
Elevated CO2 and Reactive Oxygen Species in Stomatal Closure



State Key Laboratory of Crop Stress Adaptation and Improvement, School of Life Sciences, Henan University, Kaifeng 475004, China
*
Author to whom correspondence should be addressed.
Plants 2021, 10(2), 410; https://doi.org/10.3390/plants10020410
Submission received: 26 January 2021
/
Revised: 10 February 2021
/
Accepted: 16 February 2021
/
Published: 23 February 2021
(This article belongs to the Special Issue Specificity of ROS and Redox Regulatory Systems)
Abstract
:Plant guard cell is essential for photosynthesis and transpiration. The aperture of stomata is sensitive to various environment factors. Carbon dioxide (CO2) is an important regulator of stomatal movement, and its signaling includes the perception, transduction and gene expression. The intersections with many other signal transduction pathways make the regulation of CO2 more complex. High levels of CO2 trigger stomata closure, and reactive oxygen species (ROS) as the key component has been demonstrated function in this regulation. Additional research is required to understand the underlying molecular mechanisms, especially for the detailed signal factors related with ROS in this response. This review focuses on Arabidopsis stomatal closure induced by high-level CO2, and summarizes current knowledge of the role of ROS involved in this process.

{kind=link}
{kind=link}
1. Introduction
Stomata, the tiny pores formed by pairs of guard cells on the surfaces of plant leaves, are the main portals for gas exchange, water transpiration and pathogen invasion in plants. Plants adapt to changes in the external environment by regulating the aperture of stomata. The movement of guard cells is regulated by a variety of environmental factors, including water status, light, carbon dioxide (CO2) concentration and pathogen attack, as well as internal signals such as phytohormones, calcium and reactive oxygen species [1,2].
CO2, as an important regulator of stomatal movement and development, greatly affects the growth and biomass of plants. With the development of modern industry and the increase in human activities, the CO2 content in the atmosphere is increasing year by year. In January 2021, data from the Mauna Loa Observatory show that the atmospheric background CO2 content will exceed 417 parts per million (ppm) (https://www.metoffice.gov.uk/research/climate/seasonal-to-decadal/long-range/forecasts/co2-forecast (accessed on 22 February 2021)). The continuous rise of CO2 in the atmosphere resulted in exponential increasing of intercellular CO2 levels in leaf cells [3,4,5]. The profound effect of CO2 on stomatal development has been confirmed using plant fossils with CO2 [6,7]. In addition to changes in development, the intercellular CO2 concentration also influences stomatal responses [8,9]. Although CO2 signaling is well established to regulate stomata movement, its molecular mechanisms are not fully understood.
In stomata, the CO2 signaling pathway is complex. It is generally thought that during stomatal closure induced by elevated CO2 concentration, CO2 is first converted into bicarbonate (HCO3−) by beta Carbonic Anhydrase 1 (βCA1) and beta Carbonic Anhydrase 4 (βCA4) [10]. HCO3− is perceived and transported by the antimicrobial extrusion (MATE) type transporter RESISTANCE TO HIGH CO2 (RHC1) [11]. Subsequently, Mitogen-Activate Protein Kinase 4 (MPK4) and MPK12 inhibit HIGH LEAF TEMPERATURE 1 (HT1), which then activates OPEN STOMATA 1 (OST1). Activation of OST1 facilitates the S-type anion channel to mediate anion effluxes, resulting in stomatal closure [10,11,12,13,14,15,16,17].
Reactive oxygen species (ROS) are major regulators of stomatal movement, particularly in response to abiotic and biotic stress [18,19]. ROS, particularly hydrogen peroxide (H2O2) and superoxide (O2−), are widely produced within different cellular compartments of plants. The evolution and maintenance of different sources for ROS production is most likely due to the requirement for intricate control of oxidative signaling, given that ROS can be cytotoxic and mutagenic and, for their proper function in signaling, their production must be tightly regulated both temporally and spatially [20]. Recent progress for the high-level CO2-induced stomatal closure showed that ROS function as the key factor [21]. However, components and mechanism underlying this ROS mediated CO2 signaling pathway are still not so clear, with only a few factors have been disclosed. This review summarizes on the process of high-level CO2-induced closure in Arabidopsis and focuses more on current knowledge related with ROS signal. Understanding of how ROS signal have been engaged in high-level CO2-induced stomatal movement is essential for CO2 signaling, which will demand extensive research.
2. Importance of CO2 Regulation of Stomatal Conductance
Under dark conditions, respiration leads to a rapid increase in intercellular CO2 concentration ([CO2]cys) in plant leaves, exceeding 600 ppm [22]. However, under light conditions, [CO2]cys drops below 200 ppm [22]. High environmental CO2 concentration induces stomatal closure, whereas low environmental CO2 concentration triggers stomatal opening. Despite its importance, little is known about the molecular mechanisms underlying the initial response of stomata to CO2.
CO2 negatively affects stomatal conductance, or the rate of passage of materials through stomata, by reducing both the number of stomata per unit of leaf area and the stomatal apertures. Decreases in stomatal conductance lead to decreases in transpiration, reducing the loss of water from leaves, which is beneficial to plant water conservation [23]. In the case of relatively high CO2 levels, fewer stomata and smaller apertures reduce the heat evapotranspirative cooling capacity of leaves, which aggravates heat stress under water shortage [24,25]. Notably, heat stress greatly reduces plant resistance and crop yields globally, affecting agricultural production and possibly nutrient content [26,27]. By contrast, greater stomatal conductance improves crop yield [28,29,30]. Therefore, under conditions of sufficient water, higher CO2 concentrations reduce stomatal conductance, which may contribute to suboptimal yields [25].
In some species, CO2 concentration also affects leaf development and leaf area. These responses directly affect leaf water conservation capacity and plant biomass [25]. Future research is needed to explore the molecular mechanism by which CO2 regulates plant gas exchange.
3. Mechanism of High CO2-Induced Stomatal Closure
Although our understanding of plant responses to environmental CO2 has advanced, the mechanisms underlying CO2 signal perception and transduction are still not clear. After entering the cell, CO2 is sensed by receptors on the plasma membrane. In animals, CO2 perception is believed to be a vestigial sensory mechanism [31] associated with carbonic anhydrase [32]. Adenylyl cyclase enzymes play important roles in CO2 sensing in mammalian and fungal systems [31]. Plant homologs of α- or γ- carbonic anhydrases have not been identified, while β-carbonic anhydrases have been characterized [10,33]. β-carbonic anhydrases have been proposed to carry out the CO2 receptor function in plants [31], which is important in the stomatal movement response to changes in CO2 concentration in Arabidopsis. Of the six Arabidopsis thaliana β-carbonic anhydrases, βCA4 is localized at the plasma membrane, while βCA1 is mainly targeted to chloroplasts when transiently expressed in Nicotiana benthamiana cells [10,34]. βCA1 and βCA4 convert CO2 into HCO3−, and both function in CO2-induced stomatal movement. The expression of either βCA4 at the plasma membrane or βCA1 in chloroplasts in guard cells of ca1 ca4 double-mutant plants restored CO2-induced stomatal closure [35]. The expression of animal carbonic anhydrase in the double mutant (ca1 ca4) similarly restored CO2-induced stomatal closure [10,35]. These data suggest that the catalytic activity of carbonic anhydrase is involved in the signaling pathway of CO2 stimulation of closure, and HCO3− and/or H+ may act as second messengers in this signal transduction in plants [10]. The apparent conservation of carbonic anhydrase activity between plants and animals implies that they sense CO2 by similar mechanisms.
Protein HT1 has been characterized as an important factor in CO2 signaling. As the first CO2-response mutant, ht1 has been identified by thermal imaging, which has higher leaf temperature and reduced stomatal CO2 response [12]. HT1 encodes a putative MAPKKK kinase that negatively regulates stomatal responses to changes in CO2 levels, but does not affect abscisic acid (ABA) signaling [12], suggesting that there is an ABA-independent signaling pathway in CO2-regulated stomatal response. ca1 ca4 ht1-2 triple mutants exhibit a similar CO2 response as ht1-2, suggesting that HT1 acts downstream of βCA1 and βCA4 in CO2-induced stomatal movement [10]. βCA4 interacts with PIP2;1, which might transport apoplastic CO2 [36]. The OST1 protein kinase is required for CO2-induced stomatal closing [37]. A recent study proposed that HT1 deactivates OST1 [11]. However, another study found HT1 to be epistatic to OST1 in a high-CO2-induced stomatal closing assay [38]. Since the function of OST1 in stomatal closure is mainly through regulating ion channels, the research of anion channels in response to high levels of CO2 is helpful for establishing a comprehensive signal pathway.
The role of HCO3− in regulating stomatal movement has been studied using patch clamp experiments. Stomatal closure requires the activation of anion channels in guard cells. Increasing the concentration of HCO3− in the cytoplasm activates anion channels [37]. The MATE transporter protein RHC1 is a receptor for HCO3− [11]. Increased CO2 concentration enhances anion channel activity in guard cells [10]. S-type anion channels in the plasma membrane of guard cells might provide a central control mechanism for stomatal closing [39]. The S-type anion channel SLAC1 plays a crucial role in stomatal movement during signaling [40]. SLAC1 is activated by elevated intracellular HCO3− levels in guard cells [37]. Nevertheless, the N terminus is important for ABA signaling, and the SLAC1 transmembrane domain responds to CO2 but not the N terminus or C terminus [41]. Recent studies have shown that residue R256 of SLAC1 is required for elevated CO2 induced stomatal closure, but not for ABA-induced stomatal closing [42]. Both OST1 and GUARD CELL HYDROGEN PEROXIDE-RESISTANT1 (GHR1) activate SLAC1 in oocytes [43,44]; this activation is inhibited by HT1 [14]. MPK12 counteracts this inhibition [14]. In a recent natural variation study, MPK12 was also identified as a key player in the Arabidopsis Cvi-0 accession for guard cell CO2 signaling [15]. These findings suggest that MPK12 inhibits HT1, and HT1 inhibits OST/GHR1-activated SLAC1. Unlike HT1, which mainly functions in CO2 signaling, MPK12 is also activated by ABA or H2O2 [45]. Co-expressing βCA4 and PIP2;1 with OST1-SLAC1 or CPK6/23-SLAC1 in oocytes activates SLAC1 via extracellular CO2 [36]. These findings suggest that in the presence of various protein kinases, SLAC1 can be regulated by cytosolic CO2/HCO3− that has been transported by PIP2;1 and converted by βCA4 in oocytes. Till now, a simple regulation pathway for CO2-induced stomatal closure has been disclosed.
Since stomatal movement is always associated with oscillations of cytosolic [Ca2+]cyt [46], research on the stomata response to high levels of CO2 [47] has also identified the involvement of calcium ions in Commelina communis [48,49] and Arabidopsis thaliana [2,50]. This conclusion is based on experiments in which Ca2+ accumulated in guard cells subjected to elevated CO2 concentration, and high CO2-induced stomatal closure was impaired in the presence of Ca2+ chelators, such as 1,2-Bis(2-Aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid (BAPTA) or ethylenedinitrilotetraacetic acid (EDTA) [48,49,50]. Exposure to reduced CO2 concentration triggered more [Ca2+]cyt transients in guard cells than did exposure to elevated CO2 concentration [2]. As guard cells produce “spontaneous” cytoplasmic Ca2+ transients, and Ca2+ is required for high CO2-induced stomatal closure, elevated CO2 concentration might enhance the sensitivity of stomatal closing mechanisms to [Ca2+]cyt [51]. In agreement with this hypothesis, CO2-derived bicarbonate enhances the Ca2+ sensitivity of the S-type anion channel activation in guard cells [37]. The increase in [Ca2+]cyt transients in response to higher CO2 levels implicates this second messenger in the complex mechanism underlying CO2-regulated stomatal movement, an idea that may need more extensive research.
Although an ABA-independent pathway has been demonstrated in CO2-regulated stomatal response, ABA signaling components have also been implicated, suggesting that a shared regulatory molecular mechanism may involve ABA and CO2 in stomatal movement. At both low and high CO2 concentrations, the [Ca2+]cyt of the ABA-insensitive mutant gca2 changes at a similar rate, and the mutant exhibits strongly attenuated stomatal closure in response to increased CO2 in leaves [2]. The gca2 mutant also exhibits an altered ABA-induced [Ca2+]cyt pattern in guard cells [46], suggesting that Ca2+ alteration mediated by GCA2 (GROWTH CONTROLLED BY ABSCISSIC ACID 2) functions downstream or at the convergence point of CO2 and ABA signaling. Therefore, the same factors as in ABA signaling may have also been implicated in CO2-induced stomatal closure, suggesting that the same mechanism exists for stomatal closure, which may be helpful for understanding CO2 signaling.
4. ROS in Stomatal Closure
Stomatal closure is accompanied by increased ROS level in the guard cell apoplast and chloroplasts in response to various treatments [19,52]. In Arabidopsis, apoplastic ROS are mainly produced by plasma membrane-localized NADPH oxidases (RESPIRATORY BURST OXIDASE HOMOLOGS, RBOHs) and cell-wall peroxidases [19,53,54], whose activities are strongly inhibited by diphenylene iodonium (DPI) and salicylhydroxamic acid (SHAM), respectively [55,56,57,58,59].
In Arabidopsis guard cells, there are two main isoforms of NADPH oxidases, AtRBOHF and AtRBOHD, which are regulated by ABA-dependent processes [60]. ABA-triggered stomatal response is significantly reduced in the atrbohF mutant, a phenotype that is enhanced in the atrbohD atrbohF double mutant but absent in the atrbohD single mutant [60]. Due to its obvious role in pathogen-triggered ROS burst, RBOHD is more commonly recognized to function in plant immune defense [61] and might therefore play a role in stomatal movement. Both NADPH oxidases have been demonstrated to function in guard cell CO2 responses by staining atrbohD atrbohF with H2DCFDA, and the CO2-induced ROS burst requires ABA signaling [21], which implies that the two oxidases are essential for both ABA and CO2-induced stomatal response.
Apoplastic ROS are also produced by other oxidases [62]. For example, in Vicia faba and Arabidopsis thaliana, copper amine oxidase and polyamine oxidases are involved in the production of the ROS in the process of ABA- and ethylene-induced stomatal closure [63,64]. The involvement of these oxidases in apoplastic ROS production comes from inhibitor studies, and further research is needed to understand their specific and detailed function and significance in ROS-induced stomatal regulation.
Apoplastic ROS production initiates the activation of plasma membrane Ca2+ channels, leading to an increase in [Ca2+]cyt levels [51]. The molecular identity of these inducible plasma membrane Ca2+ channels is not clear. In the cytosol, Ca2+ stimulates the activation of NADPH oxidases either directly by binding to their cytoplasmic EF hands (the EF hand is a helix-loop-helix structural domain or motif found in a large family of calcium-binding proteins) [65] or indirectly by affecting their phosphorylation by CALCIUM DEPENDENT PROTEIN KINASES (CPKs) [66]. Upon Ca2+ binding, CALCINEURIN-B LIKE PROTEINS (CBLs) interact with CPKs and CBL-interacting PROTEIN KINASES (CIPKs) [66], and a particular complex CBL1/CBL9-CIPK26 is formed, which then phosphorylates and activates RBOHF [67]. The increase in [Ca2+]cyt is also sensed in the chloroplasts, where a thylakoid membrane-associated Ca2+-binding protein, CALCIUM SENSING RECEPTOR (CAS), is activated through an unidentified mechanism [51]. The activation of CAS is responsible for a chloroplast ROS burst and the release of Ca2+ from thylakoids [68,69,70,71], both of which contribute to cytoplasmic Ca2+ oscillations, apoplastic Ca2+-induced stomatal closure during plant immune defense [69]. The drought sensitivity of the Arabidopsis cas mutant is caused by the improper closure of stomata [72], which highlights the importance of chloroplastic Ca2+ signaling in stomatal regulation [51].
Although there is clear evidence for the involvement of ROS in the regulation of stomatal aperture, it is not known how the ROS signals are sensed in the guard cell apoplast. Identification of the ROS and redox sensors is a major challenge in plant ROS research. In guard cells, only a few ROS sensing mechanisms are involved in stomatal regulation: redox regulation of the GHR1 apoplastic domain [44], of OST1 [73] and of CPK1 [74]. GHR1 is proposed to be involved in the perception of apoplastic ROS, a plasma membrane-associated atypical leucine-rich repeat receptor-like protein kinase. The apoplastic C-terminal domain of GHR1 has two conserved cysteines (C-57 and C-66) that are necessary for the correct function of the protein [44]. GHR1, as an inactive pseudokinase that mediates the activation of SLAC1 via interacting with CALCIUM-DEPENDENT PROTEIN KINASE 3 (CDPK3), could potentially act in stomatal closure as a scaffolding component [75]. Therefore, GHR1 is implicated as a central regulator of guard cell CO2 and early ABA responses for stomatal movement by mediating ROS signaling. However, more detailed research is demanded to elucidate the mechanism of ROS in stomatal movement induced by different stimuli.
5. Function of ROS Signaling in CO2-Induced Stomatal Closure
Despite their importance, not much effort has been dedicated to understanding the function of ROS in CO2-regulated stomatal behavior. Treatment with high concentrations of HCO3− induces ROS production and promotes plasma membrane-localized NADPH oxidase-dependent stomatal closure [76], suggesting that ROS play a role in CO2-induced stomatal closure.
Chater et al. concluded that ROS are required for stomatal closure induced by high CO2 concentrations, as no stomatal closure was observed in rbohD rbohF double mutants [21,77], but there was a strong ROS decrease in guard cells at elevated CO2 concentrations. These observations could be explained by a reduction in the oxygenase activity of RuBisCO at high CO2 concentration, and hence a reduction in ROS production by glycolate oxidase activity linked to photorespiration [78]. These findings suggest that ROS generated by NADPH oxidase play an important role in high-CO2-induced decreases in stomatal aperture.
Cell wall-bound SHAM-sensitive peroxidases also take part in apoplastic ROS production around guard cells [57,58]. Two cell-wall peroxidase-encoding genes, PRX33 and PRX34, which are highly and preferentially expressed in guard cells, are also involved in ROS production in CO2-induced stomatal closure [77]. Pharmacological and genetic studies show that ROS generated by both NADPH oxidases and cell-wall peroxidases contribute to high-CO2-induced stomatal closure. The high-CO2-induced reduction in stomatal aperture is efficiently abolished by either the NADHP oxidase inhibitor DPI or the cell-wall peroxidase inhibitor SHAM, suggesting that cell-wall peroxidases function in high-CO2-induced stomatal closure. Similar to rbohD rbohF double mutants, stomatal apertures of both prx33-3 and prx34-2 mutant lines fail to close in response to high CO2. ROS accumulation is not triggered by high CO2 in prx33-3 prx34-2 or rbohD rbohF mutants, in marked contrast to the >50% ROS increase in wild type. These results indicate that in addition to RBOHD and RBOHF, cell-wall peroxidases including PRX33 and PRX34 play an essential role in high-CO2-induced ROS generation [77]. However, it is not certain whether other oxidase proteins also participate in CO2-induced ROS accumulation, and more high-CO2-induced ROS generation related proteins need to be identified.
Proteins that regulate ROS homeostasis in CO2-induced stomatal closure have also been screened. High-CO2-induced ROS production and stomatal closure in big mutants are compromised [77]. The BIG/CIS1 protein is involved in diverse processes, including auxin transport, light and hormonal signaling, vesicle trafficking, endocytosis, phosphate deficiency tolerance, and the dynamic adjustment of circadian period [79,80,81,82,83,84,85,86]. He et al. found that cis1 mutants are compromised in both elevated CO2-induced closure and reduction in stomatal density [87]. cis1 mutants have a significantly lower leaf surface temperature compared with the wild type when exposed for 40 min to 1500 ppm CO2. S-type anion channel activity is disrupted by HCO3− treatment compared with the wild type [87]. This work suggests that the signaling pathway for ROS in CO2-regulated stomatal movement may include other signals, and unraveling the control mechanism will depend on identifying more of these molecules.
6. ROS Are the Nodes of CO2 and ABA Signaling during Stomatal Movement
Since both ABA- and high CO2-induced stomatal closure involve the activation of SLAC1 in guard cells, there may be a convergence between ABA and CO2 signaling. Several mutations causing stomatal ABA insensitivity, such as abi1-1, abi2-1 [88,89], ost1 and ghr1 [14,88], impair stomatal responses to high CO2 concentrations, indicating that there is a relationship between ABA and CO2. Stomata of the gca2 mutant are insensitive to both ABA and high CO2 signaling [2,46]. ABA receptors, PYR/PYL/RCARs, are also involved in CO2 signaling, as the inactivation of several of these proteins impairs stomatal closure in elevated CO2 concentrations [21,88], warranting further identification of the role of these receptors in CO2 signaling.
Although several key components of ABA signaling are connected with stomatal responses to high CO2 concentration, suggesting that an ABA-dependent pathway participates in CO2 signaling, ABA-independent components also exist. ABA-induced stomatal closure is completely functional in the mutants of HT1 and MPK12, whereas these plants are deficient in CO2-induced stomatal movements [14,15]. Moreover, experiments which aimed to dissect which parts of the SLAC1 anion channel are important for ABA- and for high-CO2-induced stomatal closure showed that transgenic plants expressing the SLAC1 anion channel without both C- and N-terminal regions still respond to changes in CO2 concentration, but remain ABA-insensitive. Thus, the ABA-induced activation of SLAC1 appears to involve its C- and N-terminal regions, whereas high-CO2-induced stomatal closure appears to rely only on the transmembrane region [41].
Current knowledge of both ABA- and high-CO2-induced stomatal closure suggests at least three partially overlapping pathways: (1) the direct perception of HCO3− by SLAC1 in the presence of protein kinases that activate anion channel SLAC1 [36]; (2) the ABA-independent pathway; and (3) the ABA-dependent component that partially mediates high-CO2-induced stomatal closure [21,37,88]. ABA signaling that activates OST1 and CPKs by the suppression of PP2Cs could enhance SLAC1 sensitivity to HCO3−, and directly trigger SLAC1 anion currents, although this hypothesis needs to be verified in future research. When the CO2 supply for plants is sufficient, it is possible that plants under water stress react to increased [CO2]cyt faster and stronger than plants with satisfactory water supply, thus saving water in leaves. This could explain the importance of ABA signaling for CO2-induced stomatal movements, which would enable plants to adapt to changing environmental conditions. These reports suggest that ABA and CO2 signaling pathways in stomatal closure are not totally the same, but special factors or regulation systems are recruited.
The overlap of CO2 and ABA signaling also indicates that ROS production in guard cells can increase in response to high CO2 concentration, similar to ABA-induced stomatal closure [51]. We previously showed that ROS accumulate in guard cells treated with HCO3− or high CO2 [21,76,90]. A connection between CO2 and ABA signaling is supported by the absence of ROS accumulation in stomata under elevated CO2 concentration in the ABA-deficient double mutant nced3 nced5, the triple pyr1 pyl1 pyl4 and quadruple pyr1 pyl1 pyl2 pyl4 mutants [21]. Similar to ABA, elevated CO2 induces ROS formation by NADPH oxidases [21,90]. In rbohD rbohF double mutants, guard cells are insensitive to CO2/HCO3−. These mutants also fail to produce ROS in guard cells in response to elevated CO2 [21,76]. The impaired accumulation of ROS in guard cells and decreased stomatal closure in response to elevated CO2 concentration are also observed in the tomato mutant rboh1 [90]. Since ROS-related signal components in ABA-induced stomatal closure have been revealed, the involvement of these ABA factors in ROS signaling in response to CO2 suggests that high-CO2 and ABA signaling may converge at ROS, and similar downstream components may function in both pathways.
7. Concluding Remarks and Future Directions
This review summarizes the molecular mechanisms of the ROS signaling network in plant stomatal movement, primarily under high CO2 concentrations (Figure 1). Elevated [CO2]-mediated stomatal closure requires an increase in ROS. However, great difficulty still exists in understanding the complex interactions within the guard cell signaling networks in response to changes in CO2, which include ROS signaling. Recent research has highlighted the complex interplay between CO2, redox/ROS signaling and phytohormonal regulation in the control of stomatal movements, and as discussed in this review, there is cross-talk between ABA and CO2 signaling by ROS during stomatal closure. The importance of guard cells in photosynthesis and transpiration has attracted considerable effort toward understanding how guard cells perceive and transmit signals from the environment. The identification of proteins that can sense and transmit changes in CO2/HCO3− and molecules that function in regulating ROS signals in guard cells is urgent to improve understanding of the regulation of stomatal movement in response to CO2.
Future breakthroughs will most likely come from the development of new methods and technologies that enable real-time imaging of physiological indexes such as cellular localization of ROS, Ca2+, and pH in response to various stimuli including CO2; obtaining novel genetic mutants to identify proteins involved in CO2 response will also be important [47]. Furthermore, plant altered DNA methylation has been implicated in various environmental stress response [91], increasing evidences demonstrated that multiple epigenetic factors and their interactions with hormones, especially auxin will be another direction for understanding high CO2-induced stomatal closure [92]. The isolation of methylation-related mutants will be a useful tool for exploring molecular mechanisms. Future research should also seek to understand the complex interactions between various guard cell signaling pathways and how guard cells modulate the interactions among hormones, ROS, CO2 and Ca2+ homeostasis. In addition, the translation of such knowledge from model plants to important crop species will be increasingly important for improving stress resistance and increasing yield.
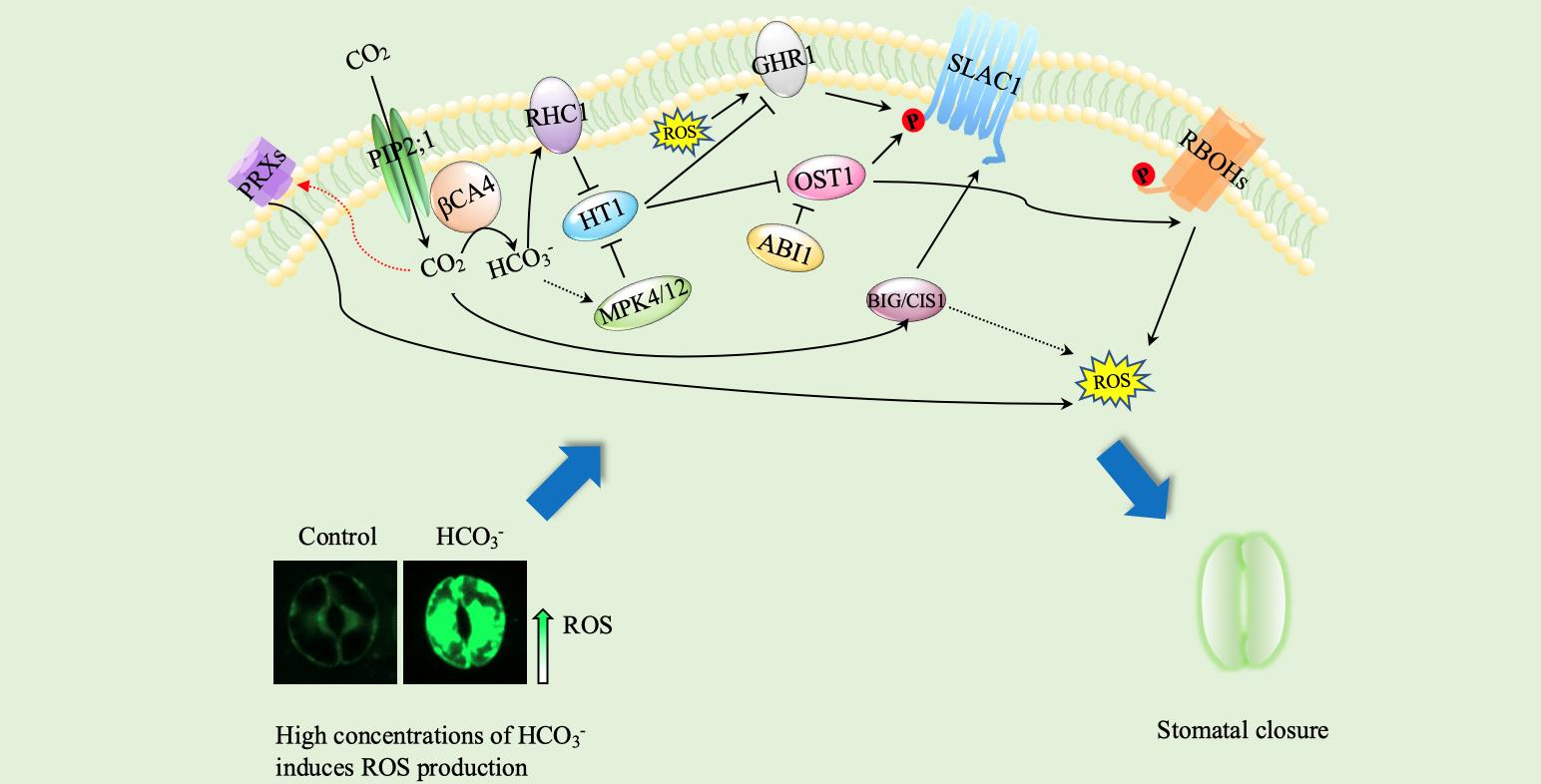
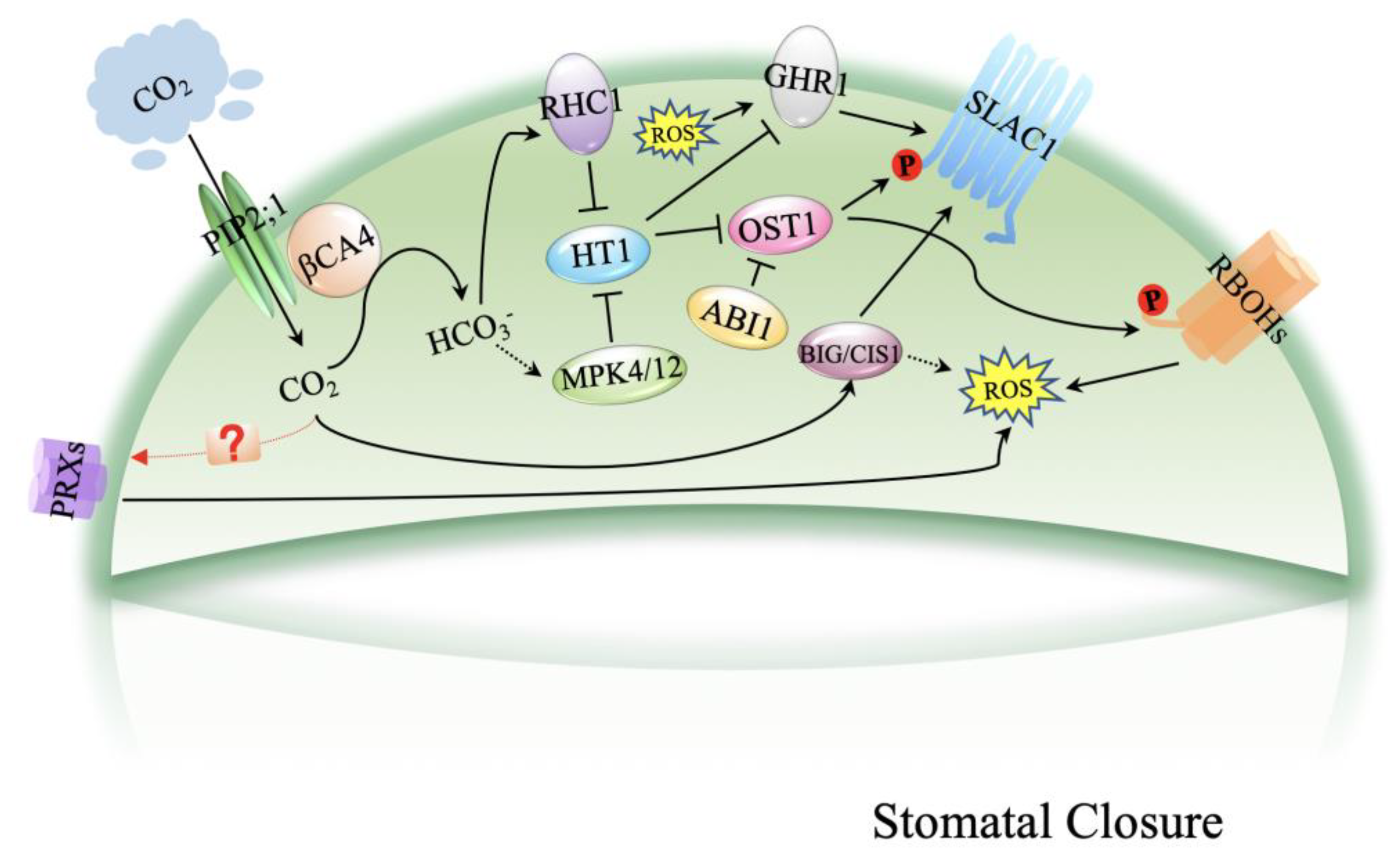
CO2 has been transported by PIP2;1 and converted into HCO3− by βCA4, which is located at the plasma membrane and interacts with PIP2;1. The accumulation of cytosolic HCO3− leads to the suppression of HT1 by MPK4/12 and RHC1. Repressed HT1 inhibits OST1, resulting in the activation of SLAC1. OST1 phosphorylates respiratory burst ROBHs to produce ROS and induces stomatal closure. GHR1 is an inactive pseudokinase that mediates the activation of SLAC1. BIG1 activates SLAC1 when cytosolic HCO3− accumulates and produces ROS that induce stomatal closure.
PRXs, peroxidases; RBOHs, respiratory burst oxidase homologs; PIP2;1, plasma membrane intrinsic protein 2;1; βCA4, beta carbonic anhydrase 4; RHC1, resistance to high CO2;HT1, high leaf temperature 1; MPK4/12, protein kinase 4/12; OST1, open stomata 1; ABI1, ABA insensitive 1; BIG1, CO2 insensitive 1 GHR1, guard cell hydrogen peroxide-resistant 1; SLAC1, slow anion channel-associated 1; ROS, reactive oxygen species; P, phosphorylation.
Author Contributions
X.M. and L.B. co-wrote this review. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31900239 and 31970198) and the Henan Science and Technology Development Program (192102110115).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schroeder, J.I.; Kwak, J.M.; Allen, G.J. Guard cell abscisic acid signalling and engineering drought hardiness in plants. Nature 2001, 410, 327–330. [Google Scholar] [CrossRef]
- Young, J.J.; Mehta, S.; Israelsson, M.; Godoski, J.; Grill, E.; Schroeder, J.I. CO2 signaling in guard cells: Calcium sensitivity response modulation, a Ca2+-independent phase, and CO2 insensitivity of the gca2 mutant. Proc. Natl. Acad. Sci. USA 2006, 103, 7506–7511. [Google Scholar] [CrossRef] [Green Version]
- Woodward, F.I. Stomatal numbers are sensitive to increases in CO2 from pre-industrial levels. Nature 1987, 327, 617–618. [Google Scholar] [CrossRef]
- Keeling, C.D.; Bacastow, R.B.; Bainbridge, A.E.; Ekdahl, C.A.; Guenther, P.R.; Waterman, L.S.; Chin, J.F.S. Atmospheric carbon-dioxide variations at Mauna-Loa Observatory, Hawaii. Tellus 1976, 28, 538–551. [Google Scholar]
- Neftel, A.; Moor, E.; Oeschger, H.; Stauffer, B. Evidence from polar ice cores for the increase in atmospheric CO2 in the past 2 centuries. Nature 1985, 315, 45–47. [Google Scholar] [CrossRef]
- Mcelwain, J.C.; Chaloner, W.G. Stomatal density and index of fossil plants track atmospheric carbon-dioxide in the Paleozoic. Ann. Bot. 1995, 76, 389–395. [Google Scholar] [CrossRef]
- Chater, C.; Gray, J.E.; Beerling, D.J. Early evolutionary acquisition of stomatal control and development gene signalling networks. Curr. Opin. Plant Biol. 2013, 16, 638–646. [Google Scholar] [CrossRef]
- Mott, K.A. Do stomata respond to CO2 concentrations other than intercellular? Plant Physiol. 1988, 86, 200–203. [Google Scholar] [CrossRef] [Green Version]
- Medlyn, B.E.; Robinson, A.P.; Clement, R.; McMurtrie, R.E. On the validation of models of forest CO2 exchange using eddy covariance data: Some perils and pitfalls. Tree Physiol. 2005, 25, 839–857. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.H.; Boisson-Dernier, A.; Israelsson-Nordström, M.; Böhmer, M.; Xue, S.; Ries, A.; Godoski, J.; Kuhn, J.M.; Schroeder, J.I. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat. Cell Biol. 2010, 12, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Hou, C.; Ren, Z.; Pan, Y.; Jia, J.; Zhang, H.; Bai, F.; Zhang, P.; Zhu, H.; He, Y.; et al. A molecular pathway for CO2 response in Arabidopsis guard cells. Nat. Commun. 2015, 6, 6057. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Negi, J.; Young, J.; Israelsson, M.; Schroeder, J.I.; Iba, K. Arabidopsis HT1 kinase controls stomatal movements in response to CO2. Nat. Cell Biol. 2006, 8, 391–397. [Google Scholar] [CrossRef]
- Hashimoto-Sugimoto, M.; Negi, J.; Monda, K.; Higaki, T.; Isogai, Y.; Nakano, T.; Hasezawa, S.; Iba, K. Dominant and recessive mutations in the Raf-like kinase HT1 gene completely disrupt stomatal responses to CO2 in Arabidopsis. J. Exp. Bot. 2016, 67, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- Hõrak, H.; Sierla, M.; Tõldsepp, K.; Wang, C.; Wang, Y.S.; Nuhkat, M.; Valk, E.; Pechter, P.; Merilo, E.; Salojärvi, J.; et al. A dominant mutation in the HT1 kinase uncovers roles of MAP kinases and GHR1 in CO2-induced stomatal closure. Plant Cell 2016, 28, 2493–2509. [Google Scholar] [CrossRef] [Green Version]
- Jakobson, L.; Vaahtera, L.; Tõldsepp, K.; Nuhkat, M.; Wang, C.; Wang, Y.S.; Hõrak, H.; Valk, E.; Pechter, P.; Sindarovska, Y.; et al. Natural variation in Arabidopsis Cvi-0 accession reveals an important role of MPK12 in guard cell CO2 signaling. PLoS Biol. 2016, 14, e2000322. [Google Scholar] [CrossRef] [Green Version]
- Tõldsepp, K.; Zhang, J.; Takahashi, Y.; Sindarovska, Y.; Hõrak, H.; Ceciliato, P.H.O.; Koolmeister, K.; Wang, Y.S.; Vaahtera, L.; Jakobson, L.; et al. Mitogen-activated protein kinases MPK4 and MPK12 are key components mediating CO2-induced stomatal movements. Plant J. 2018, 96, 1018–1035. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; De-oliveira-Ceciliato, P.; Takahashi, Y.; Schulze, S.; Dubeaux, G.; Hauser, F.; Azoulay-Shemer, T.; Tõldsepp, K.; Kollist, H.; Rappel, W.J.; et al. Insights into the molecular mechanisms of CO2-mediated regulation of stomatal movements. Curr. Biol. 2018, 28, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Song, Y.; Miao, Y.; Song, C.P. Behind the scenes: The roles of reactive oxygen species in guard cells. New Phytol. 2014, 201, 1121–1140. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Chater, C.; Peng, K.; Movahedi, M.; Dunn, J.A.; Walker, H.J.; Liang, Y.K.; McLachlan, D.H.; Casson, S.; Isner, J.C.; Wilson, I.; et al. Elevated CO2-induced responses in stomata require ABA and ABA signaling. Curr. Biol. 2015, 25, 2709–2716. [Google Scholar] [CrossRef] [Green Version]
- Hanstein, S.; Beer, D.; de Felle, H.H. Miniaturised carbon dioxide sensor designed for measurements within plant leaves. Sens. Actuators B Chem. 2001, 81, 107–114. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Ort, D.R. More than taking the heat: Crops and global change. Curr. Opin. Plant. Biol. 2010, 13, 241–248. [Google Scholar] [CrossRef]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Wroblewitz, S.; Hüther, L.; Manderscheid, R.; Weigel, H.J.; Wätzig, H.; Dänicke, S. Effect of rising atmospheric carbon dioxide concentration on the protein composition of cereal grain. J. Agric. Food Chem. 2014, 62, 6616–6625. [Google Scholar] [CrossRef]
- Weigel, H.J. Plant quality declines as CO2 levels rise. Elife 2014, 3, e03233. [Google Scholar] [CrossRef]
- Lu, Z.M.; Percy, R.G.; Qualset, C.O.; Zeiger, E. Stomatal conductance predicts yields in irrigated Pima cotton and bread wheat grown at high temperatures. J. Exp. Bot. 1998, 49, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Bahar, B.; Yildirim, M.; Barutcular, C. Relationships between stomatal conductance and yield components in spring durum wheat under Mediterranean conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 45–48. [Google Scholar]
- Prashar, A.; Yildiz, J.; McNicol, J.W.; Bryan, G.J.; Jones, H.G. Infra-red thermography for high through-put field phenotyping in Solanum tuberosum. PLoS ONE 2013, 8, e65816. [Google Scholar] [CrossRef] [PubMed]
- Frommer, W.B. CO2mmon sense. Science 2010, 327, 275–276. [Google Scholar] [CrossRef]
- Chandrashekar, J.; Yarmolinsky, D.; Buchholtz, L.; von Oka, Y.; Sly, W.; Ryba, N.J.P.; Zuker, C.S. The taste of carbonation. Science 2009, 326, 443–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroney, J.V.; Bartlett, S.G.; Samuelsson, G. Carbonic anhydrases in plants and algae. Plant. Cell Environ. 2001, 24, 141–153. [Google Scholar] [CrossRef]
- Fabre, N.; Reiter, I.M.; Becuwe-Linka, N.; Genty, B.; Rumeau, D. Characterization and expression analysis of genes encoding alpha and beta carbonic anhydrases in Arabidopsis. Plant. Cell Environ. 2007, 30, 617–629. [Google Scholar] [CrossRef]
- Hu, H.; Rappel, W.J.; Occhipinti, R.; Ries, A.; Böhmer, M.; You, L.; Xiao, C.; Engineer, C.B.; Boron, W.F.; Schroeder, J.I. Distinct cellular locations of carbonic anhydrases mediate CO2 control of stomatal movements. Plant Physiol. 2015, 169, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hu, H.H.; Qin, X.; Zeise, B.; Xu, D.Y.; Rappel, W.J.; Boron, W.F.; Schroeder, J.I. Reconstitution of CO2 regulation of SLAC1 anion channel and function of CO2-permeable PIP2;1 aquaporin as CARBONIC ANHYDRASE 4 interactor. Plant Cell 2016, 28, 568–582. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.W.; Hu, H.H.; Ries, A.; Merilo, E.; Kollist, H.; Schroeder, J.I. Central functions of bicarbonate in S-type anion channel activation and OST1 protein kinase in CO2 signal transduction in guard cell. Embo J. 2011, 30, 1645–1658. [Google Scholar] [CrossRef] [Green Version]
- Matrosova, A.; Bogireddi, H.; Mateo-Peñas, A.; Hashimoto-Sugimoto, M.; Iba, K.; Schroeder, J.I.; Israelsson-Nordström, M. The HT1 protein kinase is essential for red light-induced stomatal opening and genetically interacts with OST1 in red light and CO2-induced stomatal movement responses. New Phytol. 2015, 208, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.I.; Hagiwara, S. Cytosolic calcium regulates ion channels in the plasma-membrane of Vicia faba guard-cells. Nature 1989, 338, 427–430. [Google Scholar] [CrossRef]
- Negi, J.; Matsuda, O.; Nagasawa, T.; Oba, Y.; Takahashi, H.; Kawai-Yamada, M.; Uchimiya, H.; Hashimoto, M.; Iba, K. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature 2008, 452, 483–486. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Negi, J.; Wang, C.; Isogai, Y.; Schroeder, J.I.; Iba, K. The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis. Plant. Cell 2016, 28, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, N.; Miao, Y.; Hauser, F.; McCammon, J.A.; Rappel, W.J.; Schroeder, J.I. Identification of SLAC1 anion channel residues required for CO2/bicarbonate sensing and regulation ofstomatal movements. Proc. Natl. Acad. Sci. USA 2018, 115, 11129–11137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, D.; Scherzer, S.; Mumm, P.; Stange, A.; Marten, I.; Bauer, H.; Ache, P.; Matschi, S.; Liese, A.; Al-Rasheid, K.A.S.; et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc. Natl. Acad. Sci. USA 2009, 106, 21425–21430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, D.P.; Wang, C.; He, J.N.; Liao, H.; Duan, Y.; Zhu, Z.Q.; Guo, Y.; Chen, Z.Z.; Gong, Z.Z. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell 2012, 24, 2546–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jammes, F.; Song, C.; Shin, D.J.; Munemasa, S.; Takeda, K.; Gu, D.; Cho, D.; Lee, S.; Giordo, R.; Sritubtim, S.; et al. MAP kinases MPK9 and MPK12 are preferentially expressed in guard cells and positively regulate ROS-mediated ABA signaling. Proc. Natl Acad. Sci. USA 2009, 106, 20520–20525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, G.J.; Chu, S.P.; Harrington, C.L.; Schumacher, K.; Hoffmann, T.; Tang, Y.Y.; Grill, E.; Schroeder, J.I. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 2001, 411, 1053–1057. [Google Scholar] [CrossRef]
- Tang, M.; Zhao, X.; Hu, Y.; Zeng, M.; Wang, K.; Dong, N.; Ma, X.; Bai, L.; Song, C.P. Arabidopsis guard cell CO2/HCO3− response mutant screening by an aequorin-based calcium imaging system. Plant. Methods 2020, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.A.R.; McAinsh, M.R.; Mansfield, T.A.; Hetherington, A.M. Carbon dioxide induces increases in guard cell cytosolic free calcium. Plant. J. 1996, 9, 297–304. [Google Scholar] [CrossRef]
- Schwartz, A.; Ilan, N.; Grantz, D.A. Calcium Effects on Stomatal movement in Commelina communis L. use of EGTA to modulate stomatal response to light, KCl and CO2. Plant. Physiol. 1988, 87, 583–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, K.E.; Siegel, R.S.; Valerio, G.; Brandt, B.; Schroeder, J.I. Abscisic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus–response analyses. Ann. Bot. 2012, 109, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Ehonen, S.; Yarmolinsky, D.; Kollist, H.; Kangasjärvi, J. Reactive Oxygen Species, Photosynthesis, and Environment in the Regulation of Stomata. Antioxid Redox Signal. 2019, 30, 1220–1237. [Google Scholar] [CrossRef] [PubMed]
- Sierla, M.; Waszczak, C.; Vahisalu, T.; Kangasjärvi, J. Reactive oxygen species in the regulation of stomatal movements. Plant Physiol. 2016, 171, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Mori, I.C.; Munemasa, S. Diverse stomatal signaling and the signal integration mechanism. Annu. Rev. Plant Biol. 2015, 66, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Parihar, P.; Singh, S.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Reactive oxygen species signaling and stomatal movement: Current updates and future perspectives. Redox. Biol. 2017, 11, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Fluhr, R. Two distinct sources of elicited reactive oxygen species in tobacco epidermal cells. Plant Cell 1997, 9, 1559–1572. [Google Scholar] [CrossRef]
- Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Mori, I.C.; Pinontoan, R.; Kawano, T.; Muto, S. Involvement of superoxide generation in salicylic acid-induced stomatal closure in Vicia faba. Plant Cell Physiol. 2001, 42, 1383–1388. [Google Scholar] [CrossRef]
- Khokon, M.A.R.; Okuma, E.I.J.I.; Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of extracellular oxidative burst in salicylic acid-induced stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Okamoto, H.; Okuma, E.; Shiba, H.; Kamada, H.; Hasegawa, P.M.; Murata, Y. SIZ1 deficiency causes reduced stomatal aperture and enhanced drought tolerance via controlling salicylic acid-induced accumulation of reactive oxygen species in Arabidopsis. Plant. J. 2013, 73, 91–104. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhard, N.; Angel Torres, M.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. Embo J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Boutrot, F.; Rathjen, J.P.; Zipfel, C. ASPARTATE OXIDASE plays an important role in Arabidopsis stomatal immunity. Plant. Physiol 2012, 159, 1845–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Zepeda-Jazo, I.; Shabala, S.; Dobrovinskaya, O. Cross-talk between reactive oxygen species and polyamines in regulation of ion transport across the plasma membrane: Implications for plant adaptive responses. J. Exp. Bot. 2014, 65, 1271–1283. [Google Scholar] [CrossRef] [Green Version]
- An, Z.; Jing, W.; Liu, Y.; Zhang, W. Hydrogen peroxide generated by copper amine oxidase is involved in abscisic acid-induced stomatal closure in Vicia faba. J. Exp. Bot. 2008, 59, 815–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Liu, G.; Hou, L.; Wang, L.; Liu, X. Regulatory function of polyamine oxidase- generated hydrogen peroxide in ethylene-induced stomatal closure in Arabidopsis thaliana. J. Integr. Agric. 2013, 12, 251–262. [Google Scholar] [CrossRef]
- Kimura, S.; Kaya, H.; Kawarazaki, T.; Hiraoka, G.; Senzaki, E.; Michikawa, M.; Kuchitsu, K. Protein phosphorylation is a prerequisite for the Ca2+-dependent activation of Arabidopsis NADPH oxidases and may function as a trigger for the positive feedback regulation of Ca2+ and reactive oxygen species. Biochim Biophys Acta 2012, 1823, 398–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhorst, L.; Kudla, J. Calcium and reactive oxygen species rule the waves of signaling. Plant. Physiol. 2013, 163, 471–485. [Google Scholar] [CrossRef] [Green Version]
- Drerup, M.M.; Schlücking, K.; Hashimoto, K.; Manishankar, P.; Steinhorst, L.; Kuchitsu, K.; Kudla, J. The calcineurin B-like calcium sensors CBL1 and CBL9 together with their interacting protein kinase CIPK26 regulate the Arabidopsis NADPH oxidase RBOHF. Mol. Plant 2013, 6, 559–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, H.; Komori, T.; Kobori, M.; Nakahira, Y.; Shiina, T. Evidence for chloroplast control of external Ca2+-induced cytosolic Ca2+ transients and stomatal closure. Plant J. 2008, 53, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Nomura, H.; Komori, T.; Uemura, S.; Kanda, Y.; Shimotani, K.; Nakai, K.; Furuichi, T.; Takebayashi, K.; Sugimoto, T.; Sano, S.; et al. Chloroplast-mediated activation of plant immune signalling in Arabidopsis. Nat. Commun. 2012, 3, 926. [Google Scholar] [CrossRef] [Green Version]
- Vainonen, J.P.; Sakuragi, Y.; Stael, S.; Tikkanen, M.; Allahverdiyeva, Y.; Paakkarinen, V.; Aro, E.; Suorsa, M.; Scheller, H.V.; Vener, A.V.; et al. Light regulation of CaS, a novel phosphoprotein in the thylakoid membrane of Arabidopsis thaliana. Febs J. 2008, 275, 1767–1777. [Google Scholar] [CrossRef]
- Weinl, S.; Held, K.; Schlcking, K.; Steinhorst, L.; Kuhlgert, S.; Hippler, M.; Kudla, J. A plastid protein crucial for Ca2+-regulated stomatal responses. New Phytol. 2008, 179, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Chen, J.; Liu, T.W.; Han, A.D.; Simon, M.; Dong, X.J.; He, J.X.; Zheng, H.L. Regulation of the calcium-sensing receptor in both stomatal movement and photosynthetic electron transport is crucial for water use efficiency and drought tolerance in Arabidopsis. J. Exp. Bot. 2014, 65, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Du, Y.; Hou, Y.J.; Zhao, Y.; Hsu, C.C.; Yuan, F.; Zhu, X.; Tao, W.A.; Song, C.P.; Zhu, J.K. Nitric oxide negatively regulates abscisic acid signaling in guard cells by S- nitrosylation of OST1. Proc. Natl. Acad. Sci. USA 2015, 112, 613–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueoka-Nakanishi, H.; Sazuka, T.; Nakanishi, Y.; Maeshima, M.; Mori, H.; Hisabori, T. Thioredoxin h regulates calcium dependent protein kinases in plasma membranes. Febs J. 2013, 280, 3220–3231. [Google Scholar] [CrossRef]
- Sierla, M.; Hõrak, H.; Overmyer, K.; Waszczak, C.; Yarmolinsky, D.; Maierhofer, T.; Vainonen, J.P.; Salojärvi, J.; Denessiouk, K.; Laanemets, K.; et al. The Receptor-like pseudokinase GHR1 is required for stomatal closure. Plant. Cell 2018, 30, 2813–2837. [Google Scholar] [CrossRef] [Green Version]
- Kolla, V.A.; Vavasseur, A.; Raghavendra, A.S. Hydrogen peroxide production is an early event during bicarbonate induced stomatal closure in abaxial epidermis of Arabidopsis. Planta 2007, 225, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, R.X.; Kim, D.S.; Sun, P.; Liu, H.; Liu, Z.; Hetherington, A.M.; Liang, Y.K. ROS of distinct sources and salicylic acid separate elevated CO2-mediated stomatal movements in Arabidopsis. Front. Plant Sci. 2020, 11, 542. [Google Scholar] [CrossRef]
- Fahnenstich, H.; Scarpeci, T.E.; Valle, E.M.; Flügge, U.I.; Maurino, V.G. Generation of hydrogen peroxide in chloroplasts of Arabidopsis overexpressing glycolate oxidase as an inducible system to study oxidative stress. Plant Physiol. 2008, 148, 719–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.M.; Altschmied, L.; Chory, J. Arabidopsis mutants define downstream branches in the phototransduction pathway. Genes Dev. 1994, 8, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Ruegger, M.; Dewey, E.; Hobbie, L.; Brown, D.; Bernasconi, P.; Turner, J.; Muday, G.; Estelle, M. Reduced naphthylphthalamic acid binding in the tir3 mutant of Arabidopsis is associated with a reduction in polar auxin transport and diverse morphological defects. Plant Cell 1997, 9, 745–757. [Google Scholar]
- Gil, P.; Dewey, E.; Friml, J.; Zhao, Y.; Snowden, K.C.; Putterill, J.; Palme, K.; Estelle, M.; Chory, J. BIG: A calossin-like protein required for polar auxin transport in Arabidopsis. Genes Dev. 2001, 15, 1985–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanyuka, K.; Praekelt, U.; Franklin, K.A.; Billingham, O.E.; Hooley, R.; Whitelam, G.C.; Halliday, K.J. Mutations in the huge Arabidopsis gene BIG affect a range of hormone and light responses. Plant J. 2003, 35, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Pérez-Torres, A.; Rampey, R.A.; Bartel, B.; Herrera-Estrella, L. An auxin transport independent pathway is involved in phosphate stress-induced root architectural alterations in Arabidopsis. Identification of BIG as a mediator of auxin in pericycle cell activation. Plant Physiol. 2005, 137, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Paciorek, T.; Zažímalová, E.; Ruthardt, N.; Petrášek, J.; Stierhof, Y.D.; Kleine-Vehn, J.; Morris, D.A.; Emans, N.; Jürgens, G.; Geldner, N.; et al. Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 2005, 435, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Hearn, T.J.; Ruiz, M.C.M.; Abdul-Awal, S.M.; Wimalasekera, R.; Stanton, C.R.; Haydon, M.J.; Theodoulou, F.L.; Hannah, M.A.; Alex, A.R.W. BIG regulates dynamic adjustment of circadian period in Arabidopsis thaliana. Plant Physiol. 2018, 178, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, N.; Suzuki, M.; Fukaki, H.; Morita-Terao, M.; Tasaka, M.; Komeda, Y. CRM1/BIG-mediated auxin action regulates Arabidopsis inflorescence development. Plant. Cell Physiol. 2007, 48, 1275–1290. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Zhang, R.X.; Peng, K.; Tagliavia, C.; Li, S.; Xue, S.; Liu, A.; Hu, H.; Zhang, J.; Hubbard, K.E.; et al. The BIG protein distinguishes the process of CO2-induced stomatal closure from the inhibition of stomatal opening by CO2. New Phytol. 2018, 218, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merilo, E.; Laanemets, K.; Hu, H.; Xue, S.; Jakobson, L.; Tulva, I.; Gonzalez-Guzman, M.; Rodriguez, P.L.; Schroeder, J.I.; Broschè, M.; et al. PYR/RCAR receptors contribute to ozone-, reduced air humidity-, darkness-, and CO2-induced stomatal regulation. Plant Physiol. 2013, 162, 1652–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, A.; Hetherington, A.M. Convergence of the abscisic acid, CO2, and extracellular calcium signal transduction pathways in stomatal guard cells. Plant Physiol. 1997, 114, 1557–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, K.; Li, X.; Zhang, H.; Zhang, G.; Liu, Y.; Zhou, Y.; Xia, X.; Chen, Z.; Yu, J. Guard cell hydrogen peroxide and nitric oxide mediate elevated CO2-induced stomatal movement in tomato. New Phytol. 2015, 208, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Mithani, A.; Belfield, E.J.; Mott, R.; Hurst, L.D.; Harberd, N.P. Environmentally responsive genome-wide accumulation of de novo Arabidopsis thaliana mutations and epimutations. Genome Res. 2014, 24, 1821–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forgione, I.; Wołoszyńska, M.; Pacenza, M.; Chiappetta, A.; Greco, M.; Araniti, F.; Abenavoli, M.R.; Lijsebettens, M.V.; Bitonti, M.B.; Bruno, L. Hypomethylated drm1 drm2 cmt3 mutant phenotype of Arabidopsis thaliana is related to auxin pathway impairment. Plant Sci. 2019, 280, 383–396. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic model of CO2-induced stomatal closure.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ma, X.; Bai, L. Elevated CO2 and Reactive Oxygen Species in Stomatal Closure. Plants 2021, 10, 410. https://doi.org/10.3390/plants10020410
AMA Style
Ma X, Bai L. Elevated CO2 and Reactive Oxygen Species in Stomatal Closure. Plants. 2021; 10(2):410. https://doi.org/10.3390/plants10020410
Chicago/Turabian StyleMa, Xiaonan, and Ling Bai. 2021. "Elevated CO2 and Reactive Oxygen Species in Stomatal Closure" Plants 10, no. 2: 410. https://doi.org/10.3390/plants10020410
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.