
Multiomics Analysis Reveals Novel Genetic Determinants for Lens Differentiation, Structure, and Transparency

Abstract
:1. Introduction: The Eye Lens as a Model System for Unbiased Multiomics Analysis of Gene Regulatory Mechanisms
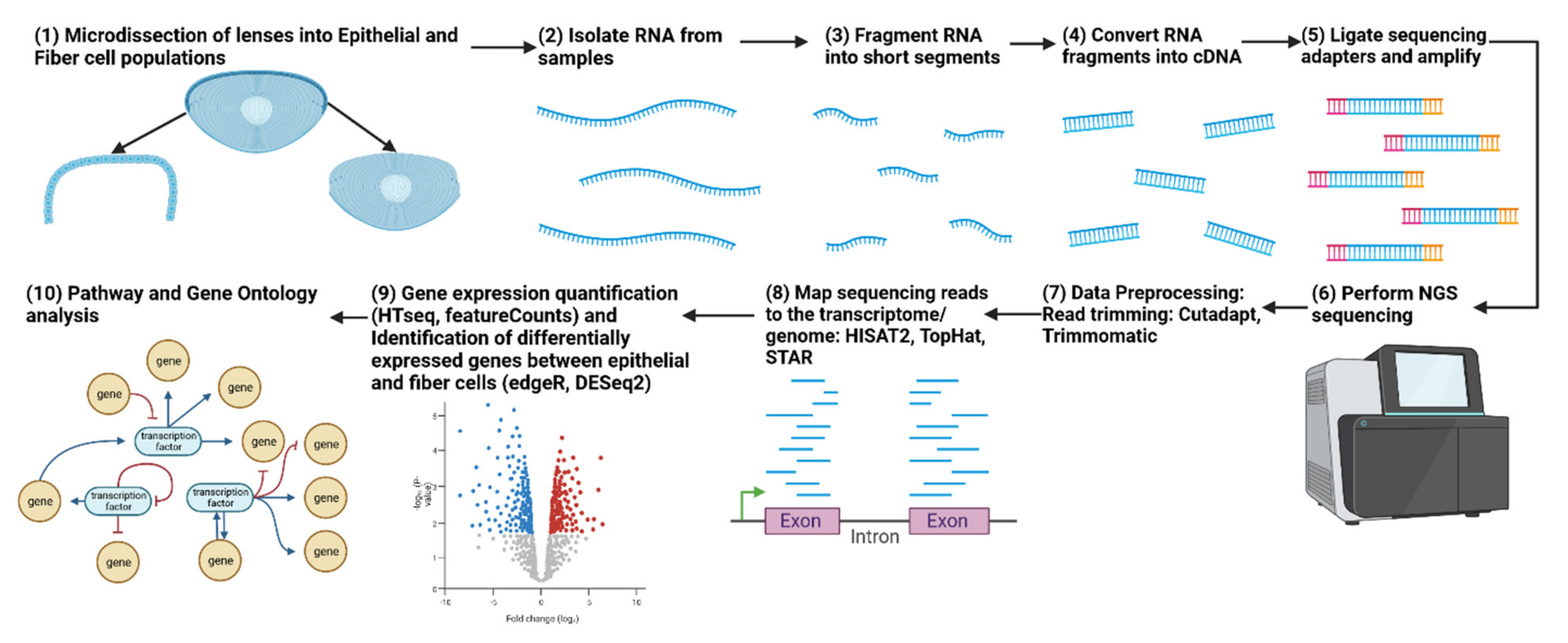
2. Bulk Transcriptome Analysis of the Lens via RNA-seq
2.1. Analysis of Lens Epithelium and Fiber Cell Compartments in Chicken and Mouse Lenses by RNA-seq
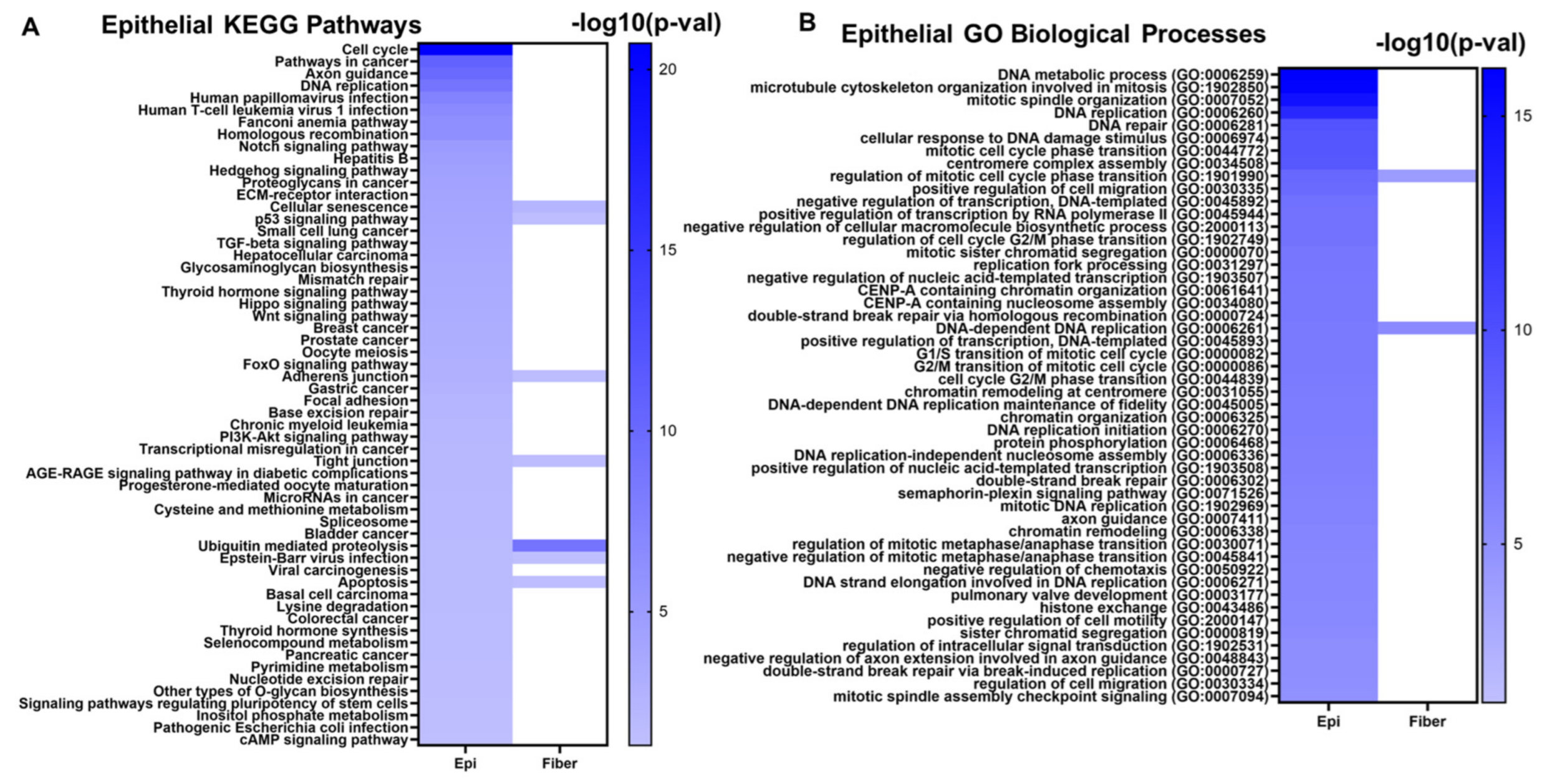
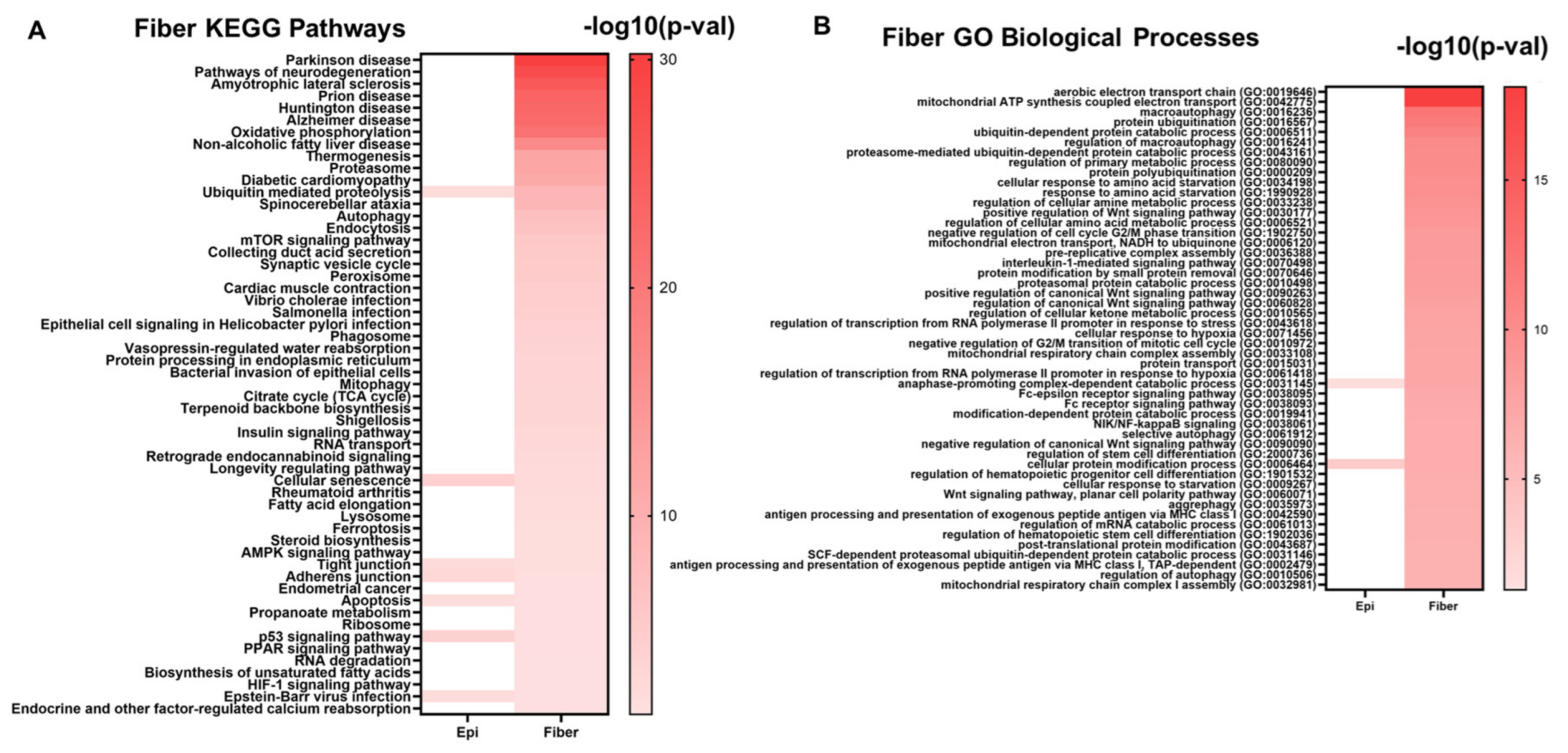
2.2. Signaling Pathways Inferred from RNA-seq Data
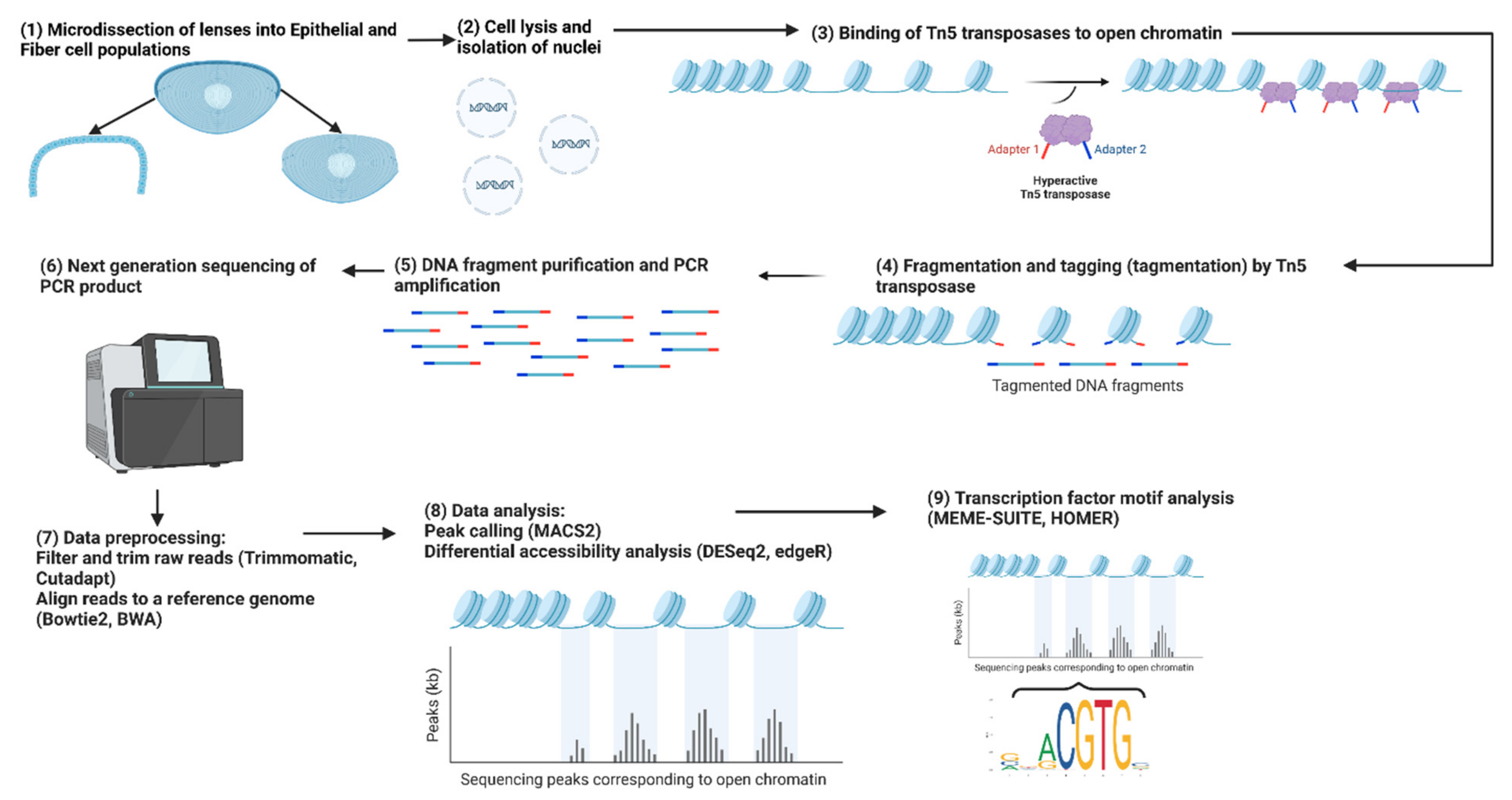
3. Analysis of Chromatin Landscape in Lens Tissues with ATAC-seq
3.1. Changes in Chromatin Accessibility in Different Chicken and Mouse Lens Compartments
3.2. Chromatin Landscape of Important Genes Encoding Lens Regulatory and Structural Proteins and Identification of Transcription Factor Binding Sequences
4. Multiomics Integration Analysis: Goals and Challenges
5. Multiomics Integration of Lens Differentiation RNA-seq and ATAC-seq: A Cross-Species Comparison
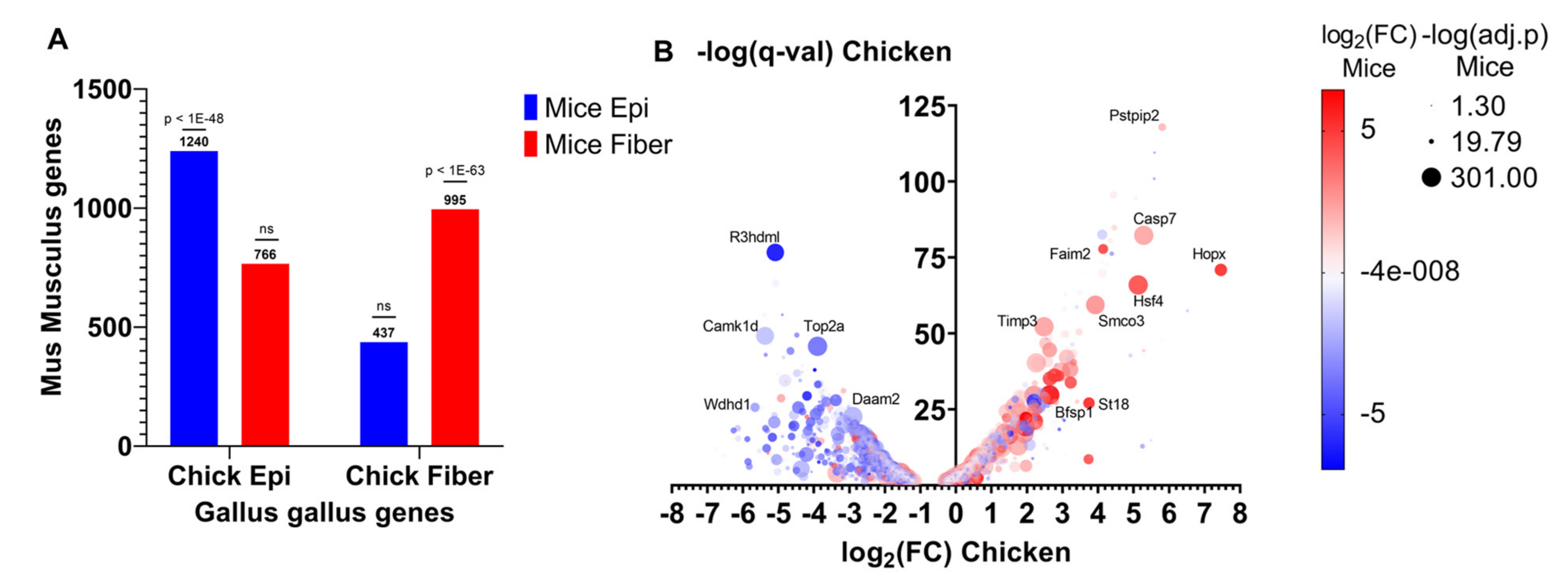
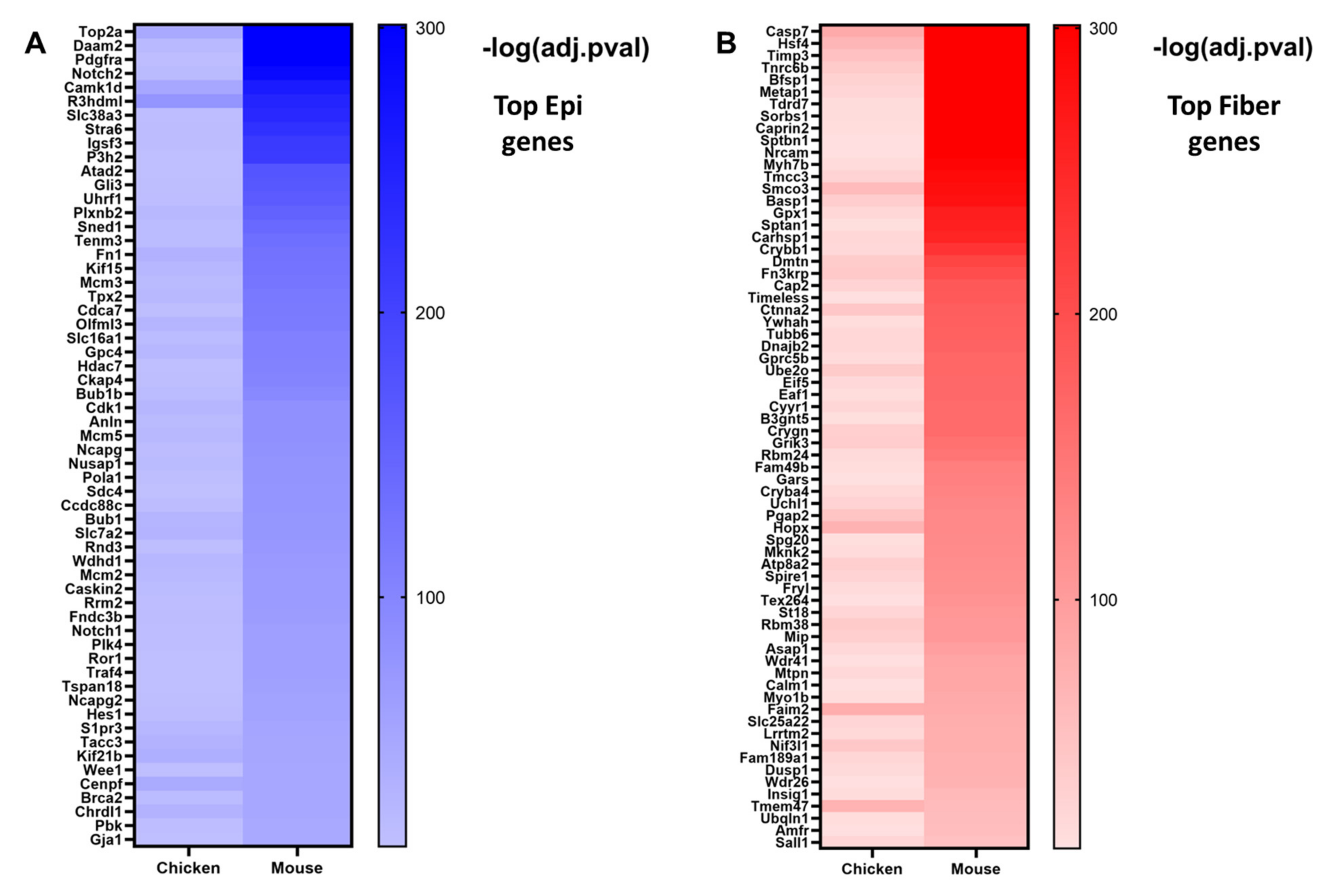
5.1. Evolutionarily Conserved Gene Expression Patterns Associated with Lens Differentiation
5.2. Evolutionarily Conserved Cell Signaling Pathways and Gene Ontologies Associated with Lens Differentiation
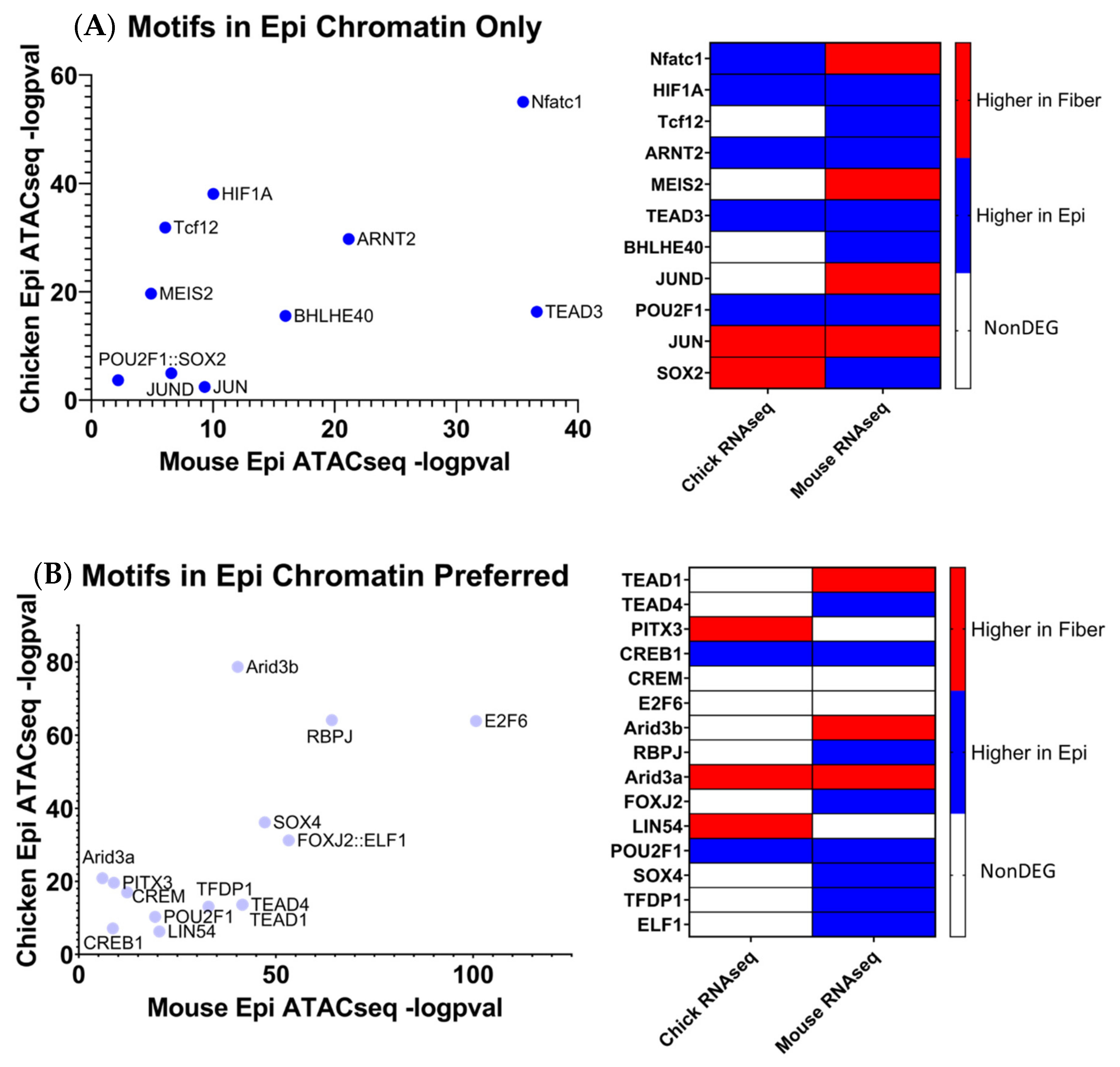
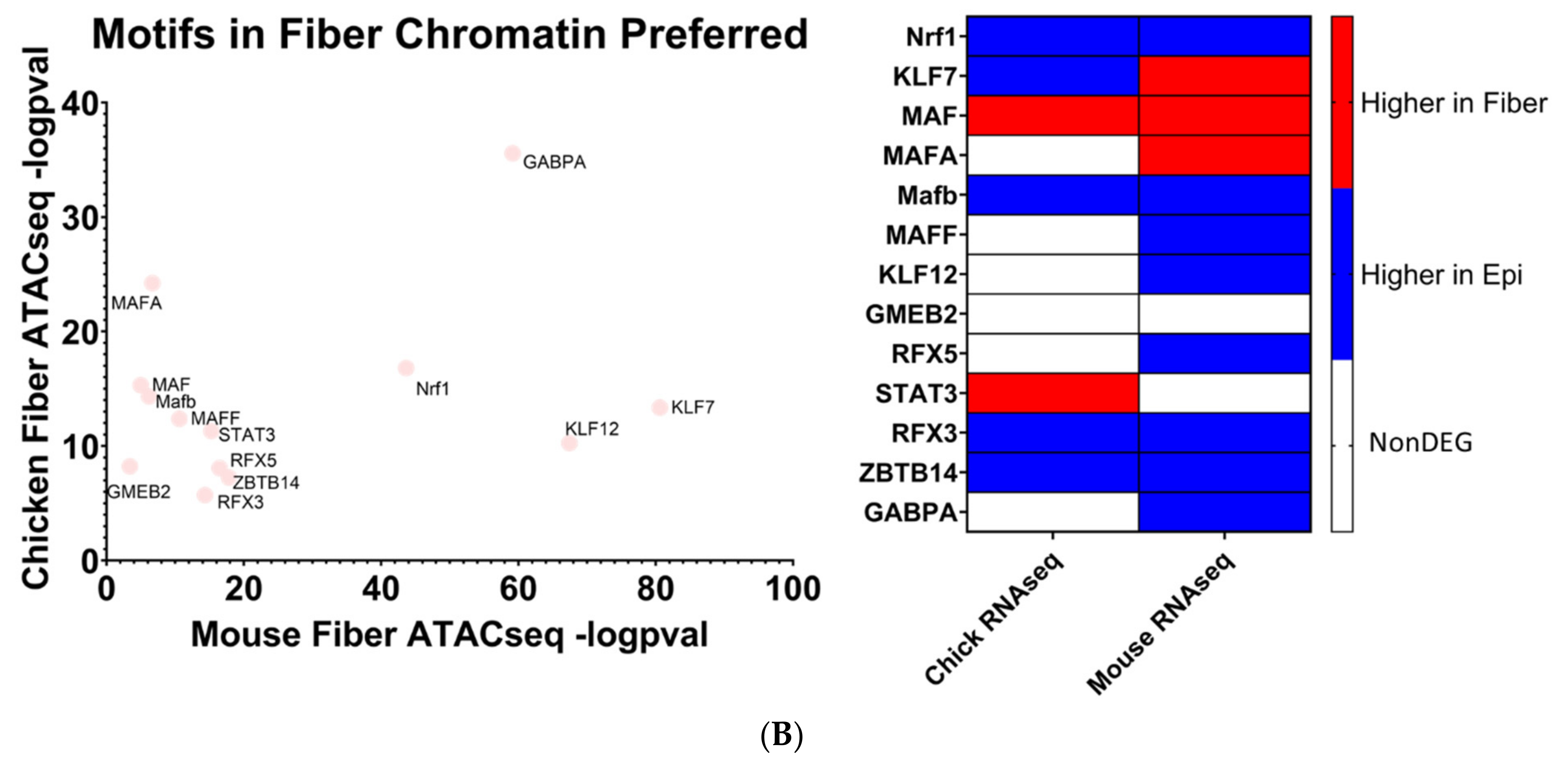
5.3. Identification of Evolutionarily Conserved Transcription Factor Binding Motifs via Multiomics Integrated Analysis
6. Whole-Genome Bisulfite Sequencing (WGBS) and Other Omics Opportunities to Study Conserved Regulatory Mechanisms of Gene Expression
7. Integrating Multiomics Analysis Results with Lens Gene Databases
8. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| ATAC-seq | Assay for transposase accessible chromatin with sequencing |
| WGBS | Whole-genome bisulfite sequencing |
| CHIP-seq | Chromatin immunoprecipitation with sequencing |
| CUT&RUN | Cleavage under targets and release using nuclease |
| EST | Expressed sequence tags |
| EC | Anterior central epithelial cells |
| EQ | Equatorial epithelial cells |
| FP | Cortical fiber cells |
| FC | Central fiber cells |
| FAIRE-seq | Formaldehyde-assisted isolation of regulatory elements |
| GO | Gene Ontology |
| Epi | Lens epithelial cells |
| Fiber | Lens fiber cells |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| AME | Analysis of motif enrichment |
References
- Cvekl, A.; Zhang, X. Signaling and Gene Regulatory Networks in Mammalian Lens Development. Trends Genet. 2017, 33, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S.; Shi, Y.; Vrensen, G.F.J.M. Biological Glass: Structural Determinants of Eye Lens Transparency. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1250–1264. [Google Scholar] [CrossRef] [PubMed]
- Menko, S.A. Lens Epithelial Cell Differentiation. Exp. Eye Res. 2002, 75, 485–490. [Google Scholar] [CrossRef]
- Wride, M.A. Lens Fibre Cell Differentiation and Organelle Loss: Many Paths Lead to Clarity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1219–1233. [Google Scholar] [CrossRef]
- Zhang, P.; Wong, C.; DePinho, R.A.; Harper, J.W.; Elledge, S.J. Cooperation between the Cdk Inhibitors P27(KIP1) and P57(KRP2) in the Control of Tissue Growth and Development. Genes Dev. 1998, 12, 3162–3167. [Google Scholar] [CrossRef]
- Kupfer, C. The National Eye Institute’s Low Vision Education Program: Improving Quality of Life. Editorial. Ophthalmology 2000, 107, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Makrides, N.; Wang, Q.; Tao, C.; Schwartz, S.; Zhang, X. Jack of All Trades, Master of Each: The Diversity of Fibroblast Growth Factor Signalling in Eye Development. Open Biol. 2022, 12, 210265. [Google Scholar] [CrossRef]
- Lovicu, F.J.; McAvoy, J.W.; de Iongh, R.U. Understanding the role of growth factors in embryonic development: Insights from the lens. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1204–1218. [Google Scholar] [CrossRef]
- Lovicu, F.J.; McAvoy, J.W. Growth Factor Regulation of Lens Development. Dev. Biol. 2005, 280, 1–14. [Google Scholar] [CrossRef]
- Smith, A.N.; Miller, L.A.D.; Song, N.; Taketo, M.M.; Lang, R.A. The duality of β-catenin function: A requirement in lens morphogenesis and signaling suppression of lens fate in periocular ectoderm. Dev. Biol. 2005, 285, 477–489. [Google Scholar] [CrossRef]
- Kreslova, J.; Machon, O.; Rusickova, J.; Lachova, J.; Wawrousek, E.F.; Kemler, R.; Krauss, S.; Piatigorsky, J.; Kozmik, Z. Abnormal lens morphogenesis and ectopic lens formation in the absence of β-catenin function. Genesis 2007, 45, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Machon, O.; Kreslova, J.; Ruzickova, J.; Vacik, T.; Klimova, L.; Fujimura, N.; Lachova, J.; Kozmik, Z. Lens Morphogenesis Is Dependent on Pax6-Mediated Inhibition of the Canonical Wnt/Beta-Catenin Signaling in the Lens Surface Ectoderm. Genesis 2010, 48, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Antosova, B.; Smolikova, J.; Borkovcova, R.; Strnad, H.; Lachova, J.; Machon, O.; Kozmik, Z. Ectopic Activation of Wnt/β-Catenin Signaling in Lens Fiber Cells Results in Cataract Formation and Aberrant Fiber Cell Differentiation. PLoS ONE 2013, 8, e78279. [Google Scholar] [CrossRef] [PubMed]
- Dawes, L.J.; Sugiyama, Y.; Tanedo, A.S.; Lovicu, F.J.; McAvoy, J.W. Wnt-Frizzled Signaling Is Part of an FGF-Induced Cascade That Promotes Lens Fiber Differentiation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1582–1590. [Google Scholar] [CrossRef]
- Chen, Y.; Stump, R.J.W.; Lovicu, F.J.; Shimono, A.; McAvoy, J.W. Wnt Signaling Is Required for Organization of the Lens Fiber Cell Cytoskeleton and Development of Lens Three-Dimensional Architecture. Dev. Biol. 2008, 324, 161–176. [Google Scholar] [CrossRef]
- Rowan, S.; Conley, K.W.; Le, T.T.; Donner, A.L.; Maas, R.L.; Brown, N.L. Notch Signaling Regulates Growth and Differentiation in the Mammalian Lens. Dev. Biol. 2008, 321, 111–122. [Google Scholar] [CrossRef]
- Saravanamuthu, S.S.; Le, T.T.; Gao, C.Y.; Cojocaru, R.I.; Pandiyan, P.; Liu, C.; Zhang, J.; Zelenka, P.S.; Brown, N.L. Conditional Ablation of the Notch2 Receptor in the Ocular Lens. Dev. Biol. 2012, 362, 219–229. [Google Scholar] [CrossRef]
- Le, T.T.; Conley, K.W.; Mead, T.J.; Rowan, S.; Yutzey, K.E.; Brown, N.L. Requirements for Jag1-Rbpj mediated Notch signaling during early mouse lens development. Dev. Dyn. 2012, 241, 493–504. Available online: https://pubmed.ncbi.nlm.nih.gov/22275127/ (accessed on 7 November 2021).
- Azimi, M.; Brown, N.L. Jagged1 protein processing in the developing mammalian lens. Biol. Open 2019, 8, bio041095. Available online: http://www.ncbi.nlm.nih.gov/pubmed/30890522 (accessed on 14 February 2021).
- Saravanamuthu, S.S.; Gao, C.Y.; Zelenka, P.S. Notch Signaling Is Required for Lateral Induction of Jagged1 during FGF-Induced Lens Fiber Differentiation. Dev. Biol. 2009, 332, 166–176. [Google Scholar] [CrossRef]
- Le, T.T.; Conley, K.W.; Brown, N.L. Jagged 1 Is Necessary for Normal Mouse Lens Formation. Dev. Biol. 2009, 328, 118–126. [Google Scholar] [CrossRef]
- Furuta, Y.; Hogan, B.L.M. BMP4 Is Essential for Lens Induction in the Mouse Embryo. Genes Dev. 1998, 12, 3764–3775. [Google Scholar] [CrossRef]
- Wawersik, S.; Purcell, P.; Rauchman, M.; Dudley, A.T.; Robertson, E.J.; Maas, R. BMP7 Acts in Murine Lens Placode Development. Dev. Biol. 1999, 207, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Pandit, T.; Jidigam, V.K.; Gunhaga, L. BMP-Induced L-Maf Regulates Subsequent BMP-Independent Differentiation of Primary Lens Fibre Cells. Dev. Dyn. 2011, 240, 1917–1928. [Google Scholar] [CrossRef]
- Jarrin, M.; Pandit, T.; Gunhaga, L. A Balance of FGF and BMP Signals Regulates Cell Cycle Exit and Equarin Expression in Lens Cells. Mol. Biol. Cell 2012, 23, 3266–3274. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, Y.; Filas, B.; Gunhaga, L.; Beebe, D.C. Negative and Positive Auto-Regulation of BMP Expression in Early Eye Development. Dev. Biol. 2015, 407, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Belecky-Adams, T.L.; Adler, R.; Beebe, D.C. Bone Morphogenetic Protein Signaling and the Initiation of Lens Fiber Cell Differentiation. Development 2002, 129, 3795–3802. [Google Scholar] [CrossRef]
- Faber, S.C.; Robinson, M.L.; Makarenkova, H.P.; Lang, R.A. Bmp Signaling Is Required for Development of Primary Lens Fiber Cells. Development 2002, 129, 3727–3737. [Google Scholar] [CrossRef] [PubMed]
- Boswell, B.A.; Overbeek, P.A.; Musil, L.S. Essential Role of BMPs in FGF-Induced Secondary Lens Fiber Differentiation. Dev. Biol. 2008, 324, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Huang, J.; Dattilo, L.K.; Kaartinen, V.; Mishina, Y.; Deng, C.X.; Umans, L.; Zwijsen, A.; Roberts, A.B.; Beebe, D.C. The Type I BMP Receptors, Bmpr1a and Acvr1, Activate Multiple Signaling Pathways to Regulate Lens Formation. Dev. Biol. 2009, 335, 305–316. [Google Scholar] [CrossRef]
- Chamberlain, C.G.; Mcavoy, J.W. Induction of Lens Fibre Differentiation by Acidic and Basic Fibroblast Growth Factor (FGF). Growth Factors 1989, 1, 125–134. [Google Scholar] [CrossRef]
- McAvoy, J.W.; Chamberlain, C.G. Fibroblast Growth Factor (FGF) Induces Different Responses in Lens Epithelial Cells Depending on Its Concentration. Development 1989, 107, 221–228. [Google Scholar] [CrossRef]
- Schulz, M.W.; Chamberlain, C.G.; De Longh, R.U.; McAvoy, J.W. Acidic and Basic FGF in Ocular Media and Lens: Implications for Lens Polarity and Growth Patterns. Development 1993, 118, 117–126. [Google Scholar] [CrossRef]
- Lovicu, F.J.; McAvoy, J.W. FGF-Induced Lens Cell Proliferation and Differentiation Is Dependent on MAPK (ERK1/2) Signalling. Development 2001, 128, 5075–5084. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.L.; MacMillan-Crow, L.A.; Thompson, J.A.; Overbeek, P.A. Expression of a Truncated FGF Receptor Results in Defective Lens Development in Transgenic Mice. Development 1995, 121, 3959–3967. [Google Scholar] [CrossRef]
- Robinson, M.L.; Overbeek, P.A.; Verran, D.J.; Grizzle, W.E.; Stockard, C.R.; Friesel, R.; Maciag, T.; Thompson, J.A. Extracellular FGF-1 Acts as a Lens Differentiation Factor in Transgenic Mice. Development 1995, 121, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mao, Y.; Bouaziz, M.; Yu, H.; Qu, X.; Wang, F.; Feng, G.S.; Shawber, C.; Zhang, X. Lens Differentiation Is Controlled by the Balance between PDGF and FGF Signaling. PLoS Biol. 2019, 17, e3000133. [Google Scholar] [CrossRef]
- Collins, T.N.; Mao, Y.; Li, H.; Bouaziz, M.; Hong, A.; Feng, G.S.; Wang, F.; Quilliam, L.A.; Chen, L.; Park, T.; et al. Crk Proteins Transduce FGF Signaling to Promote Lens Fiber Cell Elongation. Elife 2018, 7, e32586. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; McGreal, R.; Harris, R.; Gao, C.Y.; Liu, W.; Reneker, L.W.; Musil, L.S.; Cvekl, A. Regulation of C-Maf and AA-Crystallin in Ocular Lens by Fibroblast Growth Factor Signaling. J. Biol. Chem. 2016, 291, 3947–3958. [Google Scholar] [CrossRef]
- Thein, T.; de Melo, J.; Zibetti, C.; Clark, B.S.; Juarez, F.; Blackshaw, S. Control of Lens Development by Lhx2-Regulated Neuroretinal FGFs. Development 2016, 143, 3994–4002. [Google Scholar] [CrossRef]
- Qu, X.; Hertzler, K.; Pan, Y.; Grobe, K.; Robinson, M.L.; Zhang, X. Genetic Epistasis between Heparan Sulfate and FGF-Ras Signaling Controls Lens Development. Dev. Biol. 2011, 355, 12–20. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, T.; Madakashira, B.P.; Thiels, C.A.; Bechtle, C.A.; Garcia, C.M.; Zhang, H.; Yu, K.; Ornitz, D.M.; Beebe, D.C.; et al. Fibroblast Growth Factor Receptor Signaling Is Essential for Lens Fiber Cell Differentiation. Dev. Biol. 2008, 318, 276–288. [Google Scholar] [CrossRef]
- Porazinski, S.; Wang, H.; Asaoka, Y.; Behrndt, M.; Miyamoto, T.; Morita, H.; Hata, S.; Sasaki, T.; Krens, S.F.G.; Osada, Y.; et al. YAP Is Essential for Tissue Tension to Ensure Vertebrate 3D Body Shape. Nature 2015, 521, 217–221. [Google Scholar] [CrossRef]
- Song, J.Y.; Park, R.; Kim, J.Y.; Hughes, L.; Lu, L.; Kim, S.; Johnson, R.L.; Cho, S.H. Dual Function of Yap in the Regulation of Lens Progenitor Cells and Cellular Polarity. Dev. Biol. 2014, 386, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L.M.; Horsburgh, G.; Cohen, J. Small Eyes (Sey): A Homozygous Lethal Mutation on Chromosome 2 Which Affects the Differentiation of Both Lens and Nasal Placodes in the Mouse. J. Embryol. Exp. Morphol. 1986, 97, 95–110. [Google Scholar] [CrossRef]
- Ashery-Padan, R.; Marquardt, T.; Zhou, X.; Gruss, P. Pax6 Activity in the Lens Primordium Is Required for Lens Formation and for Correct Placement of a Single Retina in the Eye. Genes Dev. 2000, 14, 2701–2711. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.K.; Haynes, J.I.; Cvekl, A.; Piatigorsky, J. Dual Roles for Pax-6: A Transcriptional Repressor of Lens Fiber Cell-Specific β-Crystallin Genes. Mol. Cell Biol. 1998, 18, 5579–5586. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Cvekl, A. The Orchestration of Mammalian Tissue Morphogenesis through a Series of Coherent Feed-Forward Loops. J. Biol. Chem. 2011, 286, 43259–43271. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Rockowitz, S.; Xie, Q.; Ashery-Padan, R.; Zheng, D.; Cvekl, A. Identification of in Vivo DNA-Binding Mechanisms of Pax6 and Reconstruction of Pax6-Dependent Gene Regulatory Networks during Forebrain and Lens Development. Nucleic Acids Res. 2015, 43, 6827–6846. [Google Scholar] [CrossRef] [PubMed]
- Antosova, B.; Smolikova, J.; Klimova, L.; Lachova, J.; Bendova, M.; Kozmikova, I.; Machon, O.; Kozmik, Z. The Gene Regulatory Network of Lens Induction Is Wired through Meis-Dependent Shadow Enhancers of Pax6. PLoS Genet. 2016, 12, e1006441. [Google Scholar] [CrossRef]
- Wigle, J.T.; Chowdhury, K.; Gruss, P.; Oliver, G. Prox1 Function Is Crucial for Mouse Lens-Fibre Elongation. Nat. Genet. 1999, 21, 318–322. [Google Scholar] [CrossRef]
- Cui, W.; Tomarev, S.I.; Piatigorsky, J.; Chepelinsky, A.B.; Duncan, M.K. Mafs, Prox1, and Pax6 Can Regulate Chicken ΒB1-Crystallin Gene Expression. J. Biol. Chem. 2004, 279, 11088–11095. [Google Scholar] [CrossRef]
- Audette, D.S.; Anand, D.; So, T.; Rubenstein, T.B.; Lachke, S.A.; Lovicu, F.J.; Duncan, M.K. Prox1 and Fibroblast Growth Factor Receptors Form a Novel Regulatory Loop Controlling Lens Fiber Differentiation and Gene Expression. Development 2016, 143, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Madakashira, B.P.; Kobrinski, D.A.; Hancher, A.D.; Arneman, E.C.; Wagner, B.D.; Wang, F.; Shin, H.; Lovicu, F.J.; Reneker, L.W.; Robinson, M.L. Frs2α Enhances Fibroblast Growth Factor-Mediated Survival and Differentiation in Lens Development. Development 2012, 139, 4601–4612. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, M.; Izu, H.; Seki, K.; Fukuda, K.; Nishida, T.; Yamada, S.I.; Kato, K.; Yonemura, S.; Inouye, S.; Nakai, A. HSF4 Is Required for Normal Cell Growth and Differentiation during Mouse Lens Development. EMBO J. 2004, 23, 4297–4306. [Google Scholar] [CrossRef]
- Fujimoto, M.; Oshima, K.; Shinkawa, T.; Bei, B.W.; Inouye, S.; Hayashida, N.; Takii, R.; Nakai, A. Analysis of HSF4 Binding Regions Reveals Its Necessity for Gene Regulation during Development and Heat Shock Response in Mouse Lenses. J. Biol. Chem. 2008, 283, 29961–29970. [Google Scholar] [CrossRef] [PubMed]
- Jing, Z.; Gangalum, R.K.; Bhat, A.M.; Nagaoka, Y.; Jiang, M.; Bhat, S.P. HSF4 Mutation p.Arg116His Found in Age-Related Cataracts and in Normal Populations Produces Childhood Lamellar Cataract in Transgenic Mice. Hum. Mutat. 2014, 35, 1068–1071. [Google Scholar] [CrossRef]
- Maeda, A.; Moriguchi, T.; Hamada, M.; Kusakabe, M.; Fujioka, Y.; Nakano, T.; Yoh, K.; Lim, K.C.; Engel, J.D.; Takahashi, S. Transcription Factor GATA-3 Is Essential for Lens Development. Dev. Dyn. 2009, 238, 2280–2291. [Google Scholar] [CrossRef]
- Martynova, E.; Bouchard, M.; Musil, L.S.; Cvekl, A. Identification of Novel Gata3 Distal Enhancers Active in Mouse Embryonic Lens. Dev. Dyn. 2018, 247, 1186–1198. [Google Scholar] [CrossRef]
- Martynova, E.; Zhao, Y.; Xie, Q.; Zheng, D.; Cvekl, A. Transcriptomic Analysis and Novel Insights into Lens Fibre Cell Differentiation Regulated by Gata3. Open Biol. 2019, 9, 190220. [Google Scholar] [CrossRef] [PubMed]
- Blixt, Å.; Mahlapuu, M.; Aitola, M.; Pelto-Huikko, M.; Enerbäck, S.; Carlsson, P. A Forkhead Gene, FoxE3, Is Essential for Lens Epithelial Proliferation and Closure of the Lens Vesicle. Genes Dev. 2000, 14, 245–254. [Google Scholar] [CrossRef]
- Blixt, Å.; Landgren, H.; Johansson, B.R.; Carlsson, P. Foxe3 Is Required for Morphogenesis and Differentiation of the Anterior Segment of the Eye and Is Sensitive to Pax6 Gene Dosage. Dev. Biol. 2007, 302, 218–229. [Google Scholar] [CrossRef]
- Landgren, H.; Blixt, Å.; Carlsson, P. Persistent FoxE3 Expression Blocks Cytoskeletal Remodeling and Organelle Degradation during Lens Fiber Differentiation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4269–4277. [Google Scholar] [CrossRef]
- Medina-Martinez, O.; Shah, R.; Jamrich, M. Pitx3 Controls Multiple Aspects of Lens Development. Dev. Dyn. 2009, 238, 2193–2201. [Google Scholar] [CrossRef] [PubMed]
- Krall, M.; Htun, S.; Anand, D.; Hart, D.; Lachke, S.A.; Slavotinek, A.M. A Zebrafish Model of Foxe3 Deficiency Demonstrates Lens and Eye Defects with Dysregulation of Key Genes Involved in Cataract Formation in Humans. Hum. Genet. 2018, 137, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Kerr, C.L.; Huang, J.; Williams, T.; West-Mays, J.A. Activation of the Hedgehog Signaling Pathway in the Developing Lens Stimulates Ectopic FoxE3 Expression and Disruption in Fiber Cell Differentiation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3316–3330. [Google Scholar] [CrossRef]
- Medina-Martinez, O.; Brownell, I.; Amaya-Manzanares, F.; Hu, Q.; Behringer, R.R.; Jamrich, M. Severe Defects in Proliferation and Differentiation of Lens Cells in Foxe3 Null Mice. Mol. Cell Biol. 2005, 25, 8854–8863. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Li, T.; Ho, I.C.; Grusby, M.J.; Glimcher, L.H. Requirement for the C-Maf Transcription Factor in Crystallin Gene Regulation and Lens Development. Proc. Natl. Acad. Sci. USA 1999, 96, 3781–3785. [Google Scholar] [CrossRef]
- Kawauchi, S.; Takahashi, S.; Nakajima, O.; Ogino, H.; Morita, M.; Nishizawa, M.; Yasuda, K.; Yamamoto, M. Regulation of Lens Fiber Cell Differentiation by Transcription Factor C- Maf. J. Biol. Chem. 1999, 274, 19254–19260. [Google Scholar] [CrossRef]
- Ring, B.Z.; Cordes, S.P.; Overbeek, P.A.; Barsh, G.S. Regulation of Mouse Lens Fiber Cell Development and Differentiation by the Maf Gene. Development 2000, 127, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, N.; Kerppola, T.K. Synergistic Transcription Activation by Maf and Sox and Their Subnuclear Localization Are Disrupted by a Mutation in Maf That Causes Cataract. Mol. Cell Biol. 2004, 24, 5694–5709. [Google Scholar] [CrossRef]
- Patel, S.D.; Anand, D.; Motohashi, H.; Katsuoka, F.; Yamamoto, M.; Lachke, S.A. Deficiency of the BZIP Transcription Factors Mafg and Mafk Causes Misexpression of Genes in Distinct Pathways and Results in Lens Embryonic Developmental Defects. Front. Cell Dev. Biol. 2022, 10, 981893. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.A.; Anand, D.; Siddam, A.D.; Kakrana, A.; Dash, S.; Scheiblin, D.A.; Dang, C.A.; Terrell, A.M.; Waters, S.M.; Singh, A.; et al. Compound Mouse Mutants of BZIP Transcription Factors Mafg and Mafk Reveal a Regulatory Network of Non-Crystallin Genes Associated with Cataract. Hum. Genet. 2015, 134, 717–735. [Google Scholar] [CrossRef]
- Harris, L.L.; Talian, J.C.; Zelenka, P.S. Contrasting Patterns of C-Myc and N-Myc Expression in Proliferating, Quiescent, and Differentiating Cells of the Embryonic Chicken Lens. Development 1992, 115, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, G.R.; Matos-Rodrigues, G.E.; Zhao, Y.; Gomes, A.L.; Anand, D.; Predes, D.; de Lima, S.; Abreu, J.G.; Zheng, D.; Lachke, S.A.; et al. N-Myc Regulates Growth and Fiber Cell Differentiation in Lens Development. Dev. Biol. 2017, 429, 105–117. [Google Scholar] [CrossRef]
- Semina, E.V.; Reiter, R.S.; Murray, J.C. Isolation of a New Homeobox Gene Belonging to the Pitx/Rieg Family: Expression during Lens Development and Mapping to the Aphakia Region on Mouse Chromosome 19. Hum. Mol. Genet. 1997, 6, 2109–2116. [Google Scholar] [CrossRef]
- Semina, E.V.; Murray, J.C.; Reiter, R.; Hrstka, R.F.; Graw, J. Deletion in the Promoter Region and Altered Expression of Pitx3 Homeobox Gene in Aphakia Mice. Hum. Mol. Genet. 2000, 9, 1575–1585. [Google Scholar] [CrossRef]
- Wada, K.; Matsushima, Y.; Tada, T.; Hasegawa, S.; Obara, Y.; Yoshizawa, Y.; Takahashi, G.; Hiai, H.; Shimanuki, M.; Suzuki, S.; et al. Expression of Truncated PITX3 in the Developing Lens Leads to Microphthalmia and Aphakia in Mice. PLoS ONE 2014, 9, e111432. [Google Scholar] [CrossRef]
- Ahmad, N.; Aslam, M.; Muenster, D.; Horsch, M.; Khan, M.A.; Carlsson, P.; Beckers, J.; Graw, J. Pitx3 Directly Regulates Foxe3 during Early Lens Development. Int. J. Dev. Biol. 2013, 57, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.Y.; Chang, K.H.; Nichols, J.; Li, M. Homeodomain Protein Pitx3 Maintains the Mitotic Activity of Lens Epithelial Cells. Mech. Dev. 2009, 126, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, E.A.; Muheisen, S.; Mlodik, N.; Semina, E.V. MIP/Aquaporin 0 Represents a Direct Transcriptional Target of PITX3 in the Developing Lens. PLoS ONE 2011, 6, e21122. [Google Scholar] [CrossRef] [PubMed]
- Shui, Y.B.; Arbeit, J.M.; Johnson, R.S.; Beebe, D.C. HIF-1: An Age-Dependent Regulator of Lens Cell Proliferation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4961–4970. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Doughman, Y.Q.; Gu, S.; Jarrell, A.; Aota, S.I.; Cvekl, A.; Watanabe, M.; Dunwoodie, S.L.; Johnson, R.S.; van Heyningen, V.; et al. Cited2 Is Required for the Proper Formation of the Hyaloid Vasculature and for Lens Morphogenesis. Development 2008, 135, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Disatham, J.; Kantorow, M. Hypoxia Regulates the Degradation of Non-Nuclear Organelles during Lens Differentiation through Activation of HIF1a. Exp. Eye Res. 2020, 198, 108129. [Google Scholar] [CrossRef] [PubMed]
- Disatham, J.; Brennan, L.; Chauss, D.; Kantorow, J.; Afzali, B.; Kantorow, M. A Functional Map of Genomic HIF1α-DNA Complexes in the Eye Lens Revealed through Multiomics Analysis. BMC Genom. 2021, 22, 497. [Google Scholar] [CrossRef]
- Lang, R.A. Pathways Regulating Lens Induction in the Mouse. Int. J. Dev. Biol. 2004, 48, 783–791. [Google Scholar] [CrossRef]
- Greiling, T.M.S.; Clark, J.I. The Transparent Lens and Cornea in the Mouse and Zebra Fish Eye. Semin. Cell Dev. Biol. 2008, 19, 94–99. [Google Scholar] [CrossRef]
- Cvekl, A.; Ashery-Padan, R. The Cellular and Molecular Mechanisms of Vertebrate Lens Development. Development 2014, 141, 4432–4447. [Google Scholar] [CrossRef] [PubMed]
- Hejtmancik, J.F.; Riazuddin, S.A.; McGreal, R.; Liu, W.; Cvekl, A.; Shiels, A. Lens Biology and Biochemistry. In Proceedings of the Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2015; Volume 134, pp. 169–201. [Google Scholar]
- Piatigorsky, J. Lens Crystallins and Their Genes: Diversity and Tissue-specific Expression. FASEB J. 1989, 3, 1933–1940. [Google Scholar] [CrossRef]
- Graw, J. Genetics of Crystallins: Cataract and Beyond. Exp. Eye Res. 2009, 88, 173–189. [Google Scholar] [CrossRef]
- Cvekl, A.; McGreal, R.; Liu, W. Lens Development and Crystallin Gene Expression. Prog. Mol. Biol. Transl. Sci. 2015, 134, 129–167. [Google Scholar] [CrossRef]
- Piatigorsky, J.; Wistow, G.J. Enzyme/Crystallins: Gene Sharing as an Evolutionary Strategy. Cell 1989, 57, 197–199. [Google Scholar] [CrossRef]
- Piatigorsky, J.; O’Brien, W.E.; Norman, B.L.; Kalumuck, K.; Wistow, G.J.; Borras, T.; Nickerson, J.M.; Wawrousek, E.F. Gene Sharing by δ-Crystallin and Argininosuccinate Lyase. Proc. Natl. Acad. Sci. USA 1988, 85, 3479–3483. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Goto, K.; Okada, T.S.; Kondoh, H. Lens-Specific Enhancer in the Third Intron Regulates Expression of the Chicken Delta 1-Crystallin Gene. Genes Dev. 1987, 1, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Uchikawa, M.; Tanouchi, A.; Sekido, R.; Kondoh, H. Pax6 and SOX2 Form a Co-DNA-Binding Partner Complex That Regulates Initiation of Lens Development. Genes. Dev. 2001, 15, 1272–1286. [Google Scholar] [CrossRef] [PubMed]
- Muta, M.; Kamachi, Y.; Yoshimoto, A.; Higashi, Y.; Kondoh, H. Distinct Roles of SOX2, Pax6 and Maf Transcription Factors in the Regulation of Lens-Specific Δ1-Crystallin Enhancer. Genes Cells 2002, 7, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Cvekl, A.; Sax, C.M.; Li, X.; Mcdermott, J.B.; Piatigorsky, J. Pax-6 and Lens-Specific Transcription of the Chicken Δ1-Crystallin Gene. Proc. Natl. Acad. Sci. USA 1995, 92, 4681–4685. [Google Scholar] [CrossRef]
- Shimada, N.; Aya-Murata, T.; Reza, H.M.; Yasuda, K. Cooperative Action between L-Maf and Sox2 on δ-Crystallin Gene Expression during Chick Lens Development. Mech. Dev. 2003, 120, 455–465. [Google Scholar] [CrossRef]
- Ogino, H.; Yasuda, K. Induction of Lens Differentiation by Activation of a BZIP Transcription Factor, L-Maf. Science 1998, 280, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Disatham, J.; Chauss, D.; Gheyas, R.; Brennan, L.; Blanco, D.; Daley, L.; Menko, A.S.; Kantorow, M. Lens Differentiation Is Characterized by Stage-Specific Changes in Chromatin Accessibility Correlating with Differentiation State-Specific Gene Expression. Dev. Biol. 2019, 453, 86–104. [Google Scholar] [CrossRef]
- Disatham, J.; Brennan, L.; Jiao, X.; Ma, Z.; Hejtmancik, J.F.; Kantorow, M. Changes in DNA Methylation Hallmark Alterations in Chromatin Accessibility and Gene Expression for Eye Lens Differentiation. Epigenet. Chromatin 2022, 15, 1–27. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, D.; Cvekl, A. Profiling of Chromatin Accessibility and Identification of General Cis-Regulatory Mechanisms That Control Two Ocular Lens Differentiation Pathways. Epigenet. Chromatin 2019, 12, 1–23. [Google Scholar] [CrossRef]
- Chang, W.; Zhao, Y.; Rayêe, D.; Xie, Q.; Suzuki, M.; Zheng, D.; Cvekl, A. Dynamic Changes in Whole Genome DNA Methylation, Chromatin and Gene Expression during Mouse Lens Differentiation. Epigenet. Chromatin 2023, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and Quantifying Mammalian Transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential Analysis of Gene Regulation at Transcript Resolution with RNA-Seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential Gene and Transcript Expression Analysis of RNA-Seq Experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Chaitankar, V.; Karakülah, G.; Ratnapriya, R.; Giuste, F.O.; Brooks, M.J.; Swaroop, A. Next Generation Sequencing Technology and Genomewide Data Analysis: Perspectives for Retinal Research. Prog. Retin. Eye Res. 2016, 55, 1–31. [Google Scholar] [CrossRef]
- Hrdlickova, R.; Toloue, M.; Tian, B. RNA-Seq Methods for Transcriptome Analysis. Wiley Interdiscip. Rev. RNA 2017, 8, e1364. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Wang, Z. Next-Generation Transcriptome Assembly. Nat. Rev. Genet. 2011, 12, 671–682. [Google Scholar] [CrossRef]
- Chauss, D.; Basu, S.; Rajakaruna, S.; Ma, Z.; Gau, V.; Anastas, S.; Brennan, L.A.; Hejtmancik, J.F.; Menko, A.S.; Kantorow, M. Differentiation State-Specific Mitochondrial Dynamic Regulatory Networks Are Revealed by Global Transcriptional Analysis of the Developing Chicken Lens. G3 Genes Genomes Genet. 2014, 4, 1515–1527. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, D.; Cvekl, A. A Comprehensive Spatial-Temporal Transcriptomic Analysis of Differentiating Nascent Mouse Lens Epithelial and Fiber Cells. Exp. Eye Res. 2018, 175, 56–72. [Google Scholar] [CrossRef]
- Hoang, T.V.; Raj Kumar, P.K.; Sutharzan, S.; Tsonis, P.A.; Liang, C.; Robinson, M.L. Comparative Transcriptome Analysis of Epithelial and Fiber Cells in Newborn Mouse Lenses with RNA Sequencing. Mol. Vis. 2014, 20, 1491–1517. [Google Scholar]
- Padula, S.L.; Anand, D.; Hoang, T.V.; Chaffee, B.R.; Liu, L.; Liang, C.; Lachke, S.A.; Robinson, M.L. High-Throughput Transcriptome Analysis Reveals That the Loss of Pten Activates a Novel NKX6-1/RASGRP1 Regulatory Module to Rescue Microphthalmia Caused by Fgfr2-Deficient Lenses. Hum. Genet. 2019, 138, 1391–1407. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; An, J.; Wang, J.; Cao, X.; Cao, Y.; Huang, C.; Jiao, S.; Yan, D.; Lin, X.; Zhou, X. Znhit1 Regulates P21Cip1 to Control Mouse Lens Differentiation. Investig. Ophthalmol. Vis. Sci. 2022, 63, 18. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.; Chen, Y.; Davidson, E.A.; Garcia-Milian, R.; Golla, J.P.; Apostolopoulos, N.; Orlicky, D.J.; Schey, K.; Thompson, D.C.; Vasiliou, V. Impaired GSH Biosynthesis Disrupts Eye Development, Lens Morphogenesis and PAX6 Function. Ocul. Surf. 2021, 22, 190–203. [Google Scholar] [CrossRef]
- Zhao, M.; Mei, T.; Shang, B.; Zou, B.; Lian, Q.; Xu, W.; Wu, K.; Lai, Y.; Liu, C.; Wei, L.; et al. Defect of LSS Disrupts Lens Development in Cataractogenesis. Front. Cell. Dev. Biol. 2021, 9, 788422. [Google Scholar] [CrossRef]
- Fan, J.; Lerner, J.; Wyatt, M.K.; Cai, P.; Peterson, K.; Dong, L.; Wistow, G. The Klotho-Related Protein KLPH (Lctl) Has Preferred Expression in Lens and Is Essential for Expression of Clic5 and Normal Lens Suture Formation. Exp. Eye Res. 2018, 169, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.Y.; Ali, M.; Kabir, F.; Na, C.H.; Delannoy, M.; Ma, Y.; Qiu, C.; Costello, M.J.; Hejtmancik, J.F.; Riazuddin, S.A. The Role of FYCO1-Dependent Autophagy in Lens Fiber Cell Differentiation. Autophagy 2022, 18, 2198–2215. [Google Scholar] [CrossRef]
- PIATIGORSKY, J. Lens Differentiation in Vertebrates: A Review of Cellular and Molecular Features. Differentiation 1981, 19, 134–153. [Google Scholar] [CrossRef] [PubMed]
- Audette, D.S.; Scheiblin, D.A.; Duncan, M.K. The Molecular Mechanisms Underlying Lens Fiber Elongation. Exp. Eye Res. 2017, 156, 41–49. [Google Scholar] [CrossRef]
- Cheng, C.; Nowak, R.B.; Fowler, V.M. The Lens Actin Filament Cytoskeleton: Diverse Structures for Complex Functions. Exp. Eye Res. 2017, 156, 58–71. [Google Scholar] [CrossRef]
- Costello, M.J.; Brennan, L.A.; Basu, S.; Chauss, D.; Mohamed, A.; Gilliland, K.O.; Johnsen, S.; Menko, A.S.; Kantorow, M. Autophagy and Mitophagy Participate in Ocular Lens Organelle Degradation. Exp. Eye Res. 2013, 116, 141–150. [Google Scholar] [CrossRef]
- FitzGerald, P.G. Lens Intermediate Filaments. Exp. Eye Res. 2009, 88, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.T.; White, T.W.; Gong, X. Lens Gap Junctions in Growth, Differentiation, and Homeostasis. Physiol. Rev. 2010, 90, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Perng, M.D.; Zhang, Q.; Quinlan, R.A. Insights into the Beaded Filament of the Eye Lens. Exp. Cell Res. 2007, 10, 2180–2188. [Google Scholar] [CrossRef]
- Rao, P.V.; Maddala, R. The Role of the Lens Actin Cytoskeleton in Fiber Cell Elongation and Differentiation. Semin. Cell Dev. Biol. 2006, 17, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.L. An Essential Role for FGF Receptor Signaling in Lens Development. Semin. Cell Dev. Biol. 2006, 17, 726–740. [Google Scholar] [CrossRef]
- Brennan, L.A.; McGreal-Estrada, R.; Logan, C.M.; Cvekl, A.; Menko, A.S.; Kantorow, M. BNIP3L/NIX Is Required for Elimination of Mitochondria, Endoplasmic Reticulum and Golgi Apparatus during Eye Lens Organelle-Free Zone Formation. Exp. Eye Res. 2018, 174, 173–184. [Google Scholar] [CrossRef]
- Wolf, L.V.; Yang, Y.; Wang, J.; Xie, Q.; Braunger, B.; Tamm, E.R.; Zavadil, J.; Cvekl, A. Identification of Pax6-Dependent Gene Regulatory Networks in the Mouse Lens. PLoS ONE 2009, 4, e4159. [Google Scholar] [CrossRef]
- Zhang, J.; Ney, P.A. Role of BNIP3 and NIX in Cell Death, Autophagy, and Mitophagy. Cell Death Differ. 2009, 16, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Al-Bari, M.A.A.; Xu, P. Molecular Regulation of Autophagy Machinery by MTOR-Dependent and -Independent Pathways. Ann. N. Y. Acad. Sci. 2020, 1467, 3–20. [Google Scholar] [CrossRef]
- Upreti, A.; Padula, S.L.; Tangeman, J.A.; Wagner, B.D.; O’Connell, M.J.; Jaquish, T.J.; Palko, R.K.; Mantz, C.J.; Anand, D.; Lovicu, F.J.; et al. Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response. Cells 2023, 12, 501. [Google Scholar] [CrossRef]
- Buenrostro, J.D.; Wu, B.; Litzenburger, U.M.; Ruff, D.; Gonzales, M.L.; Snyder, M.P.; Chang, H.Y.; Greenleaf, W.J. Single-Cell Chromatin Accessibility Reveals Principles of Regulatory Variation. Nature 2015, 523, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of Native Chromatin for Fast and Sensitive Epigenomic Profiling of Open Chromatin, DNA-Binding Proteins and Nucleosome Position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef]
- Wilken, M.S.; Brzezinski, J.A.; La Torre, A.; Siebenthall, K.; Thurman, R.; Sabo, P.; Sandstrom, R.S.; Vierstra, J.; Canfield, T.K.; Hansen, R.S.; et al. DNase I Hypersensitivity Analysis of the Mouse Brain and Retina Identifies Region-Specific Regulatory Elements. Epigenet. Chromatin 2015, 8, 8. [Google Scholar] [CrossRef]
- John, S.; Sabo, P.J.; Canfield, T.K.; Lee, K.; Vong, S.; Weaver, M.; Wang, H.; Vierstra, J.; Reynolds, A.P.; Thurman, R.E.; et al. Genome-Scale Mapping of DNase I Hypersensitivity. Curr. Protoc. Mol. Biol. 2013, 103, 21.27.1–21.27.20. [Google Scholar] [CrossRef]
- Gaulton, K.J.; Nammo, T.; Pasquali, L.; Simon, J.M.; Giresi, P.G.; Fogarty, M.P.; Panhuis, T.M.; Mieczkowski, P.; Secchi, A.; Bosco, D.; et al. A Map of Open Chromatin in Human Pancreatic Islets. Nat. Genet. 2010, 42, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Rockowitz, S.; Chauss, D.; Wang, P.; Kantorow, M.; Zheng, D.; Cvekl, A. Chromatin Features, RNA Polymerase II and the Comparative Expression of Lens Genes Encoding Crystallins, Transcription Factors, and Autophagy Mediators. Mol. Vis. 2015, 21, 955–973. [Google Scholar] [PubMed]
- Grindley, J.C.; Davidson, D.R.; Hill, R.E. The Role of Pax-6 in Eye and Nasal Development. Development 1995, 121, 1433–1442. [Google Scholar] [CrossRef]
- Quinn, J.C.; West, J.D.; Hill, R.E. Multiple Functions for Pax6 in Mouse Eye and Nasal Development. Genes Dev. 1996, 10, 435–446. [Google Scholar] [CrossRef]
- Pontoriero, G.F.; Deschamps, P.; Ashery-Padan, R.; Wong, R.; Yang, Y.; Zavadil, J.; Cvekl, A.; Sullivan, S.; Williams, T.; West-Mays, J.A. Cell Autonomous Roles for AP-2α in Lens Vesicle Separation and Maintenance of the Lens Epithelial Cell Phenotype. Dev. Dyn. 2008, 237, 602–617. [Google Scholar] [CrossRef]
- Brennan, L.; Disatham, J.; Kantorow, M. Mechanisms of Organelle Elimination for Lens Development and Differentiation. Exp. Eye Res. 2021, 209, 108682. [Google Scholar] [CrossRef]
- Zhao, Y.; Wilmarth, P.A.; Cheng, C.; Limi, S.; Fowler, V.M.; Zheng, D.; David, L.L.; Cvekl, A. Proteome-Transcriptome Analysis and Proteome Remodeling in Mouse Lens Epithelium and Fibers. Exp. Eye Res. 2019, 179, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Richiert, D.; Tumminia, S.J.; Russell, P. The Tensegrity Model Applied to the Lens: A Hypothesis for the Presence of the Fiber Cell Ball and Sockets. Med. Hypotheses 2000, 55, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Barton, K.; De Maria, A.; Petrash, J.M.; Shiels, A.; Bassnett, S. The Stratified Syncytium of the Vertebrate Lens. J. Cell Sci. 2009, 122, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Cvekl, A.; Eliscovich, C. Crystallin Gene Expression: Insights from Studies of Transcriptional Bursting. Exp. Eye Res. 2021, 207, 108564. [Google Scholar] [CrossRef]
- Limi, S.; Senecal, A.; Coleman, R.; Lopez-Jones, M.; Guo, P.; Polumbo, C.; Singer, R.H.; Skoultchi, A.I.; Cvekl, A. Transcriptional Burst Fraction and Size Dynamics during Lens Fiber Cell Differentiation and Detailed Insights into the Denucleation Process. J. Biol. Chem. 2018, 293, 13176–13190. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Cheng, C.; Xia, C.H. Connexins in Lens Development and Cataractogenesis. J. Membr. Biol. 2007, 218, 9–12. [Google Scholar] [CrossRef]
- Sjödal, M.; Edlund, T.; Gunhaga, L. Time of Exposure to BMP Signals Plays a Key Role in the Specification of the Olfactory and Lens Placodes Ex Vivo. Dev. Cell 2007, 13, 141–149. [Google Scholar] [CrossRef]
- Boswell, B.A.; Musil, L.S. Synergistic Interaction between the Fibroblast Growth Factor and Bone Morphogenetic Protein Signaling Pathways in Lens Cells. Mol. Biol. Cell 2015, 26, 2561–2572. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Basu, S.; Rajakaruna, S.; Reyes, B.; Van Bockstaele, E.; Menko, A.S. Suppression of MAPK/JNK-MTORC1 Signaling Leads to Premature Loss of Organelles and Nuclei by Autophagy during Terminal Differentiation of Lens Fiber Cells. Autophagy 2014, 10, 1193–1211. [Google Scholar] [CrossRef]
- Brennan, L.; Costello, M.J.; Hejtmancik, J.F.; Menko, A.S.; Riazuddin, S.A.; Shiels, A.; Kantorow, M. Autophagy Requirements for Eye Lens Differentiation and Transparency. Cells 2023, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Beebe, D.C. Maintaining Transparency: A Review of the Developmental Physiology and Pathophysiology of Two Avascular Tissues. Semin. Cell Dev. Biol. 2008, 19, 125–133. [Google Scholar] [CrossRef]
- Weinberg, J.; Gaur, M.; Swaroop, A.; Taylor, A. Proteostasis in Aging-Associated Ocular Disease. Mol. Asp. Med. 2022, 88. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Chang, M.L.; Reznikov, N.; Taylor, A. Disassembly of the Lens Fiber Cell Nucleus to Create a Clear Lens: The P27 Descent. Exp. Eye Res. 2017, 156, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.F.; Menko, A.S. Phosphatidylinositol 3-Kinase Is Necessary for Lens Fiber Cell Differentiation and Survival. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4490–4499. [Google Scholar] [CrossRef]
- Nishiguchi, S.; Wood, H.; Kondoh, H.; Lovell-Badge, R.; Episkopou, V. Sox1 Directly Regulates the γ-Crystallin Genes and Is Essentiat for Lens Development in Mice. Genes Dev. 1998, 12, 776–781. [Google Scholar] [CrossRef]
- Gunhaga, L. The Lens: A Classical Model of Embryonic Induction Providing New Insights into Cell Determination in Early Development. Trans. R. Soc. B Biol. Sci. 2011, 366, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- McLeay, R.C.; Bailey, T.L. Motif Enrichment Analysis: A Unified Framework and an Evaluation on ChIP Data. BMC Bioinform. 2010, 11, 165. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime Cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed]
- West-Mays, J.A.; Coyle, B.M.; Piatigorsky, J.; Papagiotas, S.; Libby, D. Ectopic Expression of AP-2α Transcription Factor in the Lens Disrupts Fiber Cell Differentiation. Dev. Biol. 2002, 245, 13–27. [Google Scholar] [CrossRef]
- Taylor, A.; Clement, T.M.; Eddy, E.M.; Nagata, S.; Chaffee, B.R.; Nakahara, M.; Shang, F.; Wagner, B.D.; Chang, M.-L.; Robinson, M.L. Nuclear Removal during Terminal Lens Fiber Cell Differentiation Requires CDK1 Activity: Appropriating Mitosis-Related Nuclear Disassembly. Development 2014, 17, 3388–3398. [Google Scholar] [CrossRef]
- Taylor, A.; Gu, Y.; Chang, M.L.; Yang, W.; Francisco, S.; Rowan, S.; Bejarano, E.; Pruitt, S.; Zhu, L.; Weiss, G.; et al. Repurposing a Cyclin-Dependent Kinase 1 (CDK1) Mitotic Regulatory Network to Complete Terminal Differentiation in Lens Fiber Cells. Investig. Ophthalmol. Vis. Sci. 2023, 64, 6. [Google Scholar] [CrossRef]
- Zandy, A.J.; Lakhani, S.; Zheng, T.; Flavell, R.A.; Bassnett, S. Role of the Executioner Caspases during Lens Development. J. Biol. Chem. 2005, 280, 30263–30272. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Landsbury, A.; Dahm, R.; Liu, Y.; Zhang, Q.; Quinlan, R.A. Functions of the Intermediate Filament Cytoskeleton in the Eye Lens. J. Clin. Investig. 2009, 119, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, J.; Sun, X.; Martinez-Wittinghan, F.J.; Li, L.; Varadaraj, K.; Mathias, R.T. Effects of GPX-1 Knockout on Membrane Transport and Intracellular Homeostasis in the Lens. J. Membr. Biol. 2009, 227, 25–37. [Google Scholar] [CrossRef]
- Lachke, S.A.; Alkuraya, F.S.; Kneeland, S.C.; Ohn, T.; Aboukhalil, A.; Howell, G.R.; Saadi, I.; Cavallesco, R.; Yue, Y.; Tsai, A.C.-H.; et al. Mutations in the RNA Granule Component TDRD7 Cause Cataract and Glaucoma. Science 2011, 331, 1571–1576. [Google Scholar] [CrossRef]
- Khan, A.; Fornes, O.; Stigliani, A.; Gheorghe, M.; Castro-Mondragon, J.A.; van der Lee, R.; Bessy, A.; Chèneby, J.; Kulkarni, S.R.; Tan, G.; et al. JASPAR 2018: Update of the Open-Access Database of Transcription Factor Binding Profiles and Its Web Framework. Nucleic Acids Res. 2018, 46, D260–D266. [Google Scholar] [CrossRef]
- Vaquerizas, J.M.; Kummerfeld, S.K.; Teichmann, S.A.; Luscombe, N.M. A Census of Human Transcription Factors: Function, Expression and Evolution. Nat. Rev. Genet. 2009, 10, 252–263. [Google Scholar] [CrossRef]
- Berger, M.F.; Badis, G.; Gehrke, A.R.; Talukder, S.; Philippakis, A.A.; Peña-Castillo, L.; Alleyne, T.M.; Mnaimneh, S.; Botvinnik, O.B.; Chan, E.T.; et al. Variation in Homeodomain DNA Binding Revealed by High-Resolution Analysis of Sequence Preferences. Cell 2008, 133, 1266–1276. [Google Scholar] [CrossRef]
- Jolma, A.; Yan, J.; Whitington, T.; Toivonen, J.; Nitta, K.R.; Rastas, P.; Morgunova, E.; Enge, M.; Taipale, M.; Wei, G.; et al. DNA-Binding Specificities of Human Transcription Factors. Cell 2013, 152, 327–339. [Google Scholar] [CrossRef]
- Jolma, A.; Yin, Y.; Nitta, K.R.; Dave, K.; Popov, A.; Taipale, M.; Enge, M.; Kivioja, T.; Morgunova, E.; Taipale, J. DNA-Dependent Formation of Transcription Factor Pairs Alters Their Binding Specificity. Nature 2015, 527, 384–388. [Google Scholar] [CrossRef]
- Nitta, K.R.; Jolma, A.; Yin, Y.; Morgunova, E.; Kivioja, T.; Akhtar, J.; Hens, K.; Toivonen, J.; Deplancke, B.; Furlong, E.E.M.; et al. Conservation of Transcription Factor Binding Specificities across 600 Million Years of Bilateria Evolution. Elife 2015, 2015, e04837. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Kudo, T.; Ogata, K.; Hamada, M.; Nakamura, M.; Kito, K.; Abe, Y.; Ueda, N.; Yamamoto, M.; Engel, J.D.; et al. Neither MafA/L-Maf nor MafB Is Essential for Lens Development in Mice. Genes Cells 2009, 14, 941–947. [Google Scholar] [CrossRef]
- Johnsen, Ø.; Murphy, P.; Prydz, H.; Kolstø, A.B. Interaction of the CNC-BZIP Factor TCF11/LCR-F1/Nrf1 with MafG: Binding-Site Selection and Regulation of Transcription. Nucleic Acids Res. 1998, 26, 512–520. [Google Scholar] [CrossRef]
- Johnsen, Ø.; Skammelsrud, N.; Luna, L.; Nishizawa, M.; Prydz, H.; Kolsto, A.B. Small Maf Proteins Interact with the Human Transcription Factor TCF11/Nrf1/LCR-F1. Nucleic Acids Res. 1996, 24, 4289–4297. [Google Scholar] [CrossRef] [PubMed]
- Sans, C.L.; Satterwhite, D.J.; Stoltzman, C.A.; Breen, K.T.; Ayer, D.E. MondoA-Mlx Heterodimers Are Candidate Sensors of Cellular Energy Status: Mitochondrial Localization and Direct Regulation of Glycolysis. Mol. Cell Biol. 2006, 26, 4863–4871. [Google Scholar] [CrossRef]
- Eilers, A.L.; Sundwall, E.; Lin, M.; Sullivan, A.A.; Ayer, D.E. A Novel Heterodimerization Domain, CRM1, and 14-3-3 Control Subcellular Localization of the MondoA-Mlx Heterocomplex. Mol. Cell Biol. 2002, 22, 8514–8526. [Google Scholar] [CrossRef] [PubMed]
- Ebong, S.; Chepelinsky, A.B.; Robinson, M.L.; Zhao, H.; Yu, C.R.; Egwuagu, C.E. Characterization of the Roles of STAT1 and STAT3 Signal Transduction Pathways in Mammalian Lens Development. Mol. Vis. 2004, 10, 122–131. [Google Scholar]
- Yang, Y.; Cvekl, A. Large Maf Transcription Factors: Cousins of AP-1 Proteins and Important Regulators of Cellular Differentiation. Einstein J. Biol. Med. Albert 2016, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, H.; Igarashi, K. Expression of the Oxidative Stress-Regulated Transcription Factor Bach2 in Differentiating Neuronal Cells. J. Biochem. 2002, 132, 427–431. [Google Scholar] [CrossRef]
- Kröll-Hermi, A.; Ebstein, F.; Stoetzel, C.; Geoffroy, V.; Schaefer, E.; Scheidecker, S.; Bär, S.; Takamiya, M.; Kawakami, K.; Zieba, B.A.; et al. Proteasome Subunit PSMC3 Variants Cause Neurosensory Syndrome Combining Deafness and Cataract Due to Proteotoxic Stress. EMBO Mol. Med. 2020, 12, e11861. [Google Scholar] [CrossRef]
- Héberlé, É.; Bardet, A.F. Sensitivity of Transcription Factors to DNA Methylation. Essays Biochem. 2019, 63, 727–741. [Google Scholar] [PubMed]
- Yin, Y.; Morgunova, E.; Jolma, A.; Kaasinen, E.; Sahu, B.; Khund-Sayeed, S.; Das, P.K.; Kivioja, T.; Dave, K.; Zhong, F.; et al. Impact of Cytosine Methylation on DNA Binding Specificities of Human Transcription Factors. Science 2017, 356i, eaaj2239. [Google Scholar] [CrossRef]
- Domcke, S.; Bardet, A.F.; Adrian Ginno, P.; Hartl, D.; Burger, L.; Schübeler, D. Competition between DNA Methylation and Transcription Factors Determines Binding of NRF1. Nature 2015, 528, 575–579. [Google Scholar] [CrossRef]
- Wang, H.; Maurano, M.T.; Qu, H.; Varley, K.E.; Gertz, J.; Pauli, F.; Lee, K.; Canfield, T.; Weaver, M.; Sandstrom, R.; et al. Widespread Plasticity in CTCF Occupancy Linked to DNA Methylation. Genome Res. 2012, 22, 1680–1688. [Google Scholar] [CrossRef]
- Harris, R.A.; Wang, T.; Coarfa, C.; Nagarajan, R.P.; Hong, C.; Downey, S.L.; Johnson, B.E.; Fouse, S.D.; Delaney, A.; Zhao, Y.; et al. Comparison of Sequencing-Based Methods to Profile DNA Methylation and Identification of Monoallelic Epigenetic Modifications. Nat. Biotechnol. 2010, 28, 1097–1105. [Google Scholar] [CrossRef]
- Cedar, H.; Bergman, Y. Linking DNA Methylation and Histone Modification: Patterns and Paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA Methylation: Islands, Start Sites, Gene Bodies and Beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. DNA Methylation Patterns and Epigenetic Memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Donner, A.L.; Episkopou, V.; Maas, R.L. Sox2 and Pou2f1 Interact to Control Lens and Olfactory Placode Development. Dev. Biol. 2007, 303, 784–799. [Google Scholar] [CrossRef]
- Yoshida, T.; Yasuda, K. Characterization of the Chicken L-Maf, MafB and c-Maf in Crystallin Gene Regulation and Lens Differentiation. Genes Cells 2002, 7, 693–706. [Google Scholar] [CrossRef]
- Shiels, A.; Bennett, T.M.; Hejtmancik, J.F. Cat-Map: Putting Cataract on the Map. Mol. Vis. 2010, 16, 2007–2015. [Google Scholar] [PubMed]
- Kakrana, A.; Yang, A.; Anand, D.; Djordjevic, D.; Ramachandruni, D.; Singh, A.; Huang, H.; Ho, J.W.K.; Lachke, S.A. ISyTE 2.0: A Database for Expression-Based Gene Discovery in the Eye. Nucleic Acids. Res. 2018, 46, D875–D885. [Google Scholar] [CrossRef]
- Girelli, D.; Corrocher, R.; Bisceglia, L.; Olivieri, O.; De Franceschi, L.; Zelante, L.; Gasparini, P. Molecular Basis for the Recently Described Hereditary Hyperferritinemia- Cataract Syndrome: A Mutation in the Iron-Responsive Element of Ferritin L- Subunit Gene (the ‘Verona Mutation’). Blood 1995, 86, 4050–4053. [Google Scholar] [CrossRef] [PubMed]
- Zuercher, J.; Neidhardt, J.; Magyar, I.; Labs, S.; Moore, A.T.; Tanner, F.C.; Waseem, N.; Schorderet, D.F.; Munier, F.L.; Bhattacharya, S.; et al. Alterations of the 5’untranslated Region of SLC16A12 Lead to Age-Related Cataract. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3354–3361. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Wysocka, J. Deciphering the Multi-Scale, Quantitative Cis-Regulatory Code. Mol. Cell 2023, 83, 373–392. [Google Scholar] [CrossRef]
- Kvon, E.Z.; Waymack, R.; Gad, M.; Wunderlich, Z. Enhancer Redundancy in Development and Disease. Nat. Rev. Genet. 2021, 22, 324–336. [Google Scholar] [CrossRef]
- Long, H.K.; Prescott, S.L.; Wysocka, J. Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution. Cell 2016, 167, 1170–1187. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.G.; Farnham, P.J. Making Sense of GWAS: Using Epigenomics and Genome Engineering to Understand the Functional Relevance of SNPs in Non-Coding Regions of the Human Genome. Epigenet. Chromatin 2015, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lupski, J.R. Non-Coding Genetic Variants in Human Disease. Hum. Mol. Genet. 2015, 24, R102–R110. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-Wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef]
- Argelaguet, R.; Clark, S.J.; Mohammed, H.; Stapel, L.C.; Krueger, C.; Kapourani, C.A.; Imaz-Rosshandler, I.; Lohoff, T.; Xiang, Y.; Hanna, C.W.; et al. Multi-Omics Profiling of Mouse Gastrulation at Single-Cell Resolution. Nature 2019, 576, 487–491. [Google Scholar] [CrossRef]
- Coleman, R.A.; Liu, Z.; Darzacq, X.; Tjian, R.; Singer, R.H.; Lionnet, T. Imaging Transcription: Past, Present, and Future. In Proceedings of the Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring, NY, USA, 2016; Volume 80, pp. 1–8. [Google Scholar]
- Dahal, L.; Walther, N.; Tjian, R.; Darzacq, X.; Graham, T.G.W. Single-Molecule Tracking (SMT): A Window into Live-Cell Transcription Biochemistry. Biochem. Soc. Trans. 2023, BST20221242. [Google Scholar] [CrossRef] [PubMed]
- Misteli, T. The Self-Organizing Genome: Principles of Genome Architecture and Function. Cell 2020, 183, 28–45. [Google Scholar] [CrossRef]
- Dekker, J.; Marti-Renom, M.A.; Mirny, L.A. Exploring the Three-Dimensional Organization of Genomes: Interpreting Chromatin Interaction Data. Nat. Rev. Genet. 2013, 14, 390–403. [Google Scholar] [CrossRef]
- Stadhouders, R.; Filion, G.J.; Graf, T. Transcription Factors and 3D Genome Conformation in Cell-Fate Decisions. Nature 2019, 569, 345–354. [Google Scholar] [CrossRef]
- Nagano, T.; Lubling, Y.; Stevens, T.J.; Schoenfelder, S.; Yaffe, E.; Dean, W.; Laue, E.D.; Tanay, A.; Fraser, P. Single-Cell Hi-C Reveals Cell-to-Cell Variability in Chromosome Structure. Nature 2013, 502, 59–64. [Google Scholar] [CrossRef]
- Durand, N.C.; Shamim, M.S.; Machol, I.; Rao, S.S.P.; Huntley, M.H.; Lander, E.S.; Aiden, E.L. Juicer Provides a One-Click System for Analyzing Loop-Resolution Hi-C Experiments. Cell Syst. 2016, 3, 95–98. [Google Scholar] [CrossRef]
- McGreal-Estrada, R.S.; Wolf, L.V.; Cvekl, A. Promoter-Enhancer Looping and Shadow Enhancers of the Mouse AA-Crystallin Locus. Biol. Open 2018, 7, bio036897. [Google Scholar] [CrossRef] [PubMed]
- Lachke, S.A. RNA-Binding Proteins and Post-Transcriptional Regulation in Lens Biology and Cataract: Mediating Spatiotemporal Expression of Key Factors That Control the Cell Cycle, Transcription, Cytoskeleton and Transparency. Exp. Eye Res. 2022, 214, 108889. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Siddam, A.D.; Barnum, C.E.; Janga, S.C.; Lachke, S.A. RNA-Binding Proteins in Eye Development and Disease: Implication of Conserved RNA Granule Components. Wiley Interdiscip. Rev. RNA 2016, 7, 527–557. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
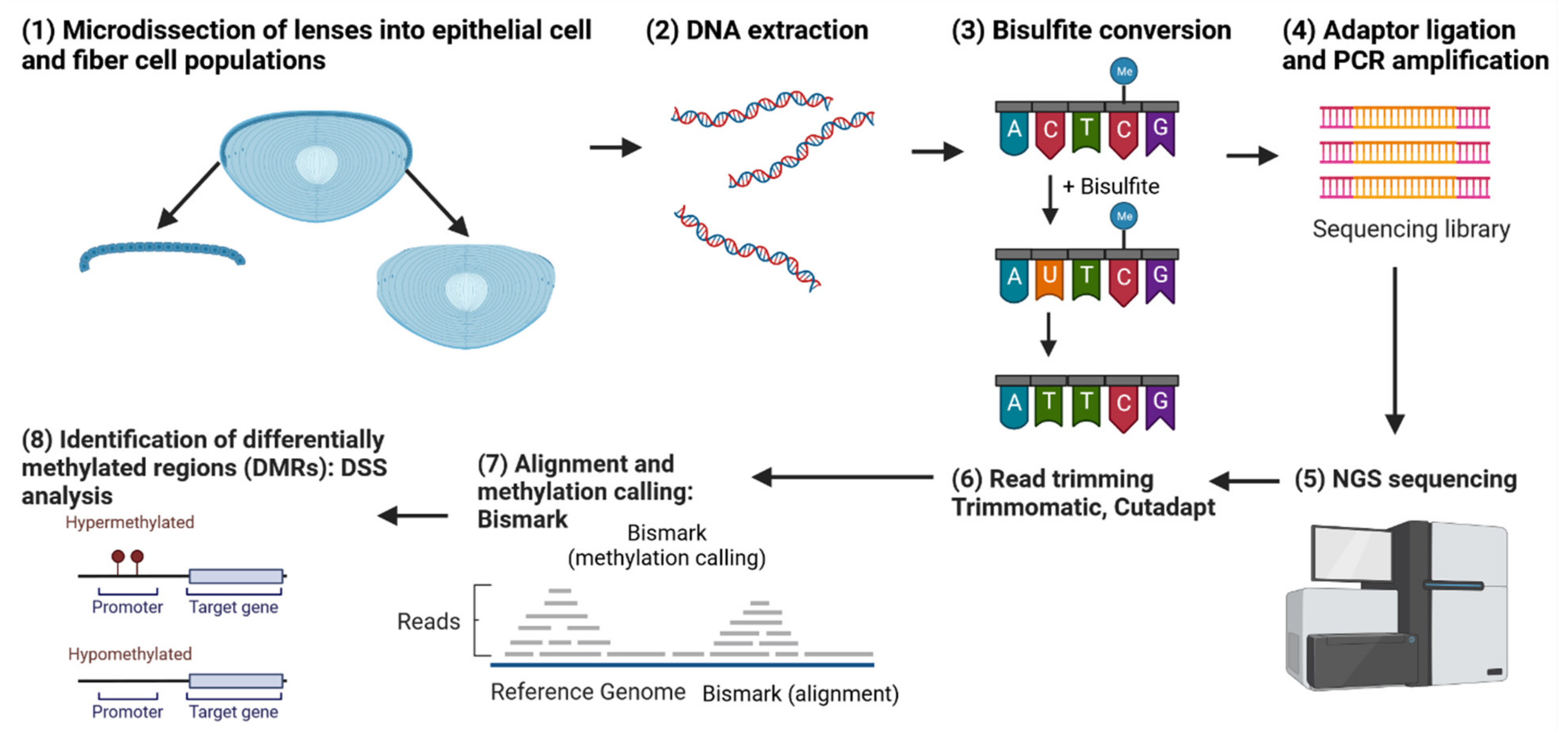{kind=link}
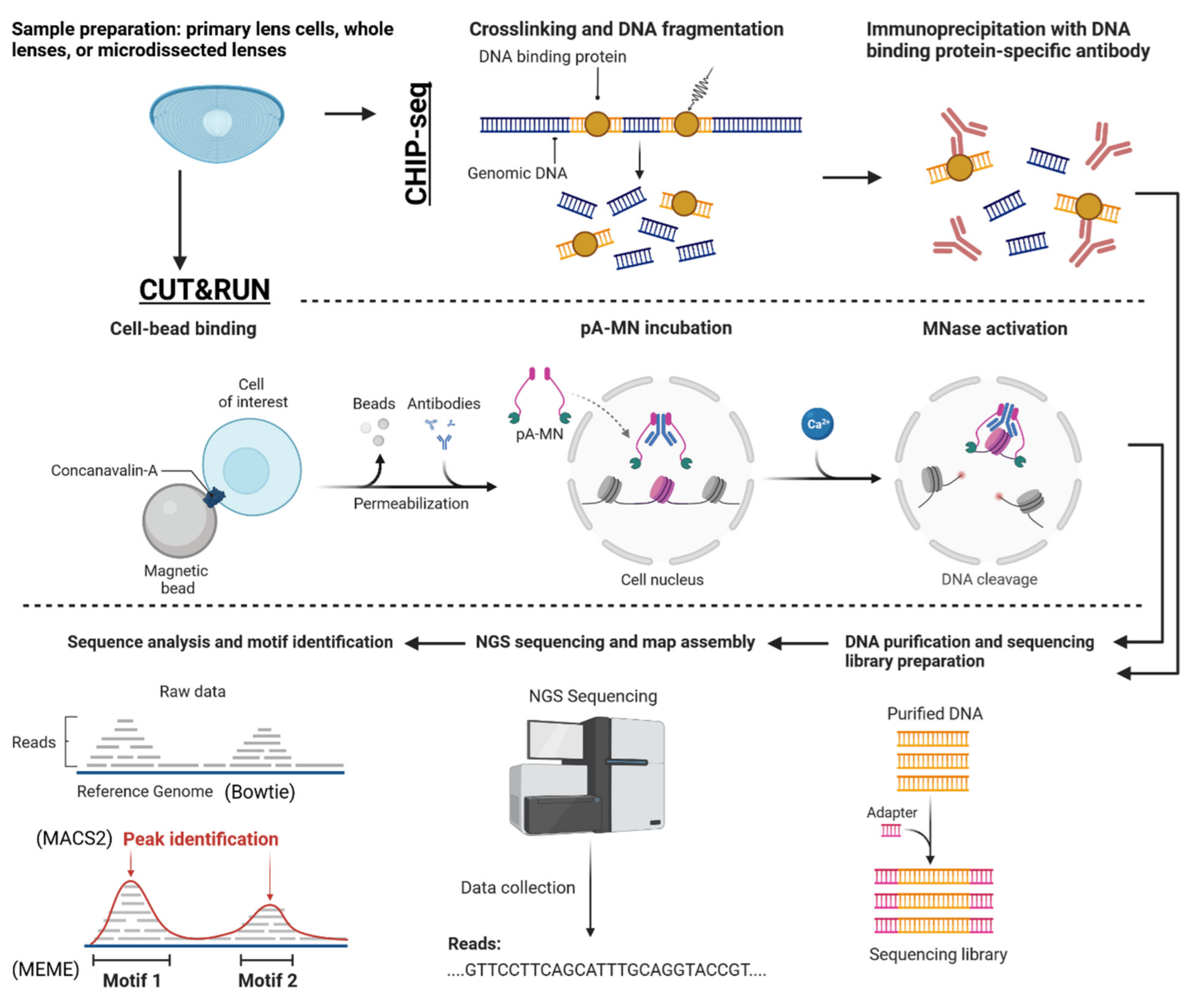{kind=link}
| Study | Model System | Notable Epithelial Preferred Genes | Notable Fiber Preferred Genes | PubmedID |
|---|---|---|---|---|
| Chauss et al., 2014 | E13 chick lenses microdissected into 4 differentiation-state specific regions. Epithelial (EC, EQ) compared to Fiber (FP, FC) | Select Lens Crystallins (CRYAA) Cell Cycle (CDK2) Lens Signaling (FGFR2) Transcription Factors (PAX6) Mitophagy (PARL, SMURF1) Mitochondrial Regulation (BID, PPARGC1A) Macroautophagy (MAP1LC3C, PIK3CB) Mitochondrial Repair/Protection (TXNRD1) | Select Lens Crystallins (ASL1, CRYAB) Actin-Capping (TMOD1) Beaded Filaments (BFSP1, BFSP2) Lens Signaling (EPHA2, CCDC80, FZD3) Transcription Factors (HSF4, SOX2, PROX1) Cataract-associated genes (LGSN) Mitophagy (BNIP3L, BNIP3) Mitochondrial Regulation (DNAJA3, DNAJC15, SLC25A22, TFB1M, SNPH) Macroautophagy (BECN1, FYCO1, MTOR, ATG3, ATG4B, SQSTM1, WIP11) Mitochondrial Repair/Protection (GLRX, MSRA, NXNL1) | 24928582 |
| Zhao et al., 2018 | E14.5, E16.5, E18.5, and P0.5 mouse lenses microdissected into Epithelial compared to Fiber | Transcription Factors (Pax6, Foxe3, Dmrta2, Mafb, Tgap2a, Tulp3) Sumoylation (Sumo1, Sumo3, Sae1, Uba2) | Crystallin Protein Translation Initiation (Eif3h, Eif3k) Transcription Factors (Stat5a, Prox1, Sox1, Maf, Hsf4, Mafg, Arid3b, Bach2, Carhsp1) Macroautophagy (Mtor, Plekhm1) Mitophagy (Bnip3) | 29883638 |
| Disatham et al., 2022 | E13 chick lenses microdissected into Epithelial compared to Fiber | Crystallins (CRYBB2) Cell Cycle (CCND1) DNA Methylases (DNMT3B) DNA demethylases (TET1) Signaling (NOTCH2) Transporters (SLC2A1) Metabolism (LDHA, ENO1) Transcription Factors (HIF1a, RBPJ, HES1, MYCN, HEY1, CREB3L1, CREB3L2, FOXP1, RREB1) | Crystallins (CRYBB3, CRYBA1, ASL1, CRYGN, CRYBB1, CRYBA4) Transcription Factors (HSF4) Beaded Filaments (BFSP1, BFSP2) Mitophagy (BNIP3L) Connexins (GJA3) mRNA processing (TDRD7) Transcription Factors (SOX2, STAT3, MLX, Arid5a) | 35246225 |
| Hoang et al., 2014 | P0 mouse lenses microdissected into Epithelial compared to Fiber | Tyrosine Kinase Receptors (Drd1, Pdgfra, Ror1, Ephb2, Ephb4, Erbb2, Ptk7, Tek, Pdgfrb, Axl, Ephb6, Epha7, Epha10, Kdr, Flt4, Egfr, Tie1, Ror2, Epha3, Epha1, Flt1) Notch Signaling (Notch1, Notch2, Notch3, Notch4, Jag2, Dll4, Dll1, Hes6, Hes1, Rbpj, Hes5, Numb) Wnt Signaling (Fzd1, Fzd7, Fzd2, Fzd4, Fzd8, Wnt5a, Dvl2) TGFB superfamily (Tgfbr1, Tgfbr2, Ncam1, Acvr1, Eng, Bmp1, Tgfb2, Bmp7, Inha) Dna degradation and repair (H2afx, Rad52, Lig1, Rad50, Nbn, Mdc1, Atm, Chek1, Rad51) Proteolysis (Casp2, Casp9, Xiap, Birc5) Aquaporins (Aqp1, Aqp4, Aqp8) Gap junctions (Gja1, Gjc1, Gja4) Intermediate filaments (Lmnb1, Lmnb2, Nes) | Tyrosine Kinase Receptors (Fgfr3, Epha2, Met, Ret) Notch Signaling (Jag1, Herpud1) Wnt Signaling (Wnt7a, Wnt7b, Wnt5b) TGFB superfamily (Bmpr1b) DNA degradation and repair (Dnase2b, Lig4) Proteolysis (Casp7, Birc7, Birc2) Aquaporins (Mip) Gap junctions (Gja3, Gja8, Gje1) Intermediate filaments (Bfsp1, Bfsp2) | 25489224 |
| Study | Model system | Notable Epithelial Preferred Signaling Pathways and Ontologies | Notable Fiber Preferred Signaling Pathways and Ontologies | PubmedID |
|---|---|---|---|---|
| Zhao et al., 2018 | E14.5, E16.5, E18.5, and P0.5 mouse lenses microdissected into Epithelial compared to Fiber | Cell cycle, cell adhesion, signal transduction, DNA repair, DNA methylation, extracellular matrix organization, cell differentiation, cell migration, Wnt signaling, Sumoylation Pathway, Notch signaling, TGFB signaling, BMP signaling, RAR activation, ATM signaling, Human embryonic stem cell pluripotency | Translation initiation, vesicle-mediated transport, oxidation-reduction process, mitochondrial translation, lipid metabolism, autophagy, protein ubiquitination, mitochondrial dysfunction, EIF2 signaling, Oxidative phosphorylation, mTOR signaling, PI3K/Akt signaling, Glycolysis | 29883638 |
| Disatham et al., 2022 | E13 chick lenses microdissected into Epithelial compared to Fiber | Extracellular matrix organization, collagen fibril organization, eye development, epithelial mesenchymal transition, unfolded protein response, G2-M checkpoint, cell cycle control, Notch signaling | Oxidative phosphorylation, Apoptosis, mTORC1 signaling, cholesterol homeostasis, TGF-beta signaling, hypoxia, heme metabolism, intermediate filament organization, hedgehog signaling | 35246225 |
| Hoang et al., 2014 | P0 mouse lenses microdissected into Epithelial compared to Fiber | Cell cycle, cell division, cell migration | Lens development in camera-type eye | 25489224 |
| Study | Model System | Transcription Factors Associated with Differentially Expressed Genes | PubmedID |
|---|---|---|---|
| Disatham et al., 2019 | E13 chick lenses microdissected into 4 differentiation-state specific regions. Epithelial (EC, EQ) compared to Fiber (FP, FC) | NFATC2, IRF1, NFAT5, ZNF384, FOXP2, CTCF, FOXK1, FOXP1, FOXK2, Arid3b, NFATC3, RBPJ, E2F6, SP2, NFATC1, ARNT::HIF1a, Hoxd9, FOXO4, Foxj2, MEF2C, Myod1, TEAD1, SPIC, FOXI1, FOXB1 | 31136738 |
| Zhao et al., 2019 | E14.5 and P0.5 mice lenses microdissected into Epithelial compared to Fiber | Etv5, Foxk2, Foxn3, Foxo3, Gata3, Gatad1, Hsf4, Jun, Maf, Mafa, Mafg, Mafk, Meis2, Mycl, Mycn, Nf2, Nfat5, Rxra, Smad1, Smad4, Sox1, Sox13, Tead1, Prox1, Foxj3, Yy1, Nfatc1, Sox8, Sp1, CTCF, Pax6 | 31053165 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Disatham, J.; Brennan, L.; Cvekl, A.; Kantorow, M. Multiomics Analysis Reveals Novel Genetic Determinants for Lens Differentiation, Structure, and Transparency. Biomolecules 2023, 13, 693. https://doi.org/10.3390/biom13040693
Disatham J, Brennan L, Cvekl A, Kantorow M. Multiomics Analysis Reveals Novel Genetic Determinants for Lens Differentiation, Structure, and Transparency. Biomolecules. 2023; 13(4):693. https://doi.org/10.3390/biom13040693
Chicago/Turabian StyleDisatham, Joshua, Lisa Brennan, Ales Cvekl, and Marc Kantorow. 2023. "Multiomics Analysis Reveals Novel Genetic Determinants for Lens Differentiation, Structure, and Transparency" Biomolecules 13, no. 4: 693. https://doi.org/10.3390/biom13040693