
Physical Exercise Induces Significant Changes in Immunoglobulin G N-Glycan Composition in a Previously Inactive, Overweight Population
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Protocol
2.2.1. Circular Exercise Program
2.2.2. Cardio Exercise Program
2.2.3. Nordic Walking Program
2.2.4. N-Glycan Analysis from DBS
2.3. Statistical Analysis
3. Results
3.1. Effect of Medium-Intensity Exercise on Motor and Functional Abilities
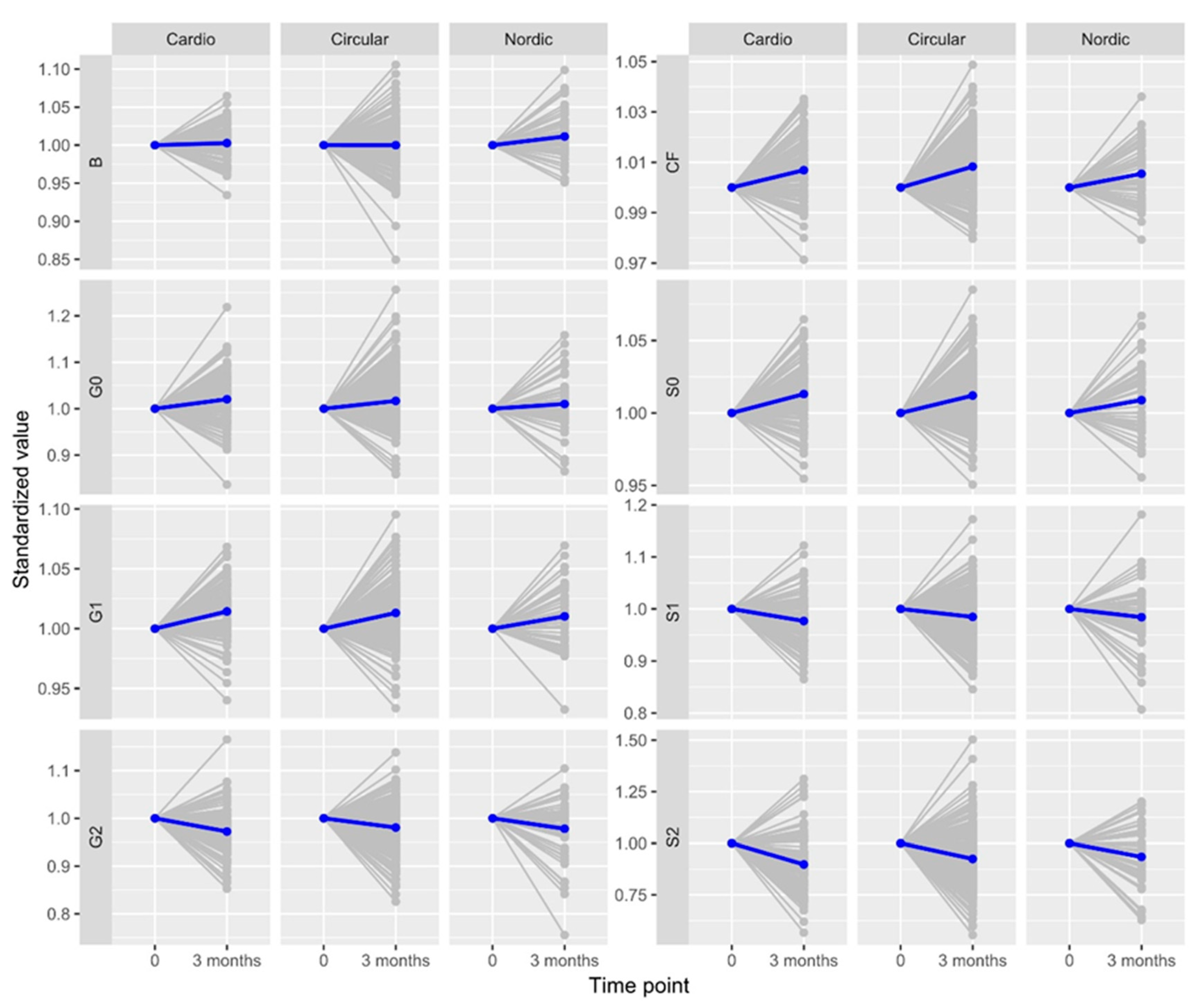
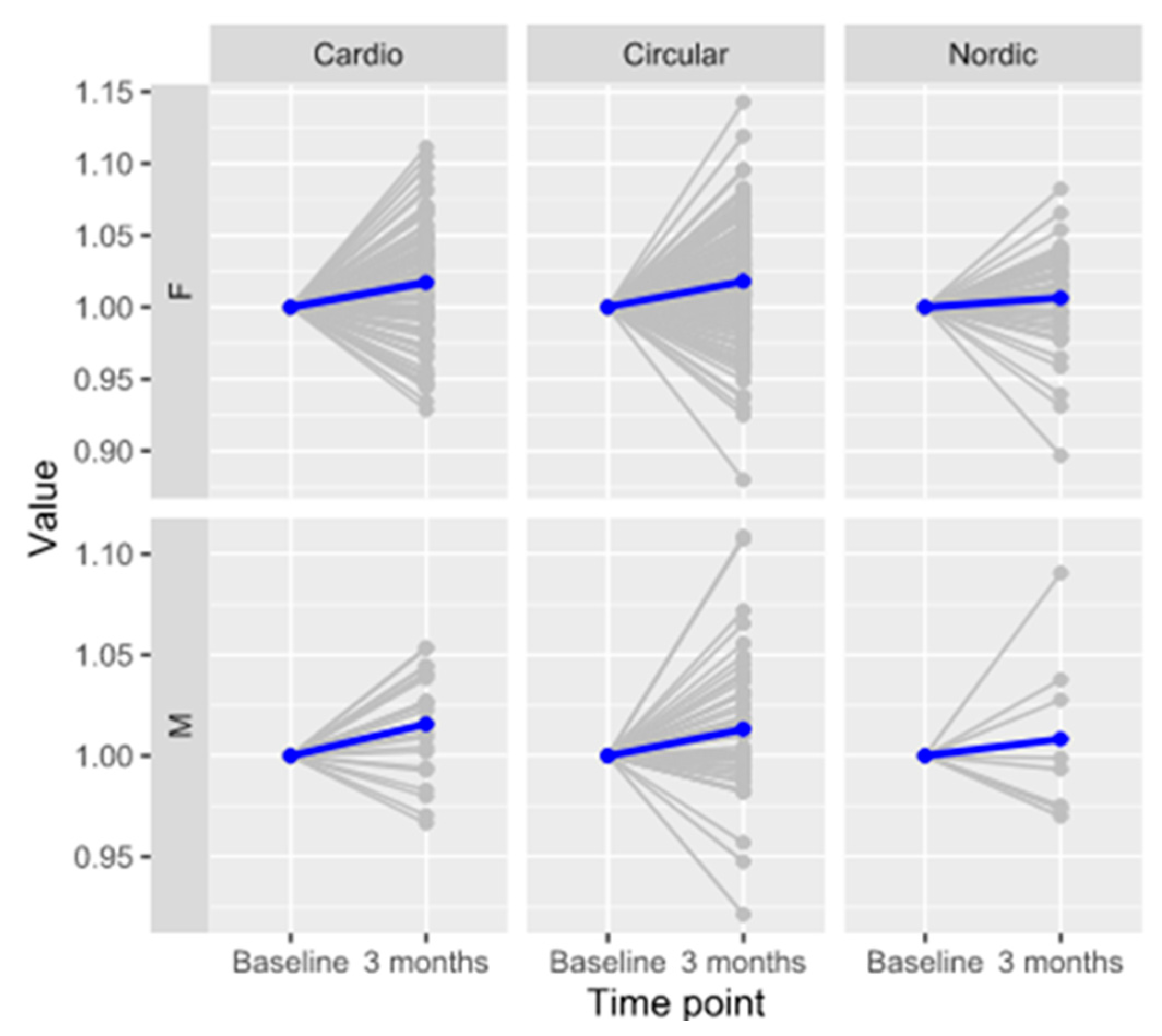
3.2. Effect of Medium-Intensity Exercise on IgG N-Glycosylation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organisation. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Blair, S.N. Physical inactivity: The biggest public health problem of the 21st century. Br. J. Sport. Med. 2009, 43, 1–2. [Google Scholar]
- Bouchard, C.; Blair, S.N.; Haskell, L. Physical Activity and Health, 2nd ed.; Hum Kinetics: Windsor, ON, USA, 2012. [Google Scholar]
- Kohl, H.W.; Lynn Craig, C.; Lambert, V.; Inoue, S.; Ramadan Alkadri, J.; Leetongin, G.; Kahlmeier, S. The pandemic of physical inactivity: Global action for public health. Lancet 2012, 380, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.; Sallis, R.; Hutber, A.; Archer, E. Exercise therapy—The public health message. Scand. J. Med. Sci. Sport. 2012, 22, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Guseman, E.H.; Cauffman, S.P.; Tucker, J.M.; Smith, L.; Eisenmann, J.C.; Stratbucker, W. The association between measures of fitness and metabolic health in treatment- seeking youth with obesity. Metab. Syndr. Relat. Disord. 2017, 15, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.G.; Guedes, D.P. Physical activity, Sedentary behaviour, Cardiorespiratory fitness and metabolic syndrome in adolescent, systematic review and meta-analysis of observational evidence. PLoS ONE 2016, 11, e0168503. [Google Scholar] [CrossRef]
- American College of Sports Medicine. Quantity and Quality of Exercise for Developing and Maintaining Cardiorespiratory, Musculoskeletal, and Neuromotor Fitness in Apparently Healthy Adults: Guidance for Prescribing Exercise. Med. Sci. Sport. Exerc. 2011, 43, 1334–1359. [Google Scholar] [CrossRef]
- Booth, F.W.; Chakravarth, M.V.; Gordon, S.E.; Spangenburg, E.E. Waging war on physical inactivity: Using modern molecular ammunition against an ancient enemy. J. Appl. Physiol. 2002, 93, 3–30. [Google Scholar] [CrossRef]
- Booth, F.W.; Roberts, C.K.; Laye, M.J. Lack of exercise is a major cause of chronic diseases. Compr. Physiol. 2012, 2, 1143–1211. [Google Scholar]
- Myers, J.; Prakash, M.; Froelicher, V.; Do, D.; Partington, S.; Atwood, J.E. Exercise capacity and mortality among men referred for exercise testing. N. Engl. J. Med. 2002, 346, 793–801. [Google Scholar] [CrossRef]
- Gupta, S.; Rohatgi, A.; Ayers, C.R.; Willis, B.L.; Haskell, W.L.; Khera, A.; Drazner, M.H.; de Lemos, J.A.; Berry, J.D. Cardiorespiratory fitness and classification of risk of cardiovascular disease mortality. Circulation 2011, 123, 1377–1383. [Google Scholar] [CrossRef]
- Schneider, J. Age dependency of oxygen uptake and related parameters in exercise testing: An expert opinion on reference values suitable for adults. Lung 2013, 191, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Gibbons, L.W.; Kampert, J.B.; Nichaman, M.Z.; Blair, S.N. Low cardiorespiratory fitness and physical inactivity as predictors of mortality in men with type 2 diabetes. Ann. Intern. Med. 2000, 132, 605–611. [Google Scholar] [CrossRef]
- Holloszy, J.O. Exercise-induced increase in muscle insulin sensitivity. J. Appl. Physiol. 2005, 99, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Lowder, T.W.; Spielmann, G.; Bigley, A.B.; LaVoy, E.C.; Kunz, H. Exercise and the aging immune system. Ageing Res. Rev. 2012, 11, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M. Effects of exercise on immune function. Sport. Sci. Exch. 2015, 28, 1–6. [Google Scholar]
- Campbell, J.P.; Turner, J.E. Debunking the Myth of Exercise- Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health Across the Lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Antunes, B.M.; Campos, E.Z.; dos Santos, R.V.T.; Rosa-Neto, J.C.; Franchini, E.; Bishop, N.C.; Lira, F.S. Anti-inflammatory response to acute exercise is related with intensity and physical fitness. J. Cell Biochem. 2018, 120, 5333–5342. [Google Scholar] [CrossRef]
- Suzuki, K. Cytokine Response to Exercise and Its Modulation. Antioxidants 2018, 7, 17. [Google Scholar] [CrossRef]
- Terra, R.; da Silva, S.A.G.; Salerno Pinto, V.; Lourenço Dutr, P.M. Effect of exercise on the immune system: Response, adaptation and cell signaling. Rev. Bras. Med. Esport 2012, 18, 208–214. [Google Scholar] [CrossRef]
- Guimarães, T.T.; Terra, R.; Lourenço Dutra, P.M. Chronic effects of exhausting exercise and overtraining on the immune response: Th1 and Th2 profile. Motricidade 2017, 13, 69–78. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G glycosylation in aging and diseases. Cell. Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Lauc, G. Precision medicine that transcends genomics: Glycans as integrators of genes and environment. BBA Gen. Subj. 2016, 1860, 1571–1573. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, X.; Dias, A.M.; Colombel, J.F.; Vermeire, S.; Van Vlierberghe, H.; Callewaert, N.; Pinho, S.S. Protein Glycosylation as a Diagnostic and Prognostic Marker of Chronic Inflammatory Gastrointestinal and Liver Diseases. Gastroenterology 2020, 158, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front. Med. 2018, 5, 61. [Google Scholar] [CrossRef]
- Štambuk, T.; Lauc, G. N-Glycans and Diabetes. In Encyclopedia of Cell Biology, 2nd ed.; Bradshaw, R., Stahl, P., Hart, G., Eds.; Academic Press: Cambridge, MA, USA, 2023; Volume 4, pp. 506–515. [Google Scholar]
- Gudelj, I.; Lauc, G. Glycans and Cardiovascular Diseases. In Encyclopedia of Cell Biology, 2nd ed.; Bradshaw, R., Stahl, P., Hart, G., Eds.; Academic Press: Cambridge, MA, USA, 2023; Volume 4, pp. 385–395. [Google Scholar]
- Vilaj, M.; Gudelj, I.; Trbojević-Akmačić, I.; Lauc, G.; Pezer, M. IgG Glycans as a biomarker of biological age. In Biomarkers of Human Aging. Healthy Ageing and Longevity; Moskalev, A., Ed.; Springer: Cham, Switzerland, 2019; Volume 10. [Google Scholar]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panagiota Panourgia, M.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Flynn, M.G.; Markofski, M.M.; Carrillo, A.E. Elevated Inflammatory Status and Increased Risk of Chronic Disease in Chronological Aging: Inflamm-aging or Inflamm-inactivity? Aging Dis. 2019, 10, 147–156. [Google Scholar] [CrossRef]
- Russell, A.C.; Kepka, A.; Trbojević-Akmačić, I.; Ugrina, I.; Song, M.; Hui, J.; Hunter, M.; Laws, S.M.; Lauc, G.; Wang, W. Increased central adiposity is associated with pro-inflammatory immunoglobulin G N-glycans. Immunobiology 2019, 224, 110–115. [Google Scholar] [CrossRef]
- Greto, V.L.; Cvetko, A.; Štambuk, T.; Dempster, N.J.; Kifer, D.; Deriš, H.; Cindrić, A.; Vučković, F.; Falchi, M.; Gillies, R.S.; et al. Extensive weight loss reduces glycan age by altering IgG N-glycosylation. Int. J. Obes. 2021, 45, 1521–1531. [Google Scholar] [CrossRef]
- Deriš, H.; Tominac, P.; Vučković, F.; Briški, N.; Astrup, A.; Blaak, E.E.; Lauc, G.; Gudelj, I. Effects of low-calorie and different weight-maintenance diets on IgG glycomecomposition. Front. Immunol. 2022, 13, 995186. [Google Scholar] [CrossRef]
- Sarin, H.V.; Gudelj, I.; Honkane, J.; Ihalainen, J.K.; Vuorela, A.; Le, J.H.; Jin, Z.; Terwilliger, J.D.; Isola, V.; Ahtiainen, J.P.; et al. Molecular Pathways MediatingImmunosuppression in Response to Prolonged Intensive Physical Training, Low-Energy Availability, and Intensive Weight Loss. Front. Immunol. 2019, 10, 907. [Google Scholar] [CrossRef]
- Tijardović, M.; Marijančević, D.; Bok, D.; Kifer, D.; Lauc, H.; Gornik, O.; Keser, T. Intense Physical Exercise Induces an Anti-inflammatory Change in IgG N-Glycosylation Profile. Front. Physiol. 2019, 10, 1522. [Google Scholar] [CrossRef] [PubMed]
- Šimunović, J.; Vilaj, M.; Trbojević-Akmačić, I.; Momčilović, A.; Vučković, F.; Gudelj, I.; Jurić, J.; Nakić, N.; Lauc, G.; Pezer, M. Comprehensive N-glycosylation analysis of immunoglobulin G from dried blood spots. Glycobiology 2019, 29, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Pučić, M.; Knežević, A.; Vidić, J.; Adamczyk, B.; Novokmet, M.; Polašek, O.; Gornik, O.; Supraha-Goreta, S.; Wormald, M.R.; Redzić, I.; et al. High throughput isolation and glycosylation analysis of IgG-variability and heritability of the IgG glycome in three isolated human populations. Mol. Cell Proteom. 2011, 10, M111.010090. [Google Scholar] [CrossRef] [PubMed]
- Trbojević-Akmačić, I.; Ugrina, I.; Lauc, G. Comparative Analysis and Validation of Different Steps in Glycomics Studies. Methods Enzymol. 2017, 586, 37–55. [Google Scholar]
- Birukov, A.; Plavša, B.; Eichelmann, F.; Kuxhaus, O.; Hoshi, R.A.; Rudman, N.; Štambuk, T.; Trbojević-Akmačić, I.; Schiborn, C.; Morze, J.; et al. Immunoglobulin G N-Glycosylation Signatures in Incident Type 2 Diabetes and Cardiovascular Disease. Diabetes Care 2022, 45, 2729–2736. [Google Scholar] [CrossRef]
- Wittenbecher, C.; Štambuk, T.; Kuxhaus, O.; Rudman, N.; Vučković, F.; Štambuk, J.; Schiborn, C.; Rahelić, D.; Dietrich, S.; Gornik, O.; et al. Plasma N-Glycans as Emerging Biomarkers of Cardiometabolic Risk: A Prospective Investigation in the EPIC-Potsdam Cohort Study. Diabetes Care 2020, 43, 661–668. [Google Scholar] [CrossRef]
- Plomp, R.; Ruhaak, L.R.; Uh, H.W.; Reiding, K.R.; Selman, M.; Houwing-Duistermaat, J.J.; Slagboom, P.E.; Beekman, M.; Wuhrer, M. Subclass-specific IgG glycosylation is associated with markers of inflammation and metabolic health. Sci. Rep. 2017, 7, 12325. [Google Scholar] [CrossRef]
- Zahavi, D.; AlDeghaither, D.; O’Connell, A.; Weiner, L.M. Enhancing antibody-dependent cell-mediated cytotoxicity: A strategy for improving antibody-based immunotherapy. Antib. Ther. 2018, 1, 7–12. [Google Scholar] [CrossRef]
- Takahashi, M.; Kuroki, Y.; Ohtsubo, K.; Taniguchi, N. Core fucose and bisecting GlcNAc, the direct modifiers of the N-glycan core: Their functions and target proteins. Carbohydr. Res. 2009, 344, 1387–1390. [Google Scholar] [CrossRef]
- Kiyoshi, M.; Tsumoto, K.; Ishii-Watabe, A.; Caaveiro, J.M.M. Glycosylation of IgG-Fc: A molecular perspective. Int. Immunol. 2017, 29, 311–317. [Google Scholar] [CrossRef]
- Feigenbaum, M.S.; Pollock, M.L. Prescription of resistance training for health and disease. Med. Sci. Sport. Exerc. 1999, 31, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Winnet, R.A.; Carpinelli, R.N. Potential Health-Related Benefits of Resistance Training. Prevent Med. 2001, 33, 503–513. [Google Scholar] [CrossRef]
- Figard- Fabre, H.; Fabre, N.; Leonardi, A. Efficacy of Nordic Walking in Obesity Management. Int. J. Sport. Med. 2011, 32, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.S.; Dhaliwal, S.S.; Hills, A.P.; Pal, S. The effect of 12 weeks of aerobic, resistance or combination exercise training on cardiovascular risk factors in the overweight and obese in a randomized trial. BMC Public Health 2012, 12, 704. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variable | Circular Training | Cardio Training | Nordic Walking | Interaction TIME*GROUP | |||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 1 | 2 | 1 | 2 | ||||
| F | p | Partial ŋ2 | |||||||
| Lunges | 12.20 ± 3.22 | 16.36 ± 3.24 | 12.65 ± 4.00 | 15.95 ± 2.83 | 11.34 ± 2.20 | 15.57 ± 3.53 | 3.80 | 0.02 * | 0.02 |
| Wall sit | 46.76 ± 27.10 | 76.20 ± 39.79 | 46.27 ± 34.02 | 76.33 ± 40.59 | 49.64 ± 24.26 | 74.12 ± 28.43 | 0.73 | 0.48 | 0.00 |
| Squats | 34.52 ± 9.45 | 46.59 ± 10.36 | 35.87 ± 9.01 | 44.86 ± 10.15 | 32.75 ± 7.81 | 46.08 ± 10.35 | 8.03 | <0.01 * | 0.04 |
| Push-ups | 23.25 ± 8.64 | 30.03 ± 8.96 | 21.30 ± 10.23 | 25.69 ± 9.86 | 23.68 ± 7.34 | 27.53 ± 7.48 | 7.22 | <0.01 * | 0.04 |
| Plank | 58.97 ± 32.57 | 118.59 ± 58.67 | 57.85 ± 28.54 | 106.09 ± 48.04 | 52.28 ± 21.15 | 86.34 ± 41.41 | 9.82 | <0.01 * | 0.05 |
| Beep test | 5.40 ± 2.13 | 6.85 ± 2.53 | 5.22 ± 1.64 | 6.38 ± 2.07 | 4.60 ± 1.38 | 5.92 ± 1.97 | 2.25 | 0.11 | 0.01 |
| Lunges | ||||||||
| Interaction | Program | Time | {1} | {2} | {3} | {4} | {5} | {6} |
| 1 | CARDIO | 1 | <0.01 * | 0.82 | <0.01 * | 0.15 | <0.01 * | |
| 2 | CARDIO | 2 | <0.01 * | <0.01 * | 0.90 | <0.01 * | 0.98 | |
| 3 | CIRCULAR | 1 | 0.82 | <0.01 * | <0.01 * | 0.52 | <0.01 * | |
| 4 | CIRCULAR | 2 | <0.01 * | 0.90 | <0.01 * | <0.01 * | 0.60 | |
| 5 | NORDIC | 1 | 0.15 | <0.01 * | 0.52 | <0.01 * | <0.01 * | |
| 6 | NORDIC | 2 | <0.01 * | 0.98 | <0.01 * | 0.60 | <0.01 * | |
| Squats | ||||||||
| Interaction | Program | Time | {1} | {2} | {3} | {4} | {5} | {6} |
| 1 | CARDIO | 1 | <0.01 * | 0.82 | <0.01 * | 0.38 | <0.01 * | |
| 2 | CARDIO | 2 | <0.01 * | <0.01 * | 0.68 | <0.01 * | 0.98 | |
| 3 | CIRCULAR | 1 | 0.82 | <0.01 * | <0.01 * | 0.84 | <0.01 * | |
| 4 | CIRCULAR | 2 | <0.01 * | 0.68 | <0.01 * | <0.01 * | 1.00 | |
| 5 | NORDIC | 1 | 0.38 | <0.01 * | 0.84 | <0.01 * | <0.01 * | |
| 6 | NORDIC | 2 | <0.01 * | 0.98 | <0.01 * | 1.00 | <0.01 * | |
| Push-ups | ||||||||
| Interaction | Program | Time | {1} | {2} | {3} | {4} | {5} | {6} |
| 1 | CARDIO | 1 | <0.01 * | 0.52 | <0.01 * | 0.68 | <0.01 * | |
| 2 | CARDIO | 2 | <0.01 * | 0.10 | <0.01 * | 0.66 | 0.89 | |
| 3 | CIRCULAR | 1 | 0.52 | 0.10 | <0.01 * | 1.00 | 0.02 * | |
| 4 | CIRCULAR | 2 | <0.01 * | <0.01 * | <0.01 * | <0.01 * | 0.44 | |
| 5 | NORDIC | 1 | 0.68 | 0.66 | 1.00 | <0.01 * | <0.01 * | |
| 6 | NORDIC | 2 | <0.01 * | 0.89 | 0.02 * | 0.44 | <0.01 * | |
| Plank | ||||||||
| Interaction | Program | Time | {1} | {2} | {3} | {4} | {5} | {6} |
| 1 | CARDIO | 1 | <0.01 * | 1.00 | <0.01 * | 0.97 | <0.01 * | |
| 2 | CARDIO | 2 | <0.01 * | <0.01 * | 0.13 | <0.01 * | 0.07 | |
| 3 | CIRCULAR | 1 | 1.00 | <0.01 * | <0.01 * | 0.92 | <0.01 * | |
| 4 | CIRCULAR | 2 | <0.01 * | 0.13 | <0.01 * | <0.01 * | <0.01 * | |
| 5 | NORDIC | 1 | 0.97 | <0.01 * | 0.92 | <0.01 * | <0.01 * | |
| 6 | NORDIC | 2 | <0.01 * | 0.07 | <0.01 * | <0.01 * | <0.01 * | |
| CIRCULAR (N = 229) | |||
| Trait | Effect (s.d.) | SE | p-value (adjusted) |
| G0 | 0.040 | 0.015 | 1.12 × 10−2 |
| G1 | 0.258 | 0.028 | 1.55 × 10−16 |
| G2 | −0.094 | 0.019 | 2.13 × 10−6 |
| B | −0.026 | 0.016 | 1.32 × 10−1 |
| CF | 0.523 | 0.050 | 1.07 × 10−20 |
| S0 | 0.258 | 0.032 | 7.01 × 10−14 |
| S1 | −0.085 | 0.020 | 5.63 × 10−5 |
| S2 | −0.500 | 0.058 | 3.06 × 10−15 |
| CARDIO (N = 115) | |||
| Trait | Effect (s.d.) | SE | p-value (adjusted) |
| G0 | 0.067 | 0.026 | 1.61 × 10−2 |
| G1 | 0.316 | 0.047 | 1.65 × 10−9 |
| G2 | −0.151 | 0.030 | 2.73 × 10−6 |
| B | 0.006 | 0.021 | 7.93 × 10−1 |
| CF | 0.521 | 0.088 | 6.55 × 10−8 |
| S0 | 0.277 | 0.044 | 1.17 × 10−8 |
| S1 | −0.128 | 0.027 | 8.74 × 10−6 |
| S2 | −0.628 | 0.076 | 9.03 × 10−13 |
| NORDIC (N = 53) | |||
| Trait | Effect (s.d.) | SE | p-value (adjusted) |
| G0 | 0.002 | 0.034 | 9.52 × 10−1 |
| G1 | 0.186 | 0.058 | 3.56 × 10−3 |
| G2 | −0.126 | 0.050 | 1.86 × 10−2 |
| B | 0.086 | 0.041 | 5.39 × 10−2 |
| CF | 0.406 | 0.118 | 1.77 × 10−3 |
| S0 | 0.181 | 0.075 | 2.54 × 10−2 |
| S1 | −0.075 | 0.054 | 1.89 × 10−1 |
| S2 | −0.445 | 0.124 | 1.16 × 10−3 |
| Program | Sex | Effect (s.d.) | SE | p (Adjusted) | N |
|---|---|---|---|---|---|
| Cardio | M | 0.260 | 0.087 | 9.26 × 10−3 | 24 |
| F | 0.139 | 0.037 | 4.20 × 10−4 | 91 | |
| Circular | M | 0.124 | 0.055 | 3.53 × 10−2 | 54 |
| F | 0.151 | 0.022 | 2.64 × 10−10 | 177 | |
| Nordic walking | M | 0.053 | 0.299 | 8.86 × 10−1 | 8 |
| F | 0.058 | 0.043 | 2.01 × 10−1 | 45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimunić-Briški, N.; Zekić, R.; Dukarić, V.; Očić, M.; Frkatović-Hodžić, A.; Deriš, H.; Lauc, G.; Knjaz, D. Physical Exercise Induces Significant Changes in Immunoglobulin G N-Glycan Composition in a Previously Inactive, Overweight Population. Biomolecules 2023, 13, 762. https://doi.org/10.3390/biom13050762
Šimunić-Briški N, Zekić R, Dukarić V, Očić M, Frkatović-Hodžić A, Deriš H, Lauc G, Knjaz D. Physical Exercise Induces Significant Changes in Immunoglobulin G N-Glycan Composition in a Previously Inactive, Overweight Population. Biomolecules. 2023; 13(5):762. https://doi.org/10.3390/biom13050762
Chicago/Turabian StyleŠimunić-Briški, Nina, Robert Zekić, Vedran Dukarić, Mateja Očić, Azra Frkatović-Hodžić, Helena Deriš, Gordan Lauc, and Damir Knjaz. 2023. "Physical Exercise Induces Significant Changes in Immunoglobulin G N-Glycan Composition in a Previously Inactive, Overweight Population" Biomolecules 13, no. 5: 762. https://doi.org/10.3390/biom13050762