Microbial Enzymes: Tools for Biotechnological Processes

Abstract
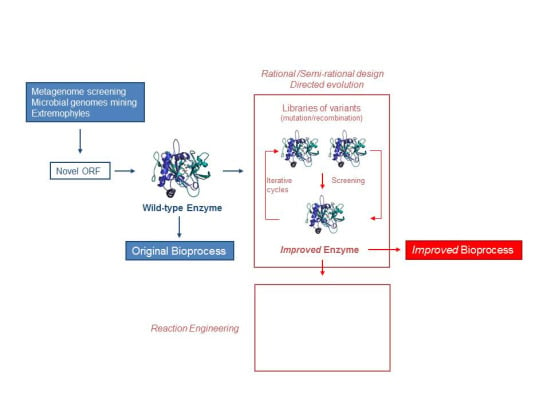
:
{kind=link}
{kind=link}
1. Introduction
2. Discovering Enzymes
2.1. Metagenomic Screening
2.2. Microbial Genomes
2.3. Extremophiles
3. Strategies to Improve Properties of Microbial Enzymes

3.1. Rational Design
3.2. Directed Evolution
4. Production of Recombinant Proteins in Microbial Hosts
5. Enzymatic Biocatalysis
5.1. Technical Applications
5.2. Enzymes in the Feed Industry
5.3. Enzymes in Food Processing
5.4. Enzymes in Chemical and Pharmaceutical Processes
6. Conclusion and Future Perspectives
Conflicts of Interest
References
- Adrio, J.L.; Demain, A.L. Microbial Cells and Enzymes—A Century of Progress. In Methods in Biotechnology. Microbial Enzymes and Biotransformations; Barredo, J.L., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 17, pp. 1–27. [Google Scholar]
- Johannes, T.W.; Zhao, H. Directed evolution of enzymes and biosynthetic pathways. Curr. Opin. Microbiol. 2006, 9, 261–267. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S. Directed evolution: Tailoring biocatalysis for industrial application. Crit. Rev. Biotechnol. 2013, 33, 365–378. [Google Scholar] [CrossRef]
- Demain, A.L.; Adrio, J.L. Contributions of microorganisms to industrial biology. Mol. Biotechnol. 2008, 38, 41–45. [Google Scholar] [CrossRef]
- Johnson, E.A. Biotechnology of non-Saccharomyces yeasts- the ascomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 503–517. [Google Scholar] [CrossRef]
- Rondon, M.R.; Goodman, R.M.; Handelsman, J. The Earth’s bounty: Assessing and accessing soil microbial diversity. Trends Biotechnol. 1999, 17, 403–409. [Google Scholar] [CrossRef]
- Verenium, A Pioneer of 21st Century Bioscience, is Working to Transform Industries. Available online: http://www.diversa.com (access on 1 July 2013).
- Uchiyama, T.; Miyazaki, K. Functional metagenomics for enzyme discovery: Challenges to efficient screening. Curr. Opin. Biotechnol. 2009, 20, 616–622. [Google Scholar] [CrossRef]
- Ferrer, M.; Beloqui, A.; Timmis, K.M.; Golyshin, P.N. Metagenomics for mining new genetic resources of microbial communities. J. Mol. Microbiol. Biotechnol. 2009, 16, 109–123. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Dupont, C.L. Microbial metagenomics: Beyond the genome. Annu. Rev. Mar. Sci. 2011, 3, 347–371. [Google Scholar] [CrossRef]
- The Institute for Genomic Research (TIGR)-JCVI. Available online: http://cmr.jcvi.org/tigr (accessed on 1 July 2013).
- The National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 1 July 2013).
- Ahmed, N. A flood of microbial genomes—Do we need more? PLoS One 2009, 4, 1–5. [Google Scholar]
- Kaul, P.; Asano, Y. Strategies for discovery and improvement of enzyme function: State of the art and opportunities. Microbiol. Biotechnol. 2012, 5, 18–33. [Google Scholar] [CrossRef]
- Schiraldini, C.; de Rosa, M. The production of biocatalysts and biomolecules from extremophiles. Trends Biotechnol. 2002, 20, 515–521. [Google Scholar] [CrossRef]
- Kumar, L.; Awasthi, G.; Singh, B. Extremophiles: A novel source of industrially important enzymes. Biotechnology 2011, 10, 1–15. [Google Scholar] [CrossRef]
- Steele, H.L.; Jaeger, K.E.; Daniel, R.; Streit, W.R. Advances in recovery of novel biocatalysts from metagenomes. J. Mol. Microbiol. Biotechnol. 2009, 16, 25–37. [Google Scholar] [CrossRef]
- Li, S.; Yang, X.; Yang, S.; Zhu, M.; Wang, X. Technology prospecting on enzymes: Application, marketing and engineering. Comput. Struct. Biotechnol. J. 2012, 2, 1–11. [Google Scholar]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef]
- Kennedy, J.; Marchesi, J.R.; Dobson, A.D. Marine metagenomics: Strategies for the discovery of novel enzymes with biotechnological applications from marine environments. Microb. Cell Fact. 2008, 7, 27–37. [Google Scholar] [CrossRef]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef]
- Jeon, J.H.; Kim, J.T.; Kim, Y.J.; Kim, H.K.; Lee, H.S.; Kang, S.G.; Kim, S.J.; Lee, J.H. Cloning and characterization of a new-cold active lipase from a deep-se sediment metagenome. Appl. Microbiol. Biotechnol. 2009, 81, 865–874. [Google Scholar] [CrossRef]
- Fernández-Álvaro, E.; Kourist, R.; Winter, J.; Böttcher, D.; Liebeton, K.; Naumer, C. Enantioselective kinetic resolution of phenylalkyl carboxylic acids using metagenome-derived esterases. Microb. Biotechnol. 2010, 3, 59–64. [Google Scholar] [CrossRef]
- Knietsch, A.; Waschkowitz, T.; Bowien, S.; Henne, A.; Daniel, R. Construction and screening of metagenomic libraries derived from enrichment cultures: Generation of a gene bank for genes conferring alcohol oxidoreductase activity on Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 1408–1416. [Google Scholar] [CrossRef]
- Gabor, E.M.; de Vries, E.J.; Janssen, D.B. Construction, characterization, and use of small-insert gene banks of DNA isolated from soil and enrichment cultures for the recovery of novel amidases. Environ. Microbiol. 2004, 6, 948–958. [Google Scholar] [CrossRef]
- Rondon, M.R.; August, P.R.; Betterman, A.D.; Brady, S.F.; Grossman, T.H.; Liles, M.R. Cloning the soil metagenome: A strategy for accesing the genetic and functional diversity of uncultured microorganisms. Appl. Environ. Microbiol. 2000, 66, 2541–2547. [Google Scholar] [CrossRef]
- Bayer, S.; Birkemeyer, C.; Ballschmiter, M. A nitrilase from ametagenomic library acts regioselectively on aliphatic dinitriles. Appl. Microbiol. Biotechnol. 2011, 89, 91–98. [Google Scholar] [CrossRef]
- Wang, K.; Li, G.; Yu, S.Q.; Zhang, C.T.; Liu, Y.H. A novel metegenome-derived β-galactosidase: Gene cloning, overexpression, purification and characterization. Appl. Microbiol. Biotechnol. 2010, 88, 155–165. [Google Scholar] [CrossRef]
- Jiang, C.; Li, S.X.; Luo, F.F.; Jin, K.; Wang, Q.; Hao, Z.Y. Biochemical characterization of two novel β-glucosidase genes by metagenome expression cloning. Bioresour. Technol. 2011, 102, 3272–3278. [Google Scholar] [CrossRef]
- Jiang, C.; Shen, P.; Yan, B.; Wua, B. Biochemical characterization of a metagenome-derived decarboxylase. Enzyme Microb. Technol. 2009, 45, 58–63. [Google Scholar] [CrossRef]
- Kotik, M.; Stepanek, V.; Grulich, M.; Kyslik, P.; Archelas, A. Access to enantiopure aromatic epoxides and diols using epoxide hydrolases derived from total biofilter DNA. J. Mol. Catal. B Enzym. 2010, 65, 41–48. [Google Scholar] [CrossRef]
- Martínez, A.; Kolvek, S.J.; Yip, C.L.T.; Hopke, J.; Brown, K.A.; MacNeil, I.A.; Osburne, M.S. Genetically modified bacterial strains and novel bacterial artificial chromosome shuttle vectors for constructing environmental libraries and detecting heterologous naturals products in multiple expression hosts. Appl. Environ. Microbiol. 2004, 70, 2452–2463. [Google Scholar] [CrossRef]
- Wexler, M.; Bond, P.L.; Richardson, D.J.; Johnston, A.W. A wide host-range metagenomic library from a waste water treatment plants yields a novel alcohol/aldehyde dehydrogenase. Environ. Microbiol. 2005, 7, 1917–1926. [Google Scholar] [CrossRef]
- Suenaga, H.; Ohnuki, T.; Miyazaki, K. Functional screening of a metagenomic library for genes involved in microbial degradation of aromatic compounds. Environ. Microbiol. 2007, 9, 2289–2297. [Google Scholar] [CrossRef]
- Mirete, S.; de Figueras , C.G.; González-Pastor, J.E. Novel nickel resistance genes from the rhizosphere metagenome of plants adapted to acid mine drainage. Appl. Environ. Microbiol. 2007, 73, 6001–6011. [Google Scholar] [CrossRef]
- Mori, T.; Mizuta, S.; Suenaga, H.; Miyazaki, K. Metagenomic screening for bleomycin resistance genes. Appl. Environ. Microbiol. 2008, 74, 6803–6805. [Google Scholar] [CrossRef]
- Uchiyama, T.; Abe, T.; Ikemura, T.; Watanabe, K. Substrate-induced gene-expression screening of environmental metagenome libraries for isolation of catabolic genes. Nat. Biotechnol. 2005, 23, 88–93. [Google Scholar] [CrossRef]
- Schipper, C.; Hornung, C.; Bijtenhoorn, P.; Quistchau, M.; Grond, S.; Streit, W.R. Metagenome-derived clones encoding two novel lactonase family proteins involved in biofilm inhibition in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2009, 75, 224–233. [Google Scholar] [CrossRef]
- Uchiyama, T.; Watanabe, K. Substrate-induced gene expression (SIGEX) screening of metagenome libraries. Nat. Protoc. 2008, 3, 1202–1212. [Google Scholar] [CrossRef]
- Van Lanen, S.; Shen, B. Microbial genomics for the improvement of natural product discovery. Curr. Opin. Microbiol. 2006, 9, 252–260. [Google Scholar] [CrossRef]
- NCBI Microbial Genomes. Available online: http://www.ncbi.nlm.nhi.gov/genomes/microbial (accessed on 10 July 2013).
- Luo, X.J.; Yu, H.L.; Xu, J.H. Genomic data mining: An efficient way to find new and better enzymes. Enzyme Eng. 2012, 1, 104–108. [Google Scholar]
- Cavicchioli, R.; Amils, R.; Wagner, D.; McGenity, T. Life and applications of extremophiles. Environ. Microbiol. 2011, 13, 1903–1907. [Google Scholar] [CrossRef] [Green Version]
- Pikuta, E.V.; Hoover, R.B.; Tang, J. Microbial Extremophiles at the limit of life. Crit. Rev. Microbiol. 2007, 33, 183–209. [Google Scholar] [CrossRef]
- Atomi, H.; Sato, T.; Kanai, T. Application of hyperthermophiles and their enzymes. Curr. Opin. Biotechnol. 2011, 22, 618–626. [Google Scholar] [CrossRef]
- De Carvalho, C.C. Enzymatic and whole cell catalysis: Finding new strategies for old processes. Biotechnol. Adv. 2011, 29, 75–83. [Google Scholar] [CrossRef]
- Hess, M. Thermoacidophilic proteins for biofuels production. Trends Microbiol. 2008, 16, 414–419. [Google Scholar] [CrossRef]
- Chen, Z.W.; Liu, Y.Y.; Wu, J.F.; She, Q.; Jiang, C.Y.; Liu, S.J. Novel bacterial sulphur oxygenase reductases from bioreactors treating gold-bearing concentrates. Appl. Microbiol. Biotechnol. 2007, 74, 688–698. [Google Scholar] [CrossRef]
- Margesin, R.; Feller, G.; Gerday, C.; Rusell, N. Cold-Adapted Microorganisms: Adaptation Strategies and Biotechnological Potential. In The Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; John Wiley and Sons: New York, NY, USA, 2002; pp. 871–885. [Google Scholar]
- Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef]
- Gomes, I.; Gomes, J.; Steiner, W. Highly thermostable amylase and pullulanase of the extreme thermophilic eubacterium Rhodothermus marinus: Production and partialcharacterization. Bioresour. Technol. 2003, 90, 207–214. [Google Scholar] [CrossRef]
- Van den Burg, B. Extremophiles as a source for novel enzymes. Curr. Opin. Microbiol. 2003, 6, 213–218. [Google Scholar] [CrossRef]
- Shukla, A.; Rana, A.; Kumar, L.; Singh, B.; Ghosh, D. Assessment of detergent activity of Streptococcus sp. AS02 protease isolated from soil of Sahastradhara, Doon Valley, Uttarakhand. Asian J. Microbiol. Biotechnol. Environ. Sci. 2009, 11, 587–591. [Google Scholar]
- Marrs, B.; Delagrave, S.; Murphy, D. Novel approaches for discovering industrial enzymes. Curr. Opin. Microbiol. 1999, 2, 241–245. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Pandy, A. Industrial Biotechnology: Sustainable Growth and Economic Success; Soetaert, W., Vandamme, E.J., Eds.; Wiley VCH Verlag GmbH: Weinheim, Germany, 2010; pp. 207–226. [Google Scholar]
- Cedrone, F.; Menez, A.; Quemeneur, E. Tailoring new enzyme functions by rational redesign. Curr. Opin. Struct. Biol. 2000, 10, 405–410. [Google Scholar] [CrossRef]
- Beppu, T. Modification of milk-clotting aspartic proteinases by recombinant DNA techniques. Ann. N. Y. Acad. Sci. 1990, 613, 14–25. [Google Scholar] [CrossRef]
- Van den Burg, B.; de Kreij, A.; van der Veek, P.; Mansfeld, J.; Venema, G. Engineering an enzyme to resist boiling. Proc. Natl. Acad. Sci. USA 1998, 95, 2056–2060. [Google Scholar] [CrossRef]
- Bolon, D.N.; Voigt, C.A.; Mayo, S.L. De novo design of biocatalysts. Curr. Opin. Struct. Biol. 2002, 6, 125–129. [Google Scholar]
- Shimaoka, M.; Shiftman, J.M.; Jing, H.; Tagaki, J.; Mayo, S.L.; Springer, T.A. Computational design of an integrin I domain stabilized in the open high affinity conformation. Nat. Struct. Biol. 2000, 7, 674–678. [Google Scholar] [CrossRef]
- Turner, N.J. Directed evolution drives the next generation of biocatalysts. Nat. Chem. Biol. 2009, 5, 567–573. [Google Scholar] [CrossRef]
- Schmidt, M.; Boettcher, D.; Bornscheuer, U.T. Industrial Biotechnology: Sustainable Growth and Economic Success; Soetaert, W., Vandamme, E.J., Eds.; Wiley VCH Verlag GmbH: Weinheim, Germany, 2010; pp. 155–187. [Google Scholar]
- Dalby, P.A. Strategy and success for the directed evolution of enzymes. Curr. Opin. Struct. Biol. 2011, 21, 473–480. [Google Scholar] [CrossRef]
- Leung, D.W.; Chen, E.; Goeddel, D.V. A method for random mutagenesis of a defined DNA segment using a modified polymerase chain reaction. Technique 1989, 1, 11–15. [Google Scholar]
- Reidhaar-Olson, J.; Bowie, J.; Breyer, R.M.; Hu, J.C.; Knight, K.L.; Lim, W.A.; Mossing, M.C.; Parsell, D.A.; Shoemaker, K.R.; Sauer, R.T. Random mutagenesis of protein sequences using oligonucleotide cassettes. Methods Enzymol. 1991, 208, 564–586. [Google Scholar]
- Taguchi, S.; Ozaki, A.; Momose, H. Engineering of a cold-adapted protease by sequential random mutagenesis and a screening system. Appl. Environ. Microbiol. 1998, 64, 492–495. [Google Scholar]
- Ness, J.E.; del Cardayre, S.B.; Minshull, J.; Stemmer, W.P. Molecular breeding: The natural approach to protein design. Adv. Protein Chem. 2000, 55, 261–292. [Google Scholar]
- Morley, K.L.; Kazlauskas, R.J. Improving enzyme properties: When are closer mutations better? Trends Biotechnol. 2005, 3, 231–237. [Google Scholar] [CrossRef]
- Reetz, M.T.; Kahakeaw, D.; Lohmer, R. Addressing the numbers problem in directed evolution. ChemBioChem 2008, 9, 1797–1804. [Google Scholar] [CrossRef]
- Muñoz, E.; Deem, M.W. Amino acid alphabet size in protein evolution experiments: Better to search a small library thoroughly or a large library sparsely? Protein Eng. Des. Sel. 2008, 1, 311–317. [Google Scholar]
- Reetz, M.T.; Wu, S. Greatly reduced amino acid alphabets in directed evolution: Making the right choice for saturation mutagenesis at homologous enzyme positions. Chem. Commun. 2008, 5499–5501. [Google Scholar] [CrossRef]
- Crameri, A.; Raillard, S.A.; Bermudez, E.; Stemmer, W.P. DNA shuffling of a family of genes from diverse species acelerates directed evolution. Nature 1998, 391, 288–291. [Google Scholar] [CrossRef]
- Kolkman, J.A.; Stemmer, W.P. Directed evolution of proteins by exon shuffling. Nat. Biotechnol. 2001, 19, 423–428. [Google Scholar] [CrossRef]
- Hackel, B.J.; Kapila, A.; Wittrup, K.D. Picomolar affinity fibronectin domain engineering utilizing loop length diversity, recursive mutagenesis, and loop shuffling. J. Mol. Biol. 2008, 381, 1238–1252. [Google Scholar] [CrossRef]
- Ostermeier, M.; Nixon, A.E.; Shim, J.H.; Benkovic, S.J. Combinatorial protein engineering by incremental truncation. Proc. Natl. Acad. Sci. USA 1999, 96, 3562–3567. [Google Scholar] [CrossRef]
- Fujii, R.; Kitakoa, M.; Hayashi, K. RAISE: A simple and novel method of generating random insertion and deletion mutations. Nucleic Acid Res. 2006, 34, e30. [Google Scholar] [CrossRef]
- Ness, J.E.; Welch, M.; Giver, L.; Bueno, M.; Cherry, J.R.; Borchert, T.V.; Stemmer, W.P.; Minshull, J. DNA shuffling of subgenomic sequences of subtilisin. Nat. Biotechnol. 1999, 17, 893–896. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Reetz, M.T. Directed evolution of enantioselective enzymes for organic chemistry. Curr. Opin. Chem. Biol. 2000, 4, 68–73. [Google Scholar] [CrossRef]
- Suenaga, H.; Mitsokua, M.; Ura, Y.; Watanabe, T.; Furukawa, K. Directed evolution of biphenyl dioxygenase: Emergence of enhanced degradation capacity for benzene, toluene, and alkylbenzenes. J. Bacteriol. 2001, 183, 5441–5444. [Google Scholar] [CrossRef]
- Song, J.K.; Rhee, J.S. Enhancement of stability and activity of phospholipase A(1) in organic solvents by directed evolution. Biochim. Biophys. Acta 2001, 1547, 370–378. [Google Scholar]
- Raillard, S.; Krebber, A.; Chen, Y.; Ness, J.E.; Bermudez, E.; Trinidad, R.; Fullem, R.; Davis, C.; Welch, M.; Seffernick, J.; et al. Novel enzyme activities and functional plasticity revealed by recombining highly homologous enzymes. Chem. Biol. 2001, 8, 891–898. [Google Scholar] [CrossRef]
- Kurtzman, A.L.; Govindarajan, S.; Vahle, K.; Jones, J.T.; Heinrichs, V.; Patten, P.A. Advances in directed protein evolution by recursive genetic recombination: Applications to therapeutic proteins. Curr. Opin. Biotechnol. 2001, 12, 361–370. [Google Scholar] [CrossRef]
- Patten, P.A.; Howard, R.J.; Stemmer, W.P. Applications of DNA shuffling to pharmaceuticals and vaccines. Curr. Opin. Biotechnol. 1997, 8, 724–733. [Google Scholar] [CrossRef]
- Siehl, D.L.; Castle, L.A.; Gorton, R.; Chen, Y.H.; Bertain, S.; Cho, H.J.; Keenan, R.; Liu, D.; Lassner, M.W. Evolution of a microbial acetyl transferase for modification of glyphosate: A novel tolerance strategy. Pest Manag. Sci. 2005, 61, 235–240. [Google Scholar] [CrossRef]
- Tobin, M.B.; Gustafsson, C.; Huisman, G.W. Directed evolution: The “rational” basis for “irrational” design. Curr. Opin. Struct. Biol. 2000, 10, 421–427. [Google Scholar] [CrossRef]
- Cherry, J.R.; Fidanstel, A.L. Directed evolution of industrial enzymes: An update. Curr. Opin. Biotechnol. 2003, 14, 438–443. [Google Scholar] [CrossRef]
- Demain, A.L.; Vaishnav, P. Production of recombinant proteins by microbes and higher organisms. Biotechnol. Adv. 2009, 27, 297–306. [Google Scholar] [CrossRef]
- Swartz, J.R. Escherichia coli Recombinant DNA Technology. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; Neidhardt, F.C., Ed.; American Society of Microbiology Press: Washington, DC, USA, 1996; pp. 1693–1771. [Google Scholar]
- Barnard, G.C.; Henderson, G.E.; Srinivasan, S.; Gerngross, T.U. High level recombinant protein expression in Ralstonia eutropha using T7 RNA polymerase based amplification. Protein Expr. Purif. 2004, 38, 264–271. [Google Scholar] [CrossRef]
- Squires, C.H.; Lucy, P. Vendor voice: A new paradigm for bacterial strain engineering. BioProcess Int. 2008, 6, 22–27. [Google Scholar]
- Romanos, M.A.; Scorer, C.A.; Clare, J.J. Foreign gene expression in yeast: A review. Yeast 1992, 8, 423–488. [Google Scholar] [CrossRef]
- Higgins, D.R.; Cregg, J.M. Introduction to Pichia pastoris. In Pichia Protocols; Higgins, D.R., Cregg, J.M., Eds.; Humana Press: Totowa, NJ, USA, 1998; pp. 1–15. [Google Scholar]
- Zhang, A.L.; Luo, X.J.; Zhang, T.Y.; Pan, Y.N. Recent advances on the GAP promoter-derived expression system of Pichia pastoris. Mol. Biol. Rep. 2009, 36, 1611–1619. [Google Scholar] [CrossRef]
- Hasslacher, M.; Schall, M.; Hayn, M.; Bora, R.; Rumbold, K.; Luecki, J.; Grieng, H.; Kohlwein, S.D.; Schwab, H. High-level intracellular expression of hydroxynitrile lyase from the tropical rubber tree Hevea brassiliensis in microbial hosts. Protein Expr. Purif. 1997, 11, 61–71. [Google Scholar] [CrossRef]
- Werton, M.W.T.; van den Bosch, T.J.; Wind, R.D.; Mooibroek, H.; de Wolf, F.A. High-yield secretion of recombinant gelatins by Pichia pastoris. Yeast 1999, 15, 1087–1096. [Google Scholar] [CrossRef]
- Morrow, K.J. Improving protein production processes. Gen. Eng. Biotechnol. News 2007, 27, 50–54. [Google Scholar]
- Bretthauer, R.K.; Castellino, F.J. Glycosylation of Pichia pastoris-derived proteins. Biotechnol. Appl. Biochem. 1999, 30, 193–200. [Google Scholar]
- Romanos, M.A. Advances in the use of Pichia pastoris for high-level expression. Curr. Opin. Biotechnol. 1995, 6, 527–533. [Google Scholar] [CrossRef]
- Giuseppin, M.; van Eijk, H.M.; Bes, B.C. Molecular regulation of methanol oxidase activity in continuous cultures of Hansenula polymorpha. Biotechnol. Bioeng. 1988, 32, 577–583. [Google Scholar] [CrossRef]
- Egli, T.; van Dijken, J.P.; Veenhuis, M.; Harder, W.; Feichter, A. Methanol metabolism in yeasts: Regulation of the synthesis of catabolite enzymes. Arch. Microbiol. 1980, 124, 115–121. [Google Scholar]
- Shuster, J.R.; Connelley, M.B. Promoter-tagged restriction enzyme-mediated insertion mutagenesis in Aspergillus niger. Mol. Gen. Genet. 1999, 262, 27–34. [Google Scholar]
- Gouka, R.J.; Gerk, C.; Hooykaas, P.J.J.; Bundock, P.; Musters, W.; Verrips, C.T.; de Groot, M.J.A. Transformation of Aspergillus awamori by Agrobacterium tumefaciens-mediated homologous recombination. Nat. Biotechnol. 1999, 6, 598–601. [Google Scholar]
- Van der Hombergh, J.P.; van de Vondervoort, P.J.; van der Heijden, N.C.; Visser, J. New protease mutants in Aspergillus niger result in strongly reduced in vitro degradation of target proteins; genetical and biochemical characterization of seven complementation groups. Curr. Genet. 1997, 28, 299–308. [Google Scholar]
- Gouka, R.J.; Punt, P.J.; van den Hondel, C.A.M.J.J. Efficient production of secreted proteins by Aspergillus: Progress, limitations and prospects. Appl. Microbiol. Biotechnol. 1997, 47, 1–11. [Google Scholar] [CrossRef]
- Moralejo, F.J.; Cardoza, R.E.; Gutierrez, S.; Martín, J.F. Thaumatin production in Aspergillus awamori by use of expression cassettes with strong fungal promoters and high gene dosage. Appl. Environ. Microbiol. 1999, 65, 1168–1174. [Google Scholar]
- Gusakov, A.V.; Salanovich, T.N.; Antonov, A.I.; Ustinov, B.B.; Okunev, O.N.; Burlingame, R.; Emalfarb, M.; Baez, M.; Sinitsyn, A.P. Design of highly efficient cellulase mixtures for enzymatic hydrolysis of cellulose. Biotechnol. Bioeng. 2007, 97, 1028–1038. [Google Scholar] [CrossRef]
- Verdoes, J.C.; Punt, P.J.; Burlingame, R.; Bartels, J.; van Dijk, R.; Slump, E.; Meens, M.; Joosten, R.; Emalfarb, M. A dedicated vector for efficient library construction and high throughput screening in the hyphal fungus Chrysosporium lucknowense. Ind. Biotechnol. 2007, 3, 48–57. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis—Key to sustainable industrial chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef]
- Gong, J.S.; Lu, Z.M.; Li, H.; Shi, J.S.; Zhou, Z.M.; Xu, Z.H. Nitrilases in nitrile biocatalysis: Recent progress and forthcoming research. Microb. Cell Fact. 2012, 11, 142–160. [Google Scholar] [CrossRef]
- Schmid, A.; Dordick, J.S.; Hauer, B.; Kiener, A.; Wubbolts, M.; Witholt, B. Industrial biocatalysis today and tomorrow. Nature 2001, 409, 258–268. [Google Scholar] [CrossRef]
- Ran, N.; Zhao, L.; Chen, Z.; Tao, J. Recent applications of biocatalysis in developing green chemistry for chemical synthesis at the industrial scale. Green Chem. 2008, 10, 361–372. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Huisman, G.W.; Kazlauskas, R.J.; Lutz, S.; Moore, J.C.; Robins, K. Engineering the third wave of biocatalysis. Nature 2012, 485, 185–194. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Enzymes and bioconversions of industrial, pharmaceutical, and biotechnological significance. Org. Process Res. Dev. 2011, 15, 224–230. [Google Scholar] [CrossRef]
- BBC Research. Report BIO030 F Enzymes in Industrial Applications: Global Markets; BBC Research: Wellesley, MA, USA, 2011. [Google Scholar]
- World Enzymes. Freedonia Group: Cleveland, OH, USA, 2011.
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial enzyme applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Araujo, R.; Casal, M.; Cavaco-Paulo, A. Application of enzymes for textiles fibers processing. Biocatal. Biotechnol. 2008, 26, 332–349. [Google Scholar] [CrossRef] [Green Version]
- Tzanov, T.; Calafell, M.; Guebitz, G.M.; Cavaco-Paulo, A. Bio-preparation of cotton fabrics. Enzyme Microb. Technol. 2001, 29, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, A.; del Río, J.C.; Martínez, A.T. Microbial and enzymatic control of pitch in the pulp and paper industry. Appl. Microbiol. Biotechnol. 2009, 82, 1005–1018. [Google Scholar] [CrossRef] [Green Version]
- Farrell, R.L.; Hata, K.; Wall, M.B. Solving pitch problems in pulp and paper processes by the use of enzymes or fungi. Adv. Biochem. Eng. Biotechnol. 1997, 57, 197–212. [Google Scholar]
- Rodriguez-Couto, S.; Toca-Herrera, J.L. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef]
- Rubin, E.M. Genomics of cellulosic biofuels. Nature 2008, 454, 841–845. [Google Scholar] [CrossRef]
- Wilson, B.D. Cellulases and biofuels. Curr. Opin. Biotechnol. 2009, 20, 295–299. [Google Scholar] [CrossRef]
- Zhang, Y.H.P. What is vital (and not vital) to advance economically-competitive biofuels production. Process Biochem. 2011, 46, 2091–2110. [Google Scholar] [CrossRef]
- Zhang, Y.H.P.; Himmmel, M.E.; Mielenz, J.R. Outlook for cellulose improvement: Screening and selection strategies. Biotechnol. Adv. 2006, 24, 452–481. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of lignocellulosic biomass: Biochemical and molecular perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19–31. [Google Scholar] [CrossRef]
- Dashtban, M.; Qin, W. Overexpression of an exotic thermotolerant β-glucosidase in Trichoderma reesei and its significant increase in cellulolytic activity and saccharification of barley straw. Microb. Cell Fact. 2012, 11, 63–78. [Google Scholar] [CrossRef]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequence analysis of the cellulolytic fungus Trichoderma reesei (syn. Hypocrea jecorina) reveals a surprisingly limited inventory of carbohydrate active enzymes. Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef]
- Kanafusa-Shinkai, S.; Wakayama, J.; Tsukamoto, K.; Hayashi, N.; Miyazaki, Y.; Ohmori, H.; Tajima, K.; Yokoyama, H. Degradation of microcrystalline cellulose and non-pretreated plant biomass by a cell-free extracellular cellulose/hemicellulase system from the extreme thermophilic bacterium Caldicellulosiruptor bescii. J. Biosci. Bioeng. 2013, 115, 64–70. [Google Scholar] [CrossRef]
- Feed Enzymes: The Global Scenario; Frost & Sullivan: London, UK, 2007.
- Choct, M. Enzymes for the feed industry: Past, present and future. World Poult. Sci. 2006, 62, 5–15. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Pariza, M.W.; Cook, M. Determining the safety of enzymes used in animal feed. Regul. Toxicol. Pharmcol. 2010, 56, 332–342. [Google Scholar] [CrossRef]
- Mendez, C.; Salas, J.A. Altering the glycosylation pattern of bioactive compounds. Trends Biotechnol. 2001, 19, 449–456. [Google Scholar] [CrossRef]
- Okanishi, M.; Suzuki, N.; Furita, T. Variety of hybrid characters among recombinants obtained by interspecific protoplast fusion in streptomycetes. Biosci. Biotechnol. Biochem. 1996, 6, 1233–1238. [Google Scholar] [CrossRef]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar]
- Dunn-Coleman, N.S.; Bloebaum, P.; Berka, R.; Bodie, E.; Robinson, N.; Armstrong, G.; Ward, M.; Przetak, M.; Carter, G.L.; LaCost, R.; et al. Commercial levels of chymosin production by Aspergillus. Biotechnology 1991, 9, 976–981. [Google Scholar] [CrossRef]
- Bodie, E.A.; Armstrong, G.L.; Dunn-Coleman, N.S. Strain improvement of chymosin-producing strains of Aspergillus niger var awamori using parasexual recombination. Enzyme Microb. Technol. 1994, 16, 376–382. [Google Scholar] [CrossRef]
- Pariza, M.W.; Johnson, E.A. Evaluating the safety of microbial enzyme preparations used in food processing: Update for a new century. Regul. Toxicol. Pharmacol. 2001, 33, 173–186. [Google Scholar] [CrossRef]
- US FDA. Center for Food Safety and Applied Nutrition. Available online: http://vm.cfsan.fda.gov (accessed on 1 July 2013).
- Vandamme, E.J.; Cerdobbel, A.; Soetaert, W. Biocatalysis on the rise. Part 1. Principles. Chem. Today 2005, 23, 57–61. [Google Scholar]
- Collection of Information on Enzymes; Austrian Federal Evironment Agency: Vienna, Austria, 2002.
- Olempska-Beer, Z.S.; Merker, R.I.; Ditto, M.D.; DiNovi, M.J. Food-processing enzymes from recombinant microorganisms—A review. Regul. Toxicol. Pharmcol. 2006, 45, 144–158. [Google Scholar] [CrossRef]
- Tufvesson, P.; Lima-Ramos, J.; Nordblad, M.; Woodley, J.M. Guidelines and cost analysis for catalyst production in biocatalytic processes. Org. Process Res. Dev. 2011, 15, 266–274. [Google Scholar] [CrossRef]
- Panke, S.; Wubbolts, M. Advances in biocatalytic synthesis of pharmaceutical intermediates. Curr. Opin. Chem. Biol. 2005, 9, 188–194. [Google Scholar] [CrossRef]
- Jäckel, C.; Hilvert, D. Biocatalysts by evolution. Curr. Opin. Biotechnol. 2010, 21, 753–759. [Google Scholar] [CrossRef]
- Lutz, S. Reengineering enzymes. Science 2010, 329, 285–287. [Google Scholar] [CrossRef]
- Kim, D.Y.; Rha, E.; Choi, S.-L.; Song, J.J.; Hong, S.-P.; Sung, M.-H.; Lee, S.-G. Development of bioreactor system for L-tyrosine synthesis using thermostable tyrosine phenol-lyase. J. Microb. Biotechnol. 2007, 17, 116–122. [Google Scholar]
- Volpato, G.; Rodrigues, R.C.; Fernandez-Lafuente, R. Use of enzymes in the production of semi-synthetic penicillins and cephalosporins: Drawbacks and perspectives. Curr. Med. Chem. 2010, 17, 3855–3873. [Google Scholar] [CrossRef]
- Kirchner, G.; Scollar, M.P.; Klibanov, A. Resolution of racemic mixtures via lipase catalysis in organic solvents. J. Am. Chem. Soc. 1995, 107, 7072–7076. [Google Scholar] [CrossRef]
- Zheng, G.W.; Xu, J.H. New opportunities for biocatalysis: Driving the synthesis of chiral chemicals. Curr. Opin. Biotechnol. 2011, 22, 784–792. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Asymmetric biocatalysis with microbial enzymes and cells. Curr. Opin. Microbiol. 2010, 13, 283–292. [Google Scholar] [CrossRef]
- Liang, J.; Mundorff, E.; Voladri, R.; Jenne, S.; Gilson, L.; Conway, A.; Krebber, A.; Wong, J.; Huisman, G.; Truesdell, S.; et al. Highly Enantioselective Reduction of a small heterocyclic ketone: Biocatalytic reduction of tetrahydrothiophene-3-one to the corresponding (R)-alcohol. Org. Process Res. Dev. 2010, 14, 185–192. [Google Scholar]
- Ma, S.K.; Gruber, J.; Davis, C.; Newman, L.; Gray, D.; Wang, A.; Grate, J.; Huisman, G.W.; Sheldon, R.A. A green-by-design biocatalytic process for atorvastatin intermediate. Green Chem. 2010, 12, 81–86. [Google Scholar] [CrossRef]
- Sheldom, R.A. E factors, green chemistry and catalysis: An odyssey. Chem. Commun. 2008, 3352–3365. [Google Scholar] [CrossRef]
- Höhne, M.; Schätzle, S.; Jochens, H.; Robins, K.; Bornscheuer, U.T. Rational assignment of key motifs for function guides in silico enzyme identification. Nat. Chem. Biol. 2010, 6, 807–813. [Google Scholar] [CrossRef]
- Koszelewski, D.; Göritzer, M.; Clay, D.; Seisser, B.; Kroutil, W. Synthesis of optically active amines employing recombinant ω-transaminases in E. coli cells. ChemCatChem 2010, 2, 73–77. [Google Scholar] [CrossRef]
- Gooding, O.; Voladri, R.; Bautista, A.; Hopkins, T.; Huisman, G.; Jenne, S.; Ma, S.; Mundorff, E.C.; Savile, M.M.; Truesdell, S.J. Development of a practical biocatalytic process for (R)-2-methylpentanol. Org. Process Res. Dev. 2010, 14, 119–126. [Google Scholar] [CrossRef]
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleits, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef]
- Liang, J.; Lalonde, J.; Borup, B.; Mitchell, V.; Mundorff, E.; Trinh, N.; Kochreckar, D.A.; Cherat, R.N.; Ganesh, P.G. Development of a biocatalytic process as an alternative to the (-)-DIP-Cl-mediated asymmetric reduction of a key intermediate of Montelukast. Org. Process Res. Dev. 2010, 14, 193–198. [Google Scholar] [CrossRef]
- Ritter, S. Green chemistry awards. Honors: Presidential challenge highlights innovations that promote sustainability. Chem. Eng. News 2012, 90, 11. [Google Scholar]
- Ema, T.; Sayaka, I.; Nobuyasu, O.; Sakai, T. Highly efficient chemoenzymatic synthesis of methyl (R)-o-chloromandelate, a key intermediate for clopidogrel, via asymmetric reduction with recombinant Escherichia coli. Adv. Synth. Catal. 2008, 350, 2039–2044. [Google Scholar] [CrossRef]
- Zaks, A.; Dodds, D.R. Application of biocatalysis and biotransformations to the synthesis of pharmaceuticals. Drug Discov. Today 1997, 2, 513–531. [Google Scholar] [CrossRef]
- Lee, M.Y.; Dordick, J.S. Enzyme activation for nonaqueous media. Curr. Opin. Biotechnol. 2002, 13, 376–384. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Adrio, J.L.; Demain, A.L. Microbial Enzymes: Tools for Biotechnological Processes. Biomolecules 2014, 4, 117-139. https://doi.org/10.3390/biom4010117
Adrio JL, Demain AL. Microbial Enzymes: Tools for Biotechnological Processes. Biomolecules. 2014; 4(1):117-139. https://doi.org/10.3390/biom4010117
Chicago/Turabian StyleAdrio, Jose L., and Arnold L. Demain. 2014. "Microbial Enzymes: Tools for Biotechnological Processes" Biomolecules 4, no. 1: 117-139. https://doi.org/10.3390/biom4010117