
VEGFA Isoforms as Pro-Angiogenic Therapeutics for Cerebrovascular Diseases


Abstract
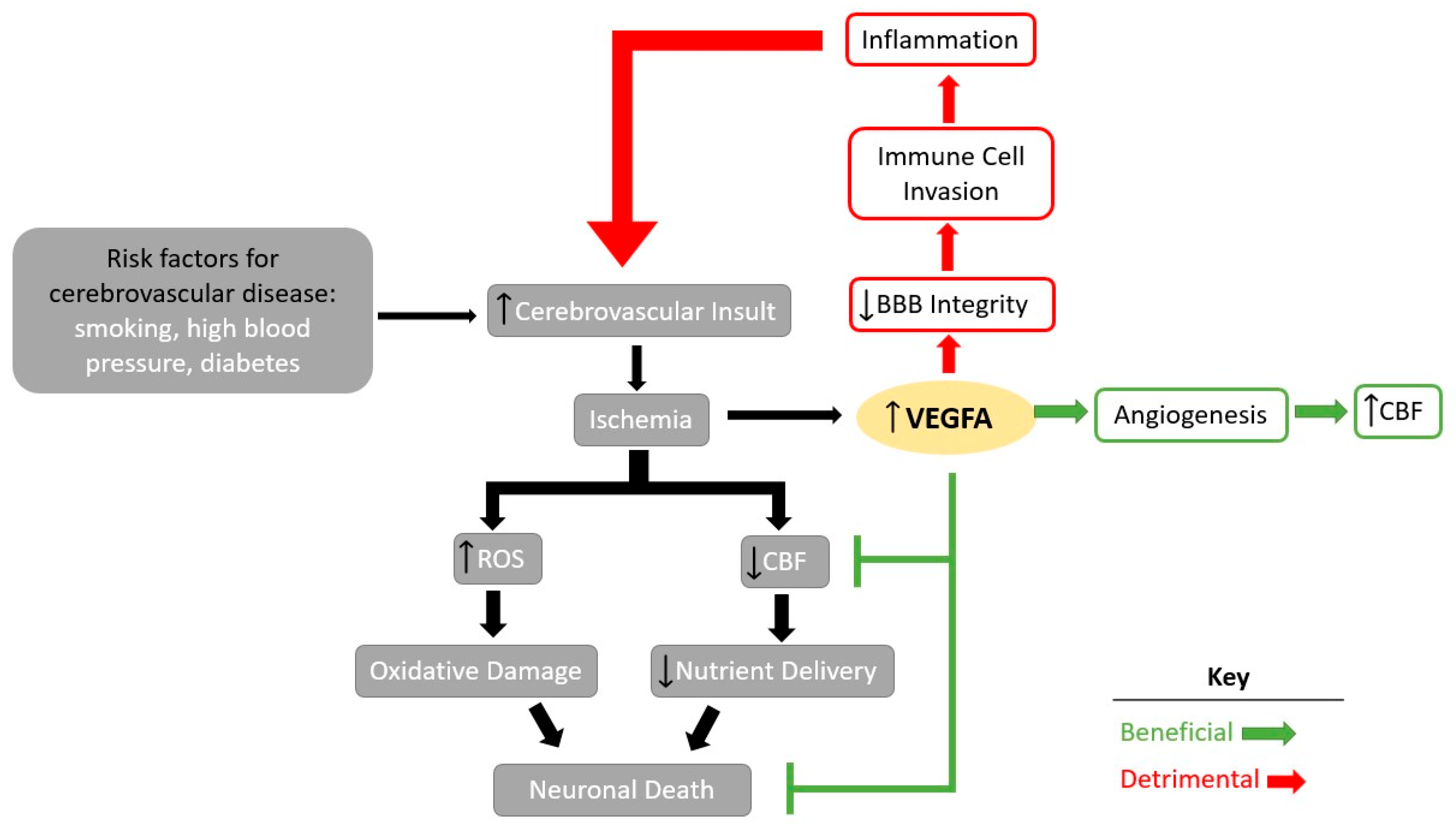
:1. Introduction
2. Therapeutic Use of VEGFA
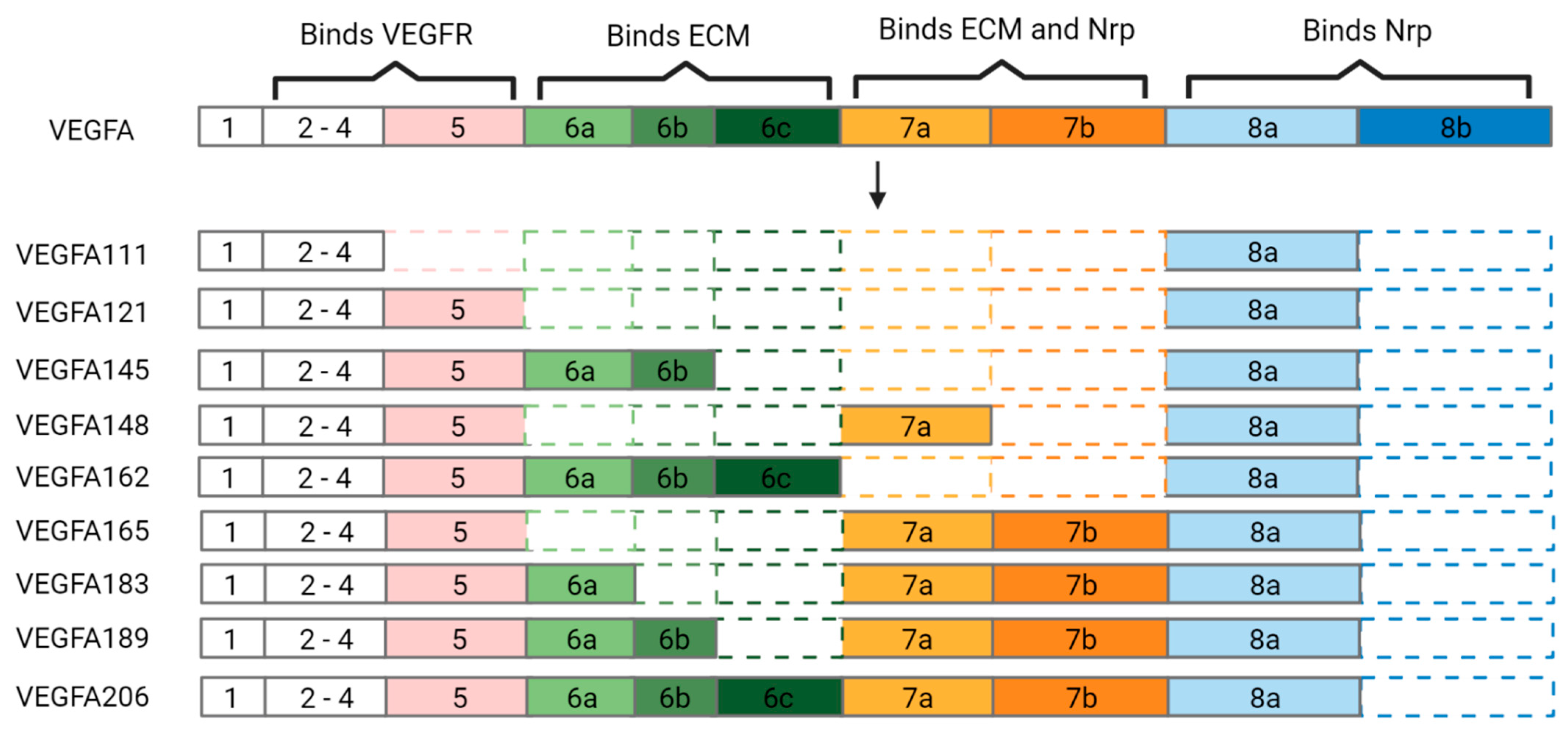
3. VEGFA Isoforms as Therapeutics
3.1. VEGFA111
3.2. VEGFA121
3.3. VEGFA145
3.4. VEGFA148
3.5. VEGFA162
3.6. VEGFA165
3.7. VEGFA183
3.8. VEGFA189
3.9. VEGFA206
4. VEGFA Pro-Angiogenic Clinical Trials
5. Improving VEGFA Administration: Timing, Route, and Dosage
6. Considerations for the Future
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goyal, M.; Raichle, M.E. Glucose Requirements of the Developing Human Brain. J. Craniofacial Surg. 2018, 66 (Suppl. 3), S46–S49. [Google Scholar] [CrossRef] [PubMed]
- Mergenthaler, P.; Lindauer, U.; Dienel, G.A.; Meisel, A. Sugar for the brain: The role of glucose in physiological and pathological brain function. Trends Neurosci. 2013, 36, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Role of vascular endothelial growth factor in the regulation of angiogenesis. Kidney Int. 1999, 56, 794–814. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Vascular endothelial growth factor and its receptor system: Physiological functions in angiogenesis and pathological roles in various diseases. J. Biochem. 2012, 153, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Brain angiogenesis in developmental and pathological processes: Therapeutic aspects of vascular endothelial growth factor. FEBS J. 2009, 276, 4636–4643. [Google Scholar] [CrossRef]
- Geiseler, S.; Morland, C. The Janus Face of VEGF in Stroke. Int. J. Mol. Sci. 2018, 19, 1362. [Google Scholar] [CrossRef]
- Henry, T.D.; Annex, B.H.; McKendall, G.R.; Azrin, M.A.; Lopez, J.J.; Giordano, F.J.; Shah, P.; Willerson, J.T.; Benza, R.L.; Berman, D.S.; et al. The VIVA trial: Vascular endothelial growth factor in Ischemia for Vascular Angiogenesis. Circulation 2003, 107, 1359–1365. [Google Scholar] [CrossRef]
- Lange, C.; Storkebaum, E.; de Almodovar, C.R.; Dewerchin, M.; Carmeliet, P. Vascular endothelial growth factor: A neurovascular target in neurological diseases. Nat. Rev. Neurol. 2016, 12, 439–454. [Google Scholar] [CrossRef]
- Peach, C.J.; Mignone, V.W.; Arruda, M.A.; Alcobia, D.C.; Hill, S.J.; Kilpatrick, L.E.; Woolard, J. Molecular Pharmacology of VEGF-A Isoforms: Binding and Signalling at VEGFR2. Int. J. Mol. Sci. 2018, 19, 1264. [Google Scholar] [CrossRef]
- Mamer, S.B.; Wittenkeller, A.; Imoukhuede, P.I. VEGF-A splice variants bind VEGFRs with differential affinities. Sci. Rep. 2020, 10, 14413. [Google Scholar] [CrossRef]
- Cudmore, M.J.; Hewett, P.W.; Ahmad, S.; Wang, K.-Q.; Cai, M.; Al-Ani, B.; Fujisawa, T.; Ma, B.; Sissaoui, S.; Ramma, W.; et al. The role of heterodimerization between VEGFR-1 and VEGFR-2 in the regulation of endothelial cell homeostasis. Nat. Commun. 2012, 3, 972. [Google Scholar] [CrossRef] [PubMed]
- Kühn, C.; Checa, S. Computational Modeling to Quantify the Contributions of VEGFR1, VEGFR2, and Lateral Inhibition in Sprouting Angiogenesis. Front. Physiol. 2019, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Favier, B.; Alam, A.; Barron, P.; Bonnin, J.; Laboudie, P.; Fons, P.; Mandron, M.; Herault, J.-P.; Neufeld, G.; Savi, P.; et al. Neuropilin-2 interacts with VEGFR-2 and VEGFR-3 and promotes human endothelial cell survival and migration. Blood 2006, 108, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Kofler, N.; Simons, M. The expanding role of neuropilin. Curr. Opin. Hematol. 2016, 23, 260–267. [Google Scholar] [CrossRef]
- Dallinga, M.G.; Habani, Y.I.; Schimmel, A.W.M.; Dallinga-Thie, G.M.; van Noorden, C.J.F.; Klaassen, I.; Schlingemann, R.O. The Role of Heparan Sulfate and Neuropilin 2 in VEGFA Signaling in Human Endothelial Tip Cells and Non-Tip Cells during Angiogenesis In Vitro. Cells 2021, 10, 926. [Google Scholar] [CrossRef]
- Domingues, A.; Fantin, A. Neuropilin 1 Regulation of Vascular Permeability Signaling. Biomolecules 2021, 11, 666. [Google Scholar] [CrossRef]
- Becker, P.M.; Waltenberger, J.; Yachechko, R.; Mirzapoiazova, T.; Sham, J.S.; Lee, C.G.; Elias, J.A.; Verin, A.D. Neuropilin-1 Regulates Vascular Endothelial Growth Factor–Mediated Endothelial Permeability. Circ. Res. 2005, 96, 1257–1265. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Gan, L.; Fagerholm, P.; Palmblad, J. Vascular endothelial growth factor (VEGF) and its receptor VEGFR-2 in the regulation of corneal neovascularization and wound healing. Acta Ophthalmol. Scand. 2004, 82, 557–563. [Google Scholar] [CrossRef]
- Liu, S.-M.; Xiao, Z.-F.; Li, X.; Zhao, Y.-N.; Wu, X.-M.; Han, J.; Chen, B.; Li, J.-Y.; Fan, C.-X.; Xu, B.; et al. Vascular endothelial growth factor activates neural stem cells through epidermal growth factor receptor signal after spinal cord injury. CNS Neurosci. Ther. 2018, 25, 375–385. [Google Scholar] [CrossRef]
- Tong, X.; Yang, Q.; Ritchey, M.D.; George, M.G.; Jackson, S.L.; Gillespie, C.; Merritt, R.K. The Burden of Cerebrovascular Disease in the United States. Prev. Chronic Dis. 2019, 16, E52. [Google Scholar] [CrossRef] [PubMed]
- Olmez, I.; Ozyurt, H. Reactive oxygen species and ischemic cerebrovascular disease. Neurochem. Int. 2011, 60, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Slevin, M.; Krupinski, J.; Slowik, A.; Kumar, P.; Szczudlik, A.; Gaffney, J. Serial measurement of vascular endothelial growth factor and transforming growth factor-β1 in serum of patients with acute ischemic stroke. Stroke 2000, 31, 1863–1870. [Google Scholar] [CrossRef] [PubMed]
- Lennmyr, F.; Ata, K.A.; Funa, K.; Olsson, Y.; Terént, A. Expression of Vascular Endothelial Growth Factor (VEGF) and its Receptors (Flt-1 and Flk-1) following Permanent and Transient Occlusion of the Middle Cerebral Artery in the Rat. J. Neuropathol. Exp. Neurol. 1998, 57, 874–882. [Google Scholar] [CrossRef]
- Grammas, P.; Sanchez, A.; Tripathy, D.; Luo, E.; Martinez, J. Vascular signaling abnormalities in Alzheimer disease. Clevel. Clin. J. Med. 2011, 78, S50–S53. [Google Scholar] [CrossRef]
- Thau-Zuchman, O.; Shohami, E.; Alexandrovich, A.G.; Leker, R.R. Vascular Endothelial Growth Factor Increases Neurogenesis after Traumatic Brain Injury. J. Cereb. Blood Flow Metab. 2010, 30, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Alrafiah, A.; Alofi, E.; Almohaya, Y.; Hamami, A.; Qadah, T.; Almaghrabi, S.; Hakami, N.; Alrawaili, M.S.; Tayeb, H.O. Angiogenesis Biomarkers in Ischemic Stroke Patients. J. Inflamm. Res. 2021, 14, 4893–4900. [Google Scholar] [CrossRef]
- Pikula, A.; Beiser, A.S.; Chen, T.C.; Preis, S.R.; Vorgias, D.; DeCarli, C.; Au, R.; Kelly-Hayes, M.; Kase, C.S.; Wolf, P.A.; et al. Serum brain-derived neurotrophic factor and vascular endothelial growth factor levels are associated with risk of stroke and vascular brain injury: Framingham Study. Stroke 2013, 44, 2768–2775. [Google Scholar] [CrossRef]
- Ghori, A.; Prinz, V.; Nieminen-Kehlä, M.; Bayerl, S.H.; Kremenetskaia, I.; Riecke, J.; Krechel, H.; Broggini, T.; Scherschinski, L.; Licht, T.; et al. Vascular Endothelial Growth Factor Augments the Tolerance Towards Cerebral Stroke by Enhancing Neurovascular Repair Mechanism. Transl. Stroke Res. 2022, 13, 774–791. [Google Scholar] [CrossRef]
- Dzietko, M.; Derugin, N.; Wendland, M.F.; Vexler, Z.S.; Ferriero, D.M. Delayed VEGF Treatment Enhances Angiogenesis and Recovery After Neonatal Focal Rodent Stroke. Transl. Stroke Res. 2012, 4, 189–200. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, K.S.; Park, I.H.; Kim, S.U. Human Neural Stem Cells Over-Expressing VEGF Provide Neuroprotection, Angiogenesis and Functional Recovery in Mouse Stroke Model. PLoS ONE 2007, 2, e156. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Su, H.; Fan, Y.; Chen, Y.; Zhu, Y.; Liu, W.; Young, W.L.; Yang, G.-Y. Adeno-associated viral-vector-mediated hypoxia-inducible vascular endothelial growth factor gene expression attenuates ischemic brain injury after focal cerebral ischemia in mice. Stroke 2006, 37, 2601–2606. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-P.; Liu, H.-J.; Wang, Z.-L.; Cheng, S.-M.; Cheng, X.; Xu, G.-L.; Liu, X.-F. The dose-effectiveness of intranasal VEGF in treatment of experimental stroke. Neurosci. Lett. 2009, 461, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Manoonkitiwongsa, P.S.; Schultz, R.L.; Whitter, E.F.; Lyden, P.D. Contraindications of VEGF-based therapeutic angiogenesis: Effects on macrophage density and histology of normal and ischemic brains. Vasc. Pharmacol. 2006, 44, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Feigin, V.L.; Brainin, M.; Norrving, B.; Martins, S.; Sacco, R.L.; Hacke, W.; Fisher, M.; Pandian, J.; Lindsay, P. World Stroke Organization (WSO): Global Stroke Fact Sheet 2022. Int. J. Stroke 2022, 17, 18–29. [Google Scholar] [CrossRef]
- Barthels, D.; Das, H. Current advances in ischemic stroke research and therapies. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1866, 165260. [Google Scholar] [CrossRef]
- National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group Tissue Plasminogen Activator for Acute Ischemic Stroke. N. Engl. J. Med. 1995, 333, 1581–1588. [CrossRef]
- Kaya, D.; Gürsoy-Ozdemir, Y.; Yemisci, M.; Tuncer, N.; Aktan, S.; Dalkara, T. VEGF Protects Brain against Focal Ischemia without Increasing Blood–Brain Permeability When Administered Intracerebroventricularly. J. Cereb. Blood Flow Metab. 2005, 25, 1111–1118. [Google Scholar] [CrossRef]
- Li, Z.; Wang, R.; Li, S.; Wei, J.; Zhang, Z.; Li, G.; Dou, W.; Wei, Y.; Feng, M. Intraventricular pre-treatment with rAAV-VEGF induces intracranial hypertension and aggravates ischemic injury at the early stage of transient focal cerebral ischemia in rats. Neurol. Res. 2008, 30, 868–875. [Google Scholar] [CrossRef]
- Hayashi, T.; Abe, K.; Itoyama, Y. Reduction of Ischemic Damage by Application of Vascular Endothelial Growth Factor in Rat Brain after Transient Ischemia. J. Cereb. Blood Flow Metab. 1998, 18, 887–895. [Google Scholar] [CrossRef]
- Liu, Y.; Long, L.; Zhang, F.; Hu, X.; Zhang, J.; Hu, C.; Wang, Y.; Xu, J. Microneedle-mediated vascular endothelial growth factor delivery promotes angiogenesis and functional recovery after stroke. J. Control. Release 2021, 338, 610–622. [Google Scholar] [CrossRef]
- Herrán, E.; Ruiz-Ortega, J.; Aristieta, A.; Igartua, M.; Requejo, C.; Lafuente, J.V.; Ugedo, L.; Pedraz, J.L.; Hernández, R.M. In vivo administration of VEGF- and GDNF-releasing biodegradable polymeric microspheres in a severe lesion model of Parkinson’s disease. Eur. J. Pharm. Biopharm. 2013, 85, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Hernández, R.M.; Herran, E.; Requejo, C.; Ruiz-Ortega, J.; Aristieta, A.; Igartua, M.; Bengoetxea, H.; Ugedo, L.; Pedraz, J.L.; Lafuente, J.V. Increased antiparkinson efficacy of the combined administration of VEGF- and GDNF-loaded nanospheres in a partial lesion model of Parkinson’s disease. Int. J. Nanomed. 2014, 9, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Baker, T.L.; Wilson, C.T.; Brady, R.D.; Mychasiuk, R.; Yamakawa, G.R.; Vo, A.; Wilson, T.; McDonald, S.J.; Shultz, S.R. Treatment with vascular endothelial growth factor-A worsens cognitive recovery in a rat model of mild traumatic brain injury. Front. Mol. Neurosci. 2022, 15, 7350. [Google Scholar] [CrossRef] [PubMed]
- Anttila, V.; Saraste, A.; Knuuti, J.; Hedman, M.; Jaakkola, P.; Laugwitz, K.-L.; Krane, M.; Jeppsson, A.; Sillanmäki, S.; Rosenmeier, J.; et al. Direct intramyocardial injection of VEGF mRNA in patients undergoing coronary artery bypass grafting. Mol. Ther. 2022, 31, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Petrelis, A.M.; Stathopoulou, M.G.; Kafyra, M.; Murray, H.; Masson, C.; Lamont, J.; Fitzgerald, P.; Dedoussis, G.; Yen, F.T.; Visvikis-Siest, S. VEGF-A-related genetic variants protect against Alzheimer’s disease. Aging 2022, 14, 2524–2536. [Google Scholar] [CrossRef] [PubMed]
- Guyot, M.; Pages, G. VEGF splicing and the role of VEGF splice variants: From physiological-pathological conditions to specific pre-mRNA splicing. Methods Mol. Biol. 2015, 1332, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Jiang, A.; Pei, D. Identification and characterization of a new splicing variant of vascular endothelial growth factor: VEGF183. Biochim. Biophys. Acta Gene Struct. Expr. 1998, 1443, 400–406. [Google Scholar] [CrossRef]
- Neufeld, G.; Cohen, T.; Gitay-Goren, H.; Poltorak, Z.; Tessler, S.; Sharon, R.; Gengrinovitch, S.; Levi, B.-Z. Similarities and differences between the vascular endothelial growth factor (VEGF) splice variants. Cancer Metastasis Rev. 1996, 15, 153–158. [Google Scholar] [CrossRef]
- Mineur, P.; Colige, A.C.; Deroanne, C.F.; Dubail, J.; Kesteloot, F.; Habraken, Y.; Noel, A.; Vöö, S.; Waltenberger, J.; Lapière, C.M.; et al. Newly identified biologically active and proteolysis-resistant VEGF-A isoform VEGF111 is induced by genotoxic agents. J. Cell Biol. 2007, 179, 1261–1273. [Google Scholar] [CrossRef]
- Delcombel, R.; Janssen, L.; Vassy, R.; Gammons, M.; Haddad, O.; Richard, B.; Colige, A. New prospects in the roles of the C-terminal domains of VEGF-A and their cooperation for ligand binding, cellular signaling and vessels formation. Angiogenesis 2013, 16, 353–371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; van Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Investig. 2000, 106, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Rosengart, T.K.; Bishawi, M.M.; Halbreiner, M.S.; Fakhoury, M.; Finnin, E.; Hollmann, C.; Shroyer, A.L.; Crystal, R.G. Long-Term Follow-Up Assessment of a Phase 1 Trial of Angiogenic Gene Therapy Using Direct Intramyocardial Administration of an Adenoviral Vector Expressing the VEGF121 cDNA for the Treatment of Diffuse Coronary Artery Disease. Hum. Gene Ther. 2013, 24, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.J.; Hilton, J.D.; Arnold, J.M.; Gregoire, J.; Rivard, A.; Archer, S.L.; Charbonneau, F.; Cohen, E.; Curtis, M.; Buller, C.E.; et al. Angiogenic gene therapy in patients with nonrevascularizable ischemic heart disease: A phase 2 randomized, controlled trial of AdVEGF121 (AdVEGF121) versus maximum medical treatment. Gene Ther. 2006, 13, 1503–1511. [Google Scholar] [CrossRef]
- Kastrup, J.; Jørgensen, E.; Fuchs, S.; Nikol, S.; Bøtker, H.; Gyöngyösi, M.; Glogar, D.; Kornowski, R. A randomised, double-blind, placebo-controlled, multicentre study of the safety and efficacy of BIOBYPASS (AdGVVEGF121.10NH) gene therapy in patients with refractory advanced coronary artery disease: The NOVA trial. Eurointervention 2011, 6, 813–818. [Google Scholar] [CrossRef]
- Woolard, J.; Bevan, H.S.; Harper, S.J.; Bates, D. Molecular Diversity of VEGF-A as a Regulator of Its Biological Activity. Microcirculation 2009, 16, 572–592. [Google Scholar] [CrossRef]
- Ourradi, K.; Blythe, T.; Jarrett, C.; Barratt, S.L.; Welsh, G.I.; Millar, A.B. VEGF isoforms have differential effects on permeability of human pulmonary microvascular endothelial cells. Respir. Res. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Giacca, M.; Zacchigna, S. VEGF gene therapy: Therapeutic angiogenesis in the clinic and beyond. Gene Ther. 2012, 19, 622–629. [Google Scholar] [CrossRef]
- Trout, A.L.; Kahle, M.P.; Roberts, J.M.; Marcelo, A.; de Hoog, L.; Boychuk, J.A.; Grupke, S.L.; Berretta, A.; Gowing, E.K.; Boychuk, C.R.; et al. Perlecan Domain-V Enhances Neurogenic Brain Repair After Stroke in Mice. Transl. Stroke Res. 2020, 12, 72–86. [Google Scholar] [CrossRef]
- Lee, B.; Clarke, D.; Alahmad, A.; Kahle, M.; Parham, C.; Auckland, L.; Shaw, C.; Fidanboylu, M.; Orr, A.; Ogunshola, O.; et al. Perlecan domain V is neuroprotective and proangiogenic following ischemic stroke in rodents. J. Clin. Investig. 2011, 121, 3005–3023. [Google Scholar] [CrossRef]
- Kaji, T.; Yamamoto, C.; Oh-I, M.; Fujiwara, Y.; Yamazaki, Y.; Morita, T.; Plaas, A.H.; Wight, T.N. The vascular endothelial growth factor VEGF165 induces perlecan synthesis via VEGF receptor-2 in cultured human brain microvascular endothelial cells. Biochim. Biophys. Acta Gen. Subj. 2006, 1760, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Castillo, G.M.; Cummings, J.A.; Yang, W.; Judge, M.E.; Sheardown, M.J.; Rimvall, K.; Snow, A.D. Sulfate Content and Specific Glycosaminoglycan Backbone of Perlecan Are Critical for Perlecan’s Enhancement of Islet Amyloid Polypeptide (Amylin) Fibril Formation. Diabetes 1998, 47, 612. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.N.; Alahmad, A.; Lee, B.; Parham, C.; Auckland, L.; Fertala, A.; Kahle, M.; Shaw, C.S.; Roberts, J.; Bix, G.J. Perlecan Domain V Induces VEGf Secretion in Brain Endothelial Cells through Integrin α5β1 and ERK-Dependent Signaling Pathways. PLoS ONE 2012, 7, e45257. [Google Scholar] [CrossRef] [PubMed]
- Barć, P.; Antkiewicz, M.; Śliwa, B.; Frączkowska, K.; Guziński, M.; Dawiskiba, T.; Małodobra-Mazur, M.; Witkiewicz, W.; Kupczyńska, D.; Strzelec, B.; et al. Double VEGF/HGF Gene Therapy in Critical Limb Ischemia Complicated by Diabetes Mellitus. J. Cardiovasc. Transl. Res. 2020, 14, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Deev, R.V.; Bozo, I.Y.; Mzhavanadze, N.D.; Voronov, D.A.; Gavrilenko, A.V.; Chervyakov, Y.V.; Staroverov, I.N.; Kalinin, R.E.; Shvalb, P.G.; Isaev, A.A. pCMV-vegf165 Intramuscular Gene Transfer is an Effective Method of Treatment for Patients with Chronic Lower Limb Ischemia. J. Cardiovasc. Pharmacol. Ther. 2015, 20, 473–482. [Google Scholar] [CrossRef]
- Symes, J.F.; Losordo, D.W.; Vale, P.R.; Lathi, K.G.; Esakof, D.D.; Mayskiy, M.; Isner, J.M. Gene therapy with vascular endothelial growth factor for inoperable coronary artery disease. Ann. Thorac. Surg. 1999, 68, 830–836. [Google Scholar] [CrossRef]
- Sarkar, N.; Ruck, A.; Kallner, G.; Y-Hassan, S.; Blomberg, P.; Islam, K.B.; Van Der Linden, J.; Lindblom, D.; Nygren, A.T.; Lind, B.; et al. Effects of intramyocardial injection of phVEGF-A165 as sole therapy in patients with refractory coronary artery disease—12-month follow-up: Angiogenic gene therapy. J. Intern. Med. 2001, 250, 373–381. [Google Scholar] [CrossRef]
- Hedman, M.; Muona, K.; Hedman, A.; Kivelä, A.; Syvänne, M.; Eränen, J.; Rantala, A.; Stjernvall, J.; Nieminen, M.S.; Hartikainen, J.; et al. Eight-year safety follow-up of coronary artery disease patients after local intracoronary VEGF gene transfer. Gene Ther. 2009, 16, 629–634. [Google Scholar] [CrossRef]
- Giusti, I.I.; Rodrigues, C.G.; Salles, F.B.; Sant’Anna, R.T.; Eibel, B.; Han, S.W.; Ludwig, E.; Grossman, G.; Prates, P.R.L.; Sant’Anna, J.R.M.; et al. High Doses of Vascular Endothelial Growth Factor 165 Safely, but Transiently, Improve Myocardial Perfusion in No-Option Ischemic Disease. Hum. Gene Ther. Methods 2013, 24, 298–306. [Google Scholar] [CrossRef]
- Lord, M.S.; Ellis, A.L.; Farrugia, B.L.; Whitelock, J.M.; Grenett, H.; Li, C.; O’Grady, R.L.; DeCarlo, A.A. Perlecan and vascular endothelial growth factor-encoding DNA-loaded chitosan scaffolds promote angiogenesis and wound healing. J. Control. Release 2017, 250, 48–61. [Google Scholar] [CrossRef]
- Hayes, A.J.; Farrugia, B.L.; Biose, I.J.; Bix, G.J.; Melrose, J. Perlecan, a Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan with Roles in Tissue Development Has Relevance to Connective Tissue Repair and Regeneration. Front. Cell Dev. Biol. 2022, 10, 6261. [Google Scholar] [CrossRef] [PubMed]
- Gyöngyösi, M.; Khorsand, A.; Zamini, S.; Sperker, W.; Strehblow, C.; Kastrup, J.; Jorgensen, E.; Hesse, B.; Tägil, K.; Bøtker, H.E.; et al. NOGA-Guided Analysis of Regional Myocardial Perfusion Abnormalities Treated with Intramyocardial Injections of Plasmid Encoding Vascular Endothelial Growth Factor A-165 in Patients with Chronic Myocardial Ischemia. Circulation 2005, 112, I157–I165. [Google Scholar] [CrossRef] [PubMed]
- Favaloro, L.; Diez, M.; Mendiz, O.; Janavel, G.V.; Valdivieso, L.; Ratto, R.; Garelli, G.; Salmo, F.; Criscuolo, M.; Bercovich, A.; et al. A Phase I, Open-Label, Two-Year Follow-up Trial. Catheter. Cardiovasc. Interv. 2012, 82, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Kukuła, K.; Urbanowicz, A.; Kłopotowski, M.; Dąbrowski, M.; Pręgowski, J.; Kądziela, J.; Chmielak, Z.; Witkowski, A.; Rużyłło, W. Long-term follow-up and safety assessment of angiogenic gene therapy trial VIF-CAD: Transcatheter intramyocardial administration of a bicistronic plasmid expressing VEGF-A165/bFGF cDNA for the treatment of refractory coronary artery disease. Am. Heart J. 2019, 215, 78–82. [Google Scholar] [CrossRef]
- Ripa, R.S.; Wang, Y.; Jørgensen, E.; Johnsen, H.E.; Hesse, B.; Kastrup, J. Intramyocardial injection of vascular endothelial growth factor-A165 plasmid followed by granulocyte-colony stimulating factor to induce angiogenesis in patients with severe chronic ischaemic heart disease. Eur. Heart J. 2006, 27, 1785–1792. [Google Scholar] [CrossRef]
- Rhim, T.; Lee, D.Y.; Lee, M. Drug Delivery Systems for the Treatment of Ischemic Stroke. Pharm. Res. 2013, 30, 2429–2444. [Google Scholar] [CrossRef]
- Eswarappa, S.M.; Potdar, A.A.; Koch, W.J.; Fan, Y.; Vasu, K.; Lindner, D.; Willard, B.; Graham, L.M.; DiCorleto, P.E.; Fox, P.L. Programmed Translational Readthrough Generates Antiangiogenic VEGF-Ax. Cell 2014, 157, 1605–1618. [Google Scholar] [CrossRef]
- Xin, H.; Zhong, C.; Nudleman, E.; Ferrara, N. Evidence for Pro-angiogenic Functions of VEGF-Ax. Cell 2016, 167, 275–284.e6. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Drug. | Animal Model | Disease Model | Dosage | Timing | Route | Result | Reference |
|---|---|---|---|---|---|---|---|
| VEGFA164 | mice | MCAO | 8 ng | 1 h post | intracerebroventricular | Reduced infarct, increase BBB integrity | [38] |
| VEGFA164 | mice | MCAO | 8 ng | 3 h post | intracerebroventricular | Reduced infarct, reduced apoptosis, increase BBB integrity | [38] |
| VEGFA164 | Mice | MCAO | 15 µg/kg | 1 h post | intravenous | Increase BBB permeability | [38] |
| VEGFA | Mice | CHI | 0.84 µg | Same time | Intravenous | Decrease lesion volume, improved neurological severity score | [26] |
| VEGFA | Rats | spinal cord injury | 500 ng | same time | Intraspinal injection | Increased neuronal stem cell proliferation | [20] |
| VEGFA165 | Rats | tMCAO | --- | 8 weeks before | AAV-plasmid injection | Intracranial hypertension, secondary ischemic injury | [39] |
| VEGFA | Mice | MCAO | --- | 28 days before | Conditional activation of transgene | infarct reduction | [29] |
| VEGFA165 | Rats | tMCAO | 9 ng | Same time | Gelfoam above cerebral cortex | Reduced infarct size | [40] |
| VEGFA | Rats | photothrombotic | 1 × 1010 vg | 3 days post | microneedle in cerebral cortex | No inflammation, increase angiogenesis, increase neu | [41] |
| VEGFA165 | Rats | 6-OHDA | 2.5 µg | 4 weeks post | Microsphere implantation on cerebral cortex | No change | [42] |
| VEGFA165 and GDNF | Rats | 6-OHDA | 2.5 µg: 2.5 µg | 4 weeks post | Microsphere implantation on cerebral cortex | Improvement in neuronal density trending | [42] |
| VEGFA and BDNF | rats | 6-OHDA | 1.25 µg: 1.25 µg | 3 weeks post | Nanosphere on Right striatum | Increased neuronal density | [43] |
| VEGFA | Rats | 6-OHDA | 2.5 µg | 3 weeks post | Nanosphere on right striatum | No change | [43] |
| VEGFA | Rats | mTBI | 10 µg/mL | Same time | Intracerebroventricular cannulation | Worse cognitive deficits | [44] |
| VEGFA Forms | VEGFA | 111 | 121 | 145 | 148 | 162 | 165 | 183 | 189 | 206 |
|---|---|---|---|---|---|---|---|---|---|---|
| Pro-angiogenic | Yes [9,47] | Yes [9,47] | Yes | Yes [9,47] | Yes [9,47] | Yes [9,47] | Yes [9,47] | Yes [9,47] | Yes [9,47] | Yes [9,47] |
| Binds to VEGFR1 | Yes [31] | No [36] | Yes [9] | No [47] | n/a | n/a | Yes [3] | n/a | n/a | n/a |
| Binds to VEGFR2 | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] | Yes [47] |
| Binds to Nrp1 | Yes [13] | n/a | n/a | n/a | n/a | n/a | Yes [13] | n/a | Yes [16] | n/a |
| Binds to Nrp2 | Yes [13] | n/a | Yes [15] | Yes [13] | n/a | n/a | Yes [13] | n/a | n/a | n/a |
| Binds to ECM | Yes [3] | No [9] | No [3,9] | Yes [9,47] | n/a | Yes [47] | Yes [9,47] | Yes [47] | Yes [9,47] | Yes [9,47] |
| Freely secreted | Yes [9] | Yes [9] | Yes [3] | No [48] | n/a | n/a | Yes [3] | n/a | No [3] | No [3] |
| Pro-neovascularization | Yes [9] | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Vascular permeability | Increases [9,47] | n/a | Increases [47,49] | n/a | n/a | n/a | Increases [47,49] | n/a | n/a | n/a |
| neuroprotective | Yes [47] | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a | n/a |
| Pro-neurogenesis | Yes [47] | n/a | n/a | n/a | n/a | n/a | Yes [47] | n/a | n/a | n/a |
| Immune cell recruitment | Yes [47] | n/a | Yes [47] | n/a | n/a | n/a | Yes [47] | n/a | n/a | n/a |
| Drug | Disease Model | Dosage | Route | Result | Reference |
|---|---|---|---|---|---|
| VEGFA165 and HFG | CLI | 2.5 mL | Intramuscular injection of a plasmid | VEGF165 increase, decrease pain, increase vascularization, | [64] |
| VEGF165 | CAD | 3.8 mg | Catheter insertion of a plasmid | Decreased angina, myocardial perfusion decrease, coronary-related complications diagnosed in follow-up study | [73] |
| VEGFA | Reduced LVEF | 3 mg | Naked mRNA epicardial injection | Safe, no effect | [45] |
| VEGFA121 | CAD | 1 mL | Adenoviral epicardial injection | safe | [53] |
| VEGFA165 | Ischemic cardiomyopathy | 2 mg | Plasmid intramyocardial injection | Safe, increased cardiac perfusion and oxygen consumption | [69] |
| VEGF165 | CLI | 2.4 mg | Plasmid intramuscular injections | Pain reduction, increased blood flow | [65] |
| VEGF165 and FGF | CAD | 0.5 mg | intramyocardial | Safe, no effect | [74] |
| VEGF165 | Coronary angina | 125 µg | Plasmid intramyocardial injection | Reduced angina and ischemic defects | [66] |
| VEGF165 | Coronary angina | 250 µg | Plasmid intramyocardial injection | Reduced angina and ischemic defects | [66] |
| VEGFA165 and G-CSF | IHD | 60 µg | Plasmid intramyocardial injection | No effect | [75] |
| VEGFA121 | CAD | 4 × 1010 particle units | Adenoviral intramyocardial injection | Increased neovascularization, coronary-related complications in follow up study | [54] |
| VEGFA121 | CAD | --- | Intramyocardial injections | Safe, no effect | [55] |
| VEGFA165 | CAD | 0.25 mg | Plasmid intramyocardial injection | Reduction in angina, safe | [67] |
| VEGFA165 | IHD | 0.5 mg | Plasmid intramyocardial injection | Trend towards improvement | [72] |
| VEGFA165 and HGF | CLI | --- | Plasmid intramuscular injection | Safe, no effect | [64] |
| VEGFA165 | Coronary angina | Intravenous injection | No effect | [7] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, A.L.; Bix, G.J. VEGFA Isoforms as Pro-Angiogenic Therapeutics for Cerebrovascular Diseases. Biomolecules 2023, 13, 702. https://doi.org/10.3390/biom13040702
White AL, Bix GJ. VEGFA Isoforms as Pro-Angiogenic Therapeutics for Cerebrovascular Diseases. Biomolecules. 2023; 13(4):702. https://doi.org/10.3390/biom13040702
Chicago/Turabian StyleWhite, Amanda Louise, and Gregory Jaye Bix. 2023. "VEGFA Isoforms as Pro-Angiogenic Therapeutics for Cerebrovascular Diseases" Biomolecules 13, no. 4: 702. https://doi.org/10.3390/biom13040702