
Advances in Understanding the Genetic Mechanisms of Zebrafish Renal Multiciliated Cell Development
 ,
, 
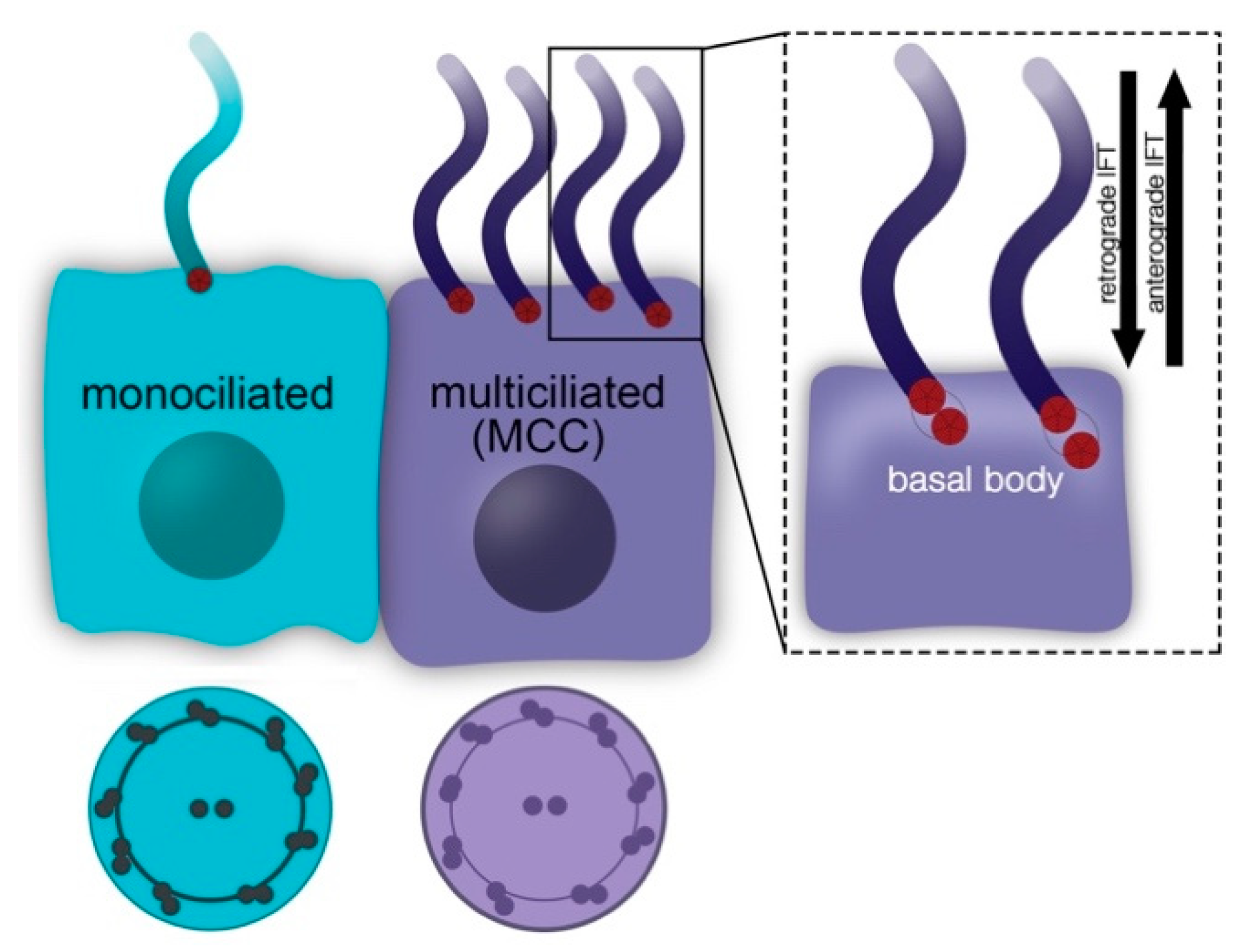{kind=link}
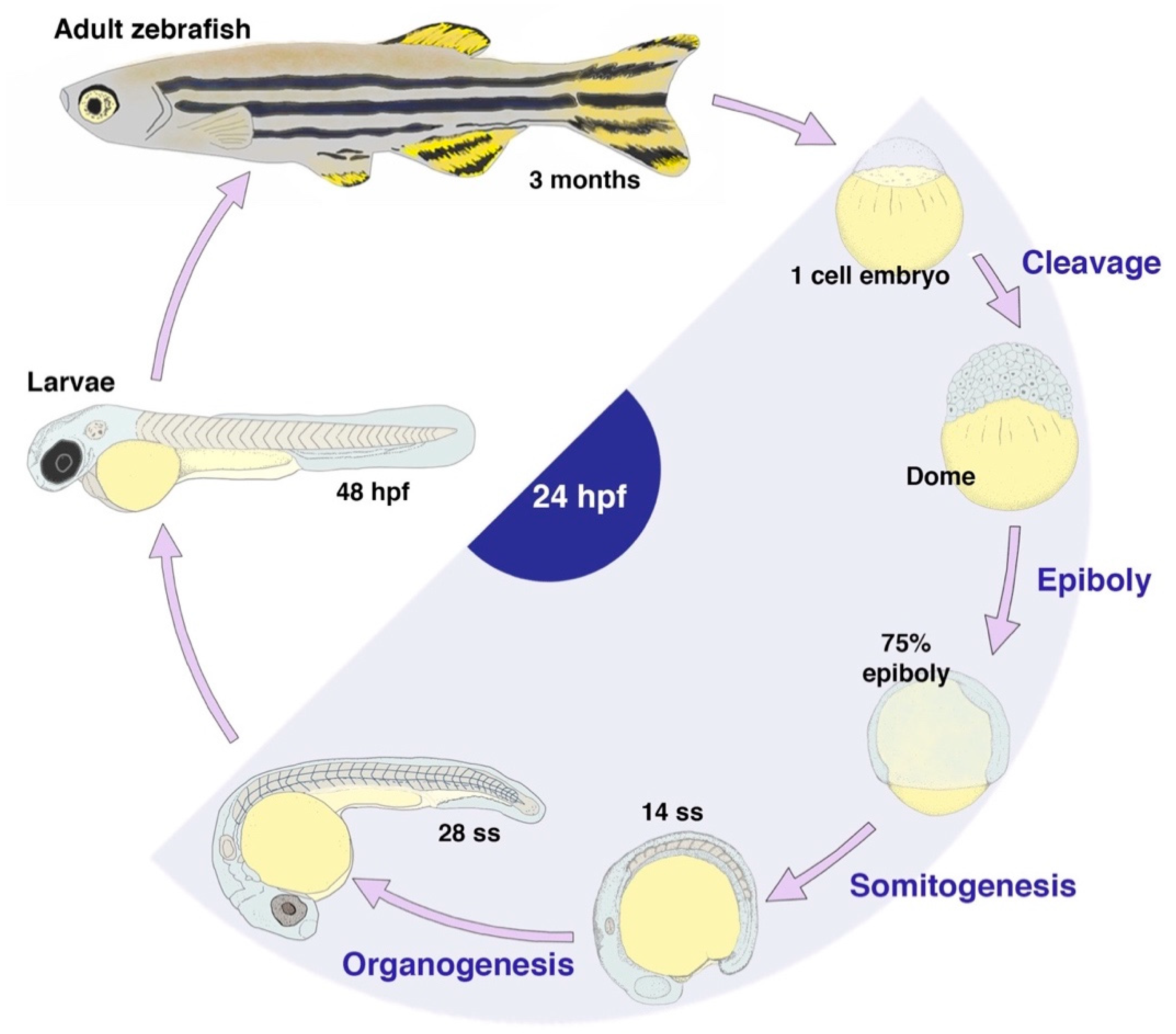{kind=link}
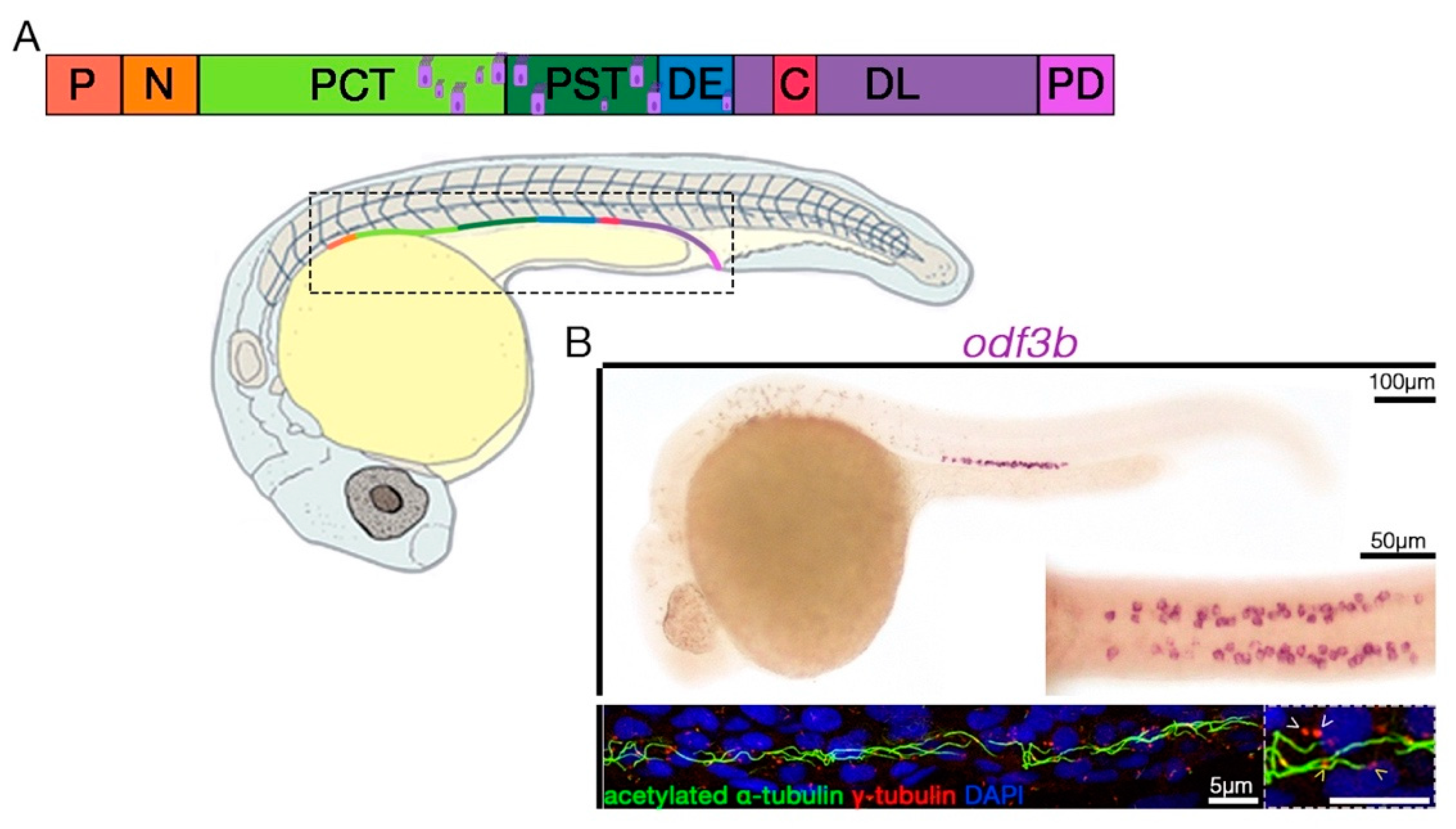{kind=link}
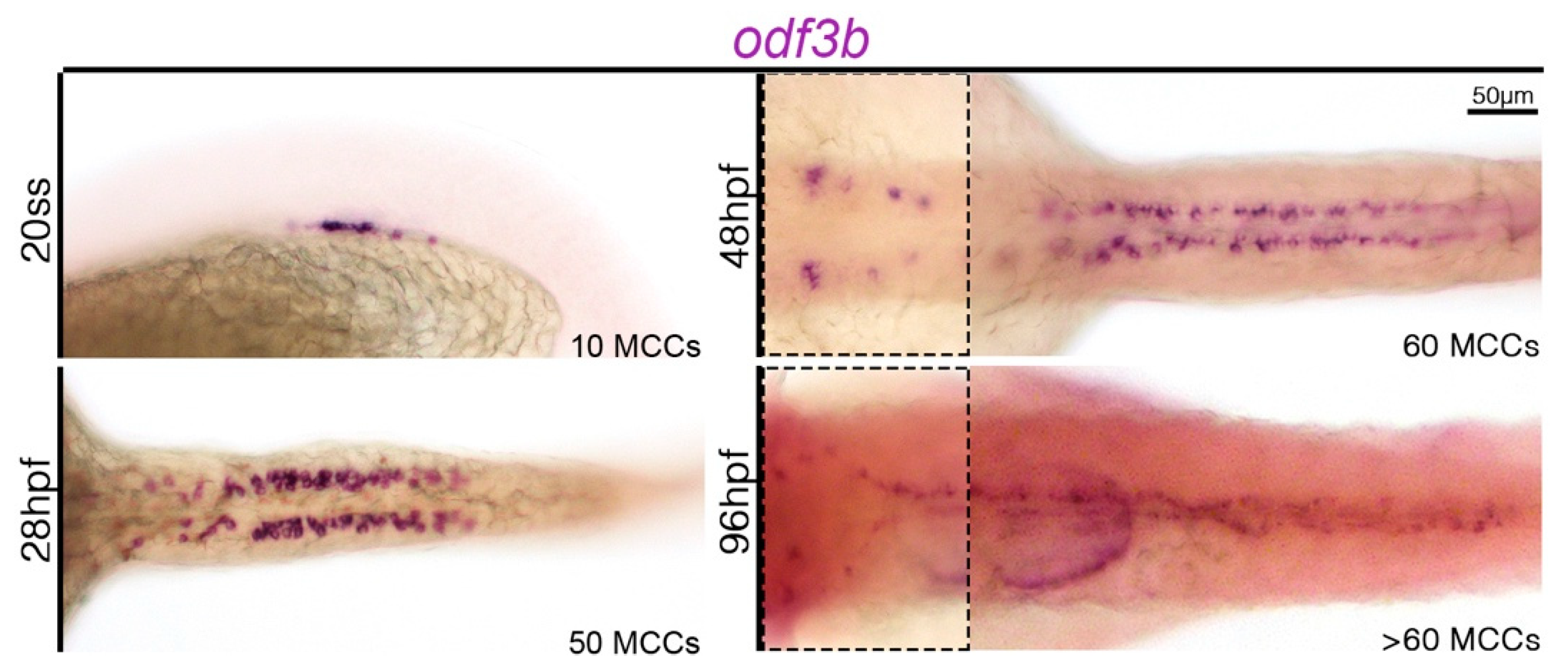{kind=link}
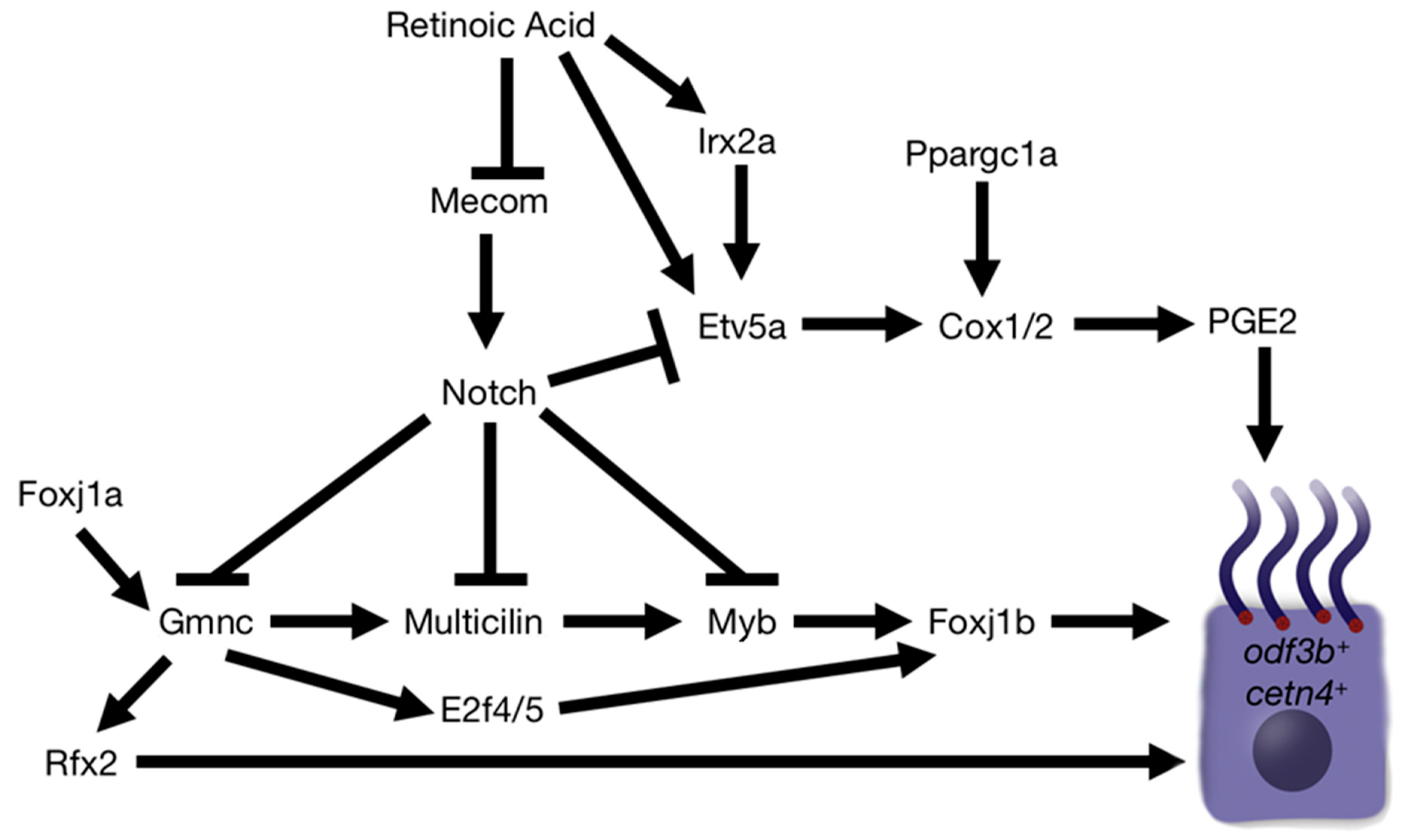{kind=link}
Abstract
:1. Introduction
2. The Zebrafish Pronephros Model of MCC Ontogeny
3. The Role of Notch Signaling in MCC Fate Choice Is Highly Conserved
4. Identification of Other Key Signaling Pathways and Transcriptional Components of the MCC Genetic Regulatory Network
4.1. Notch Is Positively Regulated by the Mecom Transcription Factor
4.2. Retinoic Acid (RA) Acts Upstream of Mecom Notch Signaling to Promote MCC Fate
4.3. Candidate Notch Targets: ETS Transcription Factors Etv5a/4
4.4. The Iroquois (irx) Transcription Factor irx2a
4.5. Prostaglandin Signaling Regulates MCC Specification and Differentiation
4.6. Modulation of Prostanoid Biosynthesis by ppargc1a
5. Swimming Ahead: Prospects and Challenges for Future Studies of MCC Development in Zebrafish
5.1. Expanding the Toolkit to Study Renal MCCs
5.2. Assembling MCC Genetic Regulatory Network(s) and Connecting the Dot(s) across Species
5.3. If and How MCCs Are Relevant to Human Kidney Disease States
5.4. Do (Renal) MCCs Regenerate?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishikawa, T.; Ueno, H.; Omori, T.; Kikuchi, K. Cilia and centrosomes: Ultrastructural and mechanical perspectives. Semin. Cell Dev. Biol. 2020, 110, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Christensen, S.T. Overview of structure and function of mammalian cilia. Annu. Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fliegauf, M.; Benzing, T.; Omran, H. When cilia go bad: Cilia defects and ciliopathies. Nat. Rev. Mol. Cell Biol. 2007, 8, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Pazour, G.J.; Quarmby, L.; Smith, A.O.; Desai, P.B.; Schmidts, M. Cilia in cystic kidney and other diseases. Cell Signal. 2020, 69, 109519. [Google Scholar] [CrossRef] [PubMed]
- Wallmeier, J.; Nielsen, K.G.; Kuehni, C.E.; Lucas, J.S.; Leigh, M.W.; Zariwala, M.A.; Omran, H. Motile ciliopathies. Nat. Rev. Dis. Prim. 2020, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.R.; Wallingford, J.B. Multiciliated cells. Curr. Biol. 2014, 24, R973–R982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutin, C.; Kodjabachian, L. Biology of multiciliated cells. Curr. Opin. Genet. Dev. 2019, 56, 1–7. [Google Scholar] [CrossRef]
- Lewis, M.; Stracker, T.H. Transcriptional regulation of multiciliated cell differentiation. Semin. Cell Dev. Biol. 2021, 110, 51–60. [Google Scholar] [CrossRef]
- Walentek, P. Signaling control of mucociliary epithelia: Stem cells, cell fates, and the plasticity of cell identity in development and disease. Cells Tissues Organs 2021, 211, 1–18. [Google Scholar] [CrossRef]
- Mahjoub, M.R.; Nanjundappa, R.; Harvey, M.N. Development of a multiciliated cell. Curr. Opin. Cell Biol. 2022, 77, 10215. [Google Scholar] [CrossRef]
- Meunier, A.; Azimzadeh, J. Multiciliated cells in animals. Cold Spring Harb. Perspect. Biol. 2016, 8, a028233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spassky, N.; Meunier, A. The development and functions of multiciliated epithelia. Nat. Rev. Mol. Cell Biol. 2017, 18, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Deblandre, G.A.; Wettstein, D.A.; Koyano-Nakagawa, N.; Kintner, C. A two-step mechanism generates the spacing pattern of the ciliated cells in the skin of Xenopus embryos. Development 1999, 126, 4715–4728. [Google Scholar] [CrossRef] [PubMed]
- Wessely, O.; Obara, T. Fish and frogs: Models for vertebrate cilia signaling. Front. Biosci. 2008, 13, 1866–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corkins, M.E.; Krneta-Stankic, V.; Kloc, M.; Miller, R.K. Aquatic models of human ciliary diseases. Genesis 2021, 59, e23410. [Google Scholar] [CrossRef] [PubMed]
- Basquin, C.; Orfila, A.M.; Azimzadeh, J. The planarian Schmidtea mediterranea as a model for studying motile cilia and multiciliated cells. Methods Cell Biol. 2015, 127, 243–262. [Google Scholar]
- Azimzadeh, J.; Basquin, C. Basal bodies across eukaryotes series: Basal bodies in the freshwater planarian Schmidtea mediterranea. Cilia 2016, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Malicki, J.; Avanesov, A.; Li, J.; Yuan, S.; Sun, Z. Analysis of cilia structure and function in zebrafish. Methods Cell Biol. 2011, 101, 39–74. [Google Scholar]
- Leventea, E.; Hazime, K.; Zhao, C.; Malicki, J. Analysis of cilia structure and function in zebrafish. Methods Cell Biol. 2016, 133, 179–227. [Google Scholar]
- Li, Y.; Xu, W.; Jerman, S.; Sun, Z. In vivo analysis of renal epithelial cells in zebrafish. Methods Cell Biol. 2019, 154, 163–181. [Google Scholar]
- Marra, A.N.; Li, Y.; Wingert, R.A. Antennas of organ morphogenesis: The roles of cilia in vertebrate kidney development. Genesis 2016, 54, 457–469. [Google Scholar] [CrossRef]
- Boon, M.; Wallmeier, J.; Ma, L.; Loges, N.T.; Jaspers, M.; Olbrich, H.; Dougherty, G.W.; Raidt, J.; Werner, C.; Amirav, I.; et al. MCIDAS mutations result in a mucociliary clearance disorder with reduced generation of multiple motile cilia. Nat. Commun. 2014, 5, 4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallmeier, J.; Al-Mutairi, D.A.; Chen, C.T.; Loges, N.T.; Pennekamp, P.; Menchen, T.; Ma, L.; Shamseldin, H.E.; Olbrich, H.; Dougherty, G.W.; et al. Mutations in CCNO result in congenital mucociliary clearance disorder with reduced generation of multiple motile cilia. Nat. Genet. 2014, 46, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Olle, M.; Jung, C.; Terre, B.; Balsiger, N.A.; Plata, C.; Roset, R.; Pardo-Pastor, C.; Garrido, M.; Rojas, S.; Alameda, F.; et al. Constitutive Cyclin O deficiency results in penetrant hydrocephalus, impaired growth and infertility. Oncotarget 2017, 8, 99261–99273. [Google Scholar] [CrossRef]
- Nanjundappa, R.; Kong, D.; Shim, K.; Stearns, T.; Brody, S.L.; Loncarek, J.; Mahjoub, M.R. Regulation of cilia abundance in multiciliated cells. eLife 2019, 8, e44039. [Google Scholar] [CrossRef] [PubMed]
- Mercey, O.; Al Jord, A.; Rostaing, P.; Mahuzier, A.; Fortoul, A.; Boudjema, A.R.; Faucourt, M.; Spassky, N.; Meunier, A. Dynamics of centriole amplification in centrosome-depleted brain multiciliated progenitors. Sci. Rep. 2019, 9, 13060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Chen, Q.; Fang, C.; Huang, Q.; Zhou, J.; Yan, X.; Zhu, X. Parental centrioles are dispensable for deuterosome formation and function during basal body amplification. EMBO Rep. 2019, 20, e46735. [Google Scholar] [CrossRef]
- Roberson, E.C.; Tran, N.K.; Konjikusic, M.J.; Fitch, R.D.; Gray, R.S.; Wallingford, J.B. A comparative study of the turnover of multi- ciliated cells in the mouse trachea, oviduct, and brain. Dev. Dynam. 2020, 249, 898–905. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.; Torroja, C.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.Y.; Choi, T.I.; Lee, Y.R.; Choe, S.K.; Kim, C.H. Zebrafish as an animal model for biomedical research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Irion, U.; Nüsslein-Volhard, C. Developmental genetics with model organisms. Proc. Natl. Acad. Sci. USA. 2022, 119, e2122148119. [Google Scholar] [CrossRef] [PubMed]
- Drummond, I.A.; Majumdar, A.; Hentschel, H.; Elger, M.; Solnica-Krezel, L.; Schier, A.F.; Neuhauss, S.C.; Stemple, D.L.; Zwartkruis, F.; Rangini, Z.; et al. Early development of the zebrafish pronephros and analysis of mutations affecting pronephric function. Development 1998, 125, 4655–4667. [Google Scholar] [CrossRef]
- Gerlach, G.F.; Wingert, R.A. Kidney organogenesis in the zebrafish: Insights into vertebrate nephrogenesis and regeneration. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 559–585. [Google Scholar] [CrossRef] [Green Version]
- Poureetezadi, S.J.; Wingert, R.A. Little fish, big catch: Zebrafish as a model for kidney disease. Kidney Int. 2016, 89, 1204–1210. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, G.F.; Wingert, R.A. Zebrafish pronephros tubulogenesis and epithelial identity maintenance are reliant on the polarity proteins prkc iota and zeta. Dev. Biol. 2014, 396, 183–200. [Google Scholar] [CrossRef] [Green Version]
- McKee, R.; Gerlach, G.F.; Jou, J.; Cheng, C.N.; Wingert, R.A. Temporal and spatial expression of tight junction genes during zebrafish pronephros development. Gene Expr. Patterns. 2014, 16, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Zucker, A.G.; Olale, F.; Haycraft, C.J.; Yoder, B.K.; Schier, A.F.; Drummond, I.A. Cilia-driven fluid flow in the zebrafish pronephros, brain and Kupffer’s vesicle is required for normal organogenesis. Development 2005, 132, 1907–1921. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.N.; Ulrich, M.; White, A.; Springer, M.; Wingert, R.A. Visualizing multiciliated cells in the zebrafish through a combined protocol of whole mount fluorescent in situ hybridization and immunofluorescence. J. Vis. Exp. 2017, 129, 56261. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.N.; Chambers, B.E.; Chambers, J.M.; Drummond, B.E.; Adeeb, B.D.; Wesselman, H.M.; Morales, E.E.; Handa, N.; Pettini, T.; Ronshaugen, M.; et al. Visualizing gene expression during zebrafish pronephros development and regeneration. Methods Cell Biol. 2019, 154, 183–215. [Google Scholar] [PubMed]
- Wesselman, H.M.; Gatz, A.; Wingert, R.A. Visualizing multiciliated cells in the zebrafish. Methods Cell Biol. 2022. [Google Scholar] [CrossRef]
- Drummond, I.A.; Davidson, A.J. Zebrafish kidney development. Methods Cell Biol. 2016, 134, 391–429. [Google Scholar] [PubMed]
- Naylor, R.W.; Qubisi, S.S.; Davidson, A.J. Zebrafish pronephros development. Results Probl. Cell Differ. 2017, 60, 27–53. [Google Scholar]
- Ma, M.; Jiang, Y.J. Jagged2a-notch signaling mediates cell fate choice in the zebrafish pronephric duct. PLoS Genet. 2007, 3, e18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pathak, N.; Kramer-Zucker, A.; Drummond, I.A. Notch signaling controls the differentiation of transporting epithelia and multiciliated cells in the zebrafish pronephros. Development 2007, 134, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The cdx genes and retinoic acid control the positioning and segmentation of the zebrafish pronephros. PLoS Genet. 2007, 3, 1922–1938. [Google Scholar] [CrossRef] [Green Version]
- Wingert, R.A.; Davidson, A.J. The zebrafish pronephros: A model to study nephron segmentation. Kidney Int. 2008, 73, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Wingert, R.A.; Davidson, A.J. Zebrafish nephrogenesis involves dynamic spatiotemporal expression changes in renal progenitors and essential signals from retinoic acid and irx3b. Dev. Dyn. 2011, 240, 2011–2027. [Google Scholar] [CrossRef] [Green Version]
- Desgrange, A.; Cereghini, S. Nephron patterning: Lessons from Xenopus, zebrafish, and mouse studies. Cells 2015, 4, 483–499. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.N.; Adeeb, B.D.; Chambers, B.E.; Drummond, B.E.; Ulrich, M.; Addiego, A.; Springer, M.; Poureetezadi, S.J.; Chambers, J.M.; Ronshaugen, M.; et al. Prostaglandin signaling regulates renal multiciliated cell specification and maturation. Proc. Natl. Acad. Sci. USA 2019, 116, 8409–8418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilyev, A.; Liu, Y.; Hellman, N.; Pathak, N.; Drummond, I.A. Mechanical stretch and PI3K signaling link cell migration and proliferation to coordinate epithelial tubule morphogenesis in the zebrafish pronephros. PLoS ONE 2012, 7, e39992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadesch, T. Notch signaling: The demise of elegant simplicity. Curr. Opin. Genet. Dev. 2004, 14, 506–5012. [Google Scholar] [CrossRef] [PubMed]
- Bray, S. Notch signalling in context. Nat. Rev. Mol. Cell Biol. 2016, 17, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, C.N.; Verdun, V.A.; Wingert, R.A. Zebrafish nephrogenesis is regulated by interactions between retinoic acid, mecom, and Notch signaling. Dev. Biol. 2014, 386, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Scheer, N.; Campos-Ortega, J.A. Use of the Ga14-UAS technique for targeted gene expression in the zebrafish. Mech. Dev. 1999, 80, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.N.; Wingert, R.A. Nephron proximal tubule patterning and corpuscles of Stannius formation are regulated by the sim1a transcription factor and retinoic acid in zebrafish. Dev. Biol. 2015, 399, 100–116. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.N.; Wingert, R.A. Epithelial cell fate in the nephron tubule is mediated by the ETS transcription factors etv5a and etv4 during zebrafish kidney development. Dev. Biol. 2016, 411, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Drummond, B.E.; Li, Y.; Marra, A.N.; Cheng, C.N.; Wingert, R.A. The tbx2a/b transcription factors direct pronephros segmentation and corpuscle of Stannius formation in zebrafish. Dev. Biol. 2016, 421, 52–66. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, P.T., Jr.; Drummond, B.E.; Miceli, R.; McKernan, M.; Gerlach, G.F.; Marra, A.N.; Fox, A.; McCampbell, K.K.; Leshchiner, I.; Rodriguez-Mari, A.; et al. The zebrafish kidney mutant zeppelin reveals that brca2/fancd1 is essential for pronephros development. Dev. Biol. 2017, 428, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.E.; Handa, N.; Drummond, B.E.; Chambers, J.M.; Marra, A.N.; Addiego, A.; Wingert, R.A. Homeogene emx1 is required for nephron distal segment development in zebrafish. Sci. Rep. 2018, 8, 18038. [Google Scholar] [CrossRef] [Green Version]
- Znosko, W.A.; Yu, S.; Thomas, K.; Molina, G.A.; Li, C.; Tsang, W.; Dawid, I.B.; Moon, A.M.; Tsang, M. Overlapping functions of Pea3 ETS transcription factors in FGF signaling during zebrafish development. Dev. Biol. 2010, 342, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavodeassi, F.; Modolell, J.; Gómez-Skarmeta, J.L. The Iroquois family of genes: From body building to neural patterning. Development 2001, 128, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Skarmeta, J.L.; Modolell, J. Iroquois genes: Genomic organization and function in vertebrate neural development. Curr. Opin. Genet. Dev. 2002, 12, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.W.; Hui, C.; Strahle, U.; Cheng, S.H. Identification and expression of zebrafish Iroquois homeobox gene irx1. Dev. Genes. Evol. 2001, 211, 442–444. [Google Scholar] [CrossRef] [PubMed]
- Lecauday, V.; Anselme, I.; Dildrop, R.; Rüther, U.; Schneider-Maunoury, S. Expression of the Iroquois genes during early nervous system formation and patterning. J. Comp. Neurol. 2005, 492, 289–302. [Google Scholar] [CrossRef]
- Naylor, R.W.; Chang, H.G.; Qubisi, S.; Davidson, A.J. A novel mechanism of gland formation in zebrafish involving transdifferentiation of renal epithelial cells and live cell extrusion. eLife 2018, 7, e38911. [Google Scholar] [CrossRef]
- Chambers, B.E.; Gerlach, G.F.; Clark, E.G.; Chen, K.H.; Levesque, A.E.; Leshchiner, I.; Goessling, W.; Wingert, R.A. Tfap2a is a novel gatekeeper of nephron differentiation during kidney development. Development 2019, 146, dev172387. [Google Scholar] [CrossRef] [Green Version]
- Chambers, B.E.; Clark, E.G.; Gatz, A.E.; Wingert, R.A. Kctd15 regulates nephron segment development by repressing Tfap2a activity. Development 2020, 147, dev191973. [Google Scholar] [CrossRef]
- Marra, A.N.; Cheng, C.N.; Adeeb, B.; Addiego, A.; Wesselman, H.M.; Chambers, B.E.; Chambers, J.M.; Wingert, R.A. Iroquois transcription factor irx2a is required for multiciliated and transporter cell fate decisions during zebrafish pronephros development. Sci. Rep. 2019, 9, 6454. [Google Scholar] [CrossRef] [Green Version]
- Kalinski, P. Regulation of immune responses by prostaglandin E2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tootle, T.L. Genetic insights into the in vivo functions of prostaglandin signaling. Int. J. Biochem. Cell Biol. 2013, 45, 1629–1632. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Folco, G.; Murphy, R.C. Transcellular biosynthesis of eicosanoids. Pharmacol. Rep. 2010, 62, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Reid, G.; Wielinga, P.; Zelcer, N.; van der Heijden, I.; Kuil, A.; de Haas, M.; Wijnholds, J.; Borst, P. The human multidrug resistance protein MRP4 functions as a prostaglandin efflux transporter and is inhibited by nonsteroidal antiinflammatory drugs. Proc. Natl. Acad. Sci. USA 2003, 100, 9244–9249. [Google Scholar] [CrossRef] [Green Version]
- Schuster, V.L.; Chi, Y.; Lu, R. The prostaglandin transporter: Eicosanoid reuptake, control of signaling, and development of high-affinity inhibitors as drug candidates. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 248–257. [Google Scholar] [PubMed]
- Jin, D.; Zhong, T.P. Prostaglandin signaling in ciliogenesis and development. J. Cell Physiol. 2022, 237, 2632–2643. [Google Scholar] [CrossRef]
- Kiriyama, M.; Ushikubi, F.; Kobayashi, T.; Hirata, M.; Sugimoto, Y.; Narumiya, S. Ligand binding specificities of the eight types and subtypes of the mouse prostanoid receptors expressed in Chinese hamster ovary cells. Br. J. Pharmacol. 1997, 122, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Chadee, K. Prostaglandin E2 stimulates IL-8 gene expression in human colonic epithelial cells by a posttranscriptional mechanism. J. Immunol. 1998, 161, 3746–3752. [Google Scholar]
- Nakayama, T.; Mutsuga, N.; Yao, L.; Tosato, G. Prostaglandin E2 promotes degranulation- independent release of MCP-1 from mast cells. J. Leukoc. Biol. 2006, 79, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Lau, H.Y. Prostaglandin E potentiates the immunologically stimulated histamine release from human peripheral blood-derived mast cells through EP1/EP3 receptors. Allergy 2006, 61, 503–506. [Google Scholar] [CrossRef]
- Weller, C.L.; Collington, S.J.; Hartnell, A.; Conroy, D.M.; Kaise, T.; Barker, J.E.; Wilson, M.S.; Taylor, G.W.; Jose, P.J.; Williams, T.J. Chemotactic action of prostaglandin E2 on mouse mast cells acting via the PGE2 receptor 3. Proc. Natl. Acad. Sci. USA 2007, 104, 11712–11717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambeau, G.; Lazdunski, M. Receptors for a growing family of secreted phospholipases A2. Trends Pharmacol. Sci. 1999, 20, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.I.; Kim, S.H.; Sepich, D.; Buchanan, F.G.; Solnica-Krezel, L.; DuBois, R.N. Cyclooxygenase-1-derived PGE2 promotes cell motility via the G-protein-coupled EP4 receptor during vertebrate gastrulation. Genes Dev. 2006, 20, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosser, T.; Yusuff, S.; Cheskis, E.; Pack, M.A.; FitzGerald, G.A. Developmental expression of functional cyclooxygenases in zebrafish. Proc. Natl. Acad. Sci. USA 2002, 99, 8418–8423. [Google Scholar] [CrossRef] [Green Version]
- Teraoka, H.; Okuno, Y.; Nijoukubo, D.; Yamakoshi, A.; Peterson, R.E.; Stegeman, J.J.; Kitazawa, T.; Hiraga, T.; Kubota, A. Involvement of COX2-thromboxane pathway in TCDD-induced precardiac edema in developing zebrafish. Aquat. Toxicol. 2014, 154, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Galus, M.; Fraz, S.; Gugilla, A.; Jönsson, M.; Wilson, J.Y. Prostaglandins prevent acetaminophen induced embryo toxicity in zebrafish (Danio rerio). Environ. Toxicol. Pharmacol. 2020, 80, 103463. [Google Scholar] [CrossRef]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin signaling regulates nephron segment patterning of renal progenitors during zebrafish kidney development. eLife 2016, 5, e17551. [Google Scholar] [CrossRef]
- Cha, Y.I.; Kim, S.H.; Solnica-Krezel, L.; Dubois, R.N. Cyclooxygenase-1 signaling is required for vascular tube formation during development. Dev. Biol. 2005, 282, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, R.; Tsuge, K.; Kishimoto, K.; Hayashi, Y.; Iwaana, T.; Hohjoh, H.; Inazumi, T.; Kawahara, A.; Tsuchiya, S.; Sugimoto, Y. Essential role of prostaglandin E2 and the EP3 receptor in lymphatic vessel development during zebrafish embryogenesis. Sci. Rep. 2019, 9, 7650. [Google Scholar] [CrossRef] [Green Version]
- Chambers, J.M.; Addiego, A.; Flores-Mireles, A.L.; Wingert, R.A. Ppargc1a controls ciliated cell development by regulating prostaglandin biosynthesis. Cell Rep. 2020, 33, 108370. [Google Scholar] [CrossRef]
- Verdugo, P. Ca2+-dependent hormonal stimulation of ciliary activity. Nature 1980, 283, 764–765. [Google Scholar] [CrossRef]
- Bonin, S.R.; Phillips, P.P.; McCaffrey, T.V. The effect of arachidonic acid metabolites on the ciliary beat frequency of human nasal mucosa in vitro. Acta Otolaryngol. 1992, 112, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Roth, Y.; Kronenberg, J. The effect of prostaglandins E2 and F2a on brain and tracheal ciliary activity. Laryngoscope 1994, 104, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Schuil, P.J.; Ten Berge, M.; Van Gelder, J.M.; Graamans, K.; Huizing, E.H. Effects of prostaglandins D2 and E2 on ciliary beat frequency of human upper respiratory cilia in vitro. Acta Otolaryngol. 1995, 115, 438–442. [Google Scholar] [CrossRef]
- Gayner, S.M.; McCaffrey, T.V. Muscarinic ciliostimulation requires endogenous prostaglandin production. Am. J. Rhinol. 1998, 12, 203–207. [Google Scholar] [CrossRef]
- Haxel, B.R.; Schäfer, D.; Klimek, L.; Mann, W.J. Prostaglandin E2 activates the ciliary beat frequency of cultured human nasal mucosa via the second messenger cyclic adenosine monophosphate. Eur. Arch. Otorhinolaryngol. 2001, 258, 230–235. [Google Scholar] [CrossRef]
- Jin, D.; Ni, T.T.; Sun, J.; Wan, H.; Amack, J.D.; Yu, G.; Fleming, J.; Chiang, C.; Li, W.; Papierniak, A.; et al. Prostaglandin signalling regulates ciliogenesis by modulating intraflagellar transport. Nat. Cell Biol. 2014, 16, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Mul, W.; Mitra, A.; Peterman, E.J.G. Mechanisms of regulation in intraflagellar transport. Cells 2022, 11, 2737. [Google Scholar] [CrossRef] [PubMed]
- Elberg, D.; Turman, M.A.; Pullen, N.; Elberg, G. Prostaglandin E2 stimulates cystogenesis through EP4 receptor in IMCD-3 cells. Prostaglandins Other Lipid Mediat. 2012, 98, 11–16. [Google Scholar] [CrossRef]
- Garcia, H.; Serafin, A.S.; Silbermann, F.; Porée, E.; Viau, A.; Mahaut, C.; Billot, K.; Birgy, É.; Garfa-Traore, M.; Roy, S.; et al. Agonists of prostaglandin E2 receptors as potential first in class treatment for nephronophthisis and related ciliopathies. Proc. Natl. Acad. Sci. USA 2022, 119, e2115960119. [Google Scholar] [CrossRef]
- Eo, J.; Han, K.; Murphy, K.M.; Song, H.; Lim, H.J. Etv5, an ETS transcription factor, is expressed in granulosa and cumulus cells and serves as a transcriptional regulator of the cyclooxygenase-2. J. Endocrinol. 2008, 198, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M.; Poureetezadi, S.J.; Addiego, A.; Lahne, M.; Wingert, R.A. ppargc1a controls nephron segmentation during zebrafish embryonic kidney ontogeny. eLife 2018, 7, e40266. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M.; Wingert, R.A. PGC-1α in disease: Recent renal insights into a versatile metabolic regulator. Cells 2020, 9, 2234. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Xu, X.; Lin, X. Wnt/β-catenin signaling directly regulates Foxj1 expression and ciliogenesis in zebrafish Kupffer’s vesicle. Development 2012, 139, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Chandrasekaran, G.; Li, W.; Kim, D.Y.; Jeong, I.Y.; Lee, S.H.; Liang, T.; Bae, J.Y.; Choi, I.; Kang, H.; et al. Wnt-PLC-IP3-Connexin-Ca2+ axis maintains ependymal motile cilia in zebrafish spinal cord. Nat. Commun. 2020, 11, 1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellman, N.E.; Liu, Y.; Merkel, E.; Austin, C.; Le Corre, S.; Beier, D.R.; Sun, Z.; Sharma, N.; Yoder, B.K.; Drummond, I.A. The zebrafish foxj1a transcription factor regulates cilia function in response to injury and epithelial stretch. Proc. Natl. Acad. Sci. USA 2010, 107, 18499–184504. [Google Scholar] [CrossRef]
- Xie, H.; Kang, Y.; Wang, S.; Zheng, P.; Chen, Z.; Roy, S.; Zhao, C. E2f5 is a versatile transcriptional activator required for spermatogenesis and multiciliated cell differentiation in zebrafish. PLoS Genet. 2020, 16, e1008655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, F.E.; Vladar, E.K.; Ma, L.; Fuentealba, L.C.; Hoh, R.; Espinoza, F.H.; Axelrod, J.D.; Alvarez-Buylla, A.; Stearns, T.; Kintner, C.; et al. Myb promotes centriole amplification and later steps of the multiciliogenesis program. Development 2013, 140, 4277–4286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Narasimhan, V.; Shboul, M.; Chong, Y.L.; Reversade, B.; Roy, S. Gmnc is a master regulator of the multiciliated cell differentiation program. Curr. Biol. 2015, 25, 3267–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Rayamajhi, D.; Ravi, V.; Narasimhan, V.; Chong, Y.L.; Lu, H.; Venkatesh, B.; Roy, S. Conservation as well as divergence in Mcidas function underlies the differentiation of multiciliated cells in vertebrates. Dev. Biol. 2020, 465, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.I.; Kwon, T.; Tu, F.; Brooks, E.R.; Gupta, R.; Meyer, M.; Baker, J.C.; Marcotte, E.M.; Wallingford, J.B. Coordinated genomic control of ciliogenesis and cell movement by RFX2. eLife 2014, 3, e01439. [Google Scholar] [CrossRef] [PubMed]
- Gegg, M.; Böttcher, A.; Burtscher, I.; Hasenoeder, S.; Van Campenhout, C.; Aichler, M.; Walch, A.; Grant, S.G.; Lickert, H. Flattop regulates basal body docking and positioning in mono- and multiciliated cells. eLife 2014, 3, e03842. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, J.L.; Vladar, E.K.; Axelrod, J.D.; Kintner, C. Multicilin promotes centriole assembly and ciliogenesis during multiciliate cell differentiation. Nat. Cell Biol. 2012, 14, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.N.; Li, Y.; Marra, A.N.; Verdun, V.; Wingert, R.A. Flat mount preparation for observation and analysis of zebrafish embryo specimens stained by whole mount in situ hybridization. J. Vis. Exp. 2014, 89, 51604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, Y.L.; Zhang, Y.; Zhou, F.; Roy, S. Distinct requirements of E2f4 versus E2f5 activity for multiciliated cell development in the zebrafish embryo. Dev. Biol. 2018, 443, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Arbi, M.; Pefani, D.E.; Taraviras, S.; Lygerou, Z. Controlling centriole numbers: Geminin family members as master regulators of centriole amplification and multiciliogenesis. Chromosoma 2018, 127, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Choksi, S.P.; Babu, D.; Lau, D.; Yu, X.; Roy, S. Systematic discovery of novel ciliary genes through functional genomics in the zebrafish. Development 2014, 141, 3410–3419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, F.; Sedzinski, J.; Ma, Y.; Marcotte, E.M.; Wallingford, J.B. Protein localization screening in vivo reveals novel regulators of multiciliated cell development and function. J. Cell Sci. 2018, 131, jcs206565. [Google Scholar] [CrossRef] [Green Version]
- Defosset, A.; Merlat, D.; Poidevin, L.; Nevers, Y.; Kress, A.; Poch, O.; Lecompte, O. Novel approach combining transcriptional and evolutionary signatures to identify new multiciliation genes. Genes 2021, 12, 1452. [Google Scholar] [CrossRef]
- Chambers, B.E.; Wingert, R.A. Renal progenitors: Roles in kidney disease and regeneration. World J. Stem Cells. 2016, 8, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M.; Wingert, R.A. Advances in understanding vertebrate nephrogenesis. Tissue Barriers 2020, 8, 1832844. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.D. Cilia in the fetal kidney of man. Beitr. Pathol. 1971, 143, 227–240. [Google Scholar]
- Katz, S.M.; Morgan, J.J. Cilia in the human kidney. Ultrastruct. Pathol. 1984, 6, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.L.; Suzuki, Y. Ciliated human renal proximal tubular cells. Observations in three cases of hypercalcemia. Am. J. Pathol. 1968, 53, 609–616. [Google Scholar]
- Datsis, S.A.; Boman, I.A. Ciliated renal tubular epithelium in congenital nephrosis. Beitr. Pathol. 1974, 151, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.E.; Ghadially, F.N. Cilia in lupus nephritis. J. Pathol. 1974, 114, 69–73. [Google Scholar] [CrossRef]
- Lungarella, G.; de Santi, M.M.; Tosi, P. Ultrastructural study of the ciliated cells from renal tubular epithelium in acute progressive glomerulonephritis. Ultrastruct. Pathol. 1984, 6, 1–7. [Google Scholar] [CrossRef]
- Ong, A.C.; Wagner, B. Detection of proximal tubular motile cilia in a patient with renal sarcoidosis associated with hypercalcemia. Am. J. Kidney Dis. 2005, 45, 1096–1099. [Google Scholar] [CrossRef]
- Johnson, C.S.; Holzemer, N.F.; Wingert, R.A. Laser ablation of the zebrafish pronephros to study renal epithelial regeneration. J. Vis. Exp. 2011, 54, e2845. [Google Scholar] [CrossRef] [Green Version]
- Palmyre, A.; Lee, J.; Ryklin, G.; Camarata, T.; Selig, M.K.; Duchemin, A.L.; Nowak, P.; Arnaout, M.A.; Drummond, I.A.; Vasilyev, A. Collective epithelial migration drives kidney repair after acute injury. PLoS ONE 2014, 9, e101304. [Google Scholar] [CrossRef] [Green Version]
- Yakulov, T.A.; Todkar, A.P.; Slanchev, K.; Wiegel, J.; Bona, A.; Groß, M.; Scholz, A.; Hess, I.; Wurditsch, A.; Grahammer, F.; et al. CXCL12 and MYC control energy metabolism to support adaptive responses after kidney injury. Nat. Commun. 2018, 9, 3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Boucher, R.C.; Bollig, F.; Englert, C.; Hildebrandt, F. Characterization of mesonephric development and regeneration using transgenic zebrafish. Am. J. Physiol. Ren. Physiol. 2010, 299, F1040–F1047. [Google Scholar] [CrossRef]
- Diep, C.Q.; Ma, D.; Deo, R.C.; Holm, T.M.; Naylor, R.W.; Arora, N.; Wingert, R.A.; Bollig, F.; Djordjevic, G.; Lichman, B.; et al. Identification of adult nephron progenitors capable of kidney regeneration in zebrafish. Nature 2011, 470, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, P.T., Jr.; Wingert, R.A. Using zebrafish to study podocyte genesis during kidney development and regeneration. Genesis 2014, 52, 771–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCampbell, K.K.; Springer, K.N.; Wingert, R.A. Analysis of nephron composition and function in the adult zebrafish kidney. J. Vis. Exp. 2014, 90, e51644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCampbell, K.K.; Springer, K.N.; Wingert, R.A. Atlas of cellular dynamics during zebrafish adult kidney regeneration. Stem Cells Int. 2015, 2015, 547636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, R.A.; Wingert, R.A. Zebrafish renal pathology: Emerging models of acute kidney injury. Curr. Pathobiol. Rep. 2015, 3, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Tasca, A.; Helmstädter, M.; Brislinger, M.M.; Haas, M.; Mitchell, B.; Walentek, P. Notch signaling induces either apoptosis or cell fate change in multiciliated cells during mucociliary tissue remodeling. Dev. Cell 2021, 56, 525–539.e6. [Google Scholar] [CrossRef]
- Drummond, B.E.; Chambers, B.E.; Wesselman, H.M.; Gibson, S.; Arceri, L.; Ulrich, M.N.; Gerlach, G.F.; Kroeger, P.T.; Leshchiner, I.; Goessling, W.; et al. osr1 Maintains Renal Progenitors and Regulates Podocyte Development by Promoting wnt2ba via the Antagonism of hand2. Biomedicines 2022, 10, 2868. [Google Scholar] [CrossRef]
- Weaver, N.E.; Healy, A.; Wingert, R.A. gldc Is Essential for Renal Progenitor Patterning during Kidney Development. Biomedicines 2022, 10, 3220. [Google Scholar] [CrossRef]
- Morales, E.E.; Wingert, R.A. Zebrafish as a model of kidney disease. Results Probl. Cell Differ. 2017, 60, 55–75. [Google Scholar] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wesselman, H.M.; Nguyen, T.K.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Advances in Understanding the Genetic Mechanisms of Zebrafish Renal Multiciliated Cell Development. J. Dev. Biol. 2023, 11, 1. https://doi.org/10.3390/jdb11010001
Wesselman HM, Nguyen TK, Chambers JM, Drummond BE, Wingert RA. Advances in Understanding the Genetic Mechanisms of Zebrafish Renal Multiciliated Cell Development. Journal of Developmental Biology. 2023; 11(1):1. https://doi.org/10.3390/jdb11010001
Chicago/Turabian StyleWesselman, Hannah M., Thanh Khoa Nguyen, Joseph M. Chambers, Bridgette E. Drummond, and Rebecca A. Wingert. 2023. "Advances in Understanding the Genetic Mechanisms of Zebrafish Renal Multiciliated Cell Development" Journal of Developmental Biology 11, no. 1: 1. https://doi.org/10.3390/jdb11010001