
Neurogenin 2 and Neuronal Differentiation 1 Control Proper Development of the Chick Trigeminal Ganglion and Its Nerve Branches

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chicken Embryos
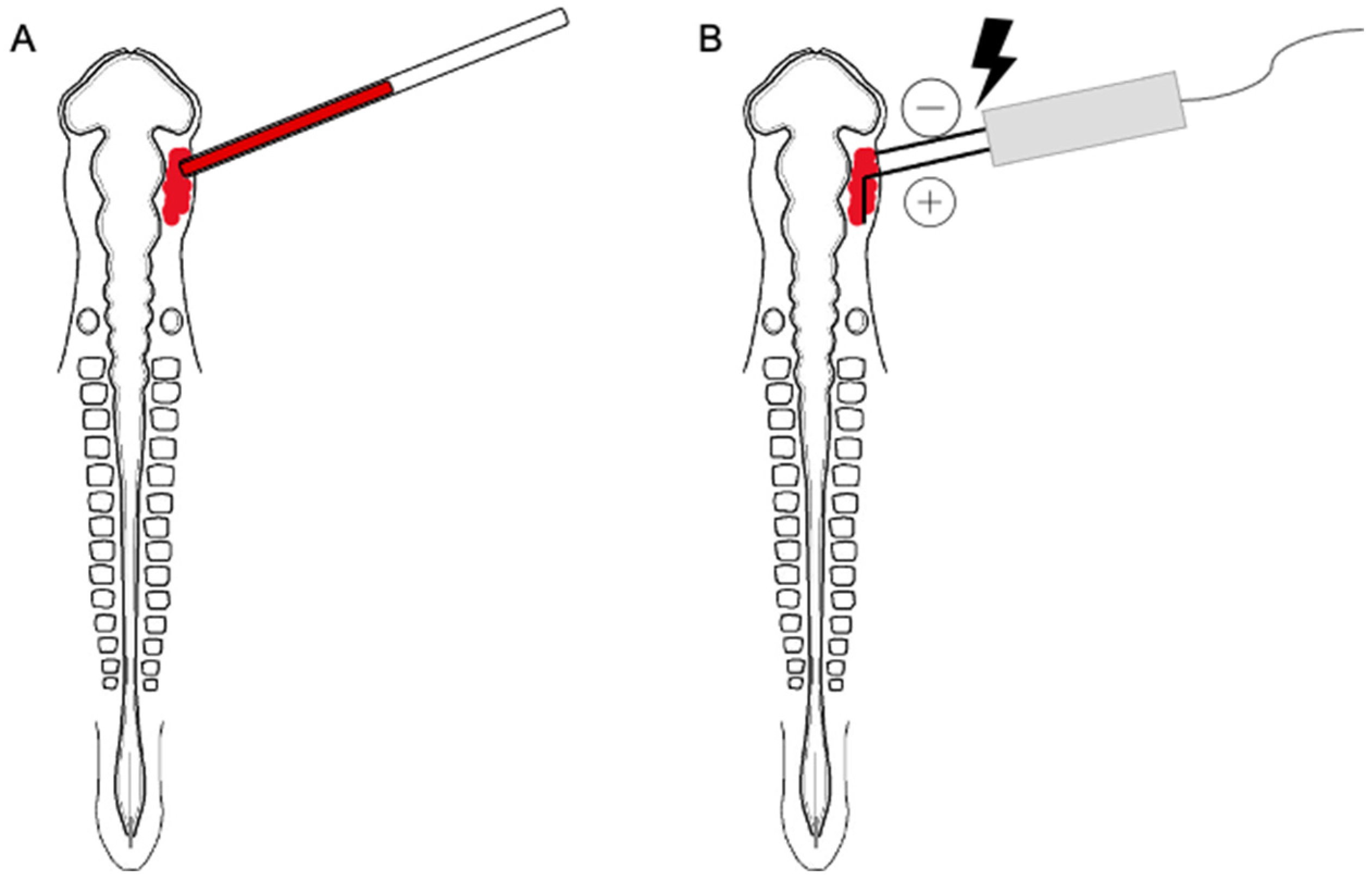
2.2. Morpholino Design and Electroporation
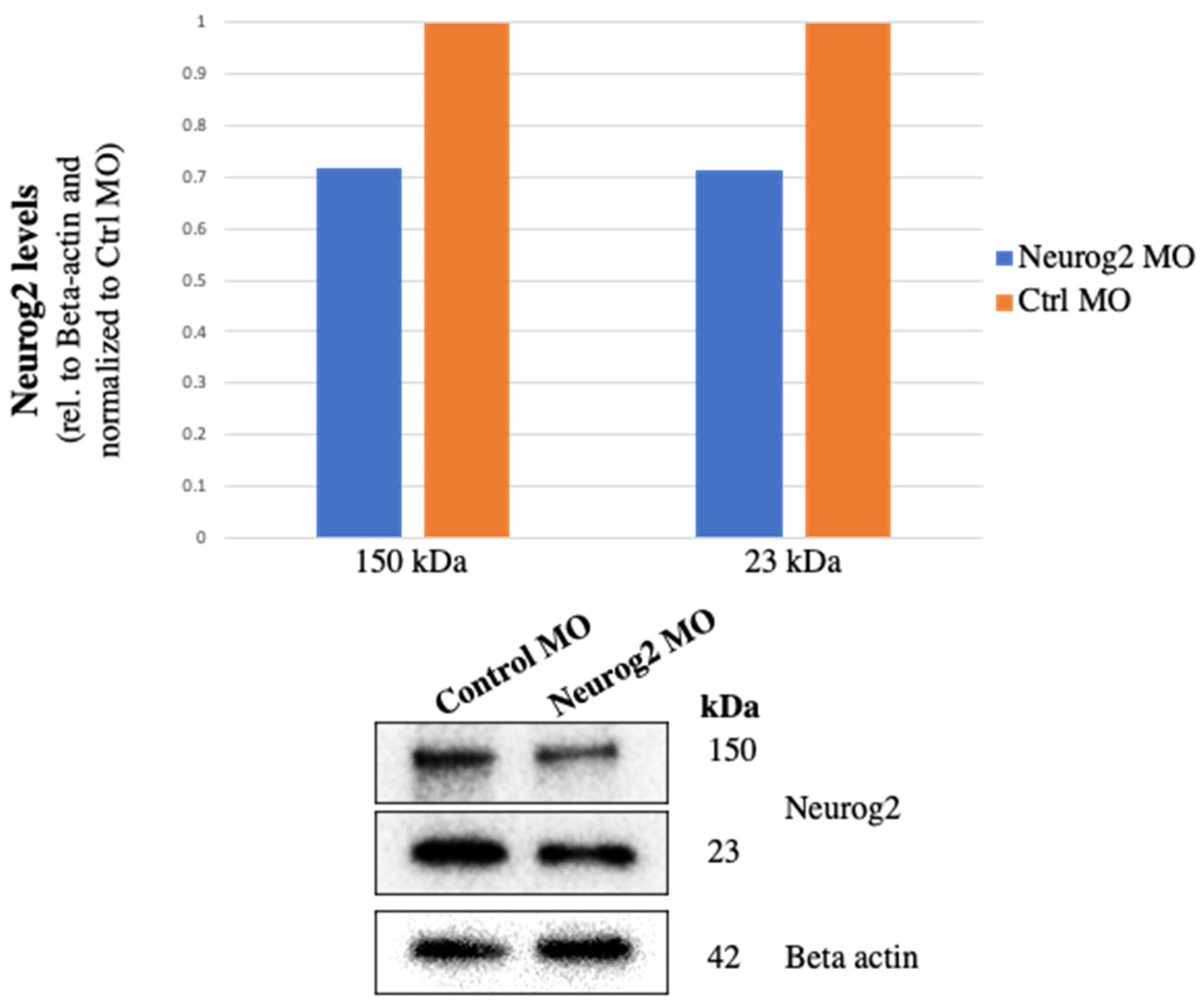
2.3. Immunoblotting
2.4. Whole-Mount Immunohistochemistry
2.5. Fructose and Urea Solution (FRUIT) Clearing
2.6. Confocal Imaging
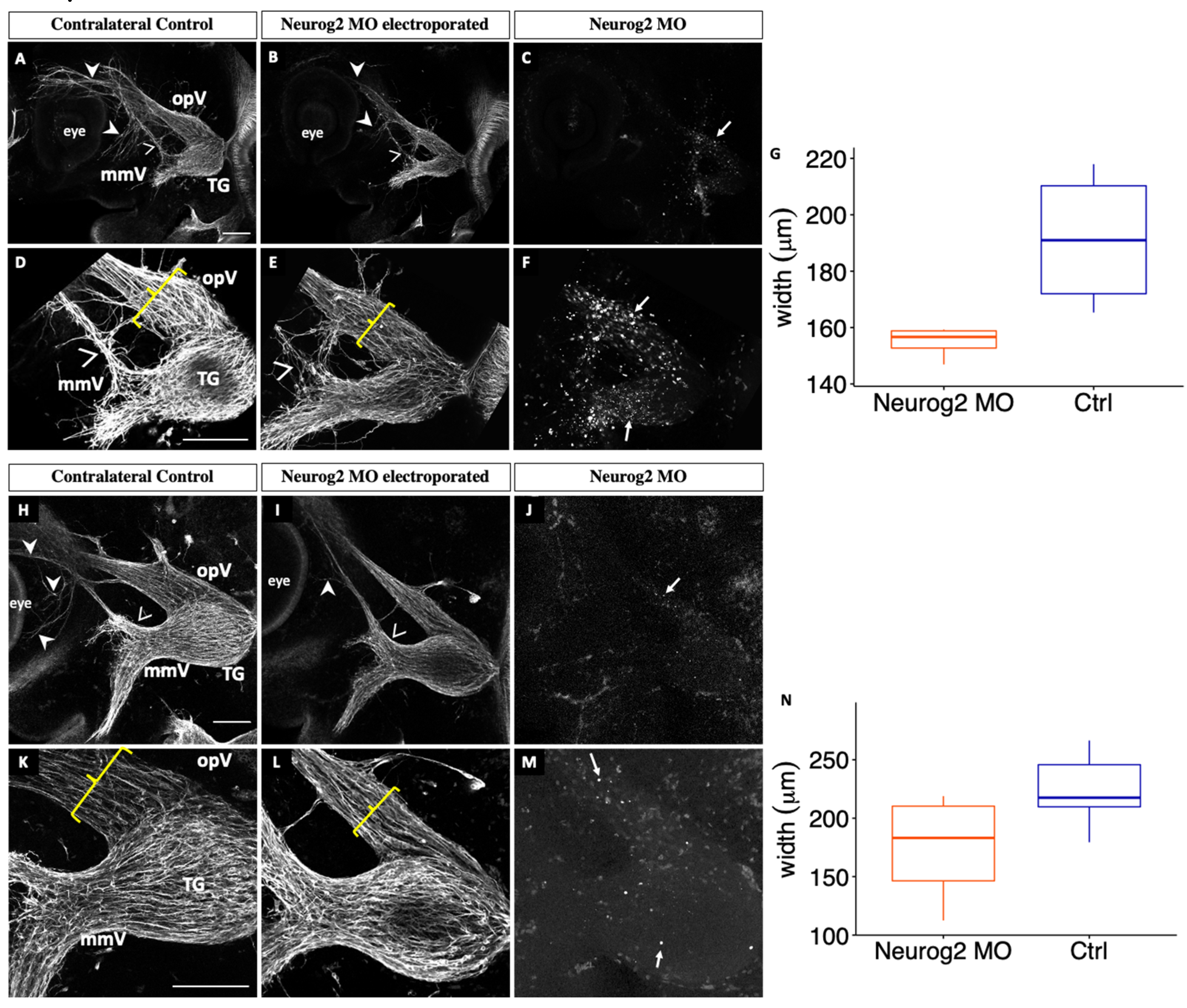
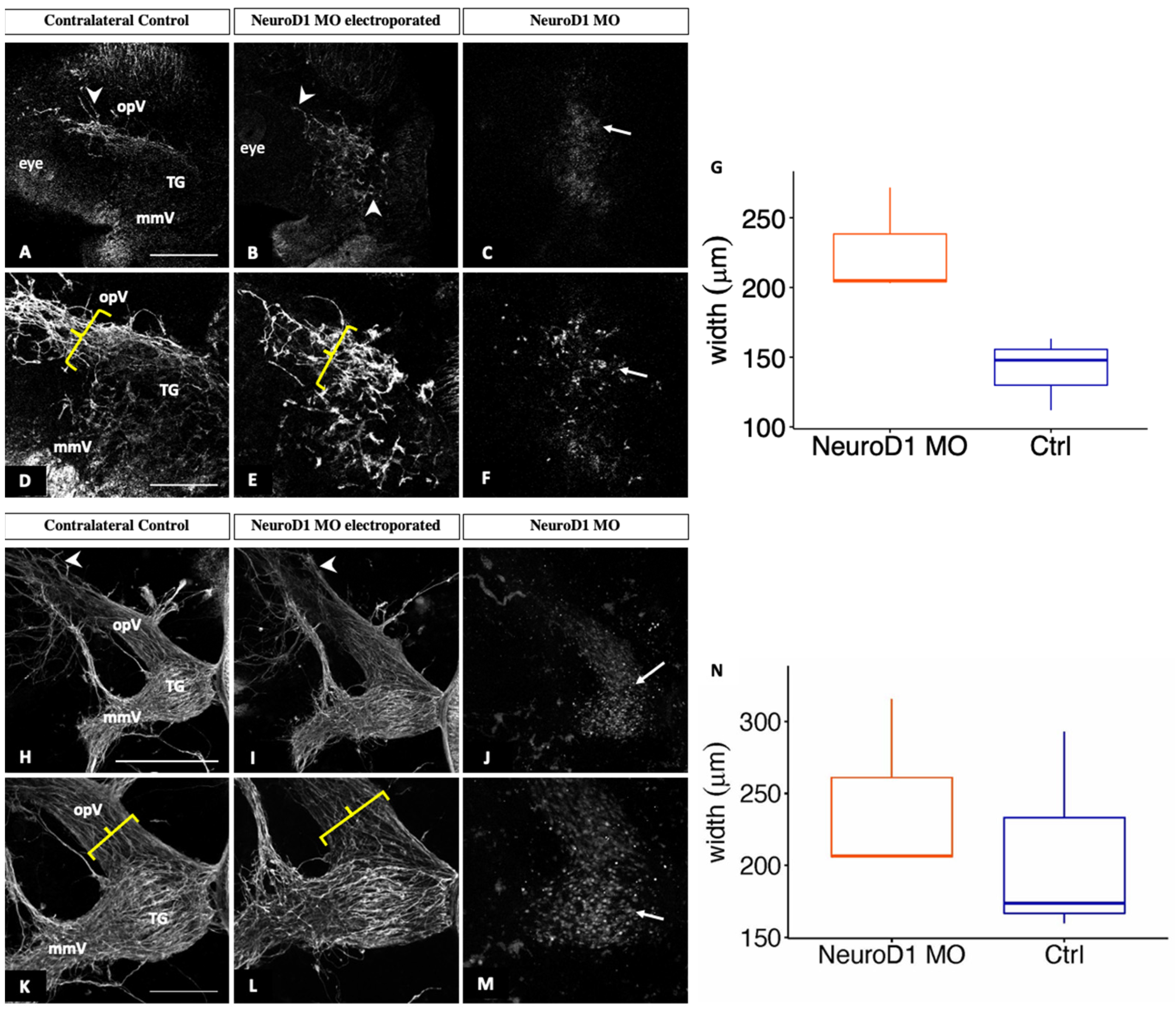
2.7. Measurement of Ophthalmic Branch Width
2.8. Statistical Analysis
3. Results
3.1. Neurog2 Controls the Proper Formation of the Trigeminal Ganglion and Its Nerve Branches
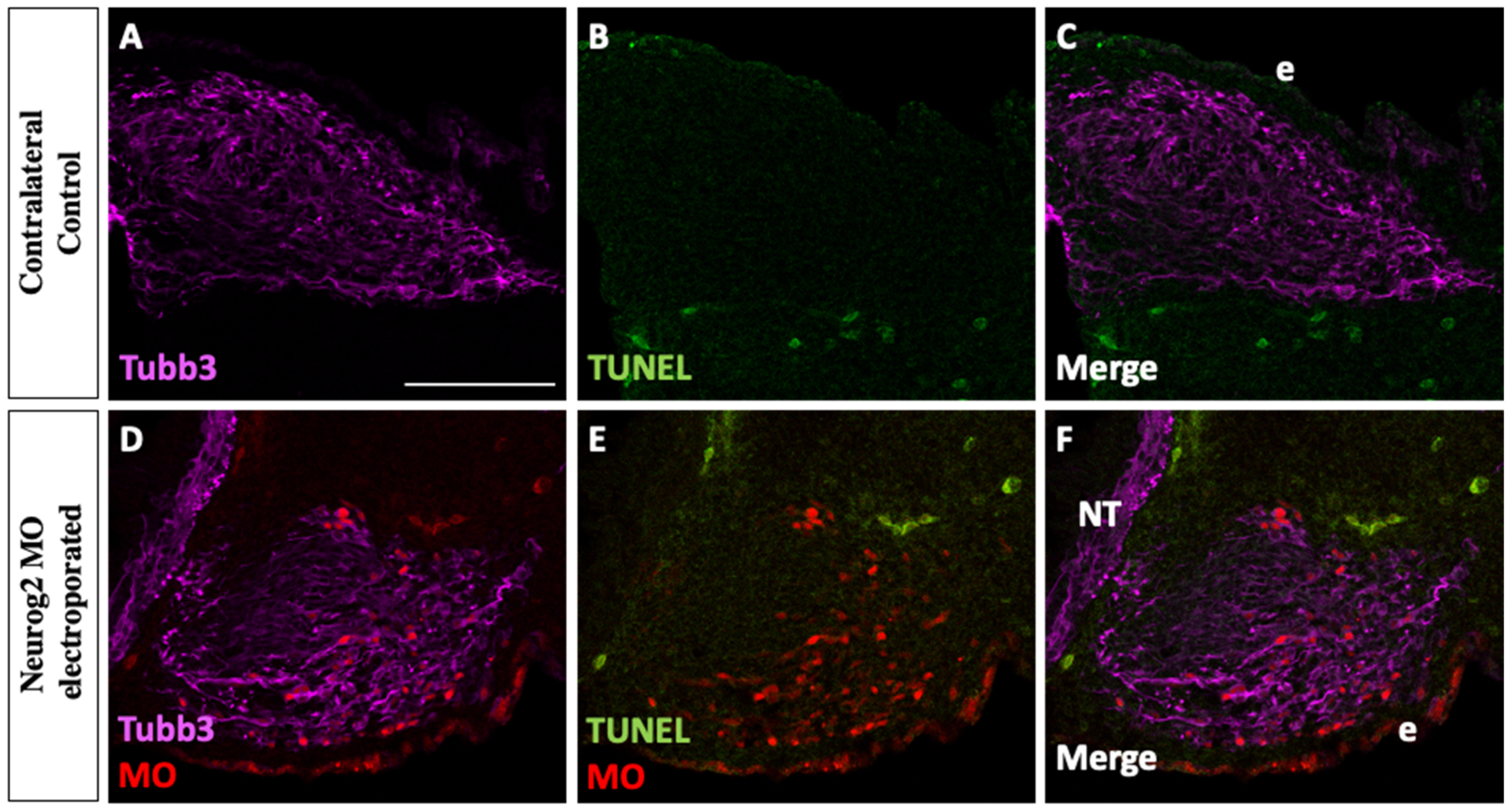
3.2. Neurog2 Depletion Does Not Cause Apoptosis of Trigeminal Placode Cells or Their Neuronal Derivatives during Trigeminal Ganglion Assembly
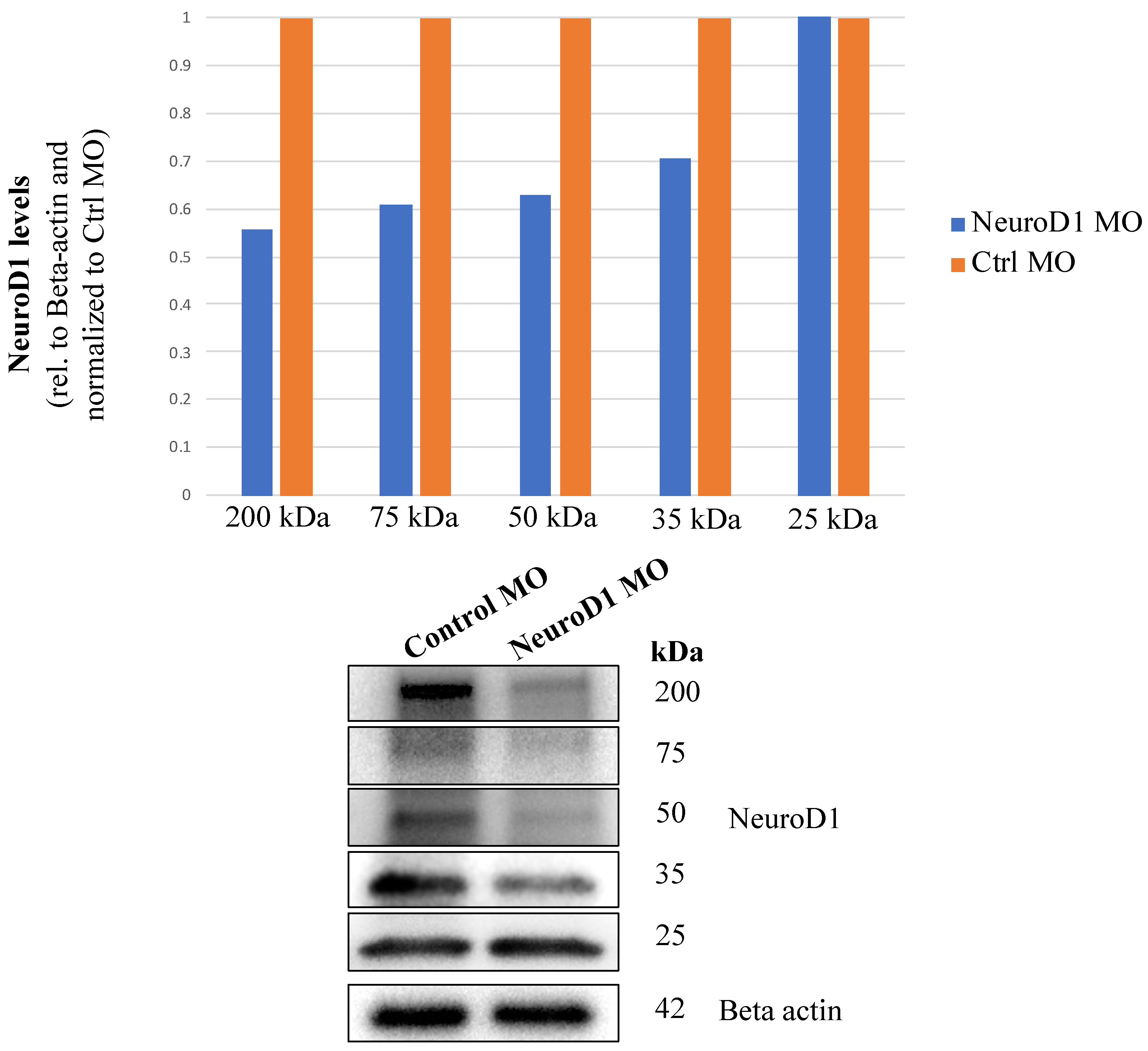
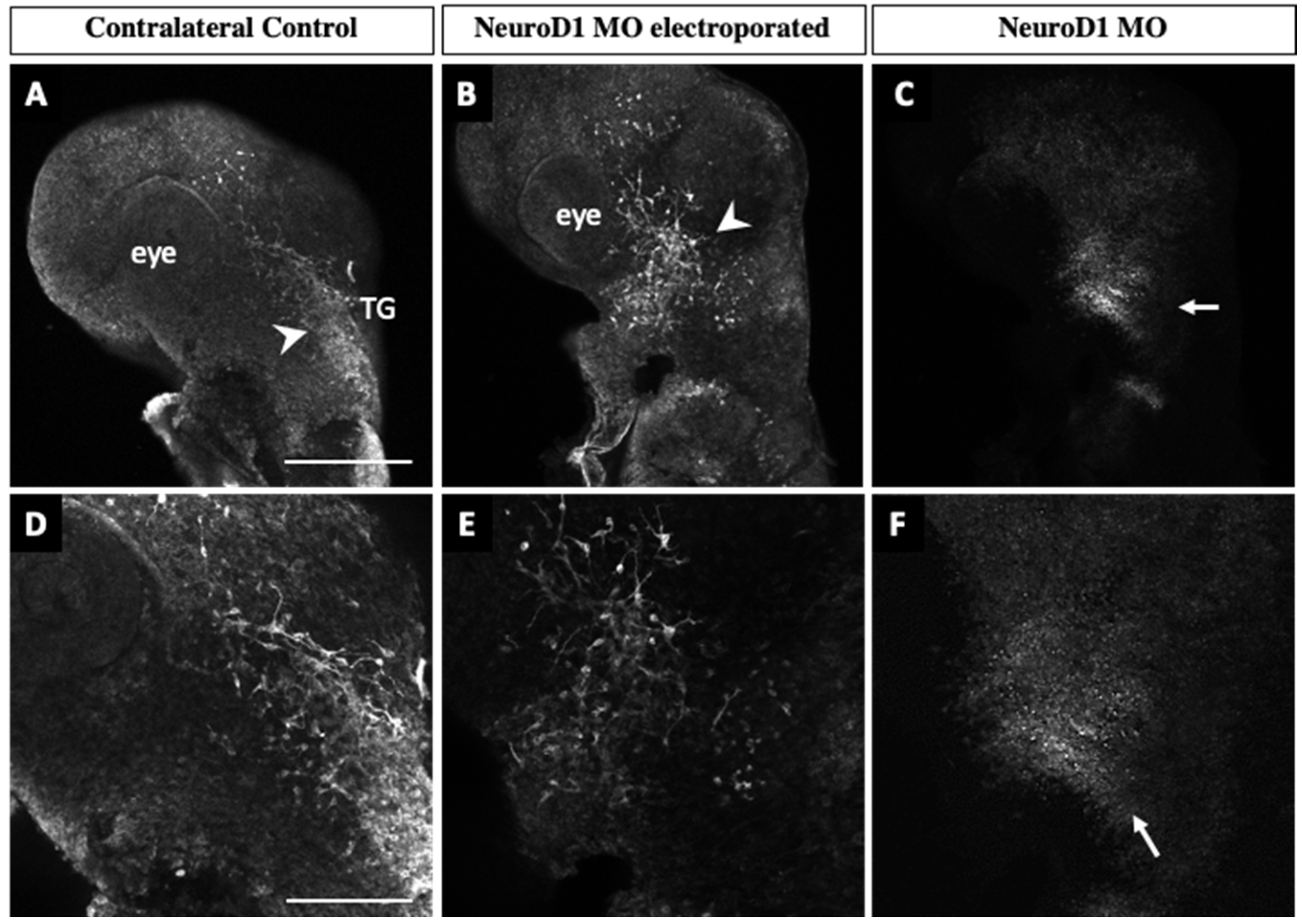
3.3. NeuroD1 Regulates Early Chick Trigeminal Ganglion Assembly
4. Discussion
4.1. Neurog2 Regulates Proper Development of the Trigeminal Ganglion, and Particularly the Forming Ophthalmic Branch
4.2. NeuroD1 Influences Trigeminal Ganglion Development
4.3. Possible Roles for Neurog2 and NeuroD1 in Trigeminal Gangliogenesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamburger, V. Experimental Analysis of the Dual Origin of the Trigeminal Ganglion in the Chick Embryo. J. Exp. Zool. 1961, 148, 91–123. [Google Scholar] [CrossRef] [PubMed]
- D’amico-Martel, A.; Noden, D.M. Contributions of Placodal and Neural Crest Cells to Avian Cranial Peripheral Ganglia. Am. J. Anat. 1983, 166, 445–468. [Google Scholar] [CrossRef] [PubMed]
- Shiau, C.E.; Lwigale, P.Y.; Das, R.M.; Wilson, S.A.; Bronner-Fraser, M. Robo2-Slit1 Dependent Cell-Cell Interactions Mediate Assembly of the Trigeminal Ganglion. Nat. Neurosci. 2008, 11, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Dude, C.M.; Baker, C.V.H. Fine-Grained Fate Maps for the Ophthalmic and Maxillomandibular Trigeminal Placodes in the Chick Embryo. Dev. Biol. 2008, 317, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.V.H.; Bronner-Fraser, M. Vertebrate Cranial Placodes I. Embryonic Induction. Dev. Biol. 2001, 232, 1–61. [Google Scholar] [CrossRef] [Green Version]
- McCabe, K.L.; Sechrist, J.W.; Bronner-Fraser, M. Birth of Ophthalmic Trigeminal Neurons Initiates Early in the Placodal Ectoderm. J. Comp. Neurol. 2009, 514, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Begbie, J.; Graham, A. Integration between the Epibranchial Placodes and the Hindbrain. Science 2001, 294, 595–598. [Google Scholar] [CrossRef]
- Freter, S.; Fleenor, S.J.; Freter, R.; Liu, K.J.; Begbie, J. Cranial Neural Crest Cells Form Corridors Prefiguring Sensory Neuroblast Migration. Development 2013, 140, 3595–3600. [Google Scholar] [CrossRef] [Green Version]
- Hardwick, L.J.A.; Philpott, A. Multi-Site Phosphorylation Regulates NeuroD4 Activity during Primary Neurogenesis: A Conserved Mechanism amongst Proneural Proteins. Neural. Dev. 2015, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Hollenberg, S.M.; Snider, L.; Turner, D.L.; Lipnick, N.; Weintraub, H. Conversion of Xenopus Ectoderm into Neurons by NeuroD, a Basic Helix-Loop-Helix Protein. Science 1995, 268, 836–844. [Google Scholar] [CrossRef]
- Huang, C.; Chan, J.A.; Schuurmans, C. Proneural BHLH Genes in Development and Disease. Curr. Top. Dev. Biol. 2014, 110, 75–127. [Google Scholar] [PubMed]
- Zirlinger, M.; Lo, L.; McMahon, J.; McMahon, A.P.; Anderson, D.J. Transient Expression of the BHLH Factor Neurogenin-2 Marks a Subpopulation of Neural Crest Cells Biased for a Sensory but Not a Neuronal Fate. Proc. Natl. Acad. Sci. USA 2002, 99, 8084–8089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Yan, R.-T.; Mao, W.; Wang, S.-Z. Neurogenin3 Promotes Early Retinal Neurogenesis. Mol. Cell. Neurosci. 2009, 40, 187–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Chen, Z.; del Barco Barrantes, I.; de La Pompa, J.L.; Anderson, D.J. Neurogenin1 Is Essential for the Determination of Neuronal Precursors for Proximal Cranial Sensory Ganglia. Neuron 1998, 20, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Fode, C.; Gradwohl, G.; Morin, X.; Dierich, A.; LeMeur, M.; Goridis, C.; Guillemot, F. The BHLH Protein NEUROGENIN 2 Is a Determination Factor for Epibranchial Placode–Derived Sensory Neurons. Neuron 1998, 20, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Abu-Elmagd, M.; Ishii, Y.; Cheung, M.; Rex, M.; le Rouedec, D.; Scotting, P.J. CSox3 Expression and Neurogenesis in the Epibranchial Placodes. Dev. Biol. 2001, 237, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, V.; Hamilton, H.L. A Series of Normal Stages in the Development of the Chick Embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Thummel, R.; Bailey, T.J.; Hyde, D.R. In Vivo Electroporation of Morpholinos into the Adult Zebrafish Retina. J. Vis. Exp. 2011, 58, 3603. [Google Scholar]
- Shah, A.; Schiffmacher, A.T.; Taneyhill, L.A. Annexin A6 Controls Neuronal Membrane Dynamics throughout Chick Cranial Sensory Gangliogenesis. Dev. Biol. 2017, 425, 85–99. [Google Scholar] [CrossRef]
- Hou, B.; Zhang, D.; Zhao, S.; Wei, M.; Yang, Z.; Wang, S.; Wang, J.; Zhang, X.; Liu, B.; Fan, L.; et al. Scalable and DiI-Compatible Optical Clearance of the Mammalian Brain. Front. Neuroanat. 2015, 9, 19. [Google Scholar] [CrossRef]
- Rasband, W.S.; ImageJ. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 15 July 2022).
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 18 July 2022).
- Koontz, A.; Urrutia, H.A.; Bronner, M.E. Making a Head: Neural Crest and Ectodermal Placodes in Cranial Sensory Development. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Begbie, J.; Ballivet, M.; Graham, A. Early Steps in the Production of Sensory Neurons by the Neurogenic Placodes. Mol. Cell. Neurosci. 2002, 21, 502–511. [Google Scholar] [CrossRef]
- Lee, T.; Cho, I.-S.; Bashyal, N.; Naya, F.J.; Tsai, M.-J.; Yoon, J.S.; Choi, J.-M.; Park, C.-H.; Kim, S.-S.; Suh-Kim, H. ERK Regulates NeuroD1-Mediated Neurite Outgrowth via Proteasomal Degradation. Exp. Neurobiol. 2020, 29, 189. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Ray, S.K.; Pan, N.; Haigh, J.; Fritzsch, B.; Leiter, A.B. Intestinal Neurod1 Expression Impairs Paneth Cell Differentiation and Promotes Enteroendocrine Lineage Specification. Sci. Rep. 2019, 9, 19489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Okamoto, I.; Kameyama, H.; Kudoh, S.; Saito, H.; Sanada, M.; Kudo, N.; Wakimoto, J.; Fujino, K.; Ikematsu, Y.; et al. Integrated Immunohistochemical Study on Small-Cell Carcinoma of the Lung Focusing on Transcription and Co-Transcription Factors. Diagnostics 2020, 10, 949. [Google Scholar] [CrossRef]
- Singh, A.; Mahesh, A.; Noack, F.; de Toledo, B.; Calegari, F.; Tiwari, V.K. Tcf12 and NeuroD1 Cooperatively Drive Neuronal Migration during Cortical Development. Development 2022, 149, dev200250. [Google Scholar] [CrossRef]
- Morimoto, Y.; Fushimi, A.; Yamashita, N.; Hagiwara, M.; Bhattacharya, A.; Cheng, J.; Frost, T.C.; Ahmad, R.; Daimon, T.; Huang, L.; et al. Addiction of Merkel Cell Carcinoma to MUC1-C Identifies a Potential New Target for Treatment. Oncogene 2022, 41, 3511–3523. [Google Scholar] [CrossRef]
- Gaudillière, B.; Konishi, Y.; de La Iglesia, N.; Yao, G.; Bonni, A. A CaMKII-NeuroD Signaling Pathway Specifies Dendritic Morphogenesis. Neuron 2004, 41, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.; Lim, J.-W.; Yellajoshyula, D.; Chang, L.-W.; Kroll, K.L. Neurogenin and NeuroD Direct Transcriptional Targets and Their Regulatory Enhancers. EMBO J. 2007, 26, 5093–5108. [Google Scholar] [CrossRef] [Green Version]
- Dent, E.W.; Gupton, S.L.; Gertler, F.B. The Growth Cone Cytoskeleton in Axon Outgrowth and Guidance. Cold Spring Harb. Perspect. Biol. 2011, 3, a001800. [Google Scholar] [CrossRef] [Green Version]
- Deinhardt, K.; Kim, T.; Spellman, D.S.; Mains, R.E.; Eipper, B.A.; Neubert, T.A.; Chao, M.v.; Hempstead, B.L. Neuronal Growth Cone Retraction Relies on Proneurotrophin Receptor Signaling through Rac. Sci. Signal 2011, 4, ra82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barde, Y.A. Neurotrophins: A Family of Proteins Supporting the Survival of Neurons. Prog. Clin. Biol. Res. 1994, 390, 45–56. [Google Scholar] [PubMed]
- Liebl, D.J.; Tessarollo, L.; Palko, M.E.; Parada, L.F. Absence of Sensory Neurons before Target Innervation in Brain-Derived Neurotrophic Factor-, Neurotrophin 3-, and TrkC-Deficient Embryonic Mice. J. Neurosci. 1997, 17, 9113–9121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digital Repository at the University of Maryland, Animal & Avian Sciences Research Works. Neurogenin 2 and Neuronal Differentiation 1 control proper development of the chick trigeminal ganglion and its nerve branches. Available online: http://hdl.handle.net/1903/29101 (accessed on 15 July 2022).







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bina, P.; Hines, M.A.; Sanyal, J.; Taneyhill, L.A. Neurogenin 2 and Neuronal Differentiation 1 Control Proper Development of the Chick Trigeminal Ganglion and Its Nerve Branches. J. Dev. Biol. 2023, 11, 8. https://doi.org/10.3390/jdb11010008
Bina P, Hines MA, Sanyal J, Taneyhill LA. Neurogenin 2 and Neuronal Differentiation 1 Control Proper Development of the Chick Trigeminal Ganglion and Its Nerve Branches. Journal of Developmental Biology. 2023; 11(1):8. https://doi.org/10.3390/jdb11010008
Chicago/Turabian StyleBina, Parinaz, Margaret A. Hines, Johena Sanyal, and Lisa A. Taneyhill. 2023. "Neurogenin 2 and Neuronal Differentiation 1 Control Proper Development of the Chick Trigeminal Ganglion and Its Nerve Branches" Journal of Developmental Biology 11, no. 1: 8. https://doi.org/10.3390/jdb11010008