
Metabolic Changes in Seed Embryos of Hypoxia-Tolerant Rice and Hypoxia-Sensitive Barley at the Onset of Germination


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
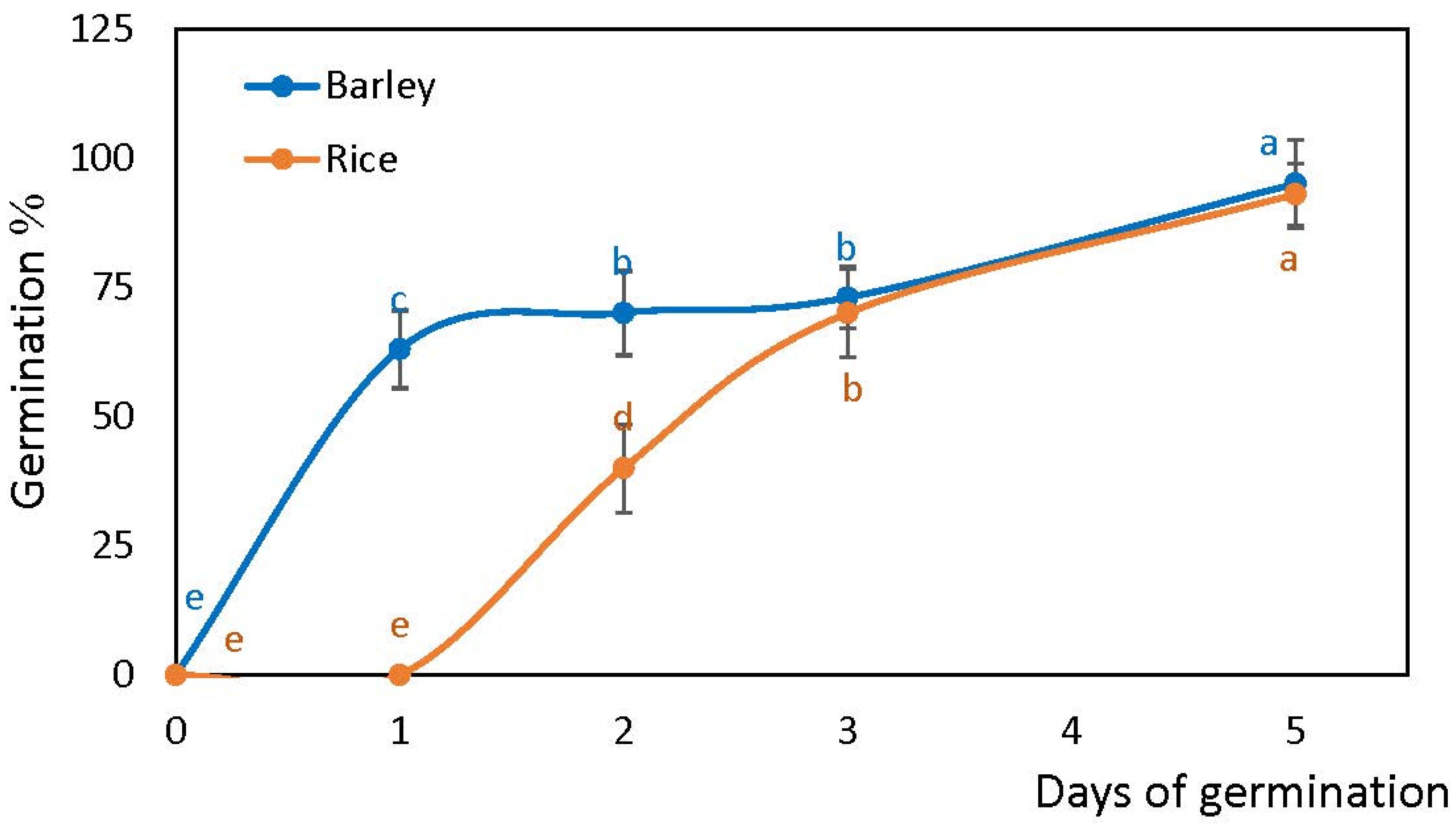
2.1. Germination Rates of Rice and Barley Seeds
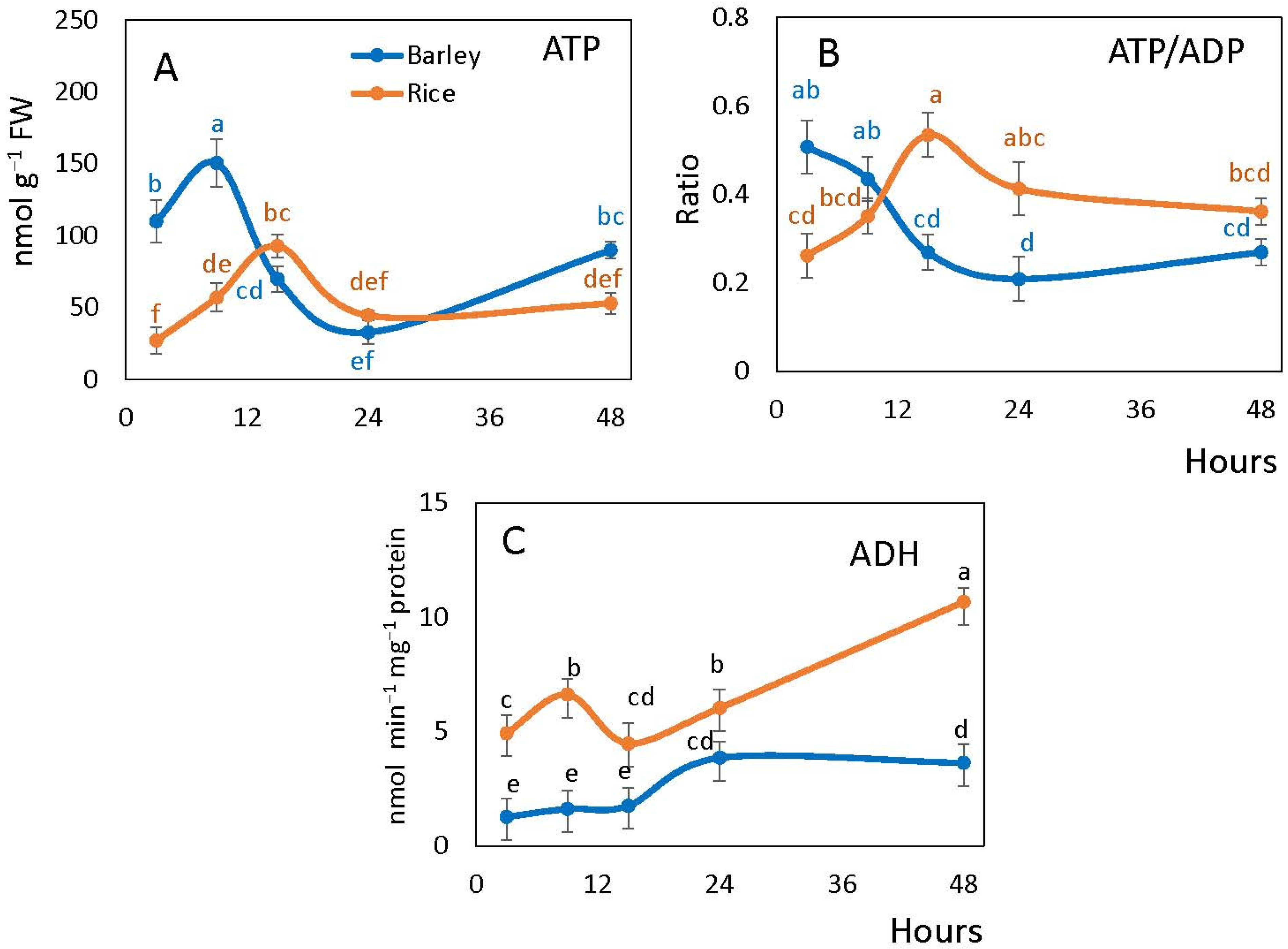
2.2. Alcohol Dehydrogenase Activity and the Level of Adenylates during Germination
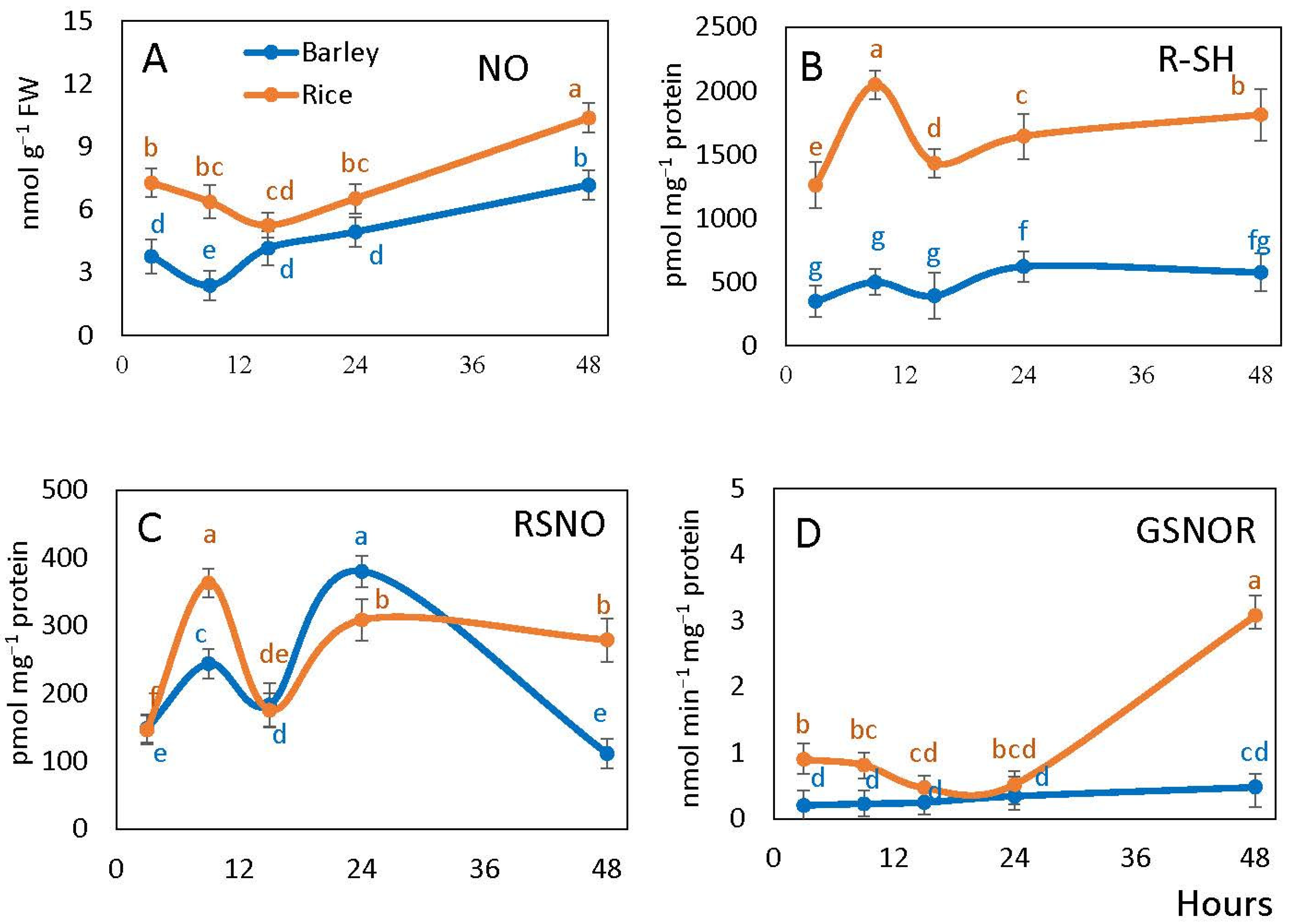
2.3. Nitric Oxide, Free Thiols, S-Nitrosoglutathione Reductase and S-Nitrosylation
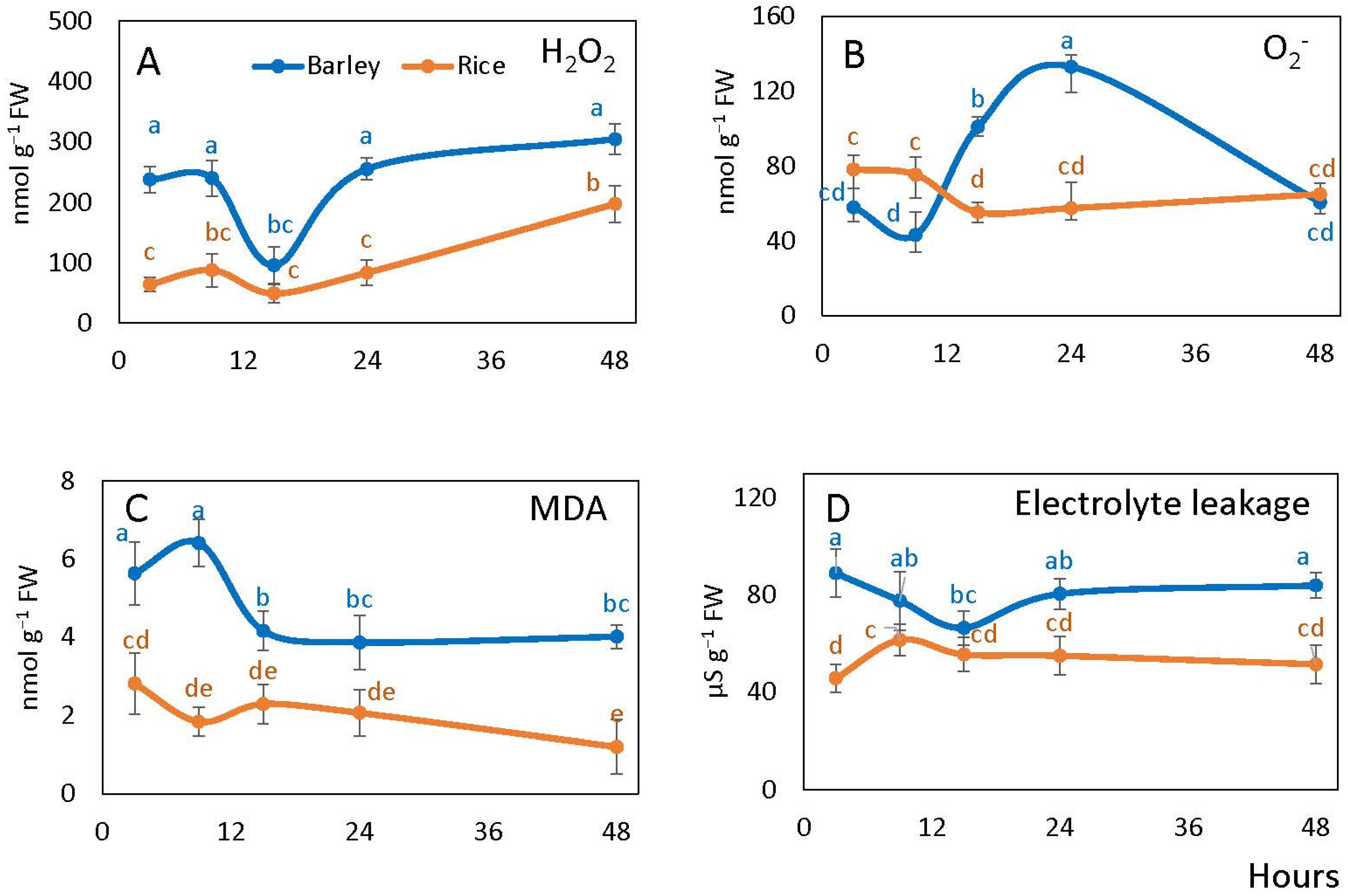
2.4. Reactive Oxygen Species, Lipid Peroxidation and Electrolyte Leakage in Rice and Barley Seeds
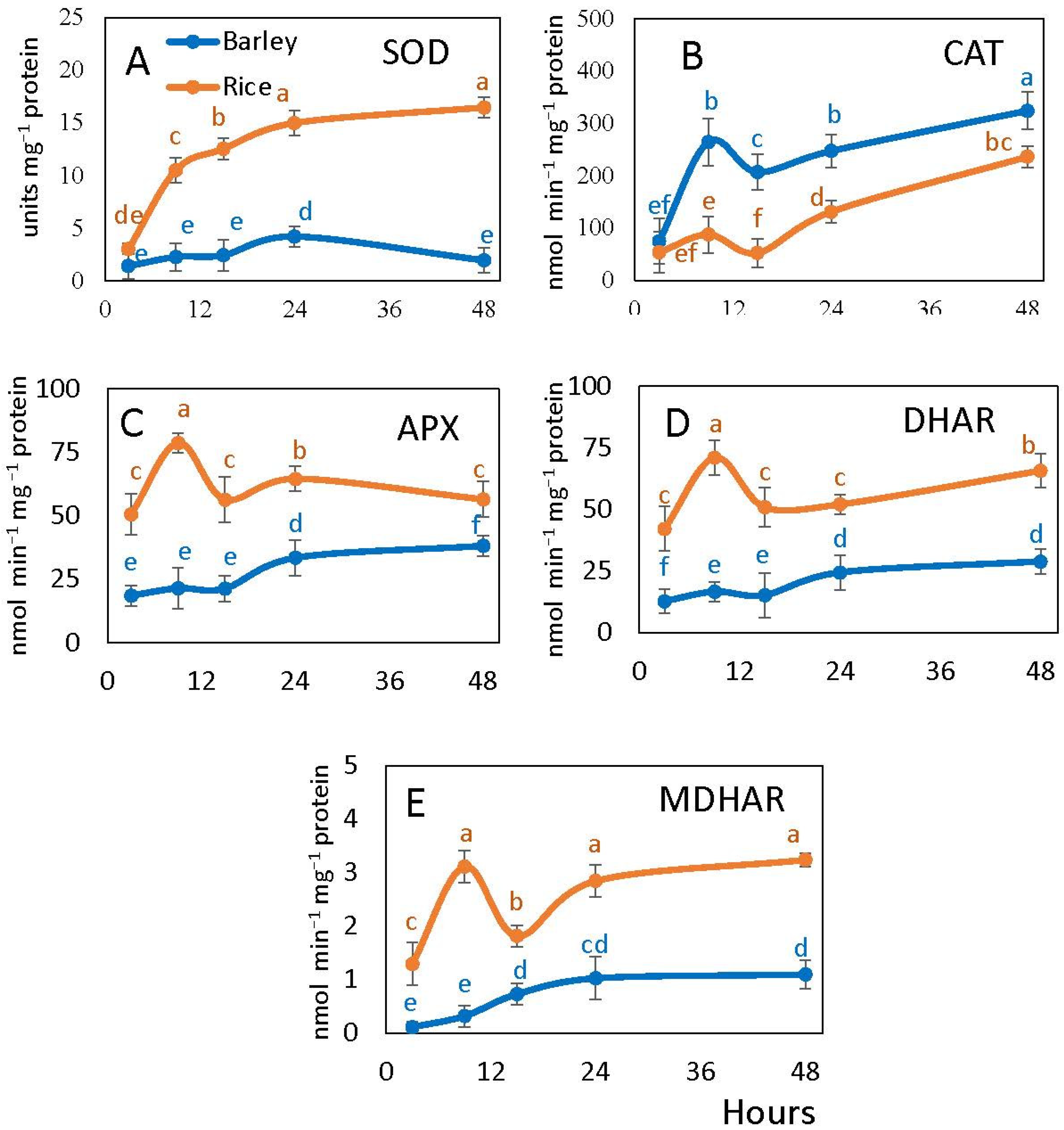
2.5. Antioxidant Enzyme Activities in Rice and Barley Seeds
3. Discussion
3.1. Seed Germination Phases and Hypoxic Conditions under Seed Coat
3.2. Respiration and Energy Availability in the Embryos of Imbibed Seeds
3.3. Nitric Oxide and ROS Scavengers in Embryo of Imbibed Seeds
3.4. Reactive Oxygen Species and Antioxidant System in Seed Embryo during Germination
4. Materials and Methods
4.1. Seed Germination and Isolation of Embryos
4.2. Alcohol Dehydrogenase and Adenylate Ratios
4.3. Nitric Oxide, Free Thiols, S-Nitrosylation and S-Nitrosoglutathione Reductase
4.4. Hydrogen Peroxide, Electrolyte Leakage and Lipid Peroxidation
4.5. Superoxide Dismutase, Catalase and Ascorbate-Glutathione Cycle Enzymes
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hockett, E.A.; Nilan, R.A. Genetics. In Barley; Rasmusson, D.C., Ed.; American Society of Agronomy: Madison, WI, USA, 1985; pp. 187–230. [Google Scholar]
- He, D.; Wang, Q.; Wang, K.; Yang, P. Genome-Wide Dissection of the MicroRNA Expression Profile in Rice Embryo during Early Stages of Seed Germination. PLoS ONE 2015, 10, e0145424. [Google Scholar] [CrossRef]
- Perata, P.; Guglielminetti, L.; Alpi, A. Mobilization of endosperm reserves in cereal under anoxia. Ann. Bot. 1997, 79, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Kamrun, N.; Masayuki, F. Plant response to Salt Stress and Role of Exogenous Protectants to Mitigate Salt-Induced Damages. In Ecophysiology and Responses of Plant under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Mwami, B.M.; Nguluu, S.N.; Kimiti, J.M.; Kimatu, J.N. Effects of water imbibition of selected bean varieties on germination. Int. J. Innov. Sci. Res. Technol. 2017, 3, 57–62. [Google Scholar]
- Bewley, J.D.; Black, M. Physiology of Development and Germination, 2nd ed.; Plenum Press: New York, NY, USA, 1994; 445 p. [Google Scholar]
- Nonogaki, H. Seed germination and dormancy: The classic story, new puzzles, and evolution. J. Integr. Plant Biol. 2019, 61, 541–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nutbeam, A.R.; Duffus, C.M. Oxygen exchange in the pericarp green layer of immature cereal grains. Plant Physiol. 1978, 62, 360–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisjuk, L.; Rolletschek, H. The oxygen status of the developing seed. New Phytol. 2009, 182, 17–30. [Google Scholar] [CrossRef]
- Larson, L.A. The effect of soaking pea seeds with or without seedcoats has on seedling growth. Plant Physiol. 1968, 43, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Bykova, N.V.; Hoehn, B.; Rampitsch, C.; Hu, J.; Stebbing, J.A.; Knox, R. Thiol redox-sensitive seed proteome in dormant and non-dormant hybrid genotypes of wheat. Phytochemistry 2011, 72, 1162–1172. [Google Scholar] [CrossRef]
- Davies, D.D. Anaerobic metabolism and the production of organic acids. In The Biochemistry of Plants; Davies, D.D., Ed.; Academic Press: London, UK, 1980; Volume 2, pp. 581–611. [Google Scholar]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic metabolism in plants. Plant Physiol. 1992, 100, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.; Greenway, H. Mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Funct. Plant Biol. 2003, 30, 999–1036. [Google Scholar] [CrossRef]
- Duff, S.M.G.; Guy, P.A.; Nie, X.; Durnin, D.C.; Hill, R.D. Hemoglobin expression in germinating barley. Seed Sci. Res. 1998, 8, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Logan, D.C.; Millar, A.H.; Sweetlove, L.J.; Hill, S.A.; Leaver, C.J. Mitochondrial biogenesis during germination in maize embryos. Plant Physiol. 2001, 125, 662–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, P.C.; Gubler, F.; Jacobsen, J.V.; Jones, R.L. Dormancy of Arabidopsis seeds barley grains can be broken by nitric oxide. Planta 2004, 219, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; El-Maarouf-Bouteau, H.; Kranner, I.; Bogatek, R.; Corbineau, F.; Bailly, C. The mechanisms involved in seed dormancy alleviation by hydrogen cyanide unravel the role of reactive oxygen species as key factors of cellular signaling during germination. Plant Physiol. 2009, 150, 494–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulff, A.; Oliveira, H.C.; Saviani, E.E.; Salgado, I. Nitrite reduction and superoxide-dependent nitricoxidedegradation by Arabidopsis mitochondria: Influence of external NAD(P)H dehydrogenases and alternative oxidase in the control of nitric oxide levels. Nitric Oxide 2009, 21, 132–139. [Google Scholar] [CrossRef]
- Tai, L.; Wang, H.J.; Xu, X.J.; Sun, W.H.; Ju, L.; Liu, W.T.; Li, W.Q.; Sun, J.; Chen, K.M. Cereal pre-harvest sprouting: A global agricultural disaster regulated by complex genetic and biochemical mechanisms. J. Exp. Bot. 2021, 72, 2857–2876. [Google Scholar] [CrossRef]
- Kranner, I.; Minibayeva, F.V.; Beckett, R.P.; Seal, C.E. What is stress? Concepts, definitions and applications in seed science. New Phytol. 2010, 188, 655–673. [Google Scholar] [CrossRef]
- Bykova, N.V.; Hoehn, B.; Rampitsch, C.; Bykova, N.V.; Hoehn, B.; Rampitsch, C.; Banks, T.; Stebbing, J.-A.; Fan, T.; alKnox, R. Redox-sensitive proteome and antioxidant strategies in wheat seed dormancy control. Proteomics 2011, 11, 865–882. [Google Scholar] [CrossRef]
- Bykova, N.V.; Hu, J.; Ma, Z.; Igamberdiev, A.U. The Role of Reactive Oxygen and Nitrogen Species in Bioenergetics, Metabolism and Signaling during Seed Germination. In Reactive Oxygen and Reactive Nitrogen Species Signaling and Communication in Plants; Gupta, K.J., Igamberdiev, A.U., Eds.; Springer: Berlin, Germany, 2015; pp. 177–195. [Google Scholar] [CrossRef]
- Debska, K.; Krasuska, U.; Budnicka, K.; Bogatek, R.; Gniazdowska, A. Dormancy removal of apple seeds by cold stratification is Associated with fluctuation in H2O2, NO production and protein carbonylation level. J. Plant Physiol. 2013, 170, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Colville, L.; Kranner, I. Desiccation tolerant plants as model systems to study redox regulation of protein thiols. Plant Growth Regul. 2010, 62, 241–255. [Google Scholar] [CrossRef]
- Davies, M.J.; Fu, S.; Wang, H.; Dean, R.T. Stable markers of oxidant damage to proteins and their application in study of human disease. Free Radic. Biol. Med. 1999, 27, 1151–1161. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu Rev Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gara, L.; de Pinto, M.C.; Arrigoni, O. Ascorbate synthesis and ascorbate peroxidase activity during the early stage of wheat germination. Physiol Plant. 1997, 100, 894–900. [Google Scholar] [CrossRef]
- Tommasi, F.; Paciolla, C.; de Pinto, M.C.; De Gara, L. A comparative study of glutathione and ascorbate metabolism during germination of Pinus Pinea L. seeds. J. Exp Bot. 2001, 52, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Job, C.; Belghazi, M.; Job, D.; Leubner-Metzger, G. Proteomics reveal tissue-specific features of the cress (Lepidium sativum L.) endosperm cap proteome and its hormone-induced changes during seed germination. Proteomics 2010, 10, 406–416. [Google Scholar] [CrossRef]
- Ma, Z.; Marsolais, F.; Bernards, M.A.; Sumarah, M.W.; Bykova, N.V.; Igamberdiev, A.U. Glyoxylate cycle and metabolism of organic acids in the scutellum of barley seeds during germination. Plant Sci. 2016, 248, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Marsolais, F.; Bykova, N.V.; Igamberdiev, A.U. Nitric Oxide and Reactive Oxygen Species Mediate Metabolic Changes in Barley Seed Embryo during Germination. Front. Plant Sci. 2016, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination–stillamystery. Plant Science 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Gruwel, M.L.H.; Chatson, B.; Yin, X.S.; Abrams, S.R. A magnetic resonance study of water uptake of whole barley kernels. Int. J. Food Sci. Technol. 2001, 36, 161–168. [Google Scholar] [CrossRef]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef]
- Geigenberger, P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol. 2013, 6, 247–256. [Google Scholar] [CrossRef]
- Rolletschek, H.; Weschke, W.; Weber, H.; Wobus, U.; Borisjuk, L. Energy state and its control on seed development: Starch accumulation is associated with high ATP and steep oxygen gradients within barley grains. J. Exp. Bot. 2004, 55, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef]
- He, D.; Yang, P. Proteomics of rice seed germination. Front. Plant Sci. 2013, 4, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benamar, A.; Tallon, C.; Macherel, D. Membrane integrity and oxidative properties of mitochondria isolated from imbibing pea seeds after priming or accelerated ageing. Seed Sci. Res. 2003, 13, 35–45. [Google Scholar] [CrossRef]
- He, D.; Han, C.; Yang, P. Gene expression profile changes in germinating rice. J. Integr. Plant Biol. 2011, 53, 835–844. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Hill, R.D. Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J. Exp. Bot. 2004, 55, 2473–2482. [Google Scholar] [CrossRef] [Green Version]
- Zafari, S.; Hebelstrup, K.H.; Igamberdiev, A.U. Transcriptional and metabolic changes associated with phytoglobin expression during germination of barley seeds. Int. J. Mol. Sci. 2020, 21, 2796. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Strbac, D.; Marschner, H. Activities of hydrogen peroxide scavenging enzymes in germinating wheat seeds. J. Exp. Bot. 1993, 44, 127–132. [Google Scholar] [CrossRef]
- Caliskan, M.; Cuming, A.C. Spatial specificity of H2O2-generating oxalate oxidase gene expression during wheat embryo germination. Plant J. 1998, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Wojtyla, Ł.; Garnczarska, M.; Zalewski, T.; Bednarski, W.; Ratajczak, L.; Jurga, S. A comparative study of water distribution, free radical production and activation of antioxidative metabolism in germinating pea seeds. J. Plant Physiol. 2006, 163, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Igamberdiev, A.U. The anoxic plant mitochondrion as a nitrite: NO reductase. Mitochondrion 2011, 11, 537–543. [Google Scholar] [CrossRef]
- Nie, X.Z.; Hill, R.D. Mitochondrial respiration and hemoglobin gene expression in barley aleurone tissue. Plant Physiol. 1997, 114, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Hunt, P.W.; Watts, R.A.; Trevaskis, B.; Llewelyn, D.J.; Burnell, J.; Dennis, E.S.; Peacock, W.J. Expression and evolution of functionally distinct hemoglobin genes in plants. Plant Mol. Biol. 2001, 47, 677–692. [Google Scholar] [CrossRef]
- Ross, E.J.H.; Shearman, L.; Mathiesen, M.; Zhou, Y.J.; Arredondo-Peter, R.; Sarath, G.; Klucas, R.V. Nonsymbiotic hemoglobins in rice are synthesized during germination and in differentiating cell types. Protoplasma 2001, 218, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Singh, P.; Kaladhar, V.C.; Bhatoee, M.; Paul, D.; Pathak, P.K.; Gupta, K.J. Phytoglobin-NO cycle and AOX pathway play a role in anaerobic germination and growth of deepwater rice. Plant Cell Environ. 2021. [Google Scholar] [CrossRef]
- Corpas, F.J.; Alché, J.D.; Barroso, J.B. Current overview of S-nitrosoglutathione (GSNO) in higher plants. Front. Plant Sci. 2013, 4, 126. [Google Scholar] [CrossRef] [Green Version]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubala, S.; Wojtyla, Ł.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced expression of the proline synthesis gene P5CSA in relations to seed osmopriming improvement of Brassica napus germination under salinity stress. J. Plant Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, N.A.; Sharma, P.; Gill, S.; Hasanuzzaman, M.; Khan, E.A.; Kachhap, K.; Mohamed, A.; Thangavel, P.; Devi, G.D.; Vasudhevan, P.; et al. Catalase and ascorbate peroxidase-representative H2O2-detoxifying heme enzymes in plants. Environ. Sci. Pollut. Res. Int. 2016, 23, 19002–19029. [Google Scholar] [CrossRef] [PubMed]
- Guha, T.; Das, H.; Mukherjee, A.; Kundu, R. Elucidating ROS signaling networks and physiological changes involved in nanoscale zero valent iron primed rice seed germination sensu stricto. Free Radic. Biol. Med. 2021, 171, 11–25. [Google Scholar] [CrossRef]
- Al-Mudaris, M. Notes on various parameters recording the speed of seed germination. Der Trop. 1998, 99, 147–154. [Google Scholar]
- Blandino, A.; Caro, I.; Cantero, D. Comparative study of alcohol dehydrogenase activity in flor yeast extracts. Biotechnol. Lett. 1997, 19, 651–654. [Google Scholar] [CrossRef]
- Joshi, A.K.; Ahmed, S.; Ames, G.F.L. Energy coupling in bacterial periplasmic transport systems. J. Biol. Chem. 1989, 264, 2126–2133. [Google Scholar] [CrossRef]
- Yuroff, A.S.; Sabat, G.; Hickey, W.J. Transporter-mediated uptake of 2-chloro- and 2-hydroxybenzoate by Pseudomonas huttiensis strain D1. Appl. Environ. Microbiol. 2003, 69, 7401–7408. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.M.; Noack, E. Nitric oxide assay using hemoglobin method. Methods Enzymol. 1994, 233, 240–250. [Google Scholar] [CrossRef]
- Cochrane, D.W.; Shah, J.K.; Hebelstrup, K.H.; Igamberdiev, A.U. Expression of phytoglobin affects nitric oxide metabolism and energy state of barley plants exposed to anoxia. Plant Sci. 2017, 265, 124–130. [Google Scholar] [CrossRef]
- Sakamoto, M.; Ueda, H.; Morikawa, H. Arabidopsis glutathione-dependent formaldehyde dehydrogenase is an S-nitrosoglutathione reductase. FEBS Lett. 2002, 515, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Tammela, P.; Vaananen, P.S.; Lakso, I.; Hopia, A.; Vourela, H.; Nygrem, M. Tocopherols, tocotrienols and fatty acids as indicators of natural ageing in Pinus sylvestris seeds. Scand. J. Forest Res. 2005, 20, 378–384. [Google Scholar] [CrossRef]
- Agarie, S.; Hanaoka, N.; Kubota, F.; Agata, W.; Kaufman, B. Measurement of cell membrane stability evaluated by electrolyte leakage as a drought and heat tolerance test in rice (Oryza sativa L.). J. Fac. Agr. Kyushu Univ. 1995, 40, 233–240. [Google Scholar] [CrossRef]
- Heath, R.L.; Parker, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–278. [Google Scholar] [CrossRef]
- Guilbault, G.G. Glucose Oxidase. In Handbook of Enzymatic Methods of Analysis; Schwartz, M.K., Ed.; Marcel Dekker: New York, NY, USA, 1976; pp. 106–109. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayawardhane, J.; Wijesinghe, M.K.P.S.; Bykova, N.V.; Igamberdiev, A.U. Metabolic Changes in Seed Embryos of Hypoxia-Tolerant Rice and Hypoxia-Sensitive Barley at the Onset of Germination. Plants 2021, 10, 2456. https://doi.org/10.3390/plants10112456
Jayawardhane J, Wijesinghe MKPS, Bykova NV, Igamberdiev AU. Metabolic Changes in Seed Embryos of Hypoxia-Tolerant Rice and Hypoxia-Sensitive Barley at the Onset of Germination. Plants. 2021; 10(11):2456. https://doi.org/10.3390/plants10112456
Chicago/Turabian StyleJayawardhane, Jayamini, M. K. Pabasari S. Wijesinghe, Natalia V. Bykova, and Abir U. Igamberdiev. 2021. "Metabolic Changes in Seed Embryos of Hypoxia-Tolerant Rice and Hypoxia-Sensitive Barley at the Onset of Germination" Plants 10, no. 11: 2456. https://doi.org/10.3390/plants10112456