Improved Cordycepin Production by Cordyceps militaris KYL05 Using Casein Hydrolysate in Submerged Conditions


 ,
,  and
and
Abstract
:
1. Introduction
2. Experimental
2.1. Microorganisms
2.2. Media and Culture Conditions
2.2.1. Seed Culture of C. Militaris
2.2.2. Main Culture of C. Militaris
2.2.3. Effects of Temperature, Initial pH, and Shaking Speed
2.2.4. Effects of Carbon Source and Nitrogen Source
2.3. Analytical Methods
3. Results and Discussion
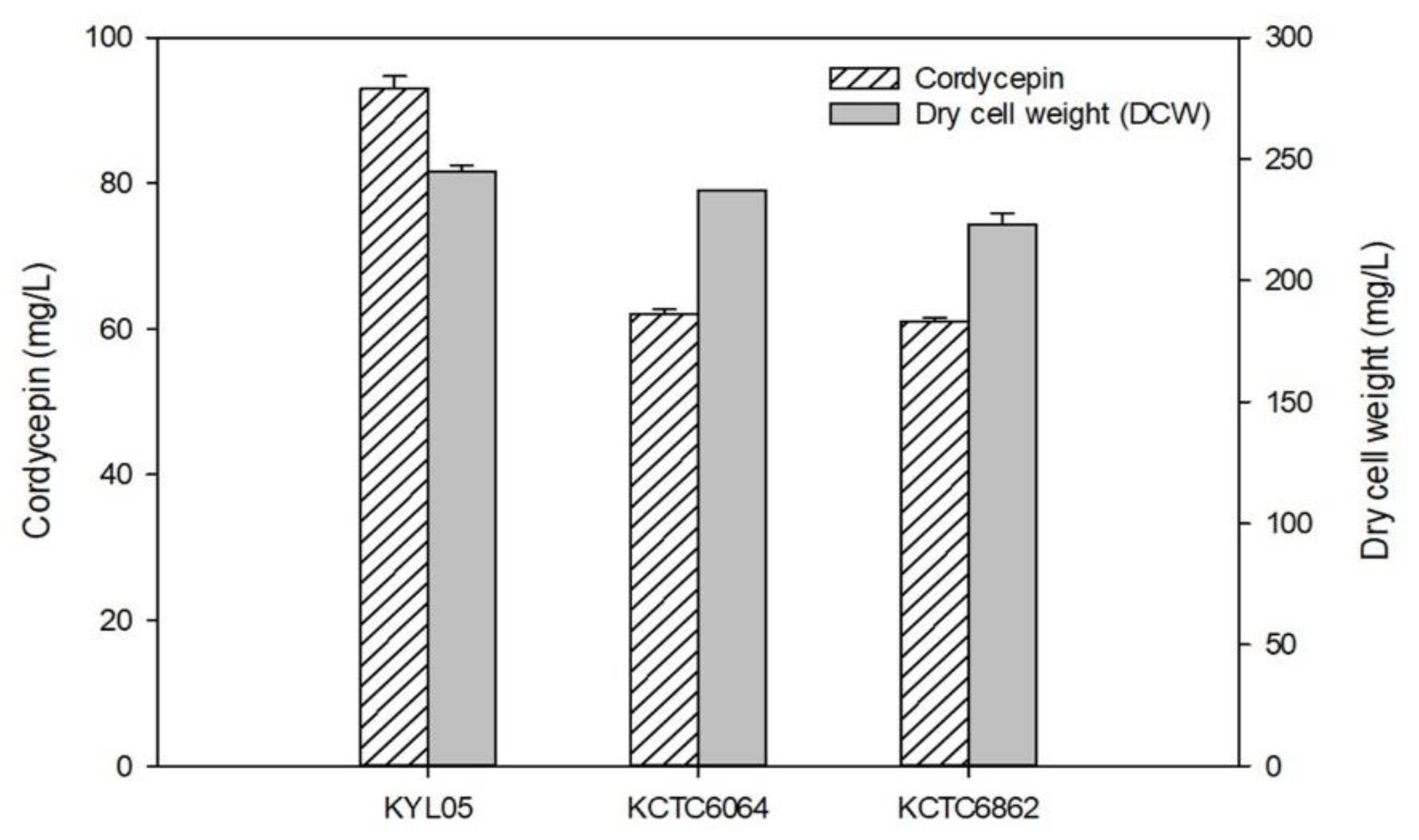
3.1. Microbial Strain Selection for Cordycepin Production
3.2. Determination of Culture Conditions for Cordycepin Production
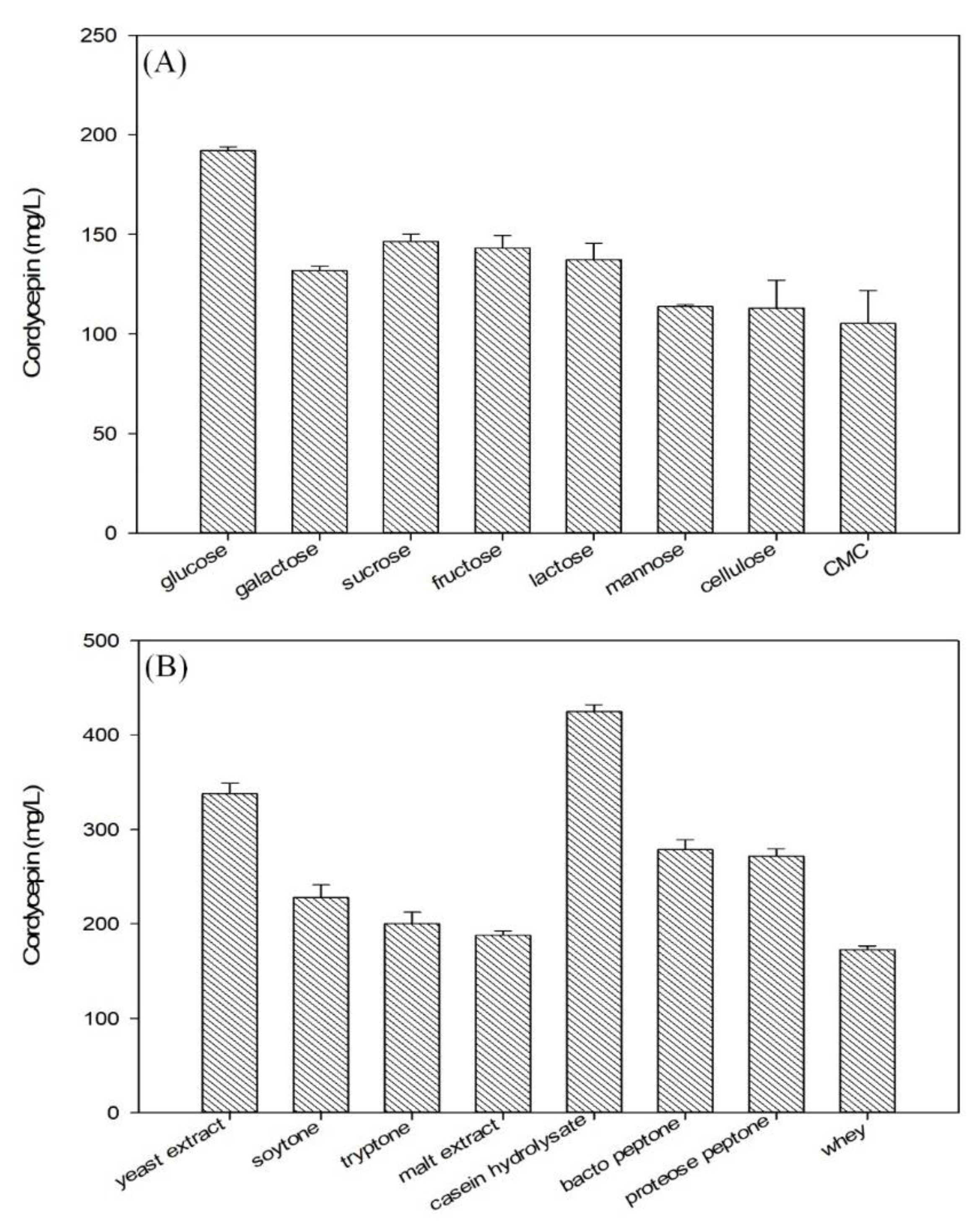
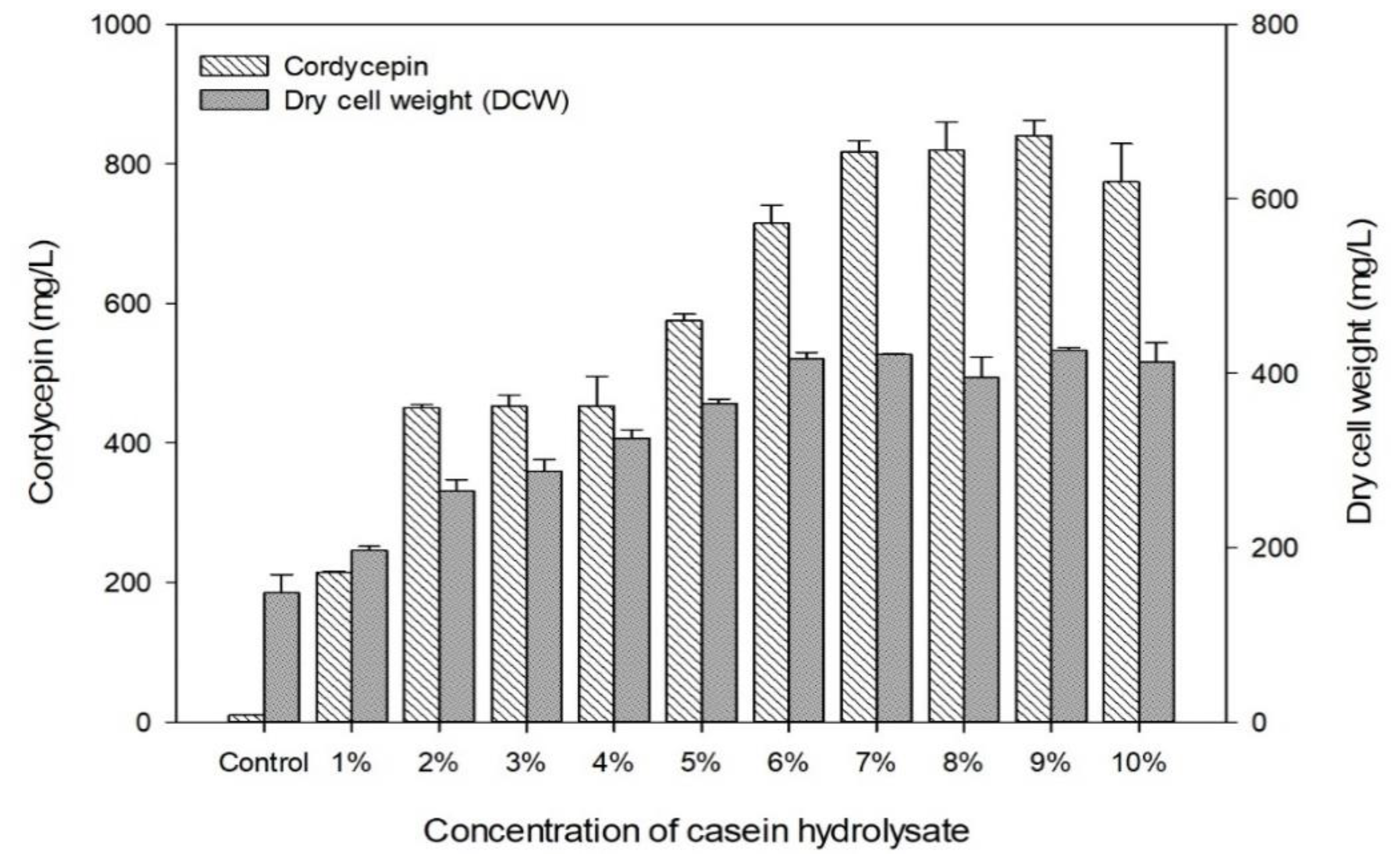
3.3. Effect of Carbon and Nitrogen Source on Cordycepin Production
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Chang, P.C.; Chou, C.J.; Chen, C.F. Protein constituent contributes to the hypotensive and vasorelaxant acttvtties of cordyceps sinensis. Life Sci. 2000, 66, 1369–1376. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of Cordyceps with special reference to cordycepin. 3 Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Zhang, W.; Zhang, Y.; Liu, X. The medicinal fungus Cordyceps militaris: Research and development. Mycol. Prog. 2012, 11, 599–614. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.W.; Dong, S.J.; Xu, F.X.; Wang, S.H. The optimization of extraction of cordycepin from fruiting body of Cordyceps militaris (L.) Link. Adv. Mater. Res. 2011, 393, 1024–1028. [Google Scholar] [CrossRef]
- Kwon, J.S.; Lee, J.S.; Shin, W.C.; Lee, K.E.; Hong, E.K. Optimization of culture conditions and medium components for the production of mycelial biomass and exo-polysaccharides with Cordyceps militaris in liquid culture. Biotechnol. Bioprocss Eng. 2009, 14, 756–762. [Google Scholar] [CrossRef]
- Fan, D.D.; Wang, W.; Zhong, J.J. Enhancement of cordycepin production in submerged cultures of Cordyceps militaris by addition of ferrous sulfate. Biochem. Eng. J. 2012, 60, 30–35. [Google Scholar] [CrossRef]
- Masuda, M.; Urabe, E.; Sakurai, A.; Sakakibara, M. Production of cordycepin by surface culture using the medicinal mushroom Cordyceps militaris. Enzym. Microb. Technol. 2006, 39, 641–646. [Google Scholar] [CrossRef]
- Shih, I.L.; Tsai, K.-L.; Hsieh, C. Effects of culture conditions on the mycelial growth and bioactive metabolite production in submerged culture of Cordyceps militaris. Biochem. Eng. J. 2007, 33, 193–201. [Google Scholar] [CrossRef]
- Oh, J.; Yoon, D.H.; Shrestha, B.; Choi, H.K.; Sung, G.H. Metabolomic profiling reveals enrichment of cordycepin in senescence process of Cordyceps militaris fruit bodies. J. Microbiol. 2019, 57, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.Y.; Lee, J.H.; Kim, D.S.; Lee, J.H.; Lee, S.K.; Lee, S.J.; Park, C.; Kim, S.W. Enhancement of glucose yield from canola agricultural residue by alkali pretreatment based on multi-regression models. J. Ind. Eng. Chem. 2017, 51, 303–311. [Google Scholar] [CrossRef]
- Siripong, P.; Doungporn, P.; Yoo, H.Y.; Kim, S.W. Improvement of sugar recovery from Sida acuta (Thailand Weed) by NaOH pretreatment and application to bioethanol production. Korean J. Chem. Eng. 2018, 35, 2413–2420. [Google Scholar] [CrossRef]
- Yang, H.J.; Lee, T.; Kim, J.R.; Choi, Y.-E.; Park, C. Improved production of bacterial cellulose from waste glycerol through investigation of inhibitory effects of crude glycerol-derived compounds by Gluconacetobacter xylinus. J. Ind. Eng. Chem. 2019, 75, 158–163. [Google Scholar]
- Lee, S.J.; Choi, H.S.; Kim, C.K.; Thapa, L.P.; Park, C.; Kim, S.W. Process strategy for 2,3-butanediol production in fed-batch culture by acetate addition. J. Ind. Eng. Chem. 2017, 56, 157–162. [Google Scholar] [CrossRef]
- Kim, H.; Yoo, H.Y.; Kim, Y.H.; Kim, I.K.; Byun, E.H.; Yang, Y.H.; Park, S.J.; Na, J.G.; Shon, H.; Lee, T.; et al. Improved reutilization of industrial crude lysine to 1,5-diaminopentane by enzymatic decarboxylation using various detergents and organic solvents. Korean J. Chem. Eng. 2018, 35, 1854–1859. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.R.; Lee, J.H.; Lee, S.K.; Chun, Y.; Han, S.O.; Yoo, H.Y.; Park, C.; Kim, S.W. Enhanced in-vitro Hemozoin polymerization by optimized process using Histidine-Rich Protein II (HRPII). Polymers 2019, 11, 1162. [Google Scholar] [CrossRef]
- Kang, S.W.; Lee, H.; Kim, D.; Lee, D.; Kim, S.; Chun, G.T.; Lee, J.; Kim, S.W.; Park, C. Strain development and medium optimization for fumaric acid production. Biotechnol. Bioprocss Eng. 2010, 15, 761–769. [Google Scholar] [CrossRef]
- Jung, Y.R.; Shin, H.Y.; Yoo, H.Y.; Um, Y.; Kim, S.W. Production of cellulases and β-glucosidase in Trichoderma reesei mutated by proton beam irradiation. Korean J. Chem. Eng. 2012, 29, 925–930. [Google Scholar] [CrossRef]
- Jung, D.U.; Yoo, H.Y.; Kim, S.B.; Lee, J.H.; Park, C.; Kim, S.W. Optimization of medium composition for enhanced cellulose production by mutant Penicillium brasilianum KUEB15 using statistical method. J. Ind. Eng. Chem. 2015, 25, 145–150. [Google Scholar] [CrossRef]
- Cha, S.H.; Lim, J.S.; Yoon, C.S.; Koh, J.H.; Chang, H.I.; Kim, S.W. Production of mycelia and exo-biopolymer from molasses by Cordyceps sinensis 16 in submerged culture. Bioresour. Technol. 2007, 98, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Kim, J.C.; Lim, J.S.; Yoon, C.S.; Koh, J.H.; Chang, H.I.; Kim, S.W. Morphological characteristics of Cordyceps sinensis 16 and production of mycelia and exo-biopolymer from molasses in submerged culture. J. Ind. Eng. Chem. 2006, 12, 115–120. [Google Scholar]
- Lee, J.Y.; Kang, S.W.; Kim, S.W. Relationship between agitation speed and the morphological characteristics of Verticillium lecanii CS-625 during spore production. Biotechnol. Bioprocess Eng. 2008, 13, 1–6. [Google Scholar] [CrossRef]
- Kunhorm, P.; Chaicharoenaudomrung, N.; Noisa, P. Enrichment of cordycepin for cosmeceutical applications: Culture systems and strategies. Appl. Microbiol. Biotechnol. 2019, 103, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Jin, L.; Ren, X.; Lu, J.; Meng, Q. Optimization of fermentation process of Cordyceps militaris and antitumor activities of polysaccharides in vitro. J. Food Drug Anal. 2014, 22, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Lai, Y.J.; Wu, S.C.; Hsu, W.H.; Tai, C.J. Optimal conditions for cordycepin production in surface liquid-cultured Cordyceps militaris treated with porcine liver extracts for suppression of oral cancer. J. Food Drug Anal. 2018, 26, 135–144. [Google Scholar] [CrossRef]
- Wanwipa, V.; Nachon, R.; Warasinee, M.; Nam Ninh, N.; Hon Wai, L.; Kobkul, L. Genome-scale metabolic network of Cordyceps militaris useful for comparative analysis of entomopathogenic fungi. Gene 2017, 626, 132–139. [Google Scholar]
- Kang, C.; Wen, T.C.; Kang, J.C.; Meng, Z.B.; Li, G.R.; Hyde, K.D. Optimization of large-scale culture conditions for the production of cordycepin with Cordyceps militaris by liquid static culture. Sci. World J. 2014, 2014, 15. [Google Scholar] [CrossRef]
- Mao, X.B.; Eksriwong, T.; Chauvatcharin, S.; Zhong, J.J. Optimization of carbon source and carbon/nitrogen ratio for cordycepin production by submerged cultivation of medicinal mushroom Cordyceps militaris. Process Biochem. 2005, 40, 1667–1672. [Google Scholar] [CrossRef]
- Tang, J.; Qian, Z.; Wu, H. Enhancing cordycepin production in liquid static cultivation of Cordyceps militaris by adding vegetable oils as the secondary carbon source. Bioresour. Technol. 2018, 268, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.B.; Zhong, J.J. Significant effect of NH4+ on cordycepin production by submerged cultivation of medicinal mushroom Cordyceps militaris. Enzym. Microb. Technol. 2006, 38, 343–350. [Google Scholar] [CrossRef]
- Jo, S.J.; Lee, T.H.; Chae, D.H.; Han, Y.H. Optimization of culture condition and media composition on the production of cordycepin by Cordyceps militaris. Korean J. Microbiol. 2004, 40, 217–220. [Google Scholar]
- Mao, X.B.; Zhong, J.J. Hyperproduction of cordycepin by two-stage dissolved oxygen control in submerged cultivation of medicinal mushroom Cordyceps militaris in bioreactors. Biotechnol. Prog. 2004, 20, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.C.; Long, F.Y.; Kang, C.; Wang, F.; Zeng, W. Effects of additives and bioreactors on cordycepin production from Cordyceps militaris in liquid static culture. Mycosphere 2017, 8, 886–898. [Google Scholar] [CrossRef]
- Wu, J.Y.; Wong, K.; Ho, K.P.; Zhou, L.G. Enhancement of saponin production in Panax ginseng cell culture by osmotic stress and nutrient feeding. Enzym. Microb. Technol. 2005, 36, 133–138. [Google Scholar] [CrossRef]
- Wang, J.; Su, Y.; Jia, F.; Jin, H. Characterization of casein hydrolysates derived from enzymatic hydrolysis. Chem. Cent. J. 2013, 7, 62. [Google Scholar] [CrossRef]
- Bister-Miel, F.; Guignard, J.L.; Bury, M.; Agier, C. Glutamine as an active component of casein hydrolysate: Its balancing effect on plant cells cultured in phosphorus deficient medium. Plant Cell Rep. 1985, 4, 161–163. [Google Scholar] [CrossRef]
- Leung, P.H.; Wu, J.Y. Effects of ammonium feeding on the production of bioactive metabolites (cordycepin and exopolysaccharides) in mycelial culture of a Cordyceps sinensis fungus. J. Appl. Microbiol. 2007, 103, 1942–1949. [Google Scholar] [CrossRef]
- Takagi, H. Metabolic regulatory mechanisms and physiological roles of functional amino acids and their applications in yeast. Biosci. Biotechnol. Biochem. 2019, 83, 1449–1462. [Google Scholar] [CrossRef]
- Raethong, N.; Laoteng, K.; Vongsangnak, W. Uncovering global metabolic response to cordycepin production in Cordyceps militaris through transcriptome and genome-scale network-driven analysis. Sci. Rep. 2018, 8, 9250. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Culture Conditions | Carbon Source (g/L) | Nitrogen Source (g/L) | Cordycepin Production (mg/L) | Cordycepin Productivity (mg/L∙day) | Ref. | |||
|---|---|---|---|---|---|---|---|---|---|
| Temp. (°C) | RPM | pH | Time (day) | ||||||
| C. militaris | 25 | 110 | 6 | 20 | Glucose 40 | Peptone 10 | 597 | 29.9 | [9] |
| C. militaris | 25 | 110 | 4.8 | 18 | Glucose 42 | Peptone 16 | 245 | 13.6 | [30] |
| C. militaris | 25 | 110 | 5 | 17 | Glucose 10 | Peptone 10 | 346 | 20.4 | [32] |
| C. militaris KCTC6862 | 24 | 120 | 6.5 | 5 | Glucose 10 | Peptone 10 | 22 | 4.4 | [33] |
| C. militaris KCTC16932 | 24 | 120 | 6.5 | 5 | Glucose 10 | Peptone 10 | 23 | 4.6 | |
| C. militaris DGUM32003 | 24 | 120 | 6.5 | 5 | Glucose 10 | Peptone 10 | 39 | 7.8 | |
| C. militaris | 25 | 200 | 4.8 | 17 | Glucose 40 | Peptone 10 | 201 | 11.8 | [34] |
| C. militaris KYL05 | 25 | 150 | 6 | 6 | Glucose 20 | CH 20 | 445 | 74.2 | This study |
| Amino Acid | Amino Acid Concentration (g-Amino Acid/100 g-Nitrogen Source) | ||
|---|---|---|---|
| CH | Yeast Extract | Peptone | |
| Aspartic acid | 6.12 | 6.54 | 5.82 |
| Threonine | 3.69 | 3.19 | 2.18 |
| Serine | 4.42 | 3.29 | 3.33 |
| Glutamate | 18.01 | 12.44 | 9.97 |
| Proline | 7.48 | 2.80 | 10.13 |
| Glycine | 1.69 | 3.07 | 20.00 |
| Alanine | 2.75 | 5.75 | 8.87 |
| Valine | 5.58 | 4.16 | 2.88 |
| Isoleucine | 4.33 | 3.45 | 1.76 |
| Leucine | 7.46 | 4.92 | 3.48 |
| Tyrosine | 3.19 | 0.70 | 0.48 |
| Phenylalanine | 3.88 | 2.90 | 2.15 |
| Lysine | 6.65 | 5.29 | 3.89 |
| Histidine | 2.11 | 1.38 | 0.82 |
| Arginine | 2.84 | 3.41 | 7.10 |
| Cysteine | 0.47 | 1.07 | 0.34 |
| Methionine | 2.46 | 1.08 | 0.84 |
| Total content | 83.13 | 65.42 | 84.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.K.; Lee, J.H.; Kim, H.R.; Chun, Y.; Lee, J.H.; Yoo, H.Y.; Park, C.; Kim, S.W. Improved Cordycepin Production by Cordyceps militaris KYL05 Using Casein Hydrolysate in Submerged Conditions. Biomolecules 2019, 9, 461. https://doi.org/10.3390/biom9090461
Lee SK, Lee JH, Kim HR, Chun Y, Lee JH, Yoo HY, Park C, Kim SW. Improved Cordycepin Production by Cordyceps militaris KYL05 Using Casein Hydrolysate in Submerged Conditions. Biomolecules. 2019; 9(9):461. https://doi.org/10.3390/biom9090461
Chicago/Turabian StyleLee, Soo Kweon, Ju Hun Lee, Hyeong Ryeol Kim, Youngsang Chun, Ja Hyun Lee, Hah Young Yoo, Chulhwan Park, and Seung Wook Kim. 2019. "Improved Cordycepin Production by Cordyceps militaris KYL05 Using Casein Hydrolysate in Submerged Conditions" Biomolecules 9, no. 9: 461. https://doi.org/10.3390/biom9090461