
The Complex Bridge between Aquatic and Terrestrial Life: Skin Changes during Development of Amphibians
Abstract
:1. Introduction
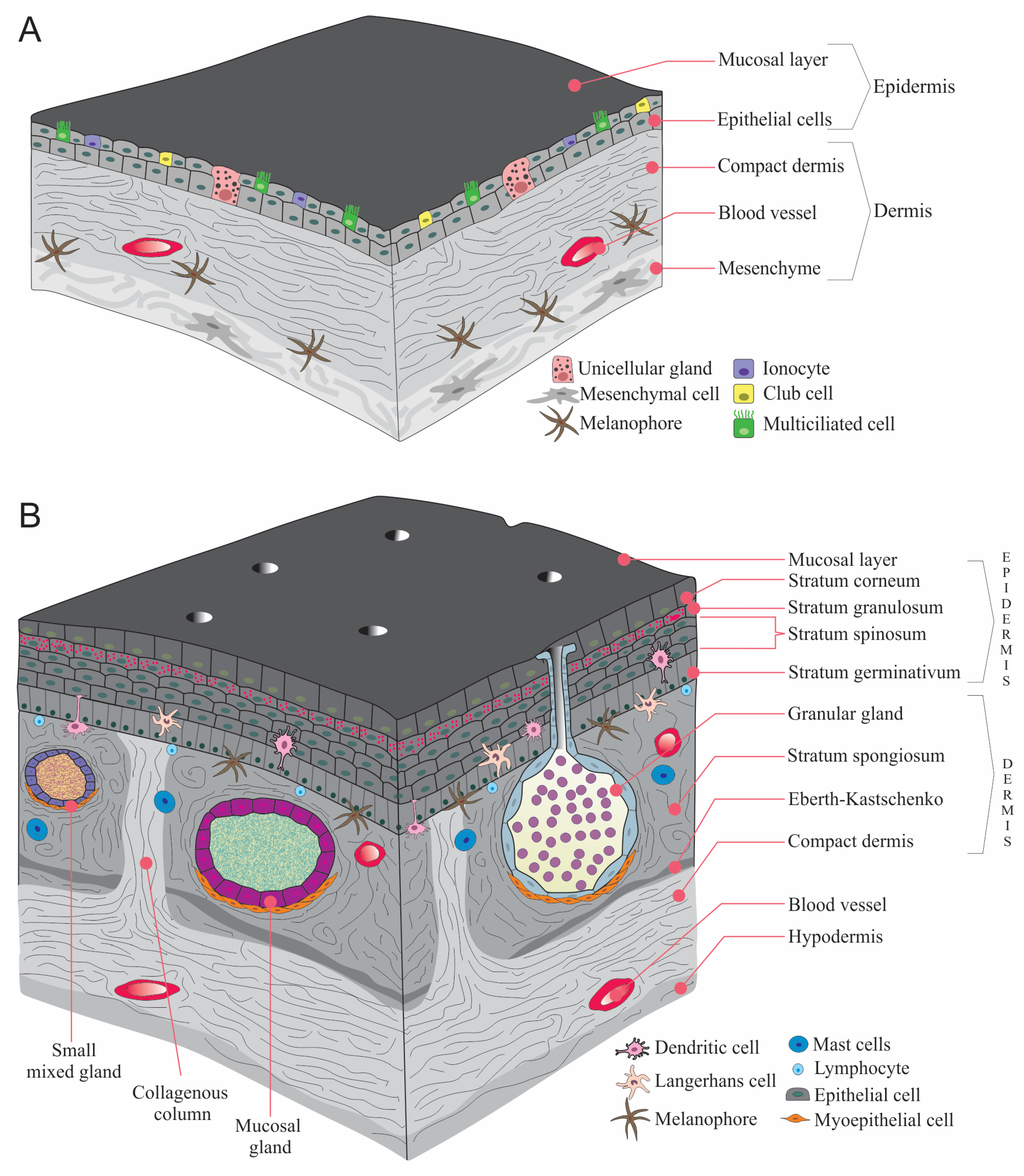
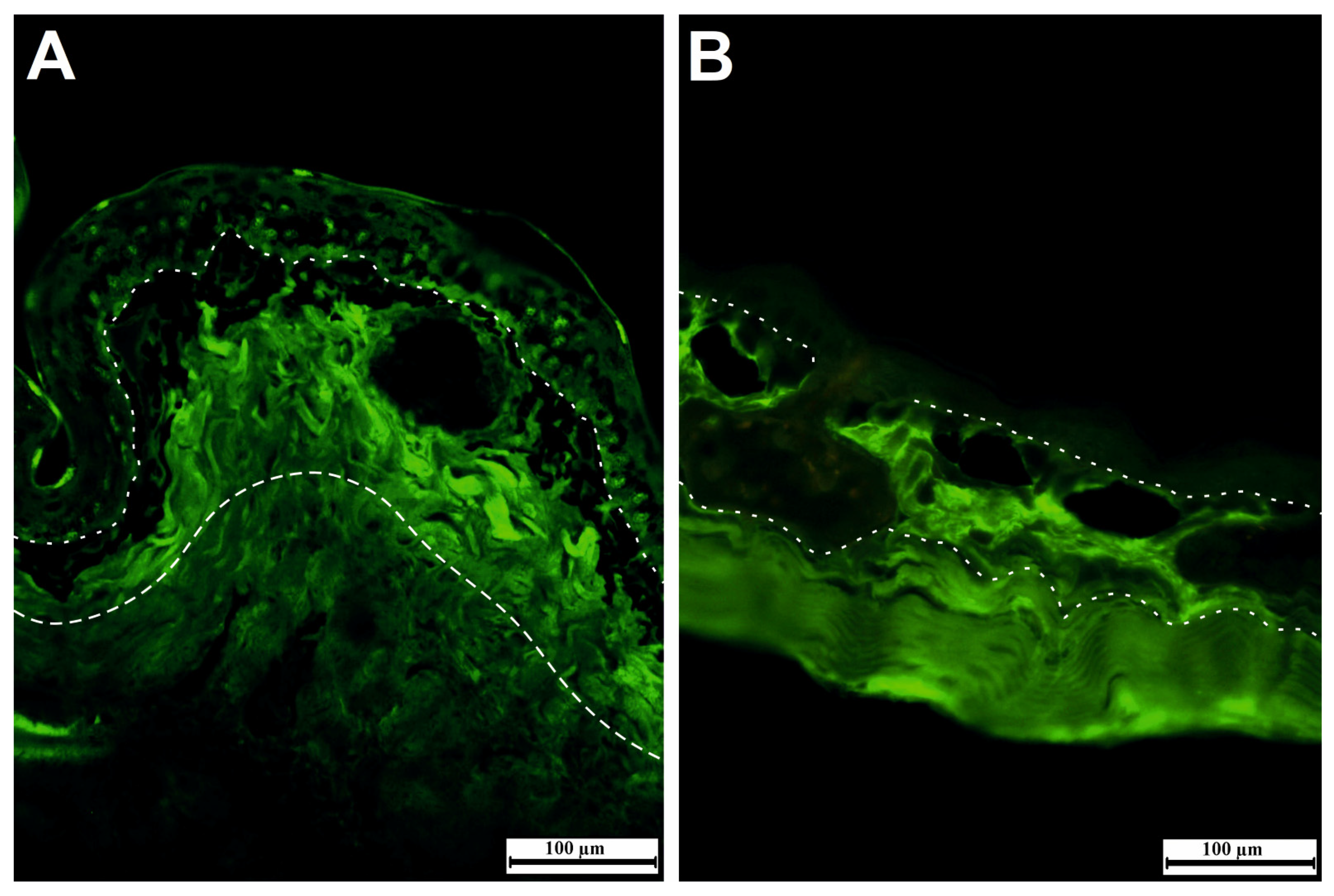
2. Skin Changes during the Transition from Larva to Adult
3. Chromatophores and Coloration
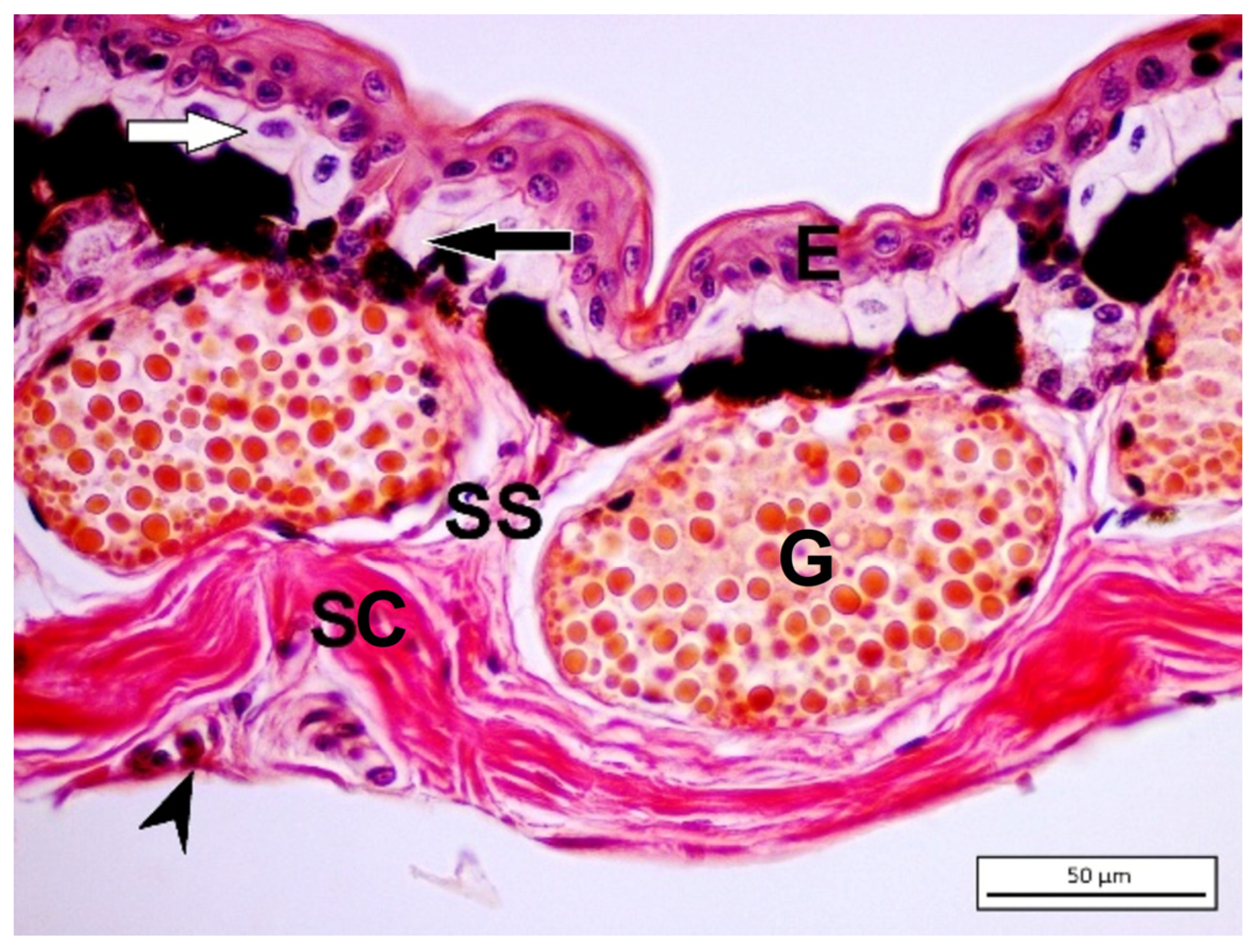
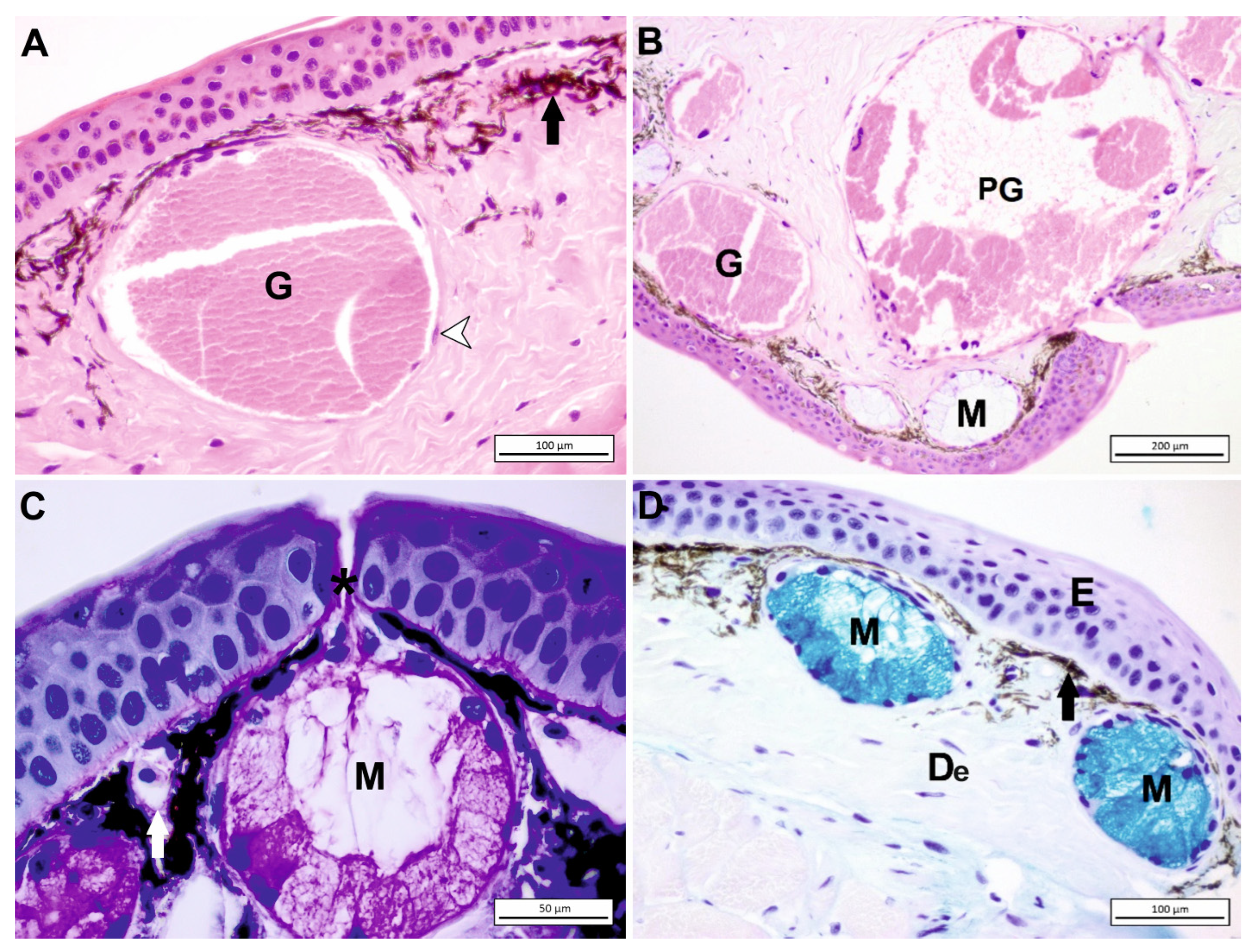
4. Amphibian Gland Formation, Types, and Function
5. Skin Immunology
6. Water Relations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blaustein, A.R.; Bancroft, B.A. Amphibian population declines: Evolutionary considerations. BioScience 2007, 57, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Cannatella, D.C.; Hillis, D.M. Amphibian relationships: Phylogenetic analysis of morphology and molecules. Herpetol. Monogr. 1993, 7, 1–7. [Google Scholar] [CrossRef]
- Cannatella, D.C.; Vieites, D.R.; Zhang, P.; Wake, M.H.; Wake, D.B. Amphibians (Lissamphibia). In The Timetree of Life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 354–356. [Google Scholar]
- Bossuyt, F.; Roelants, K. Frogs and Toads (Anura). In The Timetree of Life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 357–364. [Google Scholar]
- Vieties, D.R.; Zhang, P.; Wake, D.B. Salamanders (Caudata). In The Timetree of Life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 365–368. [Google Scholar]
- Gower, D.J.; Wilkinson, M. Caecilians (Gymnophiona). In The Timetree of Life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 369–372. [Google Scholar]
- Toledo, R.D.; Jared, C. Cutaneous granular glands and amphibian venoms. Comp. Biochem. Phys. A 1995, 111, 1–29. [Google Scholar] [CrossRef]
- Clarke, B.T. The natural history of amphibian skin secretions, their normal functioning and potential medical applications. Biol. Rev. 1997, 72, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Haslam, I.S.; Roubos, E.W.; Mangoni, M.L.; Yoshizato, K.; Vaudry, H.; Kloepper, J.E.; Pattwell, D.M.; Maderson, P.F.; Paus, R. From frog integument to human skin: Dermatological perspectives from frog skin biology. Biol. Rev. 2014, 89, 618–655. [Google Scholar] [CrossRef]
- Rakers, S.; Gebert, M.; Uppalapati, S.; Meyer, W.; Maderson, P.; Sell, A.F.; Kruse, C.; Paus, R. ‘Fish matters’: The relevance of fish skin biology to investigative dermatology. Exp. Dermatol. 2010, 19, 313–324. [Google Scholar] [CrossRef]
- Schempp, C.; Emde, M.; Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. JDDG J. Dtsch. Dermatol. Ges. 2009, 7, 750–757. [Google Scholar] [CrossRef]
- Quay, W.B. Integument and the environment glandular composition, function, and evolution. Am. Zool. 1972, 12, 95–108. [Google Scholar] [CrossRef]
- Gong, Y.; Zeng, Y.; Zheng, P.; Liao, X.; Xie, F. Structural and bio-functional assessment of the postaxillary gland in Nidirana pleuraden (Amphibia: Anura: Ranidae). Zool. Lett. 2020, 6, 7. [Google Scholar] [CrossRef]
- König, E.; Bininda-Emonds, O.R.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef]
- McNamara, M.E.; Rossi, V.; Slater, T.S.; Rogers, C.S.; Ducrest, A.L.; Dubey, S.; Roulin, A. Decoding the evolution of melanin in vertebrates. Trends Ecol. Evol. 2021, 36, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.; Candioti, F.V.; Akmentins, M.S. Direct-developing frogs: Ontogeny of Oreobates barituensis (Anura: Terrarana) and the development of a novel trait. Amphib.-Reptil. 2012, 33, 239–250. [Google Scholar] [CrossRef]
- Naumann, B.; Schweiger, S.; Hammel, J.U.; Müller, H. Parallel evolution of direct development in frogs-Skin and thyroid gland development in African Squeaker Frogs (Anura: Arthroleptidae: Arthroleptis). Dev. Dyn. 2021, 250, 584–600. [Google Scholar] [CrossRef] [PubMed]
- Ishizuya-Oka, A.; Hasebe, T.; Shi, Y.B. Apoptosis in amphibian organs during metamorphosis. Apoptosis 2010, 15, 350–364. [Google Scholar] [CrossRef] [Green Version]
- Lindemann, B.; Voute, C. Structure and functions of the epidermis. In Frog Neurobiology; Linas, R., Precht, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1976; pp. 169–210. [Google Scholar]
- Yoshizato, K. Death and transformation of larval cells during metamorphosis of anura. Dev. Growth Differ. 1992, 34, 607–612. [Google Scholar] [CrossRef]
- Potts, R.O.; Francoeur, M.L. Lipid biophysics of water loss through the skin. Proc. Natl. Acad. Sci. USA 1990, 87, 3871–3873. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Li, J.; Anboukaria, H.; Luo, Z.; Zhao, M.; Wu, H. Comparative transcriptome analyses of seven anurans reveal functions and adaptations of amphibian skin. Sci. Rep. 2016, 6, 24069. [Google Scholar] [CrossRef] [Green Version]
- Akat, E.; Yenmiş, M.; Pombal, M.A.; Molist, P.; Megías, M.; Arman, S.; Veselỳ, M.; Anderson, R.; Ayaz, D. Comparison of vertebrate skin structure at class level: A review. Anat. Rec. 2022, 305, 3543–3608. [Google Scholar] [CrossRef]
- Nishikawa, S.; Hirata, J.; Sasaki, F. Fate of ciliated epidermal cells during early development of Xenopus laevis using whole-mount immunostaining with an antibody against chondroitin 6-sulfate proteoglycan and anti-tubulin: Transdifferentiation or metaplasia of amphibian epidermis. Histochemistry 1992, 98, 355–358. [Google Scholar] [CrossRef]
- Nakajima, K.; Fujimoto, K.; Yaoita, Y. Programmed cell death during amphibian metamorphosis. Semin. Cell Dev. Biol. 2005, 16, 271–280. [Google Scholar] [CrossRef]
- Kelly, D.E. The Leydig cell in larval amphibian epidermis. Finestructure and function. Anat. Rec. 1966, 154, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Regueira, E.; Dávila, C.; Hermida, G.N. Morphological changes in skin glands during development in Rhinella arenarum (Anura: Bufonidae). Anat. Rec. 2016, 299, 141–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmura, H.; Wakahara, M. Transformation of skin from larval to adult types in normally metamorphosing and metamorphosis-arrested salamander, Hynobius retardatus. Differentiation 1998, 63, 237–246. [Google Scholar] [CrossRef]
- Faszewski, E.E.; Kaltenbach, J.C. Histology and lectin-binding patterns in the skin of the terrestrial horned frog Ceratophrys ornata. Cell Tissue Res. 1995, 281, 169–177. [Google Scholar] [CrossRef]
- McDiarmid, R.W.; Altig, R. Tadpoles: The Biology of Anuran Larvae, 1st ed.; University of Chicago Press: Chicago, IL, USA; London, UK, 1999. [Google Scholar]
- Marantelli, G.; Berger, L.; Speare, R.; Keegan, L. Distribution of the amphibian chytrid Batrachochytrium dendrobatidis and keratin during tadpole development. Pac. Conserv. Biol. 2004, 10, 173–179. [Google Scholar] [CrossRef]
- Larsen, E.H. Chloride transport by high-resistance heterocellular epithelia. Physiol. Rev. 1991, 71, 235–283. [Google Scholar] [CrossRef] [Green Version]
- Alibardi, L. Structural and immunocytochemical characterization of keratinization in vertebrate epidermis and epidermal derivatives. Int. Rev. Cytol. 2006, 253, 177–259. [Google Scholar]
- Zhang, Y.; Wang, Y.; Zeng, L.; Liu, Y.; Sun, H.; Li, S.; Wang, S.; Shu, L.; Liu, N.; Yin, S.; et al. Amphibian-derived peptide homodimer OA-GL17d promotes skin wound regeneration through the miR-663a/TGF-β1/Smad axis. Burns Trauma 2022, 10, tkac032. [Google Scholar] [CrossRef]
- Farquhar, M.G.; Palade, G.E. Cell junctions in amphibian skin. J. Cell Biol. 1965, 26, 263–291. [Google Scholar] [CrossRef] [Green Version]
- Voute, C.L.; Ussing, H.H. Some morphological aspects of active sodium transport. The epithelium of the frog skin. J. Cell Biol. 1968, 36, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Tanii, H.; Suzuki, M.; Tanaka, S. Regulation of water absorption in the frog skins by two vasotocin-dependent water-channel aquaporins, AQP-h2 and AQP-h3. Endocrinology 2003, 144, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.F.; Bui-Marinos, M.P.; Katzenback, B.A. Frog skin innate immune defences: Sensing and surviving pathogens. Front. Immunol. 2019, 9, 3128. [Google Scholar] [CrossRef] [Green Version]
- Kobelt, F.; Linsenmair, K.E. Adaptations of the reed frog Hyperolius viridiflavus (amphibia, anura, hyperoliidae) to its arid environment. I. The skin of Hyperolius viridiflavus nitidulus in wet and dry season conditions. Oecologia 1986, 68, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.C.; Jared, C. Cutaneous adaptations to water balance in amphibians. Comp. Biochem. Phys. A 1993, 105, 593–608. [Google Scholar] [CrossRef]
- Pandey, R.N.; Yaganti, S.; Coffey, S.; Frisbie, J.; Alnajjar, K.; Goldstein, D. Expression and immunolocalization of aquaporins HC-1, -2, and -3 in Cope’s gray treefrog, Hyla chrysoscelis. Comp. Bichem. Phys. A 2010, 157, 86–94. [Google Scholar] [CrossRef]
- Bagnara, J.T.; Taylor, J.D.; Hadley, M.E. The dermal chromatophore unit. J. Cell Biol. 1968, 38, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, S.K.; Epp, L.G.; Robinson, S.J. The pigmentary system of developing axolotls: I. A biochemical and structural analysis of chromatophores in wild-type axolotls. J. Embryol. Exp. Morphol. 1984, 81, 105–125. [Google Scholar]
- Polo-Cavia, N.; Gomez-Mestre, I. Pigmentation plasticity enhances crypsis in larval newts: Associated metabolic cost and background choice behaviour. Sci. Rep. 2017, 7, 39739. [Google Scholar] [CrossRef] [Green Version]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; The Johns Hopkins University Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Bagnara, J.T.; Matsumoto, J.; Ferris, W.; Frost, S.K.; Turner, W.A., Jr.; Tchen, T.T.; Taylor, J.D. Common origin of pigment cells. Science 1979, 203, 410–415. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: A comparative review. Pigment Cell Res. 2003, 16, 523–531. [Google Scholar] [CrossRef]
- Sugden, D.; Davidson, K.; Hough, K.A.; Teh, M.T. Melatonin, melatonin receptors and melanophores: A moving story. Pigment Cell Res. 2004, 17, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, C.; Cifuentes-Diaz, C.; Dunia, I.; Auber-Thomay, M.; Nicolas, P.; Amiche, M. Peptide secretion in the cutaneous glands of South American tree frog Phyllomedusa bicolor: An ultrastructural study. Eur. J. Cell Biol. 2000, 79, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Bell, S.C.; Bigler, L.; Caprioli, R.M.; Chaurand, P.; Lam, B.A.; Reinert, L.K.; Stalder, U.; Vazquez, V.M.; Schliep, K.; et al. Life history linked to immune investment in developing amphibians. Conserv. Physiol. 2016, 4, cow025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akat, E.; Arıkan, H. Morphology and biometric study of skin of Hyla orientalis Bedriaga, 1890 (Anura, Hylidae). Russ. J. Herpetol. 2013, 20, 253–258. [Google Scholar]
- Hauser, F.; Gertzen, E.-M.; Hoffmann, W. Expression of spasmolysin (FIM-A.1): An integumentary mucin from Xenopus laevis. Exp. Cell Res. 1990, 189, 157–162. [Google Scholar] [CrossRef]
- Stith, B.J. Biochemical examination of Rana pipiens epithelial mucus. J. Comp. Physiol. B 1984, 155, 89–96. [Google Scholar] [CrossRef]
- Voyles, J.; Berger, L.; Young, S.; Speare, R.; Webb, R.; Warner, J.; Rudd, D.; Campbell, R.; Skerratt, L.F. Electrolyte depletion and osmotic imbalance in amphibians with chytridiomycosis. Dis. Aquat. Organ. 2007, 77, 113–118. [Google Scholar] [CrossRef]
- Fisher, M.C.; Garner, T.W.; Walker, S.F. Global emergence of Batrachochytrium dendrobatidis and amphibian chytridiomycosis in space, time, and host. Annu. Rev. Microbiol. 2009, 63, 291–310. [Google Scholar] [CrossRef] [Green Version]
- Ohnuma, A.; Conlon, J.M.; Iwamuro, S. Differential expression of genes encoding preprobrevinin-2, prepropalustrin-2, and preproranatuerin-2 in developing larvae and adult tissues of the mountain brown frog Rana ornativentris. Comp. Biochem. Phys. C 2010, 151, 122–130. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Casciaro, B. Development of antimicrobial peptides from Amphibians. Antibiotics 2020, 9, 772. [Google Scholar] [CrossRef]
- Sang, Y.; Blecha, F. Antimicrobial peptides and bacteriocins: Alternatives to traditional antibiotics. Anim. Health Res. Rev. 2008, 9, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schadich, E.; Cole, A.L.; Squire, M.; Mason, D. Skin peptides of different life stages of Ewing’s tree frog. J. Exp. Zool. Part A 2010, 313, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Wabnitz, P.A.; Walters, H.; Tyler, M.J.; Wallace, J.C.; Bowie, J.H. First record of host defence peptides in tadpoles. The magnificent tree frog Litoria splendida. J. Pept. Res. 1998, 52, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Santana, C.J.C.; Magalhães, A.C.M.; dos Santos Júnior, A.C.; Ricart, C.A.O.; Lima, B.D.; Álvares, A.D.C.M.; Freitas, S.M.D.; Pires, O.R., Jr.; Fontes, W.; Castro, M.S. Figainin 1, a novel amphibian skin peptide with antimicrobial and antiproliferative properties. Antibiotics 2020, 9, 625. [Google Scholar] [CrossRef]
- Matsushita, O.; Okabe, A. Clostridial hydrolytic enzymes degrading extracellular components. Toxicon 2001, 39, 1769–1780. [Google Scholar] [CrossRef]
- Jedrzejas, M.J.; Mello, L.V.; De Groot, B.L.; Li, S. Mechanism of hyaluronan degradation by Streptococcus pneumoniae hyaluronate lyase structures of complexes with the substrate. J. Biol. Chem. 2002, 277, 28287–28297. [Google Scholar] [CrossRef] [Green Version]
- Necas, J.B.L.B.P.; Bartosikova, L.; Brauner, P.; Kolar, J.J.V.M. Hyaluronic acid (hyaluronan): A review. Vet. Med. 2008, 53, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Ibberson, C.B.; Jones, C.L.; Singh, S.; Wise, M.C.; Hart, M.E.; Zurawski, D.V.; Horswill, A.R. Staphylococcus aureus hyaluronidase is a CodY-regulated virulence factor. Infect. Immun. 2014, 82, 4253–4264. [Google Scholar] [CrossRef] [Green Version]
- Tracy, C.R.; Christian, K.A.; Tracy, C.R. Not just small, wet, and cold: Effects of body size and skin resistance on thermoregulation and arboreality of frogs. Ecology 2010, 91, 1477–1484. [Google Scholar] [CrossRef]
- Barbeau, T.R.; Lillywhite, H.B. Body wiping behaviors associated with cutaneous lipids in hylid tree frogs of Florida. J. Exp. Biol. 2005, 208, 2147–2156. [Google Scholar] [CrossRef] [Green Version]
- Verkman, A.S. Aquaporins at a glance. J. Cell Sci. 2011, 124, 2107–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.; Hasegawa, T.; Ogushi, Y.; Tanaka, S. Amphibian aquaporins and adaptation to terrestrial environments: A review. Comp. Biochem. Phys. A 2007, 148, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.A.; Carvalho, H.F.; de Brito-Gitirana, L. Hyaluronan in the epidermal and the dermal extracellular matrix: Its role in cutaneous hydric balance and integrity of anuran integument. Micron 2007, 38, 607–610. [Google Scholar] [CrossRef]
- Akat, E.; Arikan, H.; Göçmen, B. Investigation of dorsal/ventral skin and the parotoid region of Lyciasalamandra billae and Lyciasalamandra luschani basoglui (Urodela: Salamandridae). Biologia 2014, 69, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Croteau, M.C.; Davidson, M.A.; Lean, D.R.S.; Trudeau, V.L. Global increases in ultraviolet B radiation: Potential impacts on amphibian development and metamorphosis. Physiol. Biochem. Zool. 2008, 81, 743–761. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Ordo, Familia | Research Topic |
|---|---|---|
| Rhinella arenarum | (Anura, Bufonidae) | Development of skin |
| Arthroleptis sp. | (Anura, Arthroleptidae) | Development of skin |
| Cardioglossa sp. | (Anura, Arthroleptidae) | Development of skin |
| Hynobius retardatus | (Urodela, Hynobiidae) | Development of skin |
| Hyla savignyi | (Anura, Hylidae) | Skin histomorphology |
| Salamandra infraimmaculata | (Salamandridae, Salamandridae) | Skin histomorphology |
| Hyperolius nasutus | (Anura, Hyperoliidae) | Chromatophores in the skin |
| Hyperolius viridiflavus | (Anura, Hyperoliidae) | Skin adaptation to wet and dry season conditions |
| Dryophytes chrysoscelis (formerly Hyla chrysoscelis) | (Anura, Hylidae) | Expression and immunolocalization of aquaporins |
| Chiromantis petersi | (Anura, Rhacophoridae) | Dermal chromatophores |
| Xenopus laevis | (Anura, Pipidae) | Effects of melatonin during metamorphosis, antimicrobial peptides, aquaporins |
| Phyllomedusa bicolor | (Anura, Hylidae) | Dermal glands |
| Bufo sp. | (Anura, Bufonidae) | Dermal glands |
| Lithobates pipiens | (Anura, Ranidae) | Biochemical and function of epithelial mucus |
| Ranoidea splendida (formerly Litoria splendida) | (Anura, Hylidae) | Host-defense peptides of the skin |
| Odorrana margaretae | (Anura, Ranidae) | Antimicrobial peptides of different life stages |
| Pelophylax nigromaculatus | (Anura, Ranidae) | Antimicrobial peptides of different life stages |
| Xenopus tropicalis | (Anura, Pipidae) | The development of the mucosal barrier |
| Hyla japonica | (Anura, Hylidae) | Aquaporins |
| Rana japonica | (Anura, Ranidae) | Aquaporins |
| Rana nigromaculata | (Anura, Ranidae) | Aquaporins |
| Rana catesbeiana | (Anura, Ranidae) | Aquaporins |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akat Çömden, E.; Yenmiş, M.; Çakır, B. The Complex Bridge between Aquatic and Terrestrial Life: Skin Changes during Development of Amphibians. J. Dev. Biol. 2023, 11, 6. https://doi.org/10.3390/jdb11010006
Akat Çömden E, Yenmiş M, Çakır B. The Complex Bridge between Aquatic and Terrestrial Life: Skin Changes during Development of Amphibians. Journal of Developmental Biology. 2023; 11(1):6. https://doi.org/10.3390/jdb11010006
Chicago/Turabian StyleAkat Çömden, Esra, Melodi Yenmiş, and Berna Çakır. 2023. "The Complex Bridge between Aquatic and Terrestrial Life: Skin Changes during Development of Amphibians" Journal of Developmental Biology 11, no. 1: 6. https://doi.org/10.3390/jdb11010006