
Sensitive and Rapid Detection of Citrus Scab Using an RPA-CRISPR/Cas12a System Combined with a Lateral Flow Assay

Abstract
:1. Introduction
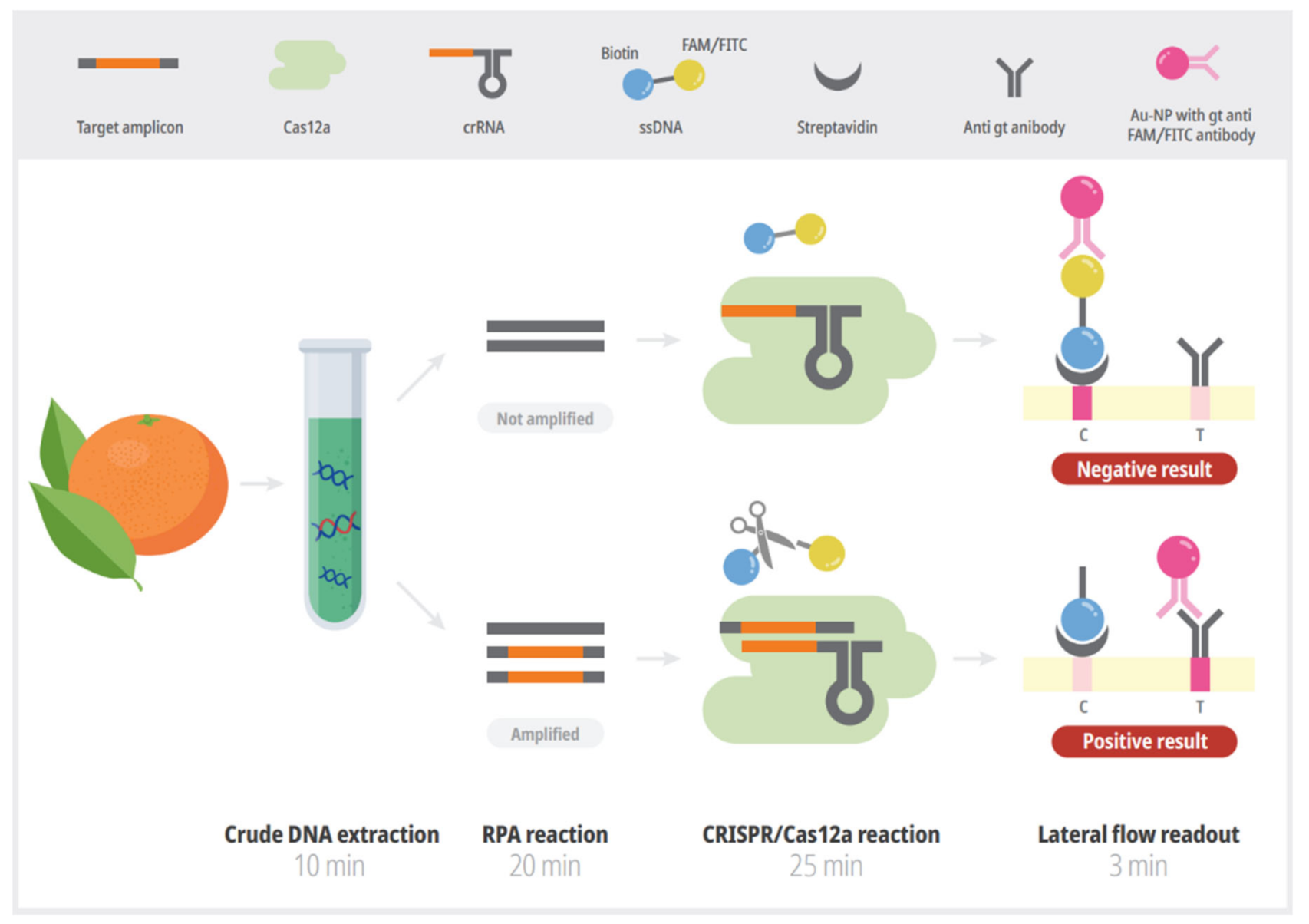
2. Materials and Methods
2.1. Preparation of Positive Control
2.2. RPA Reaction
2.3. CRISPR/Cas12a-LFA Detection
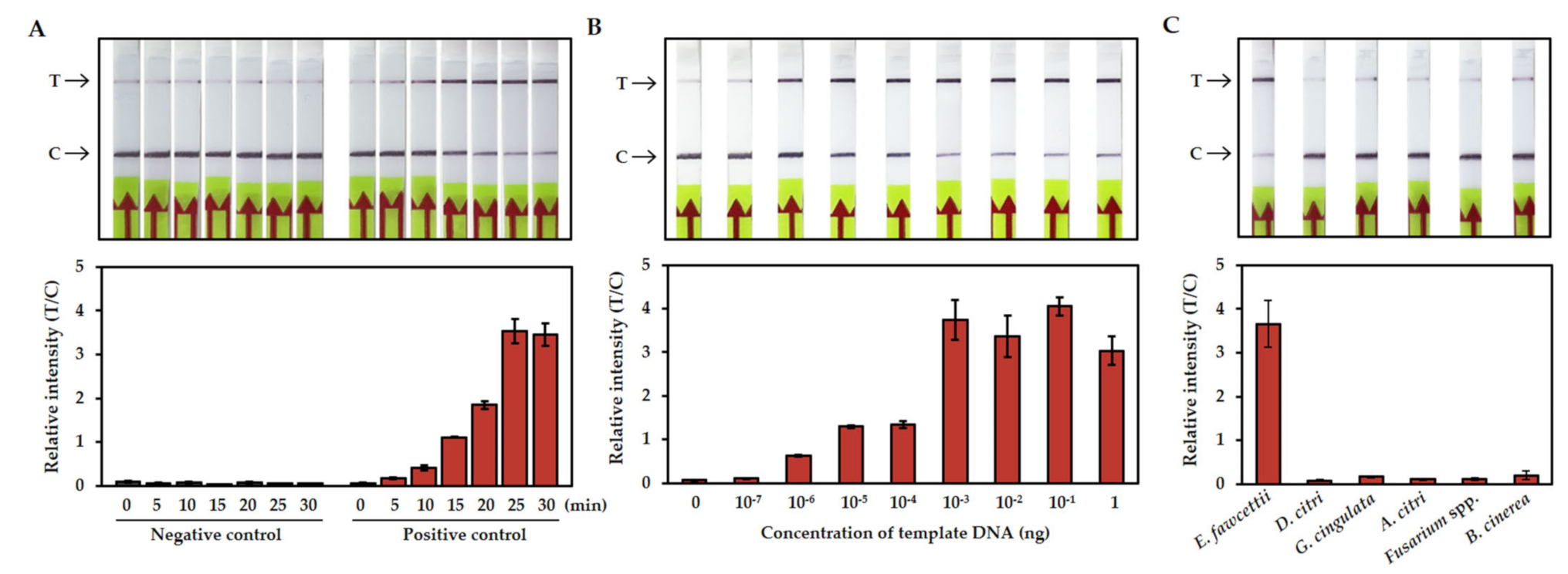
2.4. Sensitivity and Specificity of RPA/Cas12a-LFA Detection
2.5. Preparation of Crude Extract of Plant Samples
2.6. PCR for Field Sample
3. Results
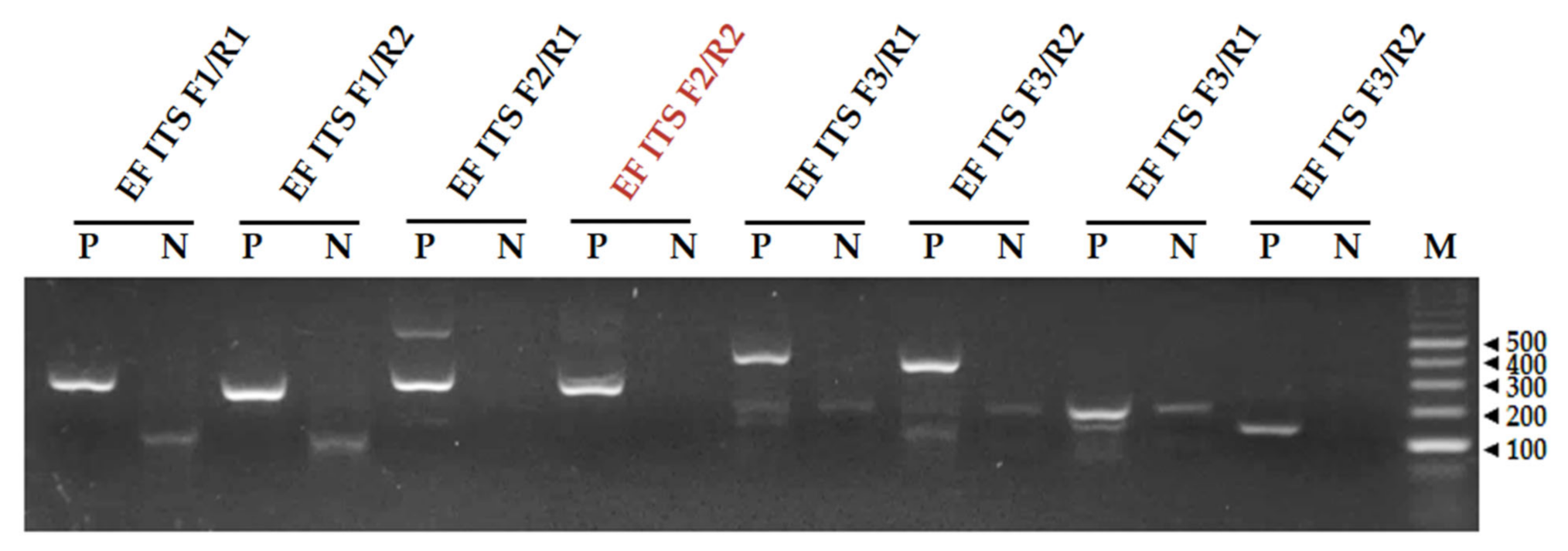
3.1. Screening of Primers for the RPA Reaction
3.2. Establishment of RPA-CRISPR/Cas12a-LFA Detection
3.3. Sensitivity and Specificity of the RPA-CRISPR/Cas12a-LFA Detection
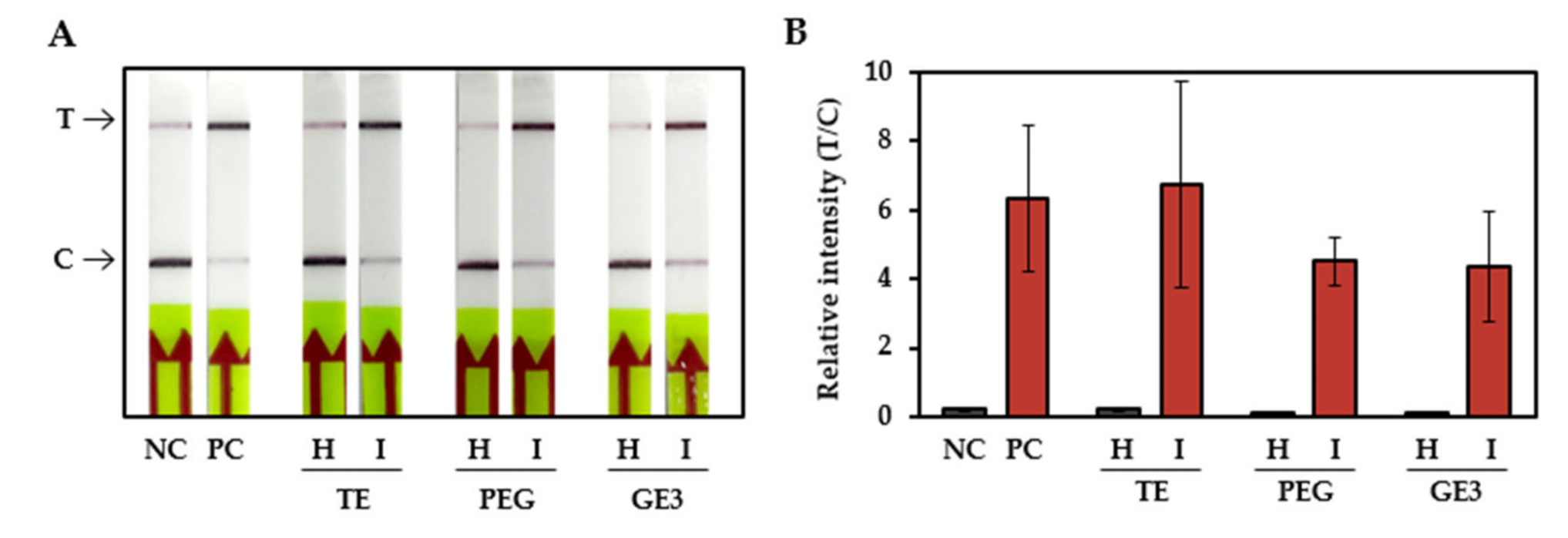
3.4. Citrus Scab Detection with Infected Field Samples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fawcett, H.S. Citrus Scab; Florida Agricultural Experiment Station: Gainesville, FL, USA, 1912. [Google Scholar]
- Bitancourt, A.A.; Jenkins, A.E. Elsinoe fawcettii, the perfect stage of the Citrus scab fungus. Phytopathology 1936, 26, 393–395. [Google Scholar]
- Timmer, L.W.; Priest, M.; Broadbent, P.; Tan, M.K. Morphological and pathological characterization of species of Elsinoe causing scab diseases of citrus. Phytopathology 1996, 86, 1032–1038. [Google Scholar] [CrossRef]
- Paudyal, D.P.; Hyun, J.W. Physical Changes in Satsuma Mandarin Leaf after Infection of Elsinoe fawcettii Causing Citrus Scab Disease. Plant Pathol. J. 2015, 31, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, H.Y.; Botella, J.R. Advanced DNA-Based Point-of-Care Diagnostic Methods for Plant Diseases Detection. Front. Plant Sci. 2017, 8, 2016. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [Green Version]
- Hyun, J.W.; Peres, N.A.; Yi, S.Y.; Timmer, L.W.; Kim, K.S.; Kwon, H.M.; Lim, H.C. Development of PCR Assays for the Identification of Species and Pathotypes of Elsinoe Causing Scab on Citrus. Plant Dis. 2007, 91, 865–870. [Google Scholar] [CrossRef]
- Ahmed, Y.; Hubert, J.; Fourrier-Jeandel, C.; Dewdney, M.M.; Aguayo, J.; Ioos, R. A Set of Conventional and Multiplex Real-Time PCR Assays for Direct Detection of Elsinoe fawcettii, E. australis, and Pseudocercospora angolensis in Citrus Fruits. Plant Dis. 2019, 103, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Hyun, J.W.; Yi, S.H.; Mackenzie, S.J.; Timmer, L.W.; Kim, K.S.; Kang, S.K.; Kwon, H.M.; Lim, H.C. Pathotypes and genetic relationship of worldwide collections of Elsinoe spp. causing scab diseases of citrus. Phytopathology 2009, 99, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Macdonald, J.; von Stetten, F. Review: A comprehensive summary of a decade development of the recombinase polymerase amplification. Analyst 2018, 144, 31–67. [Google Scholar] [CrossRef] [Green Version]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef]
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase polymerase amplification for diagnostic applications. Clin. Chem. 2016, 62, 947–958. [Google Scholar] [CrossRef]
- Zhai, J.; Wang, L.; Qiao, X.; Zhao, J.; Wang, X.; He, X. Detection of Neisseria gonorrhoeae and Chlamydia trachomatis infections in pregnant women by multiplex recombinase polymerase amplification. PLoS ONE 2021, 16, e0251119. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liao, L.; Ma, C.; Zhang, X.; Yu, J.; Yi, L.; Liu, X.; Shen, H.; Gao, S.; Lu, Q. Duplex On-Site Detection of Vibrio cholerae and Vibrio vulnificus by Recombinase Polymerase Amplification and Three-Segment Lateral Flow Strips. Biosensors 2021, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A.; Burbridge, J. Sensitive, Accurate and Rapid Detection of the Northern Root-Knot Nematode, Meloidogyne hapla, Using Recombinase Polymerase Amplification Assays. Plants 2021, 10, 336. [Google Scholar] [CrossRef] [PubMed]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L.B.; Burstein, D.; Chen, J.S.; Paez-Espino, D.; Ma, E.; Witte, I.P.; Cofsky, J.C.; Kyrpides, N.C.; Banfield, J.F.; Doudna, J.A. Programmed DNA destruction by miniature CRISPR-Cas14 enzymes. Science 2018, 362, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Ji, S.; Koh, H.R. CRISPR as a Diagnostic Tool. Biomolecules 2021, 11, 1162. [Google Scholar] [CrossRef]
- Mustafa, M.I.; Makhawi, A.M. SHERLOCK and DETECTR: CRISPR-Cas systems as potential rapid diagnostic tools for emerging infectious diseases. J. Clin. Microbiol. 2021, 59, e00745-20. [Google Scholar] [CrossRef]
- Sajid, M.; Kawde, A.-N.; Daud, M. Designs, formats and applications of lateral flow assay: A literature review. J. Saudi Chem. Soc. 2015, 19, 689–705. [Google Scholar] [CrossRef] [Green Version]
- Poulton, K.; Webster, B. Development of a lateral flow recombinase polymerase assay for the diagnosis of Schistosoma mansoni infections. Anal. Biochem. 2018, 546, 65–71. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Chen, M.; Hu, Z.; Wu, L.; Zhou, M.; Liang, D. Cas12a and Lateral Flow Strip-Based Test for Rapid and Ultrasensitive Detection of Spinal Muscular Atrophy. Biosensors 2021, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR-Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aman, R.; Mahas, A.; Marsic, T.; Hassan, N.; Mahfouz, M.M. Efficient, Rapid, and Sensitive Detection of Plant RNA Viruses With One-Pot RT-RPA-CRISPR/Cas12a Assay. Front. Microbiol. 2020, 11, 610872. [Google Scholar] [CrossRef] [PubMed]
- Alon, D.M.; Hak, H.; Bornstein, M.; Pines, G.; Spiegelman, Z. Differential Detection of the Tobamoviruses Tomato Mosaic Virus (ToMV) and Tomato Brown Rugose Fruit Virus (ToBRFV) Using CRISPR-Cas12a. Plants 2021, 10, 1256. [Google Scholar] [CrossRef]
- Mahas, A.; Hassan, N.; Aman, R.; Marsic, T.; Wang, Q.; Ali, Z.; Mahfouz, M.M. LAMP-Coupled CRISPR–Cas12a Module for Rapid and Sensitive Detection of Plant DNA Viruses. Viruses 2021, 13, 466. [Google Scholar] [CrossRef]
- Ramachandran, V.; Weiland, J.J.; Bolton, M.D. CRISPR-Based Isothermal Next-Generation Diagnostic Method for Virus Detection in Sugarbeet. Front. Microbiol. 2021, 12, 9994. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, M.; Duan, Y.-P.; Yang, Y. Highly sensitive and rapid detection of citrus Huanglongbing pathogen (Candidatus Liberibacter asiaticus) using Cas12a-based methods. Phytopathology 2021. [Google Scholar] [CrossRef]
- Zhang, Y.; Ying, J.Y. Homogeneous Immunochemical Assay on the Lateral Flow Strip for Measurement of DNase I Activity. Anal. Chem. 2015, 87, 10193–10198. [Google Scholar] [CrossRef]
- Gopal, K.; Lakshmi, L.M.; Sarada, G.; Nagalakshmi, T.; Sankar, T.G.; Gopi, V.; Ramana, K. Citrus melanose (Diaporthe citri Wolf): A review. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 113–124. [Google Scholar]
- Sreenivasaprasad, S.; Talhinhas, P. Genotypic and phenotypic diversity in Colletotrichum acutatum, a cosmopolitan pathogen causing anthracnose on a wide range of hosts. Mol. Plant Pathol. 2005, 6, 361–378. [Google Scholar] [CrossRef]
- Peever, T.; Su, G.; Carpenter-Boggs, L.; Timmer, L. Molecular systematics of citrus-associated Alternaria species. Mycologia 2004, 96, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Pers. Mol. Phylogeny Evol. Fungi 2018, 40, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawcett, H.S. Citrus diseases and their control. Citrus Dis. Control. 1936, 30, 308. [Google Scholar] [CrossRef]
- Ladanyia, M.; Ladaniya, M. Citrus Fruit: Biology, Technology and Evaluation; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Sun, L.; Nasrullah, K.F.; Nie, Z.; Wang, P.; Xu, J. Citrus Genetic Engineering for Disease Resistance: Past, Present and Future. Int. J. Mol. Sci. 2019, 20, 5256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riera, N.; Wang, H.; Li, Y.; Li, J.; Pelz-Stelinski, K.; Wang, N. Induced Systemic Resistance Against Citrus Canker Disease by Rhizobacteria. Phytopathology 2018, 108, 1038–1045. [Google Scholar] [CrossRef] [Green Version]
- Alferez, F.M.; Gerberich, K.M.; Li, J.L.; Zhang, Y.; Graham, J.H.; Mou, Z. Exogenous Nicotinamide Adenine Dinucleotide Induces Resistance to Citrus Canker in Citrus. Front. Plant Sci. 2018, 9, 1472. [Google Scholar] [CrossRef]
- Graham, J.H.; Myers, M.E. Soil Application of SAR Inducers Imidacloprid, Thiamethoxam, and Acibenzolar-S-Methyl for Citrus Canker Control in Young Grapefruit Trees. Plant Dis. 2011, 95, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Nirvan, R. Citrus scab and its control. In Annual Report of the Horticultural Research Institute; Horticultural Research Institute: Saharanpur, India, 1961; pp. 136–141. [Google Scholar]
- Ibrahim, Y.E.; Saleh, A.A.; Al-Saleh, M.A. Management of Asiatic Citrus Canker Under Field Conditions in Saudi Arabia Using Bacteriophages and Acibenzolar-S-Methyl. Plant Dis. 2017, 101, 761–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Kapur, S.; Singh, K. Management of citrus scab caused by Elsinoe fawcettii. Indian Phytopathol. 2000, 53, 461–467. [Google Scholar]
- El Kahlout, M.I.; Abu-Naser, S.S. An Expert System for Citrus Diseases Diagnosis. Int. J. Acad. Eng. Res. 2019, 3, 1–7. [Google Scholar]
- Abdulridha, J.; Batuman, O.; Ampatzidis, Y. UAV-based remote sensing technique to detect citrus canker disease utilizing hyperspectral imaging and machine learning. Remote Sens. 2019, 11, 1373. [Google Scholar] [CrossRef] [Green Version]
- Sharif, M.; Khan, M.A.; Iqbal, Z.; Azam, M.F.; Lali, M.I.U.; Javed, M.Y. Detection and classification of citrus diseases in agriculture based on optimized weighted segmentation and feature selection. Comput. Electron. Agric. 2018, 150, 220–234. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Length (bp) |
|---|---|---|
| EF ITS F1 | AACCAACTCTTGTCTTGTGAAACCTTTGCAGT | 32 |
| EF ITS F2 | CCGGGGGACCGAACCAACTCTTGTCTTGTGAAA | 33 |
| EF ITS F3 | ACTCCCCACCCTTTGCTGTTGCGAATCACGTTG | 33 |
| EF ITS R1 | AATACCAAGCGGGGCTTGATTGGTGAAATGAC | 32 |
| EF ITS R2 | GGGGCTTGATTGGTGAAATGACGCTCGAACAGG | 33 |
| crRNA | UAAUUUCUACUAAGUGUAGAUAACGCACAUGCGCCCCUUG | 40 |
| Leaves | Fruits | |||
|---|---|---|---|---|
| PCR Positive | PCR Negative | PCR Positive | PCR Negative | |
| RPA-CRISPR/Cas12a-LFA positive | 22 | 0 | 22 | 0 |
| RPA-CRISPR/Cas12a-LFA negative | 0 | 2 | 0 | 2 |
| PPA 1 (44 of 44 = 100%) | NPA 2 (4 of 4 = 100%) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, K.; Kwon, S.-H.; Lee, S.-C.; Moon, Y.-E. Sensitive and Rapid Detection of Citrus Scab Using an RPA-CRISPR/Cas12a System Combined with a Lateral Flow Assay. Plants 2021, 10, 2132. https://doi.org/10.3390/plants10102132
Shin K, Kwon S-H, Lee S-C, Moon Y-E. Sensitive and Rapid Detection of Citrus Scab Using an RPA-CRISPR/Cas12a System Combined with a Lateral Flow Assay. Plants. 2021; 10(10):2132. https://doi.org/10.3390/plants10102132
Chicago/Turabian StyleShin, Kihye, Soon-Hwa Kwon, Seong-Chan Lee, and Young-Eel Moon. 2021. "Sensitive and Rapid Detection of Citrus Scab Using an RPA-CRISPR/Cas12a System Combined with a Lateral Flow Assay" Plants 10, no. 10: 2132. https://doi.org/10.3390/plants10102132