
Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Field Sampling and Fungal Isolation
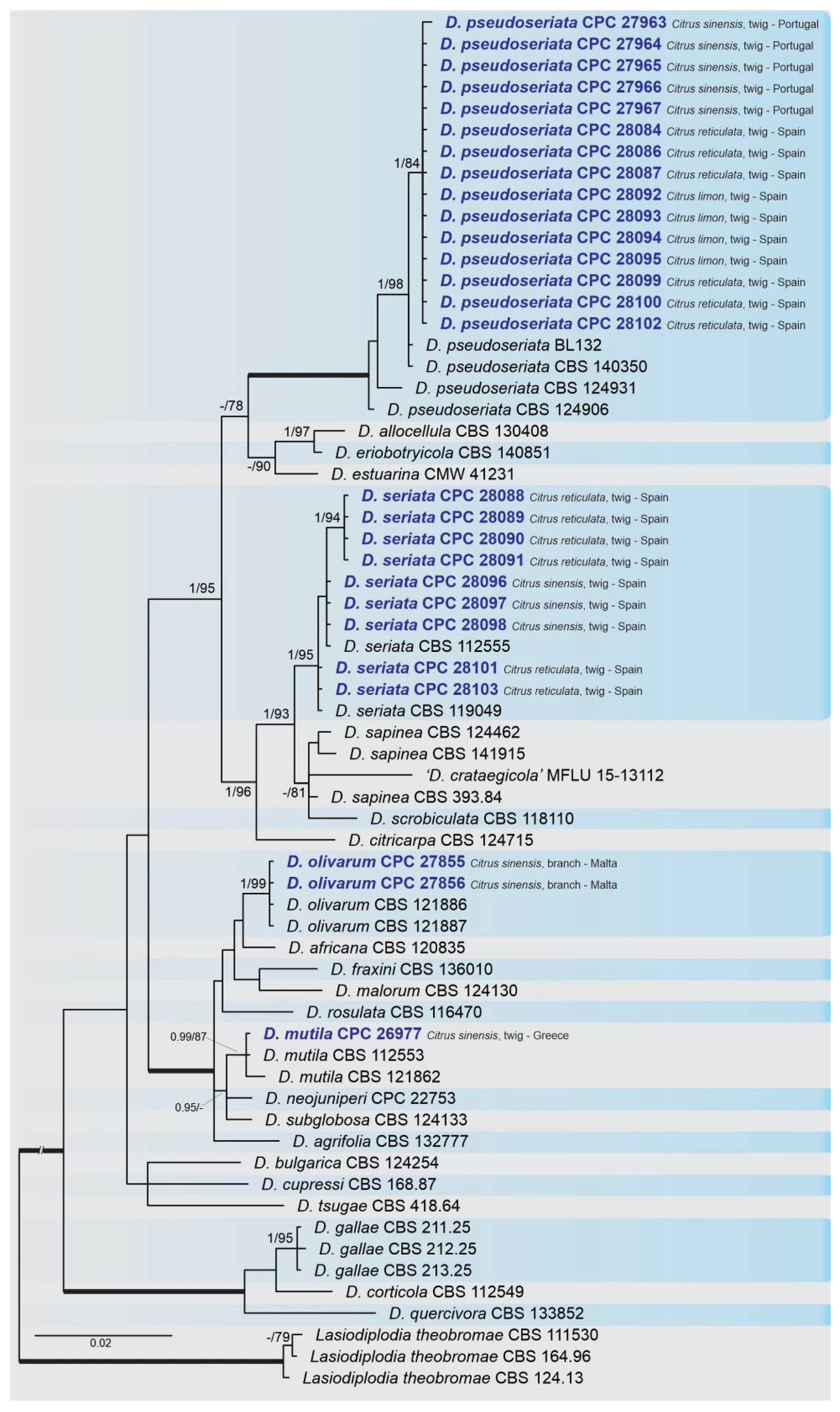
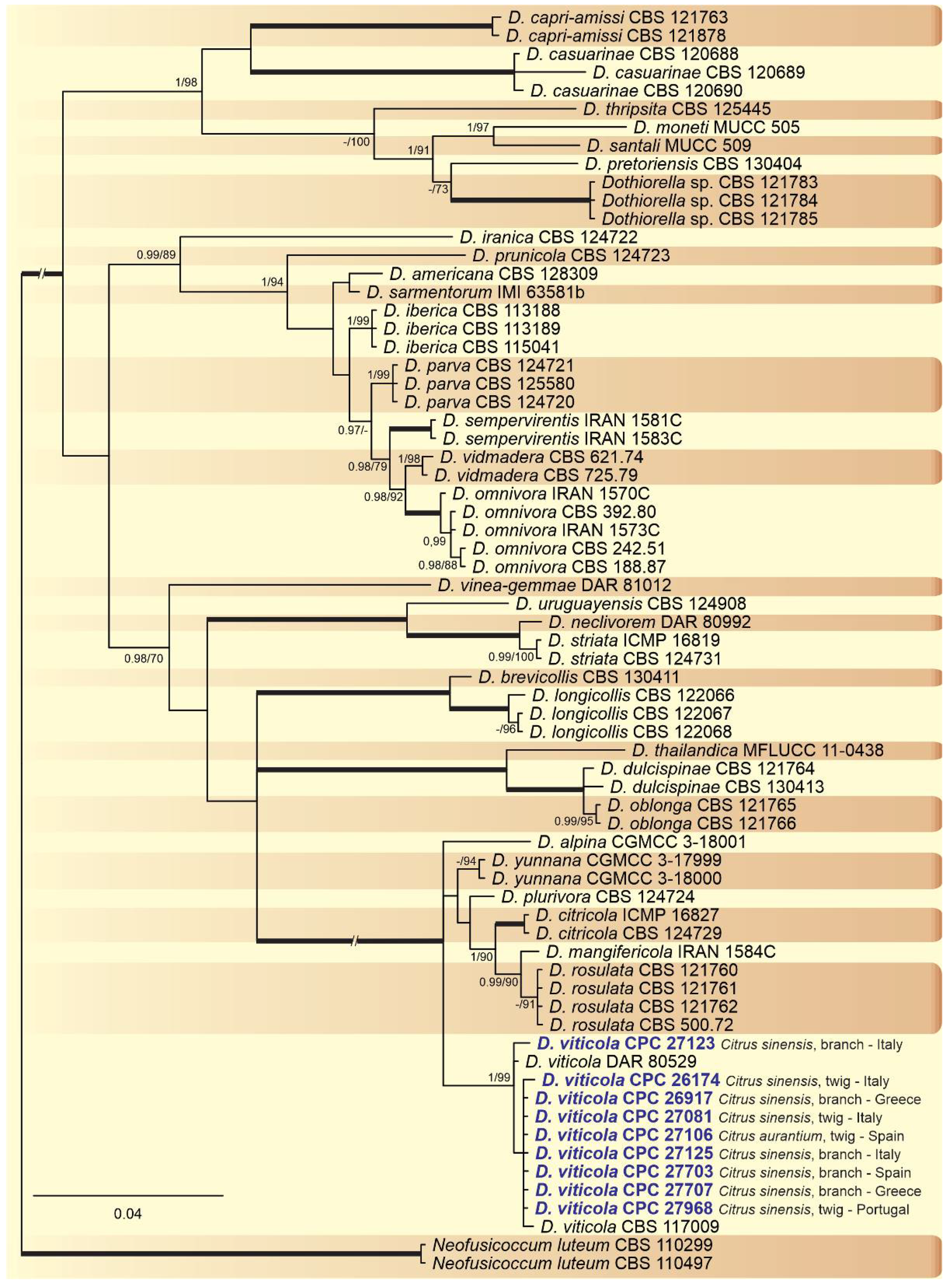
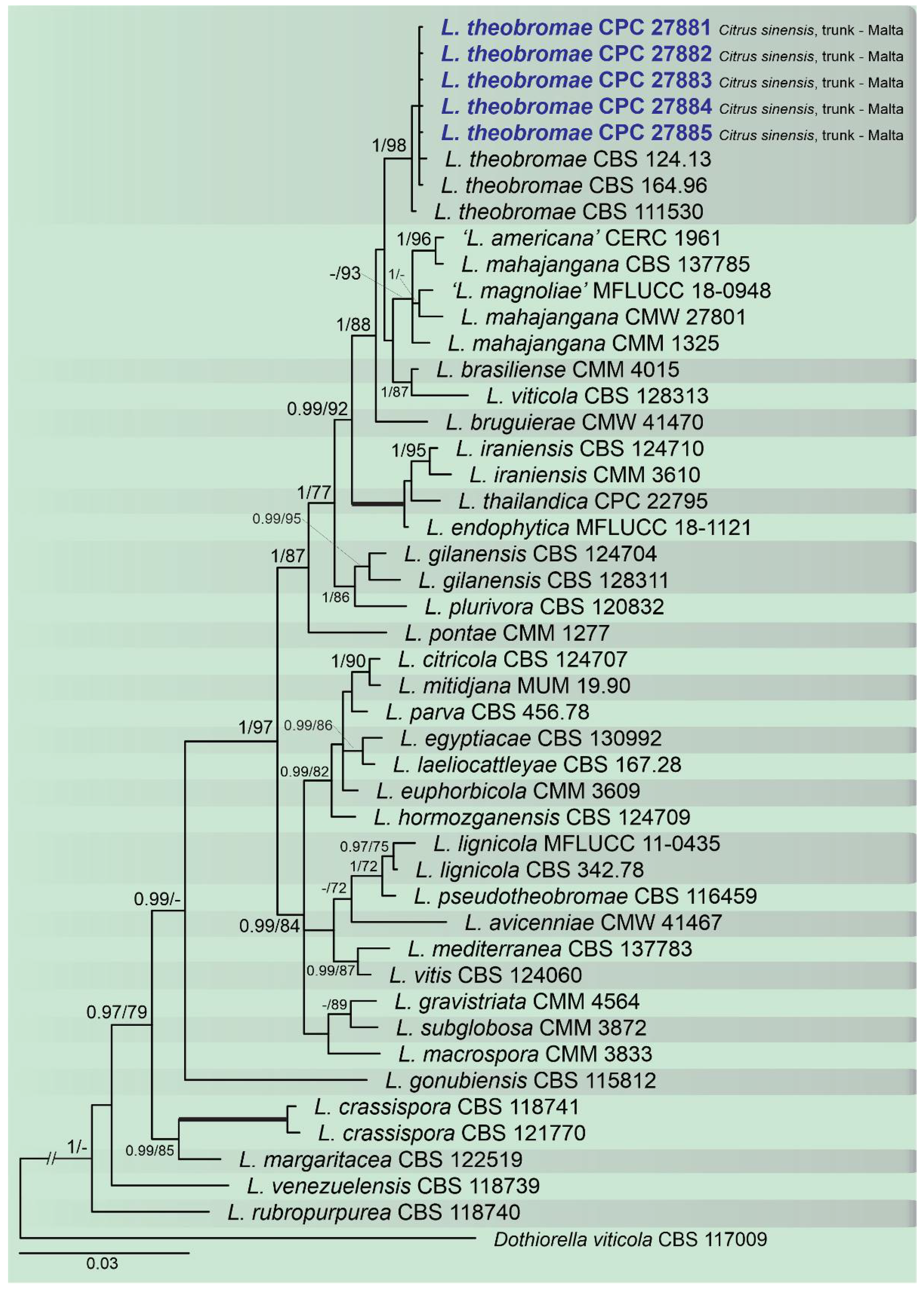
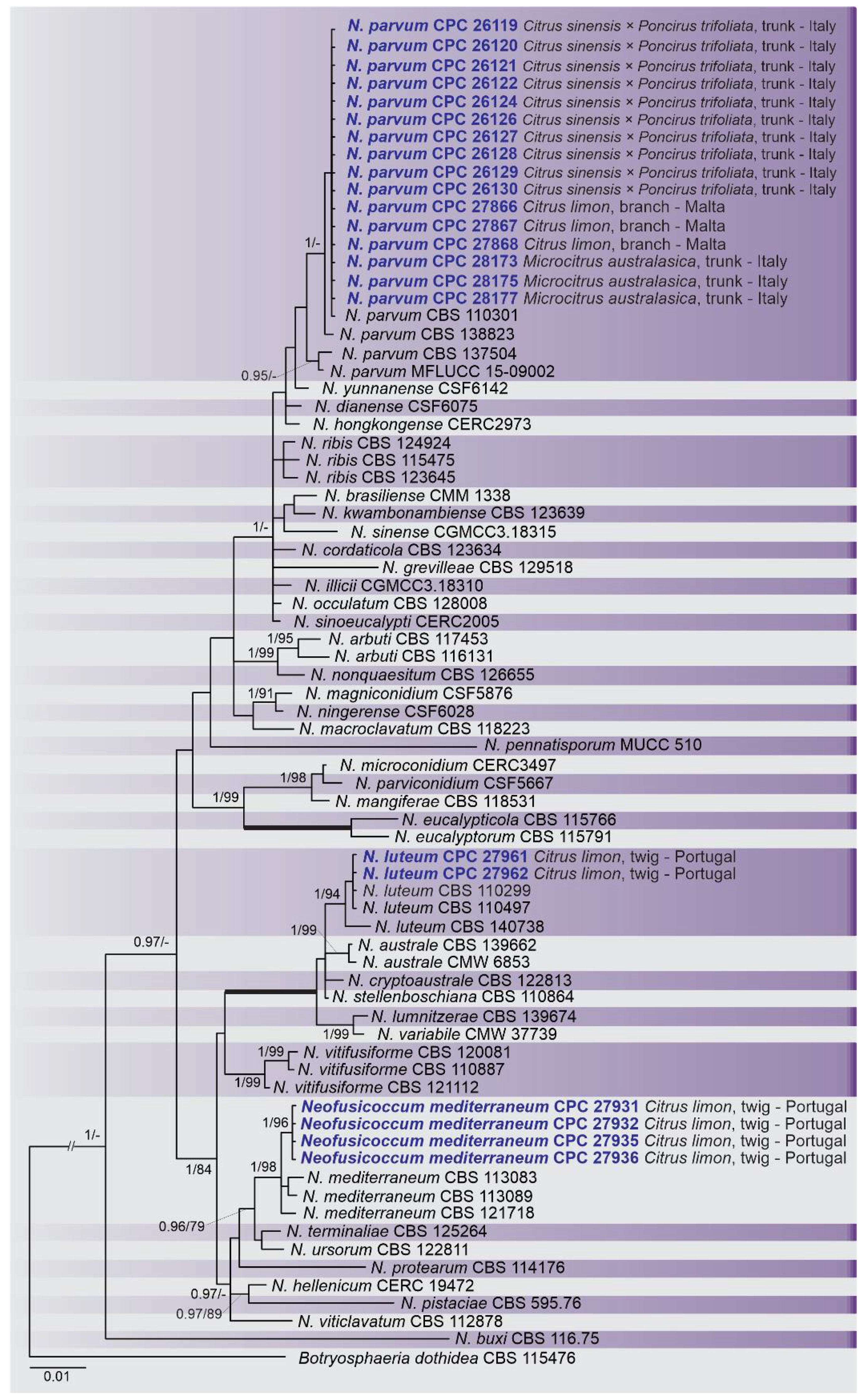
2.2. Phylogenetic Analyses
2.3. Occurrence of Botryosphaeriaceae among Countries and Citrus Species
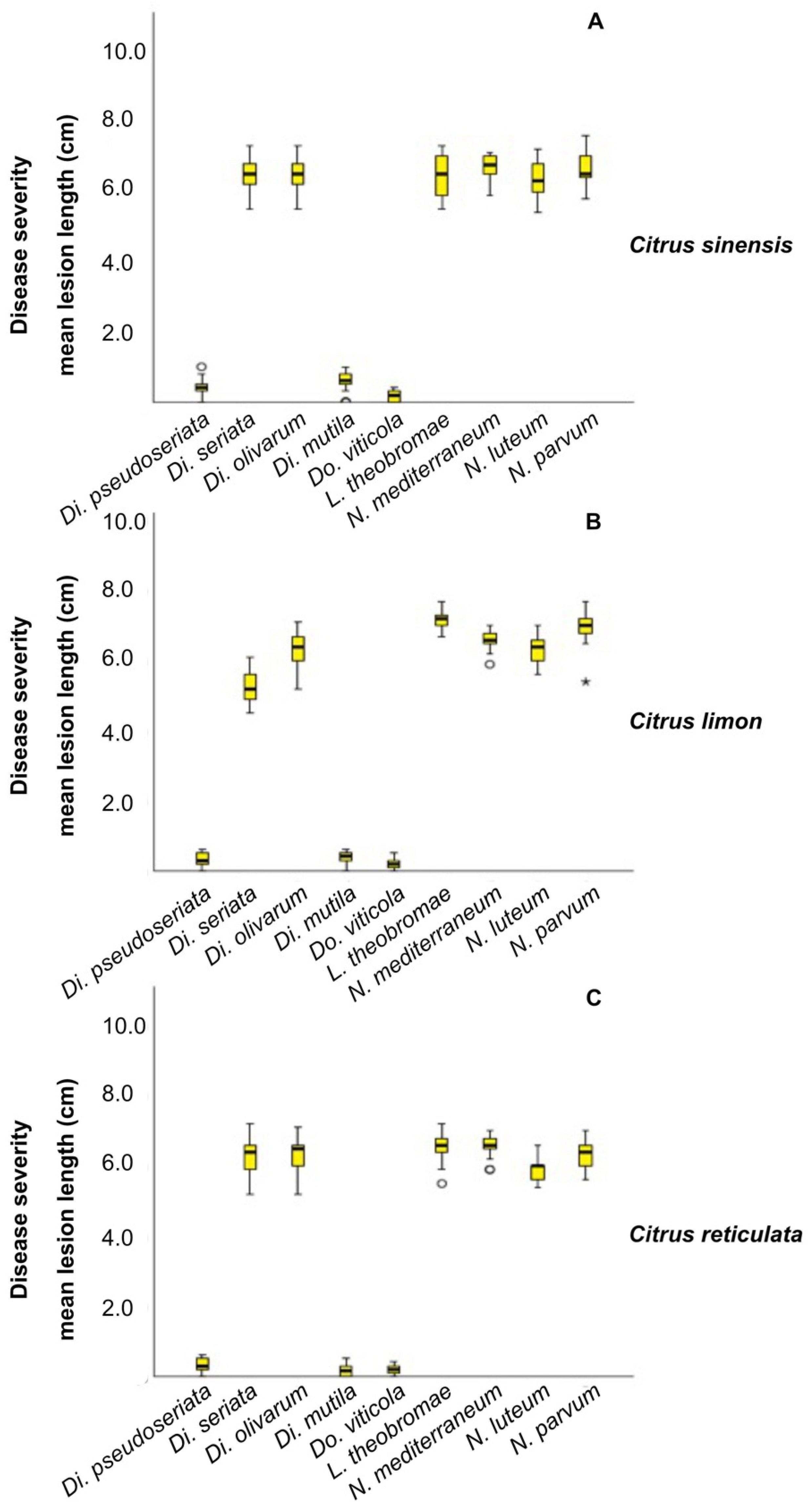
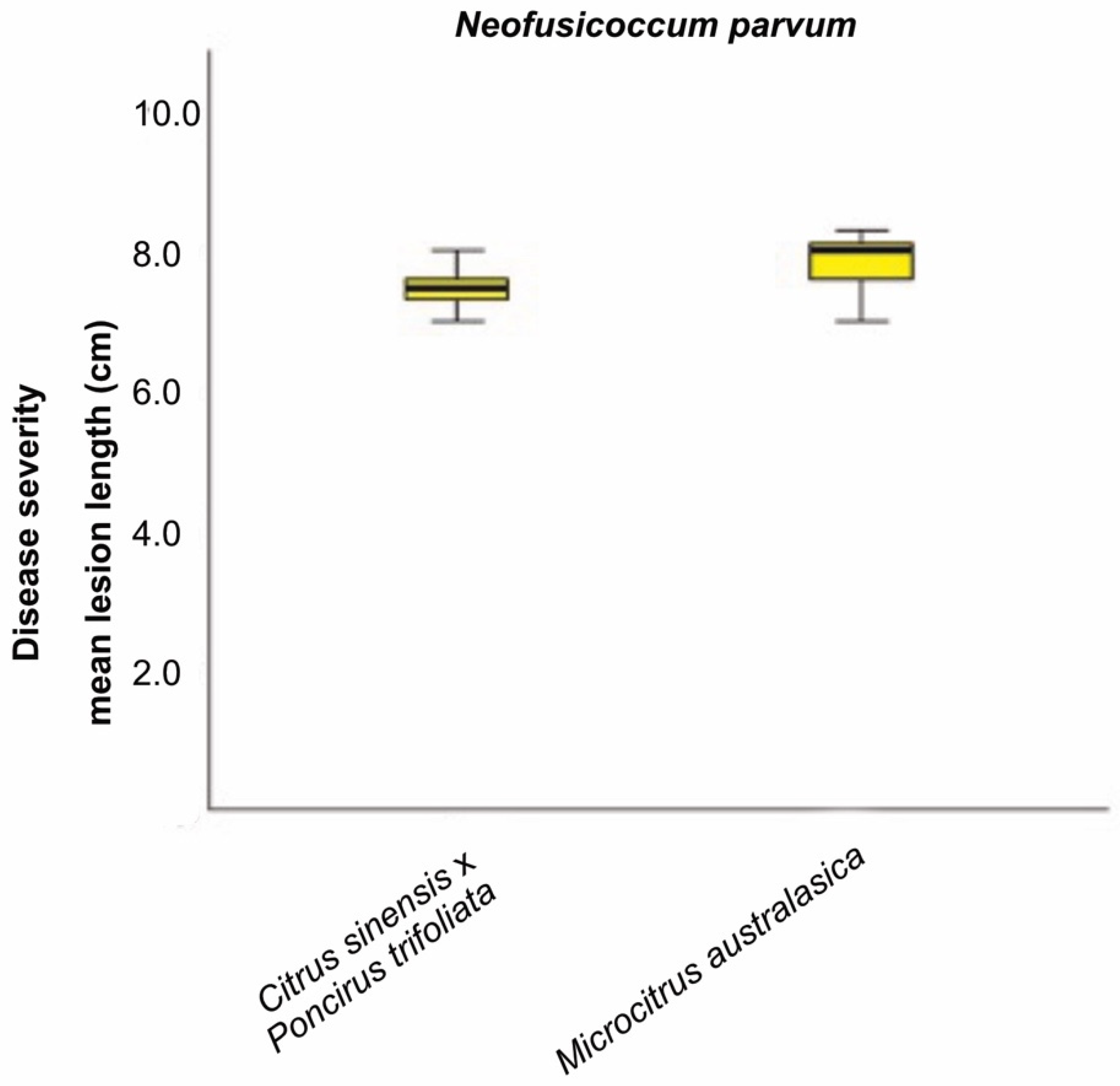
2.4. Pathogenicity Tests
3. Discussion
4. Materials and Methods
4.1. Field Sampling and Fungal Isolation
4.2. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification and Sequencing
4.3. Phylogenetic Analyses
4.4. Pathogenicity Tests
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2019. Available online: http://www.fao.org/faostat/en/#home (accessed on 26 February 2020).
- Eurostat. Citrus Fruit Statistics; Eurostat: Luxembourg, 2020. [Google Scholar]
- Crous, P.W.; Wingfield, M.J. Fungi infecting woody plants: Emerging frontiers. Persoonia 2018, 40, i–iii. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, H.S. Citrus Diseases and Their Control; McGraw-Hill: New York, NY, USA, 1936. [Google Scholar]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Huang, F.; Hou, X.; Dewdney, M.M.; Fu, Y.S.; Chen, G.Q.; Hyde, K.D.; Li, H. Diaporthe species occurring on citrus in China. Fungal Divers. 2013, 61, 237–250. [Google Scholar] [CrossRef]
- Mahadevakumar, S.; Yadav, V.; Tejaswini, G.S.; Sandeep, S.N.; Janardhana, G.R. First report of Phomopsis citri associated with dieback of Citrus lemon in India. Plant Dis. 2014, 98, 1281. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Species of Diaporthe on Camellia and Citrus in the Azores Islands. Phytopathol. Mediterr. 2018, 57, 307–319. [Google Scholar]
- Mayorquin, J.S.; Nouri, M.T.; Peacock, B.B.; Trouillas, F.P.; Douhan, G.W.; Kallsen, C.; Eskalen, A. Identification, Pathogenicity, and Spore Trapping of Colletotrichum karstii Associated with Twig and Shoot Dieback in California. Plant Dis. 2019, 103, 1464–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P.W. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia 2018, 40, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmer, L.W.; Garnsey, S.M.; Graham, J.H. Compendium of Citrus Diseases, 2nd ed.; American Phytopathological Society: Saint Paul, MN, USA, 2000. [Google Scholar]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with Citrus branch canker in the southern California desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espargham, N.; Mohammadi, H.; Gramaje, D.A. Survey of Trunk Disease Pathogens within Citrus Trees in Iran. Plants 2020, 9, 754. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Giraldo, A.; Hawksworth, D.L.; Robert, V.; Kirk, P.M.; Guarro, J.; Robbertse, B.; Schoch, C.L.; Damm, U.; Trakunyingcharoen, T.; et al. The genera of fungi: Fixing the application of type species of generic names. IMA Fungus 2014, 5, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Muller, M.M.; Sanchez, R.M.; Giordano, L.; Bianchinotti, M.V.; Anderson, F.E.; Groenewald, J.Z. Resolving Tiarosporella spp. allied to Botryosphaeriaceae and Phacidiaceae. Phytotaxa 2015, 202, 73–93. [Google Scholar] [CrossRef]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Cankers and other diseases caused by the Botryosphaeriaceae. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CAB International: Boston, MA, USA, 2013; pp. 298–317. [Google Scholar]
- Pour, F.N.; Ferreira, V.; Félix, C.; Serôdio, J.; Alves, A.; Duarte, A.S.; Esteves, A.C. Effect of temperature on the phytotoxicity and cytotoxicity of Botryosphaeriaceae fungi. Fungal Biol. 2020, 124, 571–578. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Martino, I.; Tabone, G.; Brondino, L.; Gullino, M.L. Fungal pathogens associated with stem blight and dieback of blueberry in northern Italy. Phytopathol. Mediterr. 2020, 59, 229–245. [Google Scholar]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Gusella, G.; Fiorenza, A.; Guarnaccia, V.; Polizzi, G. Identification of Neofusicoccum parvum causing canker and twig blight on Ficus carica in Italy. Phytopathol. Mediterr. 2020, 59, 213–218. [Google Scholar]
- Adesemoye, A.O.; Eskalen, A. First Report of Spencermartinsia viticola, Neofusicoccum australe, and N. parvum causing branch canker of citrus in California. Plant Dis. 2011, 95, 770. [Google Scholar] [CrossRef] [PubMed]
- Polizzi, G.; Aiello, D.; Vitale, A.; Giuffrida, F.; Groenewald, J.; Crous, P.W. First report of shoot blight, canker, and gummosis caused by Neoscytalidium dimidiatum on citrus in Italy. Plant Dis. 2009, 93, 1215. [Google Scholar] [CrossRef]
- Berraf-Tebbal, A.; Mahamedi, A.E.; Aigoun-Mouhous, W.; Špetík, M.; Čechová, J.; Pokluda, R.; Baránek, M.; Eichmeier, A.; Alves, A. Lasiodiplodia mitidjana sp. nov. and other Botryosphaeriaceae species causing branch canker and dieback of Citrus sinensis in Algeria. PLoS ONE 2020, 15, e0232448. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hamrouni, N.; Nouri, M.; Trouillas, F.; Said, A.; Sadfi-Zouaoui, N.; Hajlaoui, M. Dothiorella gummosis caused by Dothiorella viticola, first record from citrus in Tunisia. New Dis. Rep. 2018, 38, 10. [Google Scholar] [CrossRef] [Green Version]
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of species of Botryosphaeriaceae causing bot gummosis in citrus in California. Plant Dis. 2014, 98, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakalounakis, D.J.; Ntougias, S.; Kavroulakis, N.; Protopapadakis, E. Neofusicoccum parvum and Diaporthe foeniculina associated with twig and shoot blight and branch canker of citrus in Greece. J. Phytopathol. 2019, 167, 527–537. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Li, H.; Glienke, C.; Carstens, E.; Hattingh, V.; Crous, P.W. First report of Phyllosticta citricarpa and description of two new species, P. paracapitalensis and P. paracitricarpa, from citrus in Europe. Stud. Mycol. 2017, 87, 161–185. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Boutiti, M.Z.; Jamâa, M.L.B.; Phillips, A.J.L. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Mahamedi, A.E.; Phillips, A.J.L.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, distribution and host association of Botryosphaeriaceae species causing oak decline across different forest ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765. [Google Scholar] [CrossRef]
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q. Current status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2018, 48, 35–44. [Google Scholar] [CrossRef]
- Machado, A.R.; Custódio, F.A.; Cabral, P.G.C.; Capucho, A.S.; Pereira, O.L. Botryosphaeriaceae species causing dieback on Annonaceae in Brazil. Plant Pathol. 2019, 68, 1394–1406. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, H.; Zhou, Z.; Hu, T.; Wang, S.; Wang, Y.; Cao, K. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Li, G.; Slippers, B.; Wingfield, M.J.; Chen, S. Variation in Botryosphaeriaceae from Eucalyptus plantations in YunNan Province in southwestern China across a climatic gradient. IMA Fungus 2020, 11, 22. [Google Scholar] [CrossRef]
- Valencia, A.L.; Pilar, M.; Gil, B.A.; Latorre, I.; Rosales, M. Characterization and Pathogenicity of Botryosphaeriaceae Species Obtained from Avocado Trees with Branch Canker and Dieback and from Avocado Fruit with Stem End Rot in Chile. Plant Dis. 2019, 103, 996–1005. [Google Scholar] [CrossRef]
- Gusella, G.; Aiello, D.; Polizzi, G. First report of leaf and twig blight of Indian hawthorn (Rhaphiolepis indica) caused by Neofusicoccum parvum in Italy. J. Plant Pathol. 2020, 102, 275. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.; Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Phillips, A.J.L. The complex of Diplodia species associated with Fraxinus and some other woody hosts in Italy and Portugal. Fungal Divers. 2014, 67, 143–156. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Yan, J.Y.; Li, X.H. Saprobic Botryosphaeriaceae, including Dothiorella italica sp nov., associated with urban and forest trees in Italy. Mycosphere 2017, 8, 1157–1176. [Google Scholar] [CrossRef]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Loots, M.T.; Gryzenhout, M. Temporal and spatial variation of Botryosphaeriaceae associated with Acacia karroo in South Africa. Fungal Ecol. 2015, 15, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Verkley, G.J.M.; Groenewald, J.Z.; Samson, R.A. (Eds.) Fungal Biodiversity; CBS Laboratory Manual Series 1; Centraalbureau Voor Schimmelcultures: Utrecht, The Netherlands, 2009; pp. 1–269. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, A.M., Gelfard, D.H., Snindky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest v. 2 Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Locality | GPS Coordinates |
|---|---|---|
| 1 | Algemesi (Spain) | 39°11′48.8″ N, 0°28′15.0″ W |
| 2 | Alginet (Spain) | 39°15′36.3″ N, 0°27′28.9″ W |
| 3 | Alhaurin El Grande (Spain) | 36°38′43.4″ N, 4°40′37.5″ W |
| 4 | Alzira (Spain) | 39°09′25.1″ N, 0°29′26.6″ W |
| 5 | Castellò (Spain) | 39°54′14.1″ N, 0°05′10.3″ W |
| 6 | Estellencs (Spain) | 39°39′12.6″ N, 2°28′54.8″ E |
| 7 | Faro (Portugal) | 37°03′45.5″ N, 7°55′02.8″ W |
| 8 | Gozo (Malta) | 36°02′15.1″ N, 14°15′36.4″ E |
| 9 | Gozo (Malta) | 36°03′18.5″ N, 14°15′35.7″ E |
| 10 | Malaga (Spain) | 36°45′42.3″ N, 4°25′37.4″ W |
| 11 | Mascali (Italy) | 37°46′05.7″ N, 15°11′40.7″ E |
| 12 | Massafra (Italy) | 40°32′41.1″ N, 17°08′38.8″ E |
| 13 | Mastro (Greece) | 38°25′49.0″ N, 21°16′49.9″ E |
| 14 | Mesquita (Portugal) | 37°12′16.3″ N, 8°17′52.1″ W |
| 15 | Moncada (Spain) | 39°35′18.8″ N, 0°23′40.5″ W |
| 16 | Nafplio (Greece) | 37°34′56.3″ N, 22°41′48.5″ E |
| 17 | Rocca Imperiale (Italy) | 40°06′30.2″ N, 16°37′04.6″ E |
| 18 | Scordia (Italy) | 37°16′53.5″ N, 14°52′08.9″ E |
| 19 | Silves (Portugal) | 37°09′50.7″ N, 8°23′21.7″ W |
| Species | Strains 1 | Host/Substrate | Country | GenBank Numbers 2 | ||
|---|---|---|---|---|---|---|
| ITS | TEF1 | TUB2 | ||||
| Botryosphaeria dothidea | CBS 115476 = CMW 8000, ex-epitype | Prunus sp. | Switzerland | AY236949 | AY236898 | AY236927 |
| Diplodia africana | CBS 120835 = CPC 5908, ex-type | Prunus persica, stem canker | South Africa | EF445343 | EF445382 | KF766129 |
| Di. agrifolia | CBS 132777 = UCR732, ex-type | Quercus agrifolia, cankered branch | USA: California | JN693507 | JQ517317 | JQ411459 |
| Di. allocellula | CBS 130408 = CMW 36468, ex-type | Acacia karroo, healthy branches | South Africa | JQ239397 | JQ239384 | JQ239378 |
| Di. bulgarica | CBS 124254 = CAP332, ex-type | Malus sylvestris | Bulgaria | GQ923853 | GQ923821 | – |
| Di. citricarpa | CBS 124715 = CJA 131 = IRAN 1578C, ex-type | Citrus sp., twigs | Iran | KF890207 | KF890189 | KX464784 |
| Di. corticola | CBS 112549 = CAP 134, ex-type | Quercus suber | Portugal | AY259100 | AY573227 | DQ458853 |
| Di. crataegicola | MFLU 15-13112, ex-type | Crataegus sp. | Italy | KT290244 | KT290248 | KT290246 |
| Di. cupressi | CBS 168.87, ex-type | Cupressus sempervirens, canker | Israel | DQ458893 | DQ458878 | DQ458861 |
| Di. eriobotryicola | CBS 140851 = BN-21, ex-type | Eriobotrya japonica | Spain | KT240355 | KT240193 | MG015806 |
| Di. estuarina | CMW 41231 | Avicennia marina | South Africa | KP860831 | KP860676 | KP860754 |
| Di. fraxini | CBS 136010 = CAD001, ex-type | Fraxinus angustifolia | Portugal | KF307700 | KF318747 | MG015807 |
| Di. gallae | CBS 211.25 | Quercus sp., fruit | – | KX464090 | KX464564 | KX464795 |
| Di. gallae | CBS 212.25 | Quercus sp., gall | – | KX464091 | KX464565 | KX464796 |
| Di. gallae | CBS 213.25 | Quercus sp., gall | – | KX464092 | KX464566 | KX464797 |
| Di. malorum | CBS 124130 = CAP271, ex-type | Malus sylvestris | Portugal | GQ923865 | GQ923833 | – |
| Di. mutila | CPC 26977 | Citrus sinensis, twig | Greece | MW413831 | MW419149 | MW419212 |
| Di. mutila | CBS 112553 = CAP 062 | Vitis vinifera | Portugal | AY259093 | AY573219 | DQ458850 |
| Di. mutila | CBS 121862 = PD 03708098, ex-type of Di. pyri | Pyrus sp. | The Netherlands | KX464093 | KX464567 | KX464799 |
| Di. neojuniperi | CPC 22753 = B0031, ex-type | Juniperus chinensis | Thailand | KM006431 | KM006462 | – |
| Di. olivarum | CPC 27855 | Citrus sinensis,branch | Malta | MW413832 | MW419150 | MW419213 |
| Di. olivarum | CPC 27856 | Citrus sinensis,branch | Malta | MW413833 | MW419151 | MW419214 |
| Di. olivarum | CBS 121886 | Olea europaea | Italy | EU392301 | EU392278 | – |
| Di. olivarum | CBS 121887 = CAP 254, ex-type | Olea europaea, rotting drupes | Italy | EU392302 | EU392279 | HQ660079 |
| Di. pseudoseriata | CBS 124906, ex-type | Blepharocalyx salicifolius | Uruguay | EU080927 | EU863181 | MG015820 |
| Di. pseudoseriata | CPC 27963 | Citrus sinensis, twig | Portugal | MW413834 | MW419152 | MW419215 |
| Di. pseudoseriata | CPC 27964 | Citrus sinensis, twig | Portugal | MW413835 | MW419153 | MW419216 |
| Di. pseudoseriata | CPC 27965 | Citrus sinensis, twig | Portugal | MW413836 | MW419154 | MW419217 |
| Di. pseudoseriata | CPC 27966 | Citrus sinensis, twig | Portugal | MW413837 | MW419155 | MW419218 |
| Di. pseudoseriata | CPC 27967 | Citrus sinensis, twig | Portugal | MW413838 | MW419156 | MW419219 |
| Di. pseudoseriata | CPC 28084 | Citrus reticulata, twig | Spain | MW413839 | MW419157 | MW419220 |
| Di. pseudoseriata | CPC 28086 | Citrus reticulata, twig | Spain | MW413840 | MW419158 | MW419221 |
| Di. pseudoseriata | CPC 28087 | Citrus reticulata, twig | Spain | MW413841 | MW419159 | MW419222 |
| Di. pseudoseriata | CPC 28092 | Citrus limon, twig | Spain | MW413842 | MW419160 | MW419223 |
| Di. pseudoseriata | CPC 28093 | Citrus limon, twig | Spain | MW413843 | MW419161 | MW419224 |
| Di. pseudoseriata | CPC 28094 | Citrus limon, twig | Spain | MW413844 | MW419162 | MW419225 |
| Di. pseudoseriata | CPC 28095 | Citrus limon, twig | Spain | MW413845 | MW419163 | MW419226 |
| Di. pseudoseriata | CPC 28099 | Citrus reticulata, twig | Spain | MW413846 | MW419164 | MW419227 |
| Di. pseudoseriata | CPC 28100 | Citrus reticulata, twig | Spain | MW413847 | MW419165 | MW419228 |
| Di. pseudoseriata | CPC 28102 | Citrus reticulata, twig | Spain | MW413848 | MW419166 | MW419229 |
| Di. pseudoseriata | BL132 | Fraxinus angustifolia | Italy | KF307720 | KF318767 | MG015810 |
| Di. pseudoseriata | CBS 140350, ex-type of Di. insularis | Pistacia lentiscus | Italy | KX833072 | KX833073 | MG015809 |
| Di. pseudoseriata | CBS 124931 = CMW22627, ex-type of Di. alatafructa | Pterocarpus angolensis, bark wound | South Africa | FJ888460 | FJ888444 | MG015799 |
| Di. quercivora | CBS 133852 = BL8, ex-type | Quercus canariensis | Tunisia | JX894205 | JX894229 | MG015821 |
| Di. rosulata | CBS 116470, ex-type | Prunus africana | Ethiopia | EU430265 | EU430267 | EU673132 |
| Di. sapinea | CBS 393.84, ex-epitype | Pinus nigra, cones | Netherlands | DQ458895 | DQ458880 | DQ458863 |
| Di. sapinea | CBS 124462 = CAP273, ex-type of Di. intermedia | Malus sylvestris | Portugal | GQ923858 | GQ923826 | – |
| Di. sapinea | CBS 141915 = NB7, ex-type of Di. rosacearum | Eriobotrya japonica | Italy | KT956270 | KU378605 | MG015823 |
| Di. scrobiculata | CBS 118110 = CMW 189 = BOT 1195, ex-type | Pinus banksiana | USA: Wisconsin | AY253292 | AY624253 | AY624258 |
| Di. seriata | CBS 112555 = HAP 052 = CAP 063, ex-epitype | Vitis vinifera, dead stems | Portugal | AY259094 | AY573220 | DQ458856 |
| Di. seriata | CPC 28088 | Citrus reticulata,twig | Spain | MW413849 | MW419167 | MW419230 |
| Di. seriata | CPC 28089 | Citrus reticulata,twig | Spain | MW413850 | MW419168 | MW419231 |
| Di. seriata | CPC 28090 | Citrus reticulata,twig | Spain | MW413851 | MW419169 | MW419232 |
| Di. seriata | CPC 28091 | Citrus reticulata,twig | Spain | MW413852 | MW419170 | MW419233 |
| Di. seriata | CPC 28096 | Citrus sinensis,twig | Spain | MW413853 | MW419171 | MW419234 |
| Di. seriata | CPC 28097 | Citrus sinensis,twig | Spain | MW413854 | MW419172 | MW419235 |
| Di. seriata | CPC 28098 | Citrus sinensis,twig | Spain | MW413855 | MW419173 | MW419236 |
| Di. seriata | CPC 28101 | Citrus reticulata,twig | Spain | MW413856 | MW419174 | MW419237 |
| Di. seriata | CPC 28103 | Citrus reticulata,twig | Spain | MW413857 | MW419175 | MW419238 |
| Di. seriata | CBS 119049 | Vitis sp. | Italy | DQ458889 | DQ458874 | DQ458857 |
| Di. subglobosa | CBS 124133 = JL453, ex-type | Lonicera nigra | Spain | GQ923856 | GQ923824 | – |
| Di. tsugae | CBS 418.64 = IMI 197143, ex-isotype | Tsuga heterophylla | Canada | DQ458888 | DQ458873 | DQ458855 |
| Dothiorella alpina | CGMCC 3-18001, ex-type | Platycladus orientalis | China | KX499645 | KX499651 | – |
| Do. americana | CBS 128309, ex-type | Wedge-shape canker of grapevine cv. Vignoles (complex hybrid of North America Vitis species and Vitis vinifera) | USA: Missouri | HQ288218 | HQ288262 | HQ288297 |
| Do. brevicollis | CBS 130411 = CMW 36463, ex-type | Acacia karroo, healthy branches | South Africa | JQ239403 | JQ239390 | JQ239371 |
| Do. capri-amissi | CBS 121763 = CMW 25403 = CAMS 1158, ex-paratype | Acacia erioloba | South Africa | EU101323 | EU101368 | KX464850 |
| Do. capri-amissi | CBS 121878 = CMW 25404 = CAMS 1159, ex-type | Acacia erioloba | South Africa | EU101324 | EU101369 | KX464851 |
| Do. casuarinae | CBS 120688 = CMW 4855, ex-type | Casuarina sp. | Australia: Australian Capital Territory | DQ846773 | DQ875331 | DQ875340 |
| Do. casuarinae | CBS 120689 = CMW 4856, ex-paratype | Casuarina sp. | Australia: Australian Capital Territory | DQ846772 | DQ875332 | DQ875339 |
| Do. casuarinae | CBS 120690 = CMW 4857, ex-paratype | Casuarina sp. | Australia: Australian Capital Territory | DQ846774 | DQ875333 | DQ875341 |
| Do. citricola | CBS 124728 = ICMP 16827 | Citrus sinensis | New Zealand | EU673322 | EU673289 | KX464852 |
| Do. citricola | CBS 124729 = ICMP 16828, ex-type | Citrus sinensis, twigs | New Zealand | EU673323 | EU673290 | KX464853 |
| Do. dulcispinae | CBS 121764 = CMW 25406 = CAMS 1159, ex-paratype of Dothiorella oblonga | Acacia mellifera | Namibia | EU101299 | EU101344 | KX464854 |
| Do. dulcispinae | CBS 130413 = CMW 36460, ex-type | Acacia karroo, dieback branches | South Africa | JQ239400 | JQ239387 | JQ239373 |
| Do. iberica | CBS 113188 = DA-1 | Quercus suber, branch canker | Spain | AY573198 | EU673278 | EU673097 |
| Do. iberica | CBS 113189 = DE-14 | Quercus ilex, branch canker | Spain | AY573199 | AY573230 | KX464855 |
| Do. iberica | CBS 115041 = CAP 145, ex-type | Quercus ilex, dead twigs | Spain | AY573202 | AY573222 | EU673096 |
| Do. iranica | CBS 124722 = CJA 153 = IRAN 1587C | Olea sp., twigs | Iran | KC898231 | KC898214 | KX464856 |
| Do. longicollis | CBS 122066 = CMW 26164 | Terminalia sp. | Australia: Western Australia | EU144052 | EU144067 | KX464857 |
| Do. longicollis | CBS 122067 = CMW 26165 | Lysiphyllum cunninghamii | Australia: Western Australia | EU144053 | EU144068 | KX464858 |
| Do. longicollis | CBS 122068 = CMW 26166, ex-type | Lysiphyllum cunninghamii | Australia: Western Australia | EU144054 | EU144069 | KF766130 |
| Do. mangifericola | CBS 124727 = IRAN 1584C = CJA 261, ex-type | Mangifera indica, twigs | Iran | KC898221 | KX464614 | – |
| Do. moneti | WAC 13154 = MUCC 505, ex-type | Acacia rostellifera, healthy stem | Australia: Western Australia | EF591920 | EF591971 | EF591954 |
| Do. neclivorem | DAR 80992, ex-type | Vitis vinifera cv. Chardonnay, berries | Australia: New South Wales | KJ573643 | KJ573640 | KJ577551 |
| Do. oblonga | CBS 121765 = CMW 25407 = CAMS 1162, ex-type | Acacia mellifera | South Africa | EU101300 | EU101345 | KX464862 |
| Do. oblonga | CBS 121766 = CMW 25408 = CAMS 1163, ex-paratype | Acacia mellifera | South Africa | EU101301 | EU101346 | KX464863 |
| Do. omnivora | CBS 124717 = CJA 214 = IRAN 1570C | Juglans regia, twigs | Iran | KC898233 | KC898216 | KX464865 |
| Do. omnivora | CBS 392.80 | – | France | KX464133 | KX464626 | KX464897 |
| Do. omnivora | CBS 124716 = CJA 241 = IRAN 1573C | Juglans regia, twigs | Iran | KC898232 | KC898215 | KX464864 |
| Do. omnivora | CBS 242.51 | – | Italy | EU673317 | EU673284 | EU673105 |
| Do. omnivora | CBS 188.87 | Juglans regia | France | EU673316 | EU673283 | EU673119 |
| Do. parva | CBS 124720 = CJA 27 = IRAN 1579C, ex-type | Corylus sp., twigs | Iran | KC898234 | KC898217 | KX464866 |
| Do. parva | CBS 124721 = CJA 35 | Corylus sp., twigs | Iran | KX464123 | KX464615 | KX464867 |
| Do. parva | CBS 125580 | Corylus avellana, branches | Austria | KX464124 | KX464616 | KX464868 |
| Do. plurivora | CBS 124724 = CJA 254 = IRAN 1557C, ex-type | Citrus sp., twigs | Iran | KC898225 | KC898208 | KX464874 |
| Do. pretoriensis | CBS 130404 = CMW 36480, ex-type | Acacia karroo, branches with dieback | South Africa | JQ239405 | JQ239392 | JQ239376 |
| Do. prunicola | CBS 124723 = CAP 187 = IRAN 1541C, ex-type | Prunus dulcis, necrotic twigs | Portugal | EU673313 | EU673280 | EU673100 |
| Do. rosulata | CBS 121760 = CMW 25389 = CAMS 1444, ex-type | Acacia karroo | Namibia | KF766227 | EU101335 | KX464877 |
| Do. rosulata | CBS 121761 = CMW 25392 = CAMS 1147, ex-paratype | Acacia mellifera | South Africa | EU101293 | EU101338 | KX464878 |
| Do. rosulata | CBS 121762 = CMW 25395 = CAMS 1150 | Acacia mellifera | South Africa | EU101319 | EU101364 | KX464879 |
| Do. rosulata | CBS 500.72 | Medicago sativa, stubble | South Africa | EU673318 | EU673285 | EU673118 |
| Do. santali | WAC 13155 = MUCC 509, ex-type | Santalum acuminatum, healthy stem | Australia: Western Australia | EF591924 | EF591975 | EF591958 |
| Do. sarmentorum | IMI 63581b, ex-type of Bot. sarmentorum | Ulmus sp. | UK: England | AY573212 | AY573235 | EU673102 |
| Do. sempervirentis | IRAN 1581C = CBS 124719 | Cupressus sempervirens | Iran | KC898237 | KC898220 | KX464885 |
| Do. sempervirentis | IRAN 1583C = CBS 124718 = CJA 264, ex-type | Cupressus sempervirens, twigs | Iran | KC898236 | KC898219 | KX464884 |
| Do. striata | CBS 124730 = ICMP 16819 | Citrus sinensis, twigs | New Zealand | EU673320 | EU673287 | EU673142 |
| Do. striata | CBS 124731 = ICMP 16824, ex-type | Citrus sinensis, twigs | New Zealand | EU673321 | EU673288 | EU673143 |
| Do. thailandica | CBS 133991 = CPC 21557 = MFLUCC 11-0438, ex-type of Auerswaldia dothiorella | Dead bamboo culm | Thailand | JX646796 | JX646861 | JX646844 |
| Do. thripsita | CBS 125445 = BRIP 51876a, ex-type | Acacia harpophylla, dead branches, petioles & leaves | Australia: Queensland | KJ573642 | KJ573639 | KJ577550 |
| Do. uruguayensis | CBS 124908 = CMW 26763 = UY672, ex-type | Hexachlamis edulis | Uruguay | EU080923 | EU863180 | KX464886 |
| Do. vidmadera | CBS 621.74 | Pyrus communis, leaf | Switzerland | KX464129 | KX464621 | KX464887 |
| Do. vidmadera | CBS 725.79 | Pyrus malus, dead wood and buds | Switzerland | KX464130 | KX464622 | KX464888 |
| Do. vinea-gemmae | DAR 81012, ex-type | Vitis vinifera cv. Chardonnay, dormant buds | Australia: New South Wales | KJ573644 | KJ573641 | KJ577552 |
| Do. viticola | CBS 117009, ex-type | Vitis vinifera cv. Garnatxa negra, pruned canes | Spain | AY905554 | AY905559 | EU673104 |
| Do. viticola | DAR 80529, ex-type of D. westralis | Vitis vinifera cv. Cabernet Sauvignon, discarded canes | Australia: Western Australia | HM009376 | HM800511 | HM800519 |
| Do. viticola | CPC 26174 | Citrus sinensis, twig | Italy | MW413858 | MW419176 | MW419239 |
| Do. viticola | CPC 26917 | Citrus sinensis, branch | Greece | MW413859 | MW419177 | MW419240 |
| Do. viticola | CPC 27081 | Citrus sinensis, twig | Italy | MW413860 | MW419178 | MW419241 |
| Do. viticola | CPC 27106 | Citrus aurantium, twig | Spain | MW413861 | MW419179 | MW419242 |
| Do. viticola | CPC 27123 | Citrus sinensis, branch | Italy | MW413862 | MW419180 | MW419243 |
| Do. viticola | CPC 27125 | Citrus sinensis, branch | Italy | MW413863 | MW419181 | MW419244 |
| Do. viticola | CPC 27703 | Citrus sinensis, branch | Spain | MW413864 | MW419182 | MW419245 |
| Do. viticola | CPC 27707 | Citrus sinensis, branch | Greece | MW413865 | MW419183 | MW419246 |
| Do. viticola | CPC 27968 | Citrus sinensis, twig | Portugal | MW413866 | MW419184 | MW419247 |
| Do. yunnana | CGMCC 3-17999, ex-type | Camellia sp. | China | KX499643 | KX499649 | – |
| Do. yunnana | CGMCC 3-18000 | Camellia sp. | China | KX499644 | KX499650 | – |
| Dothiorella sp. | CBS 121783 = CMW 25432 = CAMS 1187 | Acacia mearnsii | South Africa | EU101333 | EU101378 | KX464859 |
| Dothiorella sp. | CBS 121784 = CMW 25430 = CAMS 1185 | Acacia mearnsii | South Africa | EU101331 | EU101376 | KX464860 |
| Dothiorella sp. | CBS 121785 = CMW 25433 = CAMS 1188 | Acacia mearnsii | South Africa | EU101334 | EU101379 | KX464861 |
| ‘Lasiodiplodia americana’ | CERC 1961 = CFCC 50065, ex-type | Pistacia vera cv. Kerman, twigs | USA: Arizona | KP217059 | KP217067 | KP217075 |
| L. avicenniae | CMW 41467 = CBS 139670, ex-type | Avicennia marina | South Africa | KP860835 | KP860680 | KP860758 |
| L. brasiliense | CMM 4015 = URM 7118, ex-type | Mangifera indica, stems | Brazil | JX464063 | JX464049 | – |
| L. bruguierae | CMW 41470 = CBS 139669, ex-type | Bruguiera gymnorrhiza | South Africa | NR_147358 | KP860678 | KP860756 |
| L. citricola | CBS 124707 = IRAN 1522C = CJA 72, ex-type | Citrus sp., twigs | Iran | GU945354 | GU945340 | KP872405 |
| L. crassispora | CBS 118741 = WAC 12533 = CMW 14691, ex-type | Santalum album | Australia: Western Australia | DQ103550 | EU673303 | EU673133 |
| L. crassispora | CBS 121770 = CMW 25414 = CAMS 1169, ex-type of L. pyriformis | Acacia mellifera | Namibia | EU101307 | EU101352 | – |
| L. endophytica | MFLUCC 18-1121 = KUMCC 17-0233, ex-type | Magnolia candolii, fresh leaves | China | MK501838 | MK584572 | MK550606 |
| L. egyptiacae | CBS 130992 = BOT-10, ex-type | Mangifera indica, leaf | Egypt | JN814397 | JN814424 | – |
| L. euphorbicola | CMM 3609, ex-type of L. euphorbicola | Jatropha curcas, collar and root rot | Brazil | KF234543 | KF226689 | KF254926 |
| L. gilanensis | CBS 124704 = IRAN 1523C, ex-type | Citrus sp., fallen twigs | Iran | GU945351 | GU945342 | KP872411 |
| L. gilanensis | CBS 128311 = UCD 2193MO, ex-type of L. missouriana | Wedge-shape canker of grapevine cv. Catawba (complex hybrid of North America Vitis species and Vitis vinifera) | USA: Missouri | HQ288225 | HQ288267 | – |
| L. gonubiensis | CBS 115812 = CMW 14077, ex-type | Syzygium cordatum, twigs and leaves | South Africa | AY639595 | DQ103566 | DQ458860 |
| L. gravistriata | CMM 4564, ex-type | Anacardium humile | Brazil | KT250949 | KT250950 | – |
| L. hormozganensis | CBS 124709 = IRAN 1500C, ex-type | Olea sp., twigs | Iran | GU945355 | GU945343 | KP872413 |
| L. iraniensis | CBS 124710 = IRAN 1520C, ex-type | Salvadora persica, twigs | Iran | GU945346 | GU945334 | KP872415 |
| L. iraniensis | CMM 3610, ex-type of L. jatrophicola | Jatropha curcas, collar and root rot | Brazil | KF234544 | KF226690 | KF254927 |
| L. laeliocattleyae | CBS 167.28, ex-type of Diplodia laeliocattleyae | Laeliocattleya | Italy | KU507487 | KU507454 | – |
| L. lignicola | MFLUCC 11-0435 = CBS 134112, ex-type | On dead wood | Thailand | JX646797 | KU887003 | JX646845 |
| L. lignicola | CBS 342.78, ex-type of L. sterculiae | Sterculia oblonga | Germany | KX464140 | KX464634 | KX464908 |
| L. macrospora | CMM 3833, ex-type | Jatropha curcas, collar and root rot | Brazil | KF234557 | KF226718 | KF254941 |
| ‘L. magnoliae’ | MFLUCC 18-0948 = KUMCC 17-0198, ex-type | Magnolia candolii, dead leaves | China | MK499387 | MK568537 | MK521587 |
| L. mahajangana | CBS 124927 = CMW27801, ex-type | Terminalia catappa, healthy branches | Madagascar | FJ900595 | FJ900641 | FJ900630 |
| L. mahajangana | CMM 1325, ex-type of L. caatinguensis | Citrus sinensis | Brazil | KT154760 | KT008006 | KT154767 |
| L. mahajangana | CBS 137785 = BL104, ex-type of L. exigua | Retama raetam, branch canker | Tunisia | KJ638317 | KJ638336 | – |
| L. margaritacea | CBS 122519 = CMW 26162 = MOZ 11A, ex-type | Adansonia gibbosa | Australia: Western Australia | EU144050 | EU144065 | KX464903 |
| L. mediterranea | CBS 137783 = BL1, ex-type | Quercus ilex, branch canker | Italy | KJ638312 | KJ638331 | – |
| L. mitidjana | MUM 19.90 = ALG111, ex-type | Citrus sinensis, branch canker | Algeria: Mitidja | MN104115 | MN159114 | – |
| L. parva | CBS 456.78, ex-type | Cassava-field soil | Colombia | EF622083 | EF622063 | KP872419 |
| L. plurivora | CBS 120832 = CPC 5803, ex-type | Prunus salicina, wood canker | South Africa | EF445362 | EF445395 | KP872421 |
| L. pontae | CMM 1277, ex-type | Spondias purpurea | Brazil | KT151794 | KT151791 | KT151797 |
| L. pseudotheobromae | CBS 116459, ex-type | Gmelina arborea | Costa Rica | EF622077 | EF622057 | EU673111 |
| L. rubropurpurea | CBS 118740 = WAC 12535 = CMW 14700, ex-type | Eucalyptus grandis, canker | Australia | DQ103553 | EU673304 | EU673136 |
| L. subglobosa | CMM 3872, ex-type | Jatropha curcas, collar and root rot | Brazil | KF234558 | KF226721 | KF254942 |
| L. thailandica | CBS 138760 = CPC 22795, ex-type | Mangifera indica, twigs | Thailand | KJ193637 | KJ193681 | – |
| L. theobromae | CBS 111530 = CPC 2095 = JT 695 | Leucospermum sp. | USA: Hawaii | EF622074 | EF622054 | – |
| L. theobromae | CPC 27881 | Citrus sinensis, trunk | Malta | MW413867 | MW419185 | MW419248 |
| L. theobromae | CPC 27882 | Citrus sinensis, trunk | Malta | MW413868 | MW419186 | MW419249 |
| L. theobromae | CPC 27883 | Citrus sinensis, trunk | Malta | MW413869 | MW419187 | MW419250 |
| L. theobromae | CPC 27884 | Citrus sinensis, trunk | Malta | MW413870 | MW419188 | MW419251 |
| L. theobromae | CPC 27885 | Citrus sinensis, trunk | Malta | MW413871 | MW419189 | MW419252 |
| L. theobromae | CBS 124.13 | – | USA | DQ458890 | DQ458875 | DQ458858 |
| L. theobromae | CBS 164.96, ex-neotype | Fruit along coral reef coast | Papua New Guinea | AY640255 | AY640258 | EU673110 |
| L. venezuelensis | CBS 118739 = WAC 12539 = CMW 13511, ex-type | Acacia mangium, wood | Venezuela | DQ103547 | EU673305 | EU673129 |
| L. viticola | CBS 128313 = UCD 2553AR, ex-type | Wedge-shape canker of grapevine cv. Vignoles (complex hybrid of North America Vitis species and Vitis vinifera) | USA: Arkansas | HQ288227 | HQ288269 | HQ288306 |
| L. vitis | CBS 124060, ex-type | Vitis vinifera | KX464148 | KX464642 | KX464917 | |
| Neofusicoccum arbuti | CBS 117453 = CMW 13455, ex-type of N. andinum | Eucalyptus sp. | Venezuela | AY693976 | AY693977 | KX464923 |
| N. arbuti | CBS 116131 = AR 4014, ex-type | Arbutus menziesii, canker | USA: Washington | AY819720 | KF531792 | KF531793 |
| N. australe | CBS 139662 = CMW 6837, ex-type | Acacia sp. | Australia: Victoria | AY339262 | AY339270 | AY339254 |
| N. australe | CMW 6853 | Sequoiadendron | Australia | AY339263 | AY339271 | AY339255 |
| N. brasiliense | CMM 1338, ex-type | Mangifera indica | Brazil | JX513630 | JX513610 | KC794030 |
| N. buxi | CBS 116.75 | Buxus sempervirens, leaf | France | KX464165 | KX464678 | – |
| N. cordaticola | CBS 123634 = CMW 13992, ex-type | Syzygium cordatum | South Africa | EU821898 | EU821868 | EU821838 |
| N. cryptoaustrale | CBS 122813 = CMW 23785, ex-type | Eucalyptus sp., living branches and leaves | South Africa | FJ752742 | FJ752713 | FJ752756 |
| N. dianense | CSF6075 = CGMCC3.20082, ex-type | Eucalyptus urophylla × E. grandis tree, twigs | China | MT028605 | MT028771 | MT028937 |
| N. eucalypticola | CBS 115679 = CMW 6539, ex-type | Eucalyptus grandis | Australia | AY615141 | AY615133 | AY615125 |
| N. eucalyptorum | CBS 115791 = CMW 10125 = BOT 24 | Eucalyptus grandis | South Africa | AF283686 | AY236891 | AY236920 |
| N. grevilleae | CBS 129518, ex-type | Grevillea aurea | Australia | JF951137 | – | – |
| N. hellenicum | CERC 1947 = CFCC 50067, ex-type | Pistacia vera cultivar Aegina | Greece | KP217053 | KP217061 | KP217069 |
| N. hongkongense | CERC2973 = CGMCC3.18749, ex-type | Araucaria cunninghamii | China | KX278052 | KX278157 | KX278261 |
| N. illicii | CGMCC3.18310, ex-type | Illicium verum | China | KY350149 | – | KY350155 |
| N. kwambonambiense | CBS 123639 = CMW 14023, ex-type | Syzygium cordatum | South Africa | EU821900 | EU821870 | EU821840 |
| N. lumnitzerae | CBS 139674 = CMW 41469, ex-type | Lumnitzera racemosa | South Africa | KP860881 | KP860724 | KP860801 |
| N. luteum | CPC 27961 | Citrus limon, twig | Portugal | MW413872 | MW419190 | MW419253 |
| N. luteum | CPC 27962 | Citrus limon, twig | Portugal | MW413873 | MW419191 | MW419254 |
| N. luteum | CBS 110497 = CPC 4594 = CAP 037 | Vitis vinifera, grape | Portugal | EU673311 | EU673277 | EU673092 |
| N. luteum | CBS 110299 = LM 926 = CAP 002, ex-type | Vitis vinifera, cane | Portugal | AY259091 | KX464688 | DQ458848 |
| N. luteum | CBS 140738 = CMW 41365, ex-type of N. mangroviorum | Avicennia marina | South Africa | KP860859 | KP860702 | KP860779 |
| N. macroclavatum | CBS 118223 = CMW 15955 = WAC 12444, ex-type | Eucalyptus globulus, wood | Australia: Western Australia | DQ093196 | DQ093217 | DQ093206 |
| N. magniconidium | CSF5876 = CGMCC3.20077, ex-type | Eucalyptus urophylla × E. grandis tree, twigs | China | MT028612 | MT028778 | MT028944 |
| N. mangiferae | CBS 118531 = CMW 7024 | Mangifera indica | Australia | AY615185 | DQ093221 | AY615173 |
| N. mediterraneum | CBS 121718 = CPC 13137, ex-type | Eucalyptus sp., branches and leaves | Greece | GU251176 | – | – |
| N. mediterraneum | CBS 113083 = CPC 5263, ex-type of N. pistaciarum | Pistacia vera | USA: California | KX464186 | KX464712 | KX464998 |
| N. mediterraneum | CBS 113089 = CPC 5274, ex-type of N. pistaciicola | Pistacia vera | USA: California | KX464199 | KX464727 | KX465014 |
| N. mediterraneum | CPC 27931 | Citrus limon, twig | Portugal | MW413874 | MW419192 | MW419255 |
| N. mediterraneum | CPC 27932 | Citrus limon, twig | Portugal | MW413875 | MW419193 | MW419256 |
| N. mediterraneum | CPC 27935 | Citrus limon, twig | Portugal | MW413876 | MW419194 | MW419257 |
| N. mediterraneum | CPC 27936 | Citrus limon, twig | Portugal | MW413877 | MW419195 | MW419258 |
| N. microconidium | CERC3497 = CGMCC3.18750, ex-type | Eucalyptus urophylla × E. grandis tree | China | KX278053 | KX278158 | KX278262 |
| N. nonquaesitum | CBS 126655 = L3IE1 = PD484, ex-type | Umbellularia californica, cankered branch | USA: California | GU251163 | GU251295 | GU251823 |
| N. ningerense | CSF6028 = CGMCC3.20078, ex-type | Eucalyptus urophylla × E. grandis tree, twigs | China | MT028613 | MT028779 | MT028945 |
| N. occulatum | CBS 128008 = MUCC 227, ex-type | Eucalyptus grandis hybrid | Australia: Queensland | EU301030 | EU339509 | EU339472 |
| N. pandanicola | MFLUCC 17-2270 = KUMCC 17-0184, ex-type | Pandanus sp. | China | MH275072 | – | – |
| N. parviconidium | CSF5667 = CGMCC3.20074, ex-type | Eucalyptus tree, twigs | China | MT028615 | MT028781 | MT028947 |
| N. parvum | CBS 138823 = ICMP 8003 = CMW 9081 = BOT2487 = ATCC 58191, ex-type | Populus nigra, bark of dead twig | New Zealand | AY236943 | AY236888 | AY236917 |
| N. parvum | CPC 26119 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413878 | MW419196 | MW419259 |
| N. parvum | CPC 26120 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413879 | MW419197 | MW419260 |
| N. parvum | CPC 26121 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413880 | MW419198 | MW419261 |
| N. parvum | CPC 26122 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413881 | MW419199 | MW419262 |
| N. parvum | CPC 26124 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413882 | MW419200 | MW419263 |
| N. parvum | CPC 26126 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413883 | MW419201 | MW419264 |
| N. parvum | CPC 26127 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413884 | MW419202 | MW419265 |
| N. parvum | CPC 26128 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413885 | MW419203 | MW419266 |
| N. parvum | CPC 26129 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413886 | MW419204 | MW419267 |
| N. parvum | CPC 26130 | Citrus sinensisx Poncirus trifoliata, trunk | Italy | MW413887 | MW419205 | MW419268 |
| N. parvum | CPC 27866 | Citrus limon, branch | Malta | MW413888 | MW419206 | MW419269 |
| N. parvum | CPC 27867 | Citrus limon, branch | Malta | MW413889 | MW419207 | MW419270 |
| N. parvum | CPC 27868 | Citrus limon, branch | Malta | MW413890 | MW419208 | MW419271 |
| N. parvum | CPC 28173 | Microcitrus australasica, twig | Italy | MW413891 | MW419209 | MW419272 |
| N. parvum | CPC 28175 | Microcitrus australasica, twig | Italy | MW413892 | MW419210 | MW419273 |
| N. parvum | CPC 28177 | Microcitrus australasica, twig | Italy | MW413893 | MW419211 | MW419274 |
| N. parvum | CBS 110301 = CAP 074 | Vitis vinifera | Portugal | AY259098 | AY573221 | EU673095 |
| N. parvum | MFLUCC 15-09002, ex-type of N. italicum | Vitis vinifera | Italy | KY856755 | KY856754 | – |
| N. parvum | CBS 137504 = ALG1, ex-type of N. algeriense | Vitis vinifera, branches | Algeria | KJ657702 | KJ657721 | – |
| N. pennatisporum | WAC 13153 = MUCC 510, ex-type | Allocasuarina fraseriana, healthy stem | Australia: Western Australia | EF591925 | EF591976 | EF591959 |
| N. pistaciae | CBS 595.76, ex-isotype of Camarosporium pistaciae | Pistacia vera, fruits | Greece | KX464163 | KX464676 | KX464953 |
| N. protearum | CBS 114176 = CPC 1775 = JT 189, ex-type | Leucadendron salignum × L. laureolum cv. Silvan Red, stems | South Africa | AF452539 | KX464720 | KX465006 |
| N. ribis | CBS 115475 = CMW 7772, ex-type | Ribes vulgare | USA | AY236935 | AY236877 | AY236906 |
| N. ribis | CBS 124924 = CMW 28363, ex-type of N. batangarum | Terminalia catappa, healthy branches | Cameroon | FJ900607 | FJ900653 | FJ900634 |
| N. ribis | CBS 123645 = CMW 14058, ex-type of N. umdonicola | Syzygium cordatum | South Africa | EU821904 | EU821874 | EU821844 |
| N. sinense | CGMCC3.18315, ex-type | Unknown woody plant | China | KY350148 | KY817755 | KY350154 |
| N. sinoeucalypti | CERC2005 = CGMCC3.18752, ex-type | Eucalyptus urophylla × E. grandis tree | China | KX278061 | KX278166 | KX278270 |
| N. stellenboschiana | CBS 110864 = STE-U 4598 = CPC 4598, ex-type | Vitis vinifera | South Africa | AY343407 | AY343348 | KX465047 |
| N. terminaliae | CBS 125264 = CMW 26683 | Terminalia sericea | South Africa | GQ471804 | GQ471782 | KX465053 |
| N. ursorum | CBS 122811 = CMW 24480, ex-type | Eucalyptus sp. | South Africa | FJ752746 | FJ752709 | KX465056 |
| N. variabile | CMW 37739, ex-type | Mimusops caffra | South Africa | MH558608 | – | MH569153 |
| N. viticlavatum | CBS 112878 = CPC 5044 = JM 86, ex-type | Vitis vinifera | South Africa | AY343381 | AY343342 | KX465058 |
| N. vitifusiforme | CBS 110887 = CPC 5252 = JM5, ex-type | Vitis vinifera | South Africa | AY343383 | AY343343 | KX465061 |
| N. vitifusiforme | CBS 120081 = CPC 12925, ex-type of N. corticosae | Eucalyptus corticosa, leaves | Australia: New South Wales | DQ923533 | KX464682 | KX464958 |
| N. vitifusiforme | CBS 121112 = CPC 5912, ex-type of N. pruni | Prunus salicina | South Africa | EF445349 | EF445391 | KX465016 |
| N. yunnanense | CSF6142 = CGMCC3.20083, ex-type | Eucalyptus globulus, twigs | China | MT028667 | MT028833 | MT028999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra, J.D.P.; Crous, P.W.; Aiello, D.; Gullino, M.L.; Polizzi, G.; Guarnaccia, V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants 2021, 10, 492. https://doi.org/10.3390/plants10030492
Bezerra JDP, Crous PW, Aiello D, Gullino ML, Polizzi G, Guarnaccia V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants. 2021; 10(3):492. https://doi.org/10.3390/plants10030492
Chicago/Turabian StyleBezerra, Jadson Diogo Pereira, Pedro Wilhelm Crous, Dalia Aiello, Maria Lodovica Gullino, Giancarlo Polizzi, and Vladimiro Guarnaccia. 2021. "Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe" Plants 10, no. 3: 492. https://doi.org/10.3390/plants10030492