
The Road to Practical Application of Cadmium Phytoremediation Using Rice

Akita Prefectural Agricultural Experiment Station, 34-1, Genpachizawa, Yuwa-Aikawa, Akita 010-1231, Japan
*
Author to whom correspondence should be addressed.
Plants 2021, 10(9), 1926; https://doi.org/10.3390/plants10091926
Submission received: 16 August 2021
/
Revised: 8 September 2021
/
Accepted: 9 September 2021
/
Published: 15 September 2021
(This article belongs to the Special Issue New Insight into Cadmium Phytoremediation)
Abstract
:Cadmium (Cd) is a toxic heavy metal that causes severe health issues in humans. Cd accumulates in the human body when foods produced in Cd-contaminated fields are eaten. Therefore, soil remediation of contaminated fields is necessary to provide safe foods. Rice is one of the primary candidates for phytoremediation. There is a genotypic variation of Cd concentration in the shoots and grains of rice. Using the world rice core collection, ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’ were observed with a significantly higher level of Cd accumulation in the shoots and grains. Moreover, OsHMA3, a heavy metal transporter, was identified as a responsive gene of quantitative trait locus (QTL) for high Cd concentration in the shoots of these three varieties likewise. However, it is difficult to apply practical phytoremediation to these varieties because of their unfavorable agricultural traits, such as shatter and easily lodged. New rice varieties and lines were bred for Cd phytoremediation using OsHMA3 as a DNA marker selection. All of them accumulated Cd in the shoots equal to or higher than ‘Cho-ko-koku’ with improved cultivation traits. Therefore, they can be used for practical Cd phytoremediation.
1. Introduction
Cadmium (Cd) is a toxic heavy metal that causes severe health issues in humans [1]. Cd may accumulate in the human body when it enters the food chain. The Codex Alimentarius Commission established a Cd limit in some agricultural products to provide safe food for human intake, e.g., 0.4 mg kg−1 for polished rice [2]. However, agricultural products harvested from Cd-contaminated fields can lead to Cd levels that exceed the Cd limit [3,4,5]. This high level of Cd can become a significant problem. Cd pollution of agricultural fields is mainly caused by the influx of Cd filled wastewater from mines and factories into agricultural fields. Cd pollution has an especially large impact on paddy fields. In addition, rice is the primary source of dietary Cd intake among Asians [3,4,5]. Therefore, soil remediation of contaminated fields, especially paddy fields, is necessary to provide safe foods for human health.
Phytoremediation is a soil remediation method that removes pollutants using plants. Phytoremediation is superior to other methods regarding its low cost and its ability to effectively purify large areas of field without any facilities. Generally, plants with large biomass and plants that accumulate high Cd levels in the aerial parts (shoots and grain) are required for effective phytoextraction. Cd hyperaccumulators, such as Solanum nigrum, Pterocypsela laciniata, Sedum plumbizincicola, are some of the candidates for soil remediation because of the high levels of Cd accumulation in the aerial parts [6,7,8]. However, most Cd hyperaccumulator have a small biomass which makes harvesting work difficult. Furthermore, a practical problem is the concern that they will spread as weed in the agricultural field after phytoremediation. On the other hand, some particular plant species, such as Vigna unguiculata, Solanum melonaena, Momordica charantia, Nicotiana tabacum, have also been reported for use with Cd phytoremediation [9]. In addition to these plants, rice (Oryza sativa L.) is also one of the primary candidates for soil remediation [10]. There is a genotypic variation of concentration of Cd ions in the shoots and rice grains, and some rice varieties showed a significantly high Cd accumulation in the shoots [11,12,13]. In addition, rice has large biomass. Furthermore, in the case of rice, there is a fully established cultivation and harvesting method, which means farmers can easily continue to work on soil remediation as they always have when cultivating and harvesting rice. Therefore, it is possible to perform phytoremediation by selecting high Cd-accumulating varieties to be cultivated without any special method or equipment.
In this review, we focus on Cd phytoremediation of paddy fields using rice. We discuss recent progress in breeding new rice varieties and future prospects for practical application.
2. Phytoremediation Using Local Rice Varieties
Among rice species, Oryza sativa subsp. indica and hybrid varieties of O. sativa subsp. japonica and indica (japonica-indica) exhibit a relatively high Cd accumulation in the shoots compared to the O. sativa subsp. japonica varieties [14]. Therefore, the candidate varieties for Cd phytoremediation were initially selected among indica or japonica-indica hybrid varieties. ‘Milyang 23’ is a japonica-indica variety that exhibits a relatively high Cd accumulation in the shoots [11]. When ‘Milyang 23’ was grown in Cd contaminated soil, it accumulated 10–15% of soil Cd in its shoots, and the decrease in Cd concentration in the soil after cultivation was the largest among the major crops, including the japonica rice variety ‘Nipponbare’ [15]. The Cd contents of soybean seeds cultivated in the same field after ‘Milyang 23’ was grown were less than those cultivated using untreated soil [16]. In a field experiment, the indica variety ‘Moretsu’ exhibited approximately 2.5-fold higher Cd accumulation than ‘Milyang 23’, and ‘IR-8’ (indica variety) also showed a high level of Cd accumulation in the shoots [17]. After performing phytoremediation using these two varieties for two years, the Cd concentration in the soil decreased by 18% compared with the soil before cultivation. Furthermore, the Cd contents in the grains of subsequently grown japonica rice were lower than those grown in a field without phytoremediation [17]. These results indicate that remediation using high Cd-accumulating rice is effective not only in the paddy field but also in the converted upland field.
The world rice core collection (WRC) covered the genetic diversity of 32,000 genotypes of cultivated rice [18]. Within WRC, ‘Jarjan’ ‘Anjana Dhan’, and ‘Cho-ko-koku’ accumulated a significantly high level of Cd in the shoots and grains [13]. Cd uptake by ‘Cho-ko-koku’ was higher than ‘IR-8’ even in the field, and the Cd concentration in the field soil and grains of subsequent cultivars decreased more than those in the control field after the cultivation of ‘Cho-ko-koku’ for 2 or 4 years [19,20]. Phytoremediation capacity of Cd removal from soil depends on Cd concentration in the aerial parts and biomass of the plants. ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’ have a large biomass, and these varieties were considered advantageous for phytoremediation. However, ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’ presented some difficulties in regard to practical phytoremediation because of their unfavorable agricultural traits, such as shatter and easily lodged. Therefore, it was necessary to breed new rice varieties, which accumulate high levels of Cd like ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’.
3. OsHMA3 Is an Important Metal Transporter for Cd Accumulation in the Shoots
Many studies have been done to identify and functionally analyze the genes related to Cd uptake and translocation in rice [21,22,23]. A primary quantitative trait locus (QTL) for increasing Cd concentration in the shoots and rice grains was located on chromosome 7 [24,25,26]. Backcrossed inbred lines (BILs) containing the QTL region (qCdp7) of ‘Jarjan’ allele reduced Cd concentration in the soil and subsequently cultivated rice shoots [27].
Heavy metal ATPase (HMA) is a metal transporter family, and some HMA in rice transports Cd [21,22,28]. OsHMA3 is localized to the vacuole membrane and functions as sequestration of Cd into vacuoles in the root cells [29,30]. One amino acid substitution causes a loss of the function of OsHMA3, and a disruption of the Cd sequestration into vacuoles leads to an increased Cd concentration in the cytoplasm (Figure 1). As a result, more Cd loads into the xylem and translocates from the roots to the shoots [31,32]. OsHMA3 from ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’ were observed with the amino acid substitution. Furthermore, the region detected qCdp7 included OsHMA3, and OsHMA3 was identified as a responsive gene of QTL for high Cd concentration in the shoots of ‘Jarjan’, ‘Anjana Dhan’, and ‘Cho-ko-koku’ [29,30,33]. Recently, it has been reported that various mutations of OsHMA3 contribute to various levels of Cd accumulation in the shoots and rice grains [34,35,36]. These results suggest that OsHMA3 plays a vital role in Cd accumulation in the shoots and rice grains. In addition, the discovery of significant genes involved in Cd translocation from the roots to the shoots and Cd accumulation in the shoots enabled the efficient breeding of rice varieties for Cd phytoremediation.
4. Breeding New Rice Varieties for Phytoremediation
Rice line ‘MJ3’ and ‘MA22’ were obtained by gamma-ray mutation of ‘Jarjan’ and ‘Anjana Dhan’, respectively [37]. ‘MJ3’ and ‘MA22’ showed the same level of Cd extraction as ‘Jarjan’ and ‘Anjana Dhan’, respectively, with a non-shattering habit. However, the culm length of ‘MJ3’ was shorter than ‘Jarjan,’ and the lodging resistance of ‘MJ3’ was improved when compared with ‘Jarjan.’ On the other hand, ‘MA22’ showed almost the same culm length and was as easily lodged as ‘Anjana Dhan’.
‘TJTT8’ was developed from BILs derived from ‘Jarjan’ and ‘Tachisugata’ [38]. ‘Tachisugata’ is a japonica-indica hybrid variety used as a livestock feed. It shows a large biomass and lodging resistance because of its thick and rigid culms [39]. The grains of ‘TJTT8’ are dark brown color and can easily be distinguished from the grains of general japonica varieties like ‘MJ3’ (Table 1). ‘TJTT8’ was selected using the qCdp7 allele and showed the same level of Cd extraction as ‘Jarjan’ at several locations of Cd-contaminated paddy fields in Japan [38]. However, the heading date and maturing date of ‘MJ3’, ‘MA22’, and ‘TJTT8’ were too late in northern parts of Japan. Late heading date and maturing date characteristics may lead to difficulty in the harvest work due to insufficient drying of the aerial parts and the need for obtaining seeds for subsequent planting. Therefore, other varieties suitable for the cultivation conditions in northern parts of Japan were required.
‘Akita 110’ is a rice line developed by the Akita Prefectural Agricultural Experiment Station located in the northern part of Japan. ‘Akita 110’ was selected from a cross between ‘Cho-ko-koku’ and ‘Akita 63’ [40]. ‘Akita 63’ showed lodging resistance and large biomass of the aerial parts [41]. In the process of breeding ‘Akita 110’, OsHMA3 was used as a DNA marker to select plants that possessed this ‘Cho-ko-koku’ allele. As a result, the Cd extraction of ‘Akita 110’ was almost the same level as ‘Cho-ko-koku’. In one year of a large-scale field trial, soil Cd concentrations in plots remediated with ‘Akita 110’ were reduced by 15.5%, whereas remediation with ‘Cho-ko-koku’ reduced soil Cd levels by 10.1% [40]. However, the Cd extraction of ‘Akita 110’ was sometimes lower than that of ‘Cho-ko-koku’, depending on the field conditions. Then, a new rice line, ‘Akita 119’, was developed to improve stable Cd accumulation in the aerial parts. ‘Akita 119’ was obtained by a soft X-ray mutation of ‘Cho-ko-koku’ [42]. ‘Akita 119’ was also selected by the OsHMA3 allele of ‘Cho-ko-koku’. The culm length of ‘Akita 119’ was around 30 cm shorter than ‘Cho-ko-koku’, and the lodging resistance of ‘Akita 119’ was improved compared to ‘Cho-ko-koku’. ‘Akita 119’ had many panicles, and its biomass was as great as that of ‘Cho-ko-koku’, even though the culm was short. The grains of ‘Akita 119’ are slender with a light brown color (Table 2). The grains can also be distinguished from the grains of the general japonica varieties and do not mix in the distribution process if separated by a sieve.

{kind=link}
{kind=link}
Table 1.
Characterization of ‘MJ3’, ‘TJTT8’, ‘Akita 110’, and ‘Akita 119’. “2*Tachisugata” means crossing twice with ‘Tachisugata.’
Table 1.
Characterization of ‘MJ3’, ‘TJTT8’, ‘Akita 110’, and ‘Akita 119’. “2*Tachisugata” means crossing twice with ‘Tachisugata.’
| MJ3 | TJTT8 | Akita 110 | Akita 119 | |
|---|---|---|---|---|
| Crossing combination (Mother/Father) | Gamma-ray mutation of ‘Jarjan’ | Tachisugata/Jarjan //2*Tachisugata | Cho-ko-koku/Akita 63 | Soft X-ray mutation of ‘Cho-ko-koku’ |
| Lodging resistance | Weak to medium | Very strong | Medium | Weak to medium |
| Shattering resistance | Strong | Strong | Strong | Strong |
| Distinguish points with general japonica varieties | Long culm length Grain shape Grain color | Long culm length Grain shape Grain color | Grain shape Grain color | Many panicle number Grain shape Grain color |
| Reference | [37] | [38] | [40] | [42] |
When ‘Akita 119’ was cultivated in a Cd-contaminated paddy field in Akita Prefecture, the Cd extraction of ‘Akita 119’ was almost the same level as ‘Cho-ko-koku’ in all experiment fields from 2011 to 2016, whereas ‘Akita 110’ was lower than ‘Cho-ko-koku’ in some fields (Table 2). We also cultivated ‘MJ3’, ‘MA22’, and ‘TJTT8’ in the same Cd-contaminated paddy field in Akita Prefecture from 2014 to 2016 (Table 2). ‘MA22’ showed the highest Cd extraction among trial plants in 2014 but lodged more severely than ‘Cho-ko-koku.’ On the other hand, Cd extraction of ‘MJ3’ and ‘TJTT8’ was about 1.5 times higher than that of ‘Cho-ko-koku’ on average, although some cultivating problems of these lines, such as late heading and maturing date, were revealed in northern parts of Japan.
5. Another Approach to Breeding New Rice Varieties
Morphological improvements are another approach to breeding new rice varieties for phytoremediation. They promise a further increase of biomass in the aerial parts by crossing varieties with large biomass, such as rice used for livestock feed. On the other hand, expanding the rhizosphere might also be an effective strategy. It is considered that deeper rooting and maximal root length enhance nutrition and water uptake from soil [43,44]. Similarly, root modification may enhance Cd absorption of phytoremediation rice.
In general, as the dry weight in the shoots increases, Cd concentration in the shoots decreases. Therefore, it is necessary to improve Cd translocation efficiency from roots to shoots to accumulate a higher Cd level in the aerial parts. OsNRAMP5 is a manganese, iron, and Cd transporter and is recognized as a major route of Cd influx into root cells [45,46]. Mutant rice with defective OsNRAMP5 showed little absorption of Cd [47]. On the other hand, rice, whose gene expression of OsNRAMP5 was suppressed by RNAi, accumulated higher amounts of Cd in their shoots than the wild type, whereas the total Cd content (roots plus shoots) was reduced [45]. This is considered to be because the expression of genes involved in metal transport was enhanced. This indicates that combining some genes related to Cd absorption and translocation is effective in enhancing Cd accumulation in the aerial parts.
A major QTL and responsive gene for increasing Cd concentration in the shoots and the grains of rice was identified as OsHMA3 within chromosome 7 [24,25]. In addition, other QTLs for Cd accumulation in the aerial parts and Cd translocation from roots to shoots have been detected in other studies [48,49,50] (Table 3). Thus, the mechanisms of Cd accumulation may be different among each high Cd-accumulating variety. Using these varieties in combination is effective for breeding new rice varieties for Cd phytoremediation (Figure 2).
6. Another Approach for Efficient Cd Phytoremediation
Cd absorption from soil to rice roots is maximized under drained and oxidative soil conditions [51]. This is because in such situations, Cd in the soil exists in a chemical form that is easily absorbed by the rice. Therefore, making more Cd in the soil into an available form affects the efficiency of phytoremediation. Some microorganisms, including bacteria and arbuscular mycorrhizal fungi, can convert unavailable Cd in the soil, facilitating its bioavailability for plants. In addition, they are well known to be able to increase the tolerance of plants to Cd stress. Numerous microbes have been reported to enhance the efficiency of Cd phytoremediation by some plant species [52,53,54]. In many cases, microorganisms interact with a limited host plant, so it is necessary to isolate microbes that operate under rice cultivation conditions.
7. Future Prospects and Conclusions
In addition to OsHMA3 and OsNRAMP5, many genes related to Cd uptake and translocation are well known in rice. For example, OsIRT1, OsIRT2, and OsNRAMP1 are involved in Cd uptake and Cd translocation within plants [55,56,57], and OsHMA2 plays a role in Cd translocation from roots to shoots and/or Cd distribution in the nodes [58,59,60]. Furthermore, OsZIP1 functions as a metal exporter under Cd excess condition to prevent Cd stress [61]. Therefore, transgenic rice development using these gene is one of the most efficient methods for breeding a new rice variety for Cd phytoremediation. OsMTP1, which is localized to the vacuole membrane, functions as sequestration of Cd into vacuoles, and overexpressing OsMTP1 enhanced Cd accumulation in transgenic tobacco [62,63]. Higher accumulation in the shoots was also observed in the OsNRAMP5-RNAi rice of ‘Anjana Dhan’, even in a Cd-contaminated field [64]. However, transgenic rice has not been practicalized in any country, even in the inedible varieties. If genetically modified crops are commercialized, it is expected that the development of new rice varieties for Cd phytoremediation will also progress dramatically.
Although a limit on Cd content in crops has been established, it is not clear how much the Cd concentration in the soil should be reduced by phytoremediation for the safe production of subsequent crops. Clarification of the change of soil Cd concentrations before and after phytoremediation is complex because it strongly depends on weather and soil conditions. Furthermore, Cd-contaminated fields, which can be used to perform field trials for phytoremediation, are limited. However, more field trial results are necessary to establish a guideline for the end of phytoremediation.
Phytoremediation is an effective method to restore Cd-contaminated soil to an agricultural field. For further amplification of Cd phytoremediation, it is necessary to establish an efficient method that also reduces the burden on farmers. It must include field work and pecuniary, because farmers usually have no income during the phytoremediation process. Therefore, it is necessary that researchers and government work together toward the practical application of Cd phytoremediation.
Author Contributions
The experiments in Cd-contaminated paddy fields in Akita Prefecture, including measurement of Cd concentration, were performed by R.T., M.I., T.K. and R.T. wrote this paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by a grant from the Ministry of Agriculture, Forestry, and Fisheries of Japan (Genome for Agricultural Innovation [QTL-4012] and Regulatory Research Projects for Food Safety, Animal Health and Plant Protection [no. 2603]).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No supporting data in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, L.; Cui, X.; Cheng, H.; Chen, F.; Wang, J.; Zhao, X.; Lin, C.; Pu, X. A review of soil cadmium contamination in China including a health risk assessment. Environ. Sci. Pollut. Res. 2015, 22, 16441–16452. [Google Scholar] [CrossRef] [PubMed]
- CODEX Alimentarius. General Standard for Contaminants and Toxins in Food and Feed. CXS 193-1995. Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%3A%2F%2Fworkspace.fao.org%2Fsites%2Fcodex%2FStandards%2FCXS+193-1995%2FCXS_193e.pdf (accessed on 10 July 2021).
- Cheng, F.; Zhao, N.; Xu, H.; Li, Y.; Zhang, W.; Zhu, Z.; Chen, M. Cadmium and lead contamination in japonica rice grains and its variation among the different locations in southeast China. Sci. Total Environ. 2006, 359, 156–166. [Google Scholar]
- Moon, C.S.; Paik, J.M.; Choi, C.S.; Kim, D.H.; Ikeda, M. Lead and cadmium levels in daily foods, blood and urine in children and their mothers in Korea. Int. Arch. Occup. Environ. Health 2003, 76, 282–288. [Google Scholar] [CrossRef]
- Watanabe, T.; Shimbo, S.; Nakatsuka, H.; Koizumi, A.; Higashikawa, K.; Matsuda-Inoguchi, N.; Ikeda, M. Gender-related difference, geographical variation and time trend in dietary cadmium intake in Japan. Sci. Total Environ. 2004, 329, 17–27. [Google Scholar] [CrossRef]
- Li, K.; Yang, B.; Wang, X.; Xu, X.; Gao, Y.; Zhu, Y. Dual effects of biochar and hyperaccumulator Solanum nigrum L. on the remediation of Cd-contaminated soil. Peer J. 2019, 7, e6631. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Lin, L.; Liao, M.; Wang, J.; Tang, Y.; Sun, G.; Liang, D.; Xia, H.; Wang, X.; Zhang, H.; et al. Phytoremediation potential of Pterocypsela laciniata as a cadmium hyperaccumulator. Environ. Sci. Pollut. Res. Int. 2019, 26, 13311–13319. [Google Scholar] [CrossRef]
- Huang, R.; Dong, M.; Mao, P.; Zhuang, P.; Paz-Ferreiro, J.; Li, Y.; Li, Y.; Hu, X.; Netherway, P.; Li, Z. Evaluation of phytoremediation potential of five Cd (hyper)accumulators in two Cd contaminated soils. Sci. Total Environ. 2020, 721, 137581. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of cadmium: Physiological, biochemical, and molecular mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ae, N.; Murakami, M.; Wagatsuma, T. Is Brassica juncea a suitable plant for phytoremediation of cadmium in soils with moderately low cadmium contamination?—Possibility of using other plant species for Cd-phytoextraction. Soil Sci. Plant Nutr. 2006, 52, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Arao, T.; Ae, N. Genotypic variations in cadmium levels of rice grain. Soil Sci. Plant Nutr. 2003, 49, 473–479. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ae, N.; Sugiyama, M.; Murakami, M.; Arao, T. Genotypic variation in shoot cadmium concentration in rice and soybean in soils with different levels of cadmium contamination. Soil Sci. Plant Nutr. 2005, 51, 101–108. [Google Scholar] [CrossRef]
- Uraguchi, S.; Mori, S.; Kuramata, M.; Kawasaki, A.; Arao, T.; Ishikawa, S. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- Morishita, T.; Fumoto, N.; Yoshizawa, T.; Kagawa, K. Varietal differences in cadmium levels of rice grains of japonica, indica, javanica, and hybrid varieties produced in the same plot of a field. Soil Sci. Plant Nutr. 1987, 33, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Ae, N.; Ishikawa, S. Phytoextraction of cadmium by rice (Oryza sativa L.), soybean (Glycine max (L.) Merr.), and maize (Zea mays L.). Environ. Pollut. 2007, 145, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Ae, N.; Ishikawa, S.; Ibaraki, T.; Ito, M. Phytoextraction by a high-Cd-accumulating rice: Reduction of Cd content of soybean seeds. Environ. Sci. Technol. 2008, 42, 6167–6172. [Google Scholar] [CrossRef]
- Ibaraki, T.; Kuroyanagi, N.; Murakami, M. Practical phytoextraction in cadmium-polluted paddy fields using a high cadmium accumulating rice plant cultured by early drainage of irrigation water. Soil Sci. Plant Nutr. 2009, 55, 421–427. [Google Scholar] [CrossRef]
- Kojima, Y.; Ebana, K.; Fukuoka, S.; Nagamine, T.; Kawase, M. Development of an RFLP-based rice diversity research set of germplasm. Breed. Sci. 2005, 55, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Nakagawa, F.; Ae, N.; Ito, M.; Arao, T. Phytoextraction by rice capable of accumulating Cd at high levels: Reduction of Cd content of rice grain. Environ. Sci. Technol. 2009, 43, 5878–5883. [Google Scholar] [CrossRef]
- Ibaraki, T.; Fujitomi, S.; Ishitsuka, A.; Yanaka, M. Phytoextraction by high-Cd-accumulating rice to reduce Cd in wheat grains grown in Cd-polluted fields. Soil Sci. Plant Nutr. 2014, 60, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: Perspectives for reducing grain cadmium accumulation. Rice 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uraguchi, S.; Fujiwara, T. Rice breaks ground for cadmium-free cereals. Curr. Opin. Plant Biol. 2013, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zou, W.; Meng, L.; Fan, X.; Xu, G.; Ye, G. Advances in the uptake and transport mechanisms and QTLs mapping of cadmium in rice. Int. J. Mol. Sci. 2019, 20, 3417. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Koyama, E.; Kono, I.; Ando, T.; Yano, M.; Ma, J.F. Identification of a novel major quantitative trait locus controlling distribution of Cd between roots and shoots in rice. Plant Cell Physiol. 2009, 50, 2223–2233. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Abe, T.; Kuramata, M.; Yamaguchi, M.; Ando, T.; Yamamoto, T.; Yano, M. A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7. J. Exp. Bot. 2010, 61, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Tezuka, K.; Miyadate, H.; Katou, K.; Kodama, I.; Matsumoto, S.; Kawamoto, T.; Masaki, S.; Satoh, H.; Yamaguchi, M.; Sakurai, K. A single recessive gene controls cadmium translocation in the cadmium hyperaccumulating rice cultivar Cho-Ko-Koku. Theor. Appl. Genet. 2010, 120, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Taguchi-Shiobara, F.; Kojima, Y.; Ebitani, T.; Kuramata, M.; Yamamoto, T.; Yano, M.; Ishikawa, S. Detection of a QTL for accumulating Cd in rice that enables efficient Cd phytoextraction from soil. Breed. Sci. 2011, 61, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Suzui, N.; Ito-Tanabata, S.; Ishii, S.; Igura, M.; Abe, T.; Kuramata, M.; Kawachi, N.; Fujimaki, S. Real-time imaging and analysis of differences in cadmium dynamics in rice cultivars (Oryza sativa) using positron-emitting 107Cd tracer. BMC Plant Biol. 2011, 11, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Koyama, E.; Yamaji, N.; Ma, J.F. Physiological, genetic, and molecular characterization of a high-Cd-accumulating rice cultivar, Jarjan. J. Exp. Bot. 2011, 62, 2265–2272. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.F.; Fujii-Kashino, M.; Yamaji, N.; Fukuoka, S.; Shen, R.F.; Ma, J.F. Isolation and characterization of a rice line with high Cd accumulation for potential use in phytoremediation. Plant Soil 2017, 410, 357–368. [Google Scholar] [CrossRef]
- Sui, F.; Zhao, D.; Zhu, H.; Gong, Y.; Tang, Z.; Huang, X.Y.; Zhang, G.; Zhao, F.J. Map-based cloning of a new total loss-of-function allele of OsHMA3 causes high cadmium accumulation in rice grain. J. Exp. Bot. 2019, 70, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yang, M.; Li, Y.; Tian, J.; Zhang, Y.; Liang, L.; Liu, Z.; Chen, K.; Lv, K.; Lian, X. Comprehensive analysis of variation of cadmium accumulation in rice and detection of a new weak allele of OsHMA3. J. Exp. Bot. 2019, 70, 6389–6400. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Kuramata, M.; Iwasaki, M.; Honma, T.; Ibaraki, T.; Yamamoto, T.; Yano, M.; Murakami, M.; Ishikawa, S. “MJ3” and “MA22”, new high-cadmium-accumulating practical rice lines with non-shattering derived from gamma ray mutation. Breed Res. 2013, 15, 17–24, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Abe, T.; Ito, M.; Takahashi, R.; Honma, T.; Sekiya, N.; Shirao, K.; Kuramata, M.; Murakami, M.; Ishikawa, S. Breeding of a practical rice line ‘TJTT8’ for phytoextraction of cadmium contamination in paddy fields. Soil Sci. Plant Nutr. 2017, 63, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Ohta, H.; Nemoto, H.; Ando, I.; Kato, H.; Sato, H.; Hirabayashi, H.; Takeuchi, Y.; Ishii, T.; Maeda, H.; Imbe, T.; et al. “Tachisugata”, a new rice cultivar for whole crop silage use. Bull. Natl. Inst. Crop Sci. 2010, 11, 67–84, (In Japanese with English Summary). [Google Scholar]
- Takahashi, R.; Ito, M.; Katou, K.; Sato, K.; Nakagawa, S.; Tezuka, K.; Akagi, H.; Kawamoto, T. Breeding and characterization of the rice (Oryza sativa L.) line “Akita 110” for cadmium phytoremediation. Soil Sci. Plant Nutr. 2016, 62, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Kodama, I.; Kawamoto, T.; Matsumoto, S.; Sato, K.; Taguchi, M.; Kyoya, K.; Kato, T.; Hatakeyama, T.; Masaki, S. Breeding of a large grain rice cultivar exhibit high yeilds, “Akita 63”. Bull. Akita Agric. Exp. Stn. 2014, 54, 3–28, (In Japanese with English Summary). [Google Scholar]
- Takahashi, R.; Ito, M.; Kato, K.; Kodama, I.; Shibata, S.; Sato, K.; Matsumoto, S.; Kawamoto, T. Breeding and characterization of the high cadmium-accumulating rice line ‘Akita 119’. Breed. Sci. 2020, 70, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Arai-Sanoh, Y.; Takai, T.; Yoshinaga, S.; Nakano, H.; Kojima, M.; Sakakibara, H.; Kondo, M.; Uga, Y. Deep rooting conferred by DEEPER ROOTING 1 enhances rice yield in paddy fields. Sci. Rep. 2014, 4, 5563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitomi, Y.; Nakao, E.; Kawai, S.; Kanno, N.; Ando, T.; Fukuoka, S.; Irie, K.; Uga, Y. Fine mapping of QUICK ROOTING 1 and 2, quantitative trait loci increasing root length in rice. G3 Genes Genomes Genet. 2018, 8, 727–735. [Google Scholar]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; et al. Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Ishimaru, Y.; Igura, M.; Kuramata, M.; Abe, T.; Senoura, T.; Hase, Y.; Arao, T.; Nishizawa, N.K.; Nakanishi, H. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. USA 2012, 109, 19166–19171. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Ae, N.; Yano, M. Chromosomal regions with quantitative trait loci controlling cadmium concentration in brown rice (Oryza sativa). New Phytol. 2005, 168, 345–350. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Shindoh, K.; Hirotsu, N.; Ishimaru, K. Evidence for separate translocation pathways in determining cadmium accumulation in grain and aerial plant parts in rice. BMC Plant Biol. 2009, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Kono, I.; Yokosho, K.; Ando, T.; Yano, M.; Ma, J.F. A major quantitative trait locus controlling cadmium translocation in rice (Oryza sativa). New Phytol. 2009, 182, 644–653. [Google Scholar] [CrossRef]
- Arao, T.; Kawasaki, A.; Baba, K.; Mori, S.; Matsumoto, S. Effects of water management on cadmium and arsenic accumulation and dimethylarsinic acid concentrations in Japanese rice. Environ. Sci. Technol. 2009, 43, 9361–9367. [Google Scholar] [CrossRef]
- Siripan, O.; Thamchaipenet, A.; Surat, W. Enhancement of the efficiency of Cd phytoextraction using bacterial endophytes isolated from Chromolaena odorata, a Cd hyperaccumulator. Int. J. Phytoremediation. 2018, 20, 1096–1105. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, L.; Liu, Q.; Sikder, M.M.; Vestergard, M.; Zhou, K.; Wang, Q.; Yang, X.; Feng, Y. Pseudomonas fluorescens promote photosynthesis, carbon fixation and cadmium phytoremediation of hyperaccumulator Sedum alfredii. Sci. Total Environ. 2020, 726, 138554. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, L.; Ma, F.; You, Y.; Wang, Y.; Yang, D. Integration of earthworms and arbuscular mycorrhizal fungi into phytoremediation of cadmium-contaminated soil by Solanum nigrum L. J. Hazard. Mater. 2020, 389, 121873. [Google Scholar] [CrossRef]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal. Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef] [Green Version]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Menguer, P.K.; Farthing, E.; Peaston, K.A.; Ricachenevsky, F.K.; Fett, J.P.; Williams, L.E. Functional analysis of the rice vacuolar zinc transporter OsMTP1. J. Exp. Bot. 2013, 64, 2871–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, N.; Bhattacharya, S.; Maiti, M.K. Enhanced cadmium accumulation and tolerance in transgenic tobacco overexpressing rice metal tolerance protein gene OsMTP1 is promising for phytoremediation. Plant Physiol. Biochem. 2016, 105, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S.; et al. From laboratory to field: OsNRAMP5-knockdown rice is a promising candidate for Cd phytoremediation in paddy fields. PLoS ONE 2014, 9, e98816. [Google Scholar] [CrossRef] [PubMed]
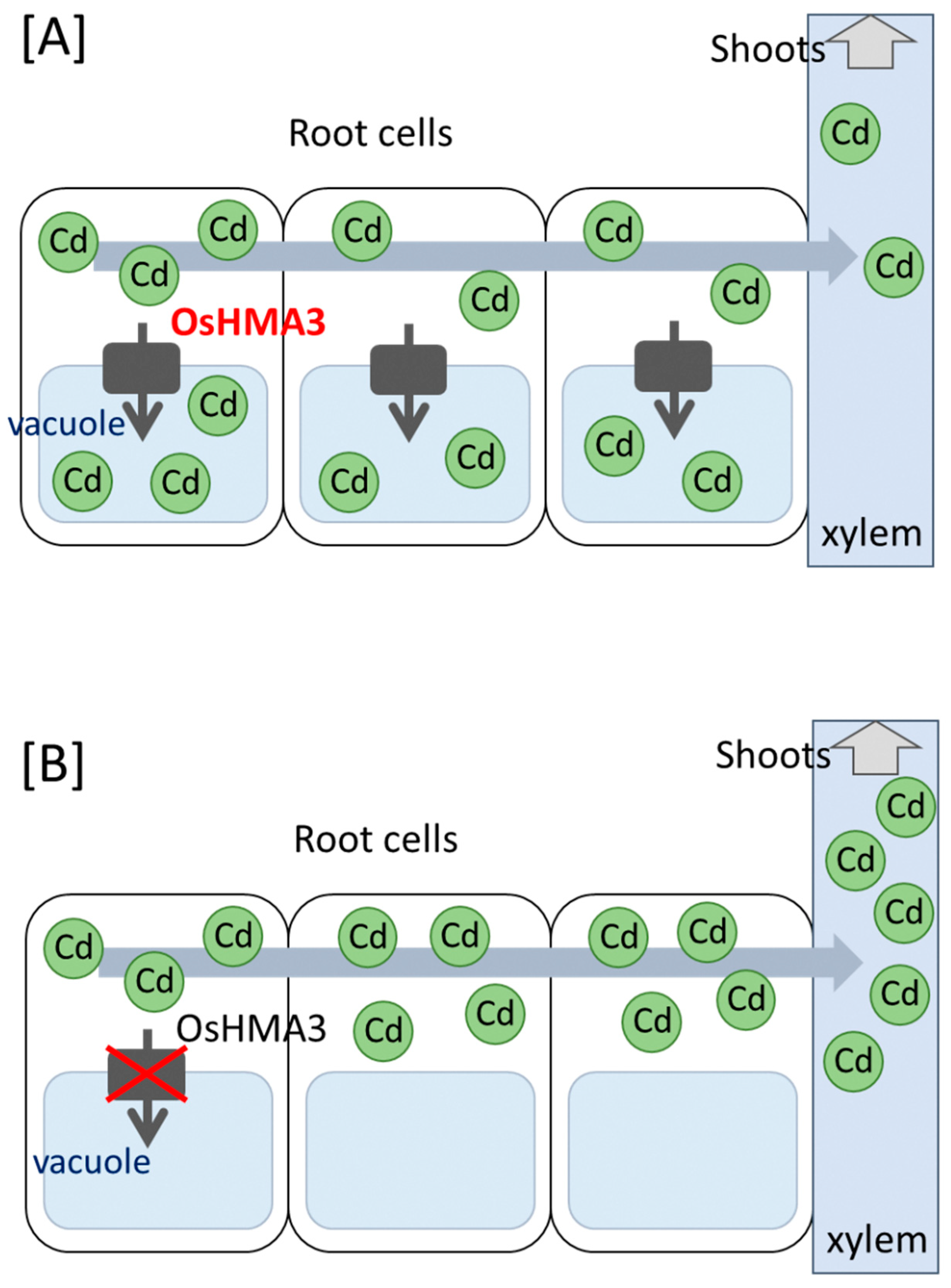
Figure 1.
Proposed model for the role of OsHMA3 in Cd translocation from the roots to the shoots. The Cd is sequestered into the vacuole via OsHMA3 in normal varieties (A), whereas not in high Cd-accumulating varieties (B). The remaining Cd in the cytoplasm is loaded into the xylem.
Figure 1.
Proposed model for the role of OsHMA3 in Cd translocation from the roots to the shoots. The Cd is sequestered into the vacuole via OsHMA3 in normal varieties (A), whereas not in high Cd-accumulating varieties (B). The remaining Cd in the cytoplasm is loaded into the xylem.

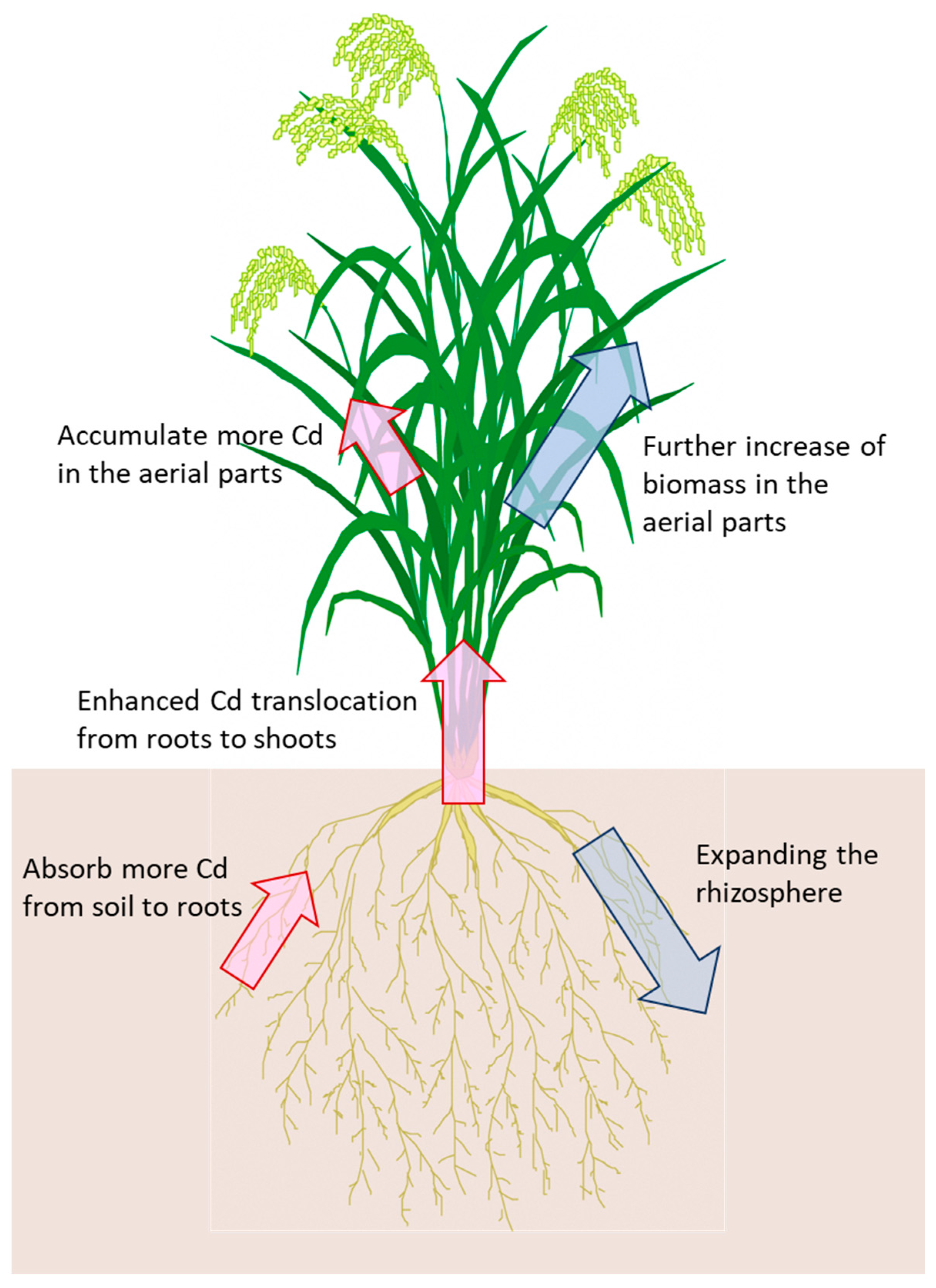
Figure 2.
The breeding goals for new rice varieties of Cd phytoremediation. Blue arrows and red arrows indicate morphological and physiological improvements, respectively.
Figure 2.
The breeding goals for new rice varieties of Cd phytoremediation. Blue arrows and red arrows indicate morphological and physiological improvements, respectively.

Table 2.
Cd extraction ratio. Each variety or line was cultivated in Cd contaminated field in Akita Prefecture. The ratio is calculated by the Cd extraction of ‘Cho-ko-koku’ cultivated in the same field as 1.00. “-” indicates no data.
Table 2.
Cd extraction ratio. Each variety or line was cultivated in Cd contaminated field in Akita Prefecture. The ratio is calculated by the Cd extraction of ‘Cho-ko-koku’ cultivated in the same field as 1.00. “-” indicates no data.
| Cd Extraction Ratio (Ratio with ‘Cho-ko-koku’ as 1.00) | Reference | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | Average | ||||
| Soil Cd Conc. (mg/kg) | 0.55 | 1.47 | 0.73 | 0.96 | 0.96 | 0.62 | 0.75 | 0.51 | 0.55 | ||
| Akita 119 | 1.10 | 1.02 | 1.23 | 1.06 | 0.92 | 1.09 | 1.16 | 0.89 | 1.00 | 1.05 | [42] |
| Akita 110 | 1.18 | 0.71 | 0.62 | 1.34 | 1.04 | 0.67 | 0.65 | - | 0.99 | 0.90 | [40] |
| MJ3 | - | - | - | 1.38 | - | 1.58 | - | - | - | 1.48 | [37] |
| MA22 | - | - | - | 1.64 | - | - | - | - | - | 1.64 | [37] |
| TJTT8 | - | - | - | 1.35 | - | 1.64 | - | 1.43 | - | 1.47 | [38] |
| Cho-ko-koku | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | [13,26] |
Table 3.
QTL region and the function reported previously.
| Function | Crossing Parents | ChromosomalRegion of QTL | Reference | |
|---|---|---|---|---|
| High-Cd | Low-Cd | |||
| Cd translocation from root to shoot | Anjana Dhan | Nipponbare | Chr. 7 | [24] |
| High Cd concentration in grain | Habataki | Sasanishiki | Chr. 7 | [25] |
| High Cd concentration in grain | Kasalath | Koshihikari | Chr. 6 | [48] |
| Low Cd concentration in grain | Kasalath | Koshihikari | Chr. 3, 8 | [48] |
| Cd translocation from root to aerial part | Kasalath | Nipponbare | Chr. 4, 11 | [49] |
| Cd translocation from root to shoot | Badari Dhan | Shwe War | Chr. 11 | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Takahashi, R.; Ito, M.; Kawamoto, T. The Road to Practical Application of Cadmium Phytoremediation Using Rice. Plants 2021, 10, 1926. https://doi.org/10.3390/plants10091926
AMA Style
Takahashi R, Ito M, Kawamoto T. The Road to Practical Application of Cadmium Phytoremediation Using Rice. Plants. 2021; 10(9):1926. https://doi.org/10.3390/plants10091926
Chicago/Turabian StyleTakahashi, Ryuichi, Masashi Ito, and Tomohiko Kawamoto. 2021. "The Road to Practical Application of Cadmium Phytoremediation Using Rice" Plants 10, no. 9: 1926. https://doi.org/10.3390/plants10091926
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.