 | <0.01 |
| Root length (cm) | −0.09 | <0.05 |
| Dry weight of shoots (g) | −0.17 | <0.01 |
| Dry weight of roots (g) | −0.12 | <0.01 |
| Leaf number | 0.25 | <0.05 |
| Photosynthesis Parameters (μmol CO2 ·m−2·s−1) | log2(AS/CK) | p-Value |
|---|---|---|
| Pn | −0.29 | <0.05 |
| Gs | −0.18 | <0.01 |
| Ci | −0.11 | <0.05 |
| E | −0.19 | <0.01 |
| SPAD value | −1.18 | <0.05 |
| Ion and Element Contents | log2(AS/CK) | p-Value | |
|---|---|---|---|
| Ion content (μmol·g−1 DW) | Na+ | 0.35 | <0.01 |
| K+ | −0.27 | <0.05 | |
| Ca2+ | −0.15 | <0.05 | |
| Mg2+ | −0.10 | NS | |
| Cl− | 0.04 | NS | |
| NO3− | −0.05 | <0.05 | |
| H2PO4− | −0.07 | <0.01 | |
| SO42− | 0.12 | NS | |
| Element content (mg·g−1) | C | −0.24 | <0.05 |
| N | 0.15 | <0.05 | |
| P | −0.02 | NS |
| Total Compound | Hits | Raw p | Log (p) | Holm Adjust | FDR | Impact | |
|---|---|---|---|---|---|---|---|
| GS/GOGAT cycle | 22 | 5 | 0.000020 | 4.6893 | 0.000859 | 0.000211 | 0.58992 |
| Phenylalanine metabolism | 12 | 1 | 0.001087 | 2.9638 | 0.025850 | 0.002038 | 0.42308 |
| Isoquinoline alkaloid biosynthesis | 6 | 1 | 0.023442 | 1.6300 | 0.155050 | 0.024532 | 0.41176 |
| Glycine, serine, and threonine metabolism | 33 | 7 | 0.000079 | 4.1039 | 0.002913 | 0.000347 | 0.39130 |
| Fatty acid biosynthesis | 14 | 1 | 0.025780 | 1.5887 | 0.155050 | 0.026366 | 0.25000 |
| Starch and sucrose metabolism | 22 | 2 | 0.015432 | 1.8116 | 0.154320 | 0.019289 | 0.22509 |
| Glyoxylate and dicarboxylate metabolism | 29 | 7 | 0.000028 | 4.5510 | 0.001125 | 0.000211 | 0.21779 |
| Tryptophan metabolism | 23 | 1 | 0.013612 | 1.8661 | 0.149740 | 0.017502 | 0.17241 |
| Arginine biosynthesis | 18 | 4 | 0.000024 | 4.6172 | 0.000990 | 0.000211 | 0.16991 |
| Tyrosine metabolism | 18 | 1 | 0.023442 | 1.6300 | 0.155050 | 0.024532 | 0.16757 |
| Pyruvate metabolism | 22 | 1 | 0.008012 | 2.0963 | 0.120170 | 0.011266 | 0.15462 |
| Arginine and proline metabolism | 28 | 2 | 0.000231 | 3.6360 | 0.007190 | 0.000677 | 0.15223 |
| Pyrimidine metabolism | 38 | 8 | 0.000272 | 3.5653 | 0.007891 | 0.000720 | 0.14246 |
| Citrate cycle (TCA cycle) | 20 | 3 | 0.000241 | 3.6186 | 0.007219 | 0.000677 | 0.13384 |
| Aminoacyl-tRNA biosynthesis | 46 | 9 | 0.000051 | 4.2909 | 0.001996 | 0.000329 | 0.11111 |
| Sulfur metabolism | 15 | 3 | 0.000009 | 5.0239 | 0.000416 | 0.000211 | 0.10774 |
| Metabolites | log2(AS/CK) | p-Value |
|---|---|---|
| Amino acid | 0.13 | <0.01 |
| Organic acid | 0.04 | <0.01 |
| Sugar | −0.13 | <0.01 |
| Nucleotide | 0.19 | <0.05 |
| ATP | 0.06 | <0.01 |
| GS | 0.10 | <0.01 |
| GOGAT | 0.11 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, G.; Shi, Y.; Chen, F.; Mu, C.; Wang, J. Physiological and Metabolic Responses of Leymus chinensis Seedlings to Alkali Stress. Plants 2022, 11, 1494. https://doi.org/10.3390/plants11111494
Yan G, Shi Y, Chen F, Mu C, Wang J. Physiological and Metabolic Responses of Leymus chinensis Seedlings to Alkali Stress. Plants. 2022; 11(11):1494. https://doi.org/10.3390/plants11111494
Chicago/Turabian StyleYan, Ge, Yujie Shi, Fangfang Chen, Chunsheng Mu, and Junfeng Wang. 2022. "Physiological and Metabolic Responses of Leymus chinensis Seedlings to Alkali Stress" Plants 11, no. 11: 1494. https://doi.org/10.3390/plants11111494