
Tyrosinase Magnetic Cross-Linked Enzyme Aggregates: Biocatalytic Study in Deep Eutectic Solvent Aqueous Solutions
 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Tyrosinase CLEAs and mCLEAs
2.2.2. Characterization of Tyrosinase CLEAs/mCLEAs
2.2.3. Preparation of DESs
2.2.4. Activity Assays in DES Aqueous Solutions
2.2.5. Kinetic Studies
2.2.6. Stability of Free Tyrosinase/Tyrosinase mCLEAs in DESs
2.2.7. Reusability of Tyrosinase mCLEAs
2.2.8. Storage Stability of Tyrosinase mCLEAs
2.2.9. Fluorescence Study
2.2.10. Circular Dichroism Analysis
2.2.11. Homogeneous Enzymatic Modification of Chitosan with Caffeic Acid
2.2.12. Characterization of Chitosan Films
2.2.13. Antioxidant Activity of Chitosan Films
3. Results and Discussion
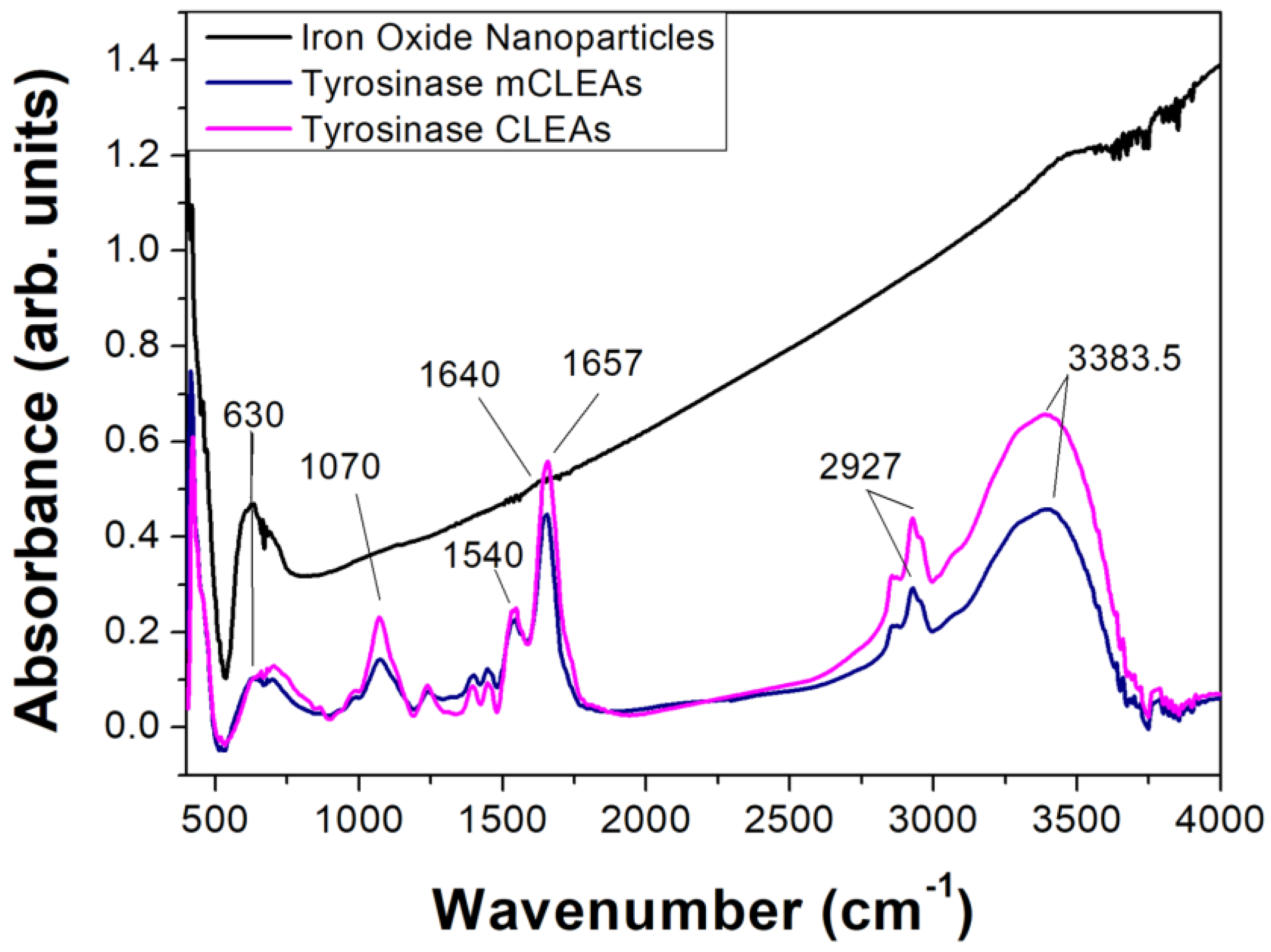
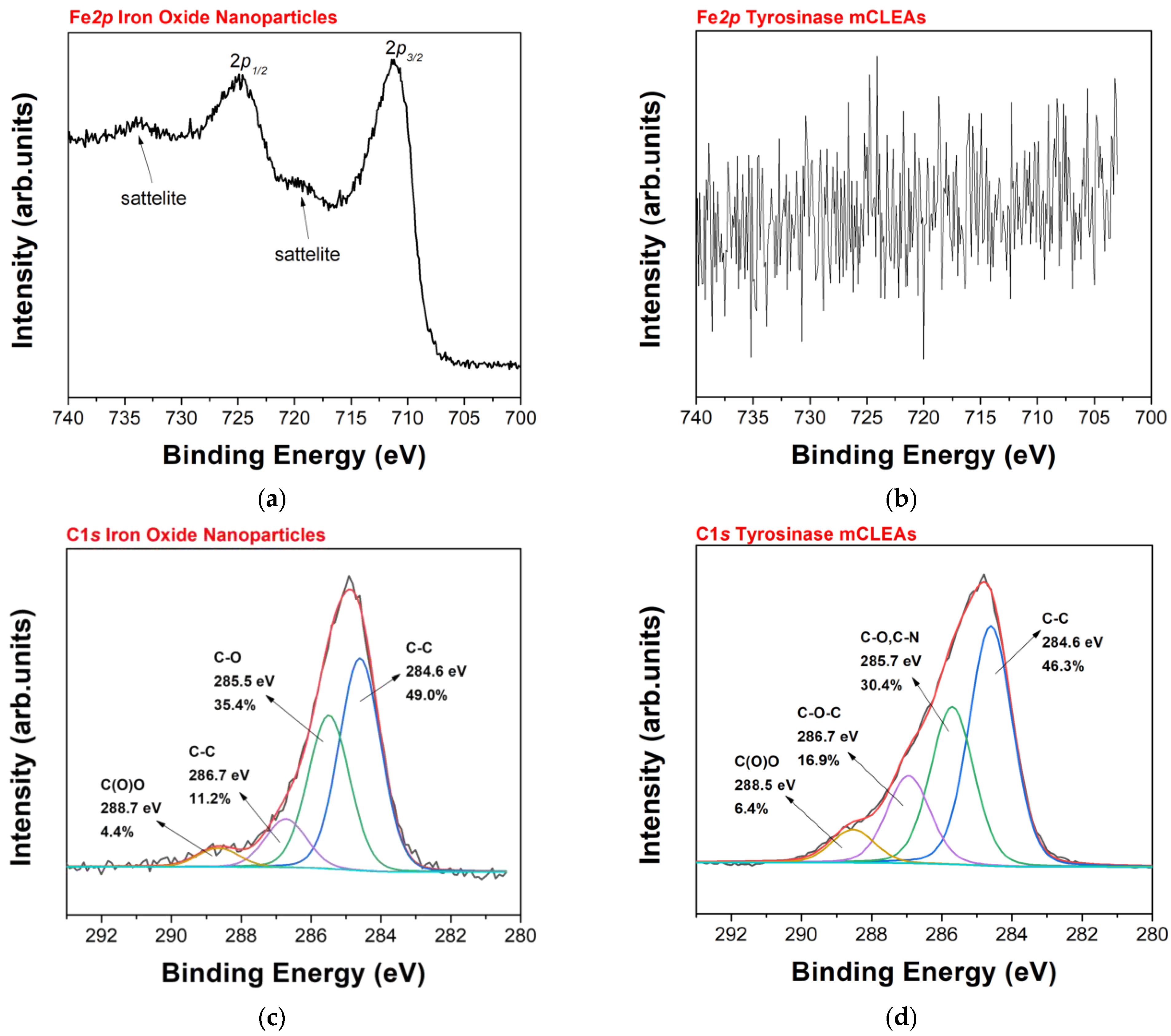
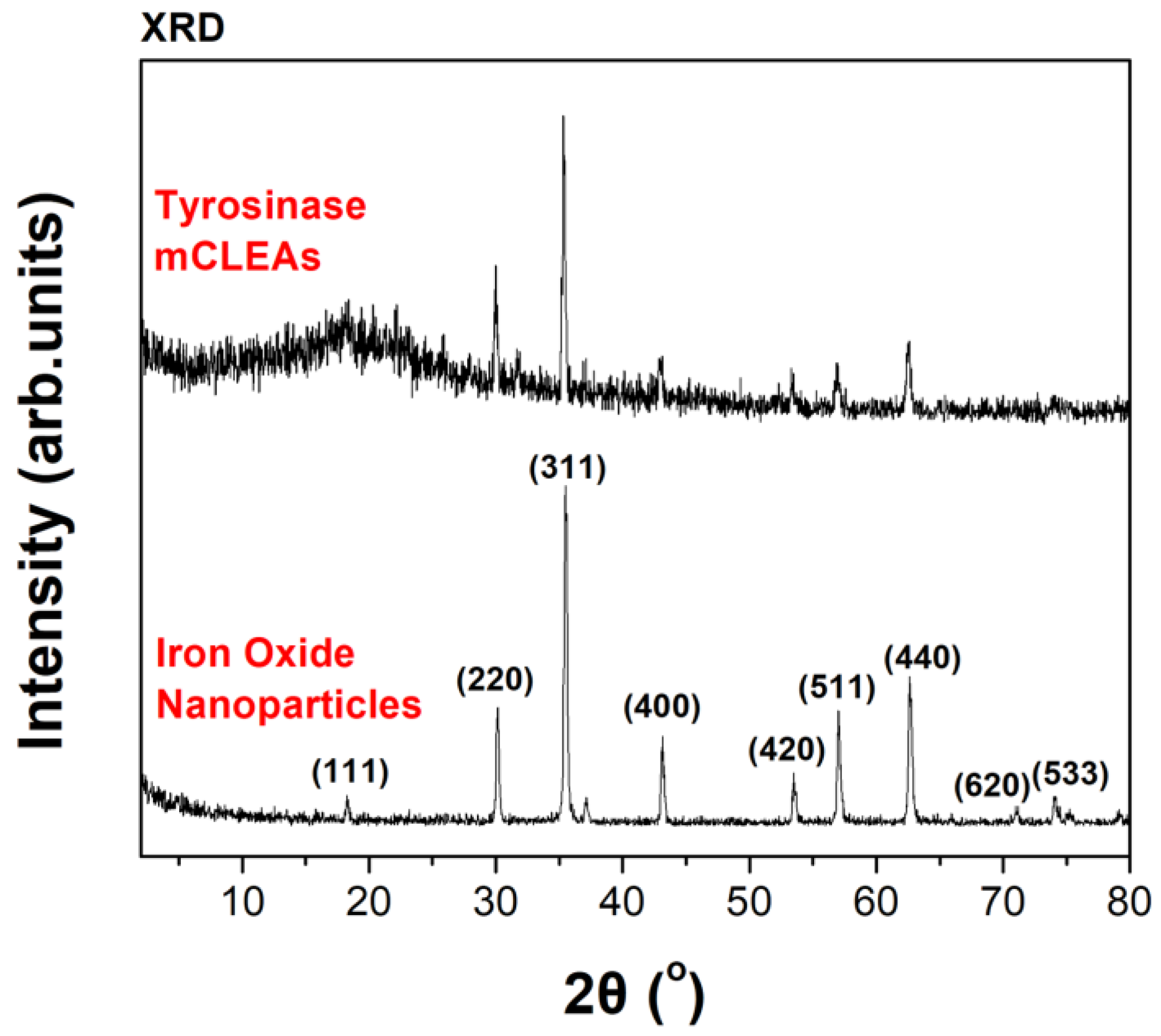
3.1. Preparation and Characterization of Tyrosinase CLEAs/mCLEAS
3.2. Biocatalytic Characteristics of Free and Immobilized Tyrosinase in DES Aqueous Solutions
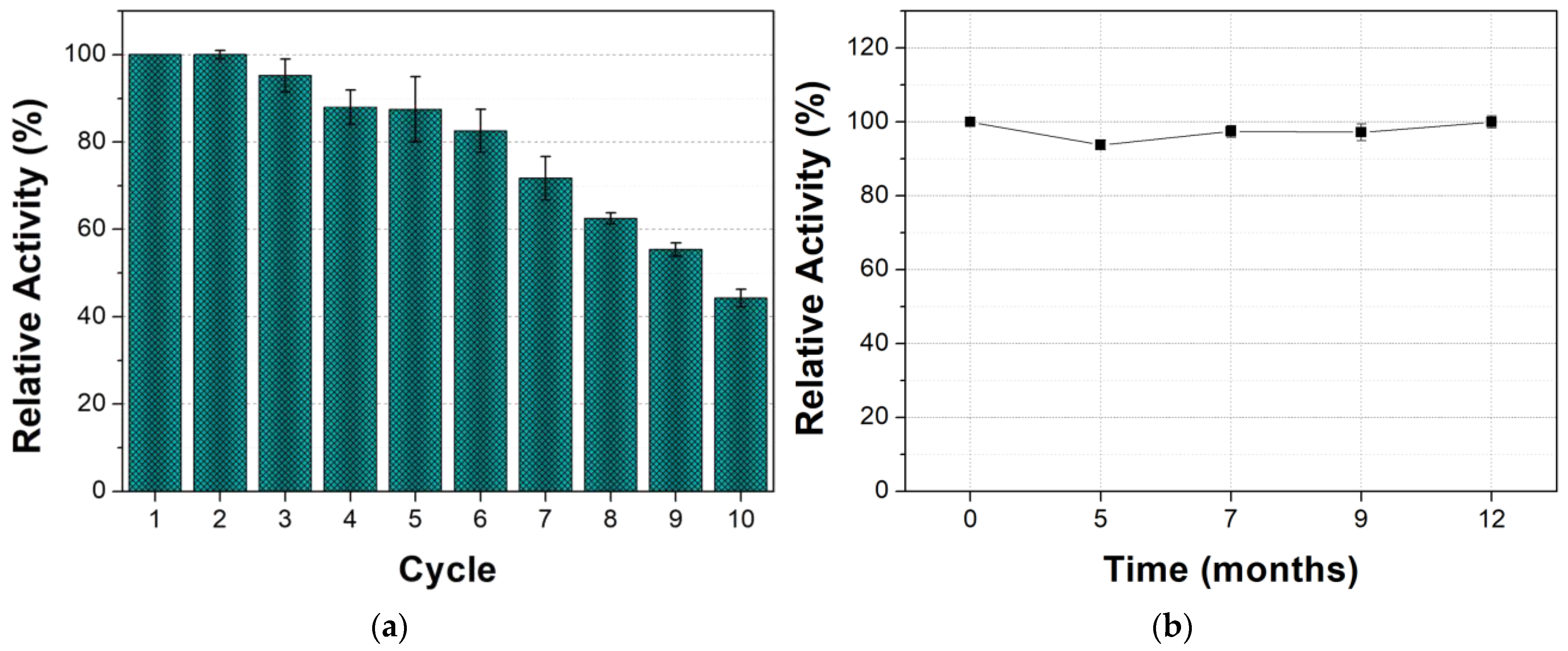
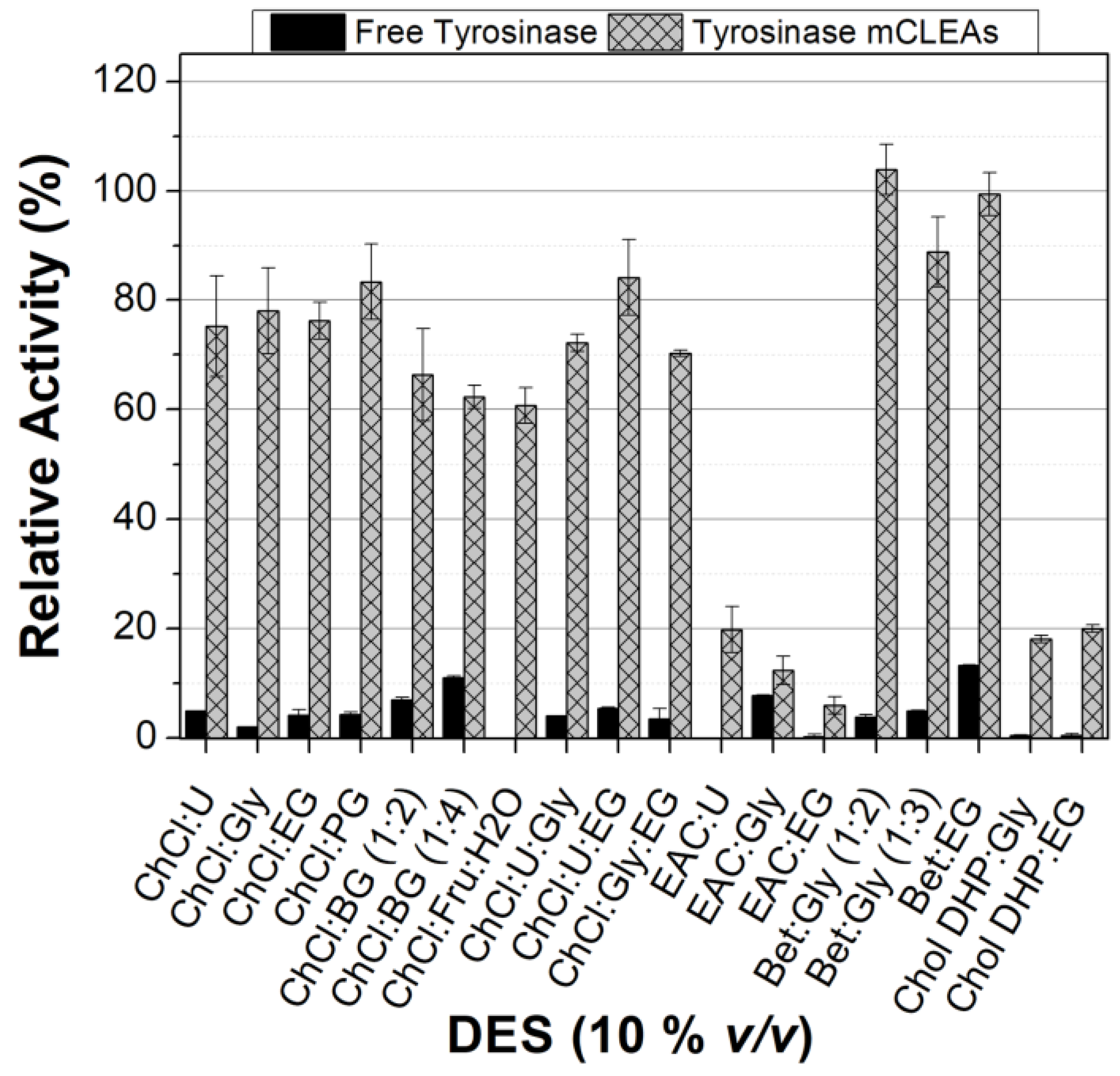
3.2.1. Reusability and Storage Stability of Tyrosinase mCLEAs
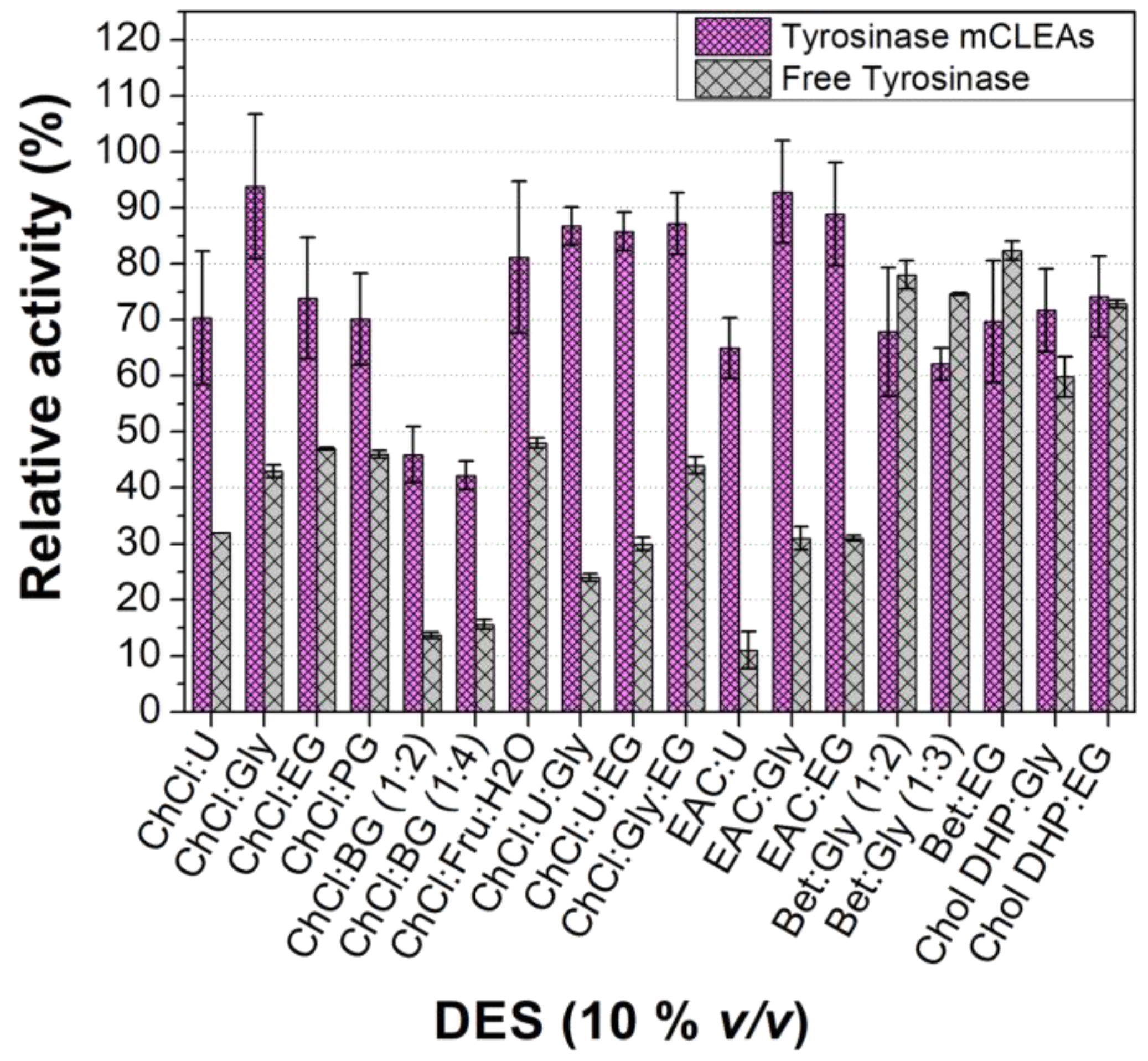
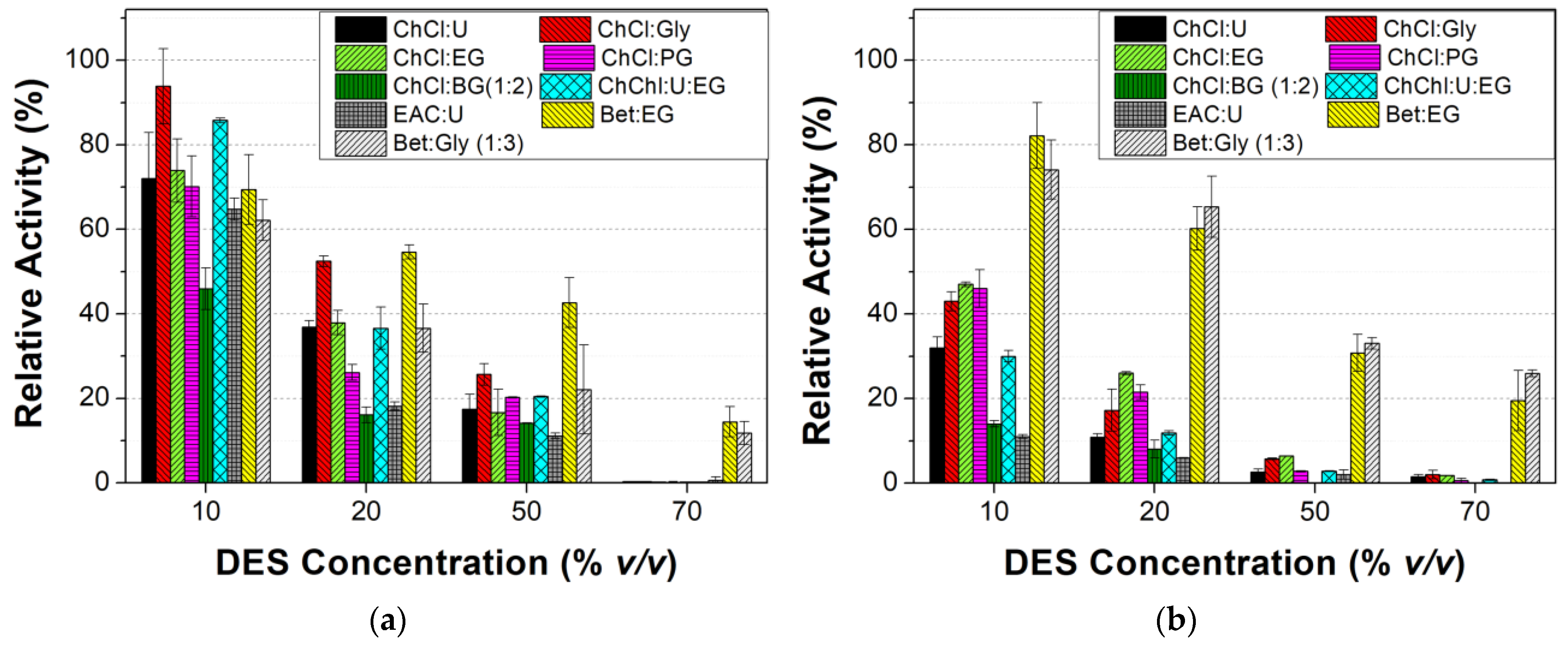
3.2.2. Effect of DES Nature and Concentration on the Tyrosinase Activity
3.2.3. Kinetic Study of Tyrosinase mCLEAs in the Presence of Various DESs
3.2.4. Effect of DESs on the Stability of Tyrosinase
3.3. Structural Studies of Free Tyrosinase
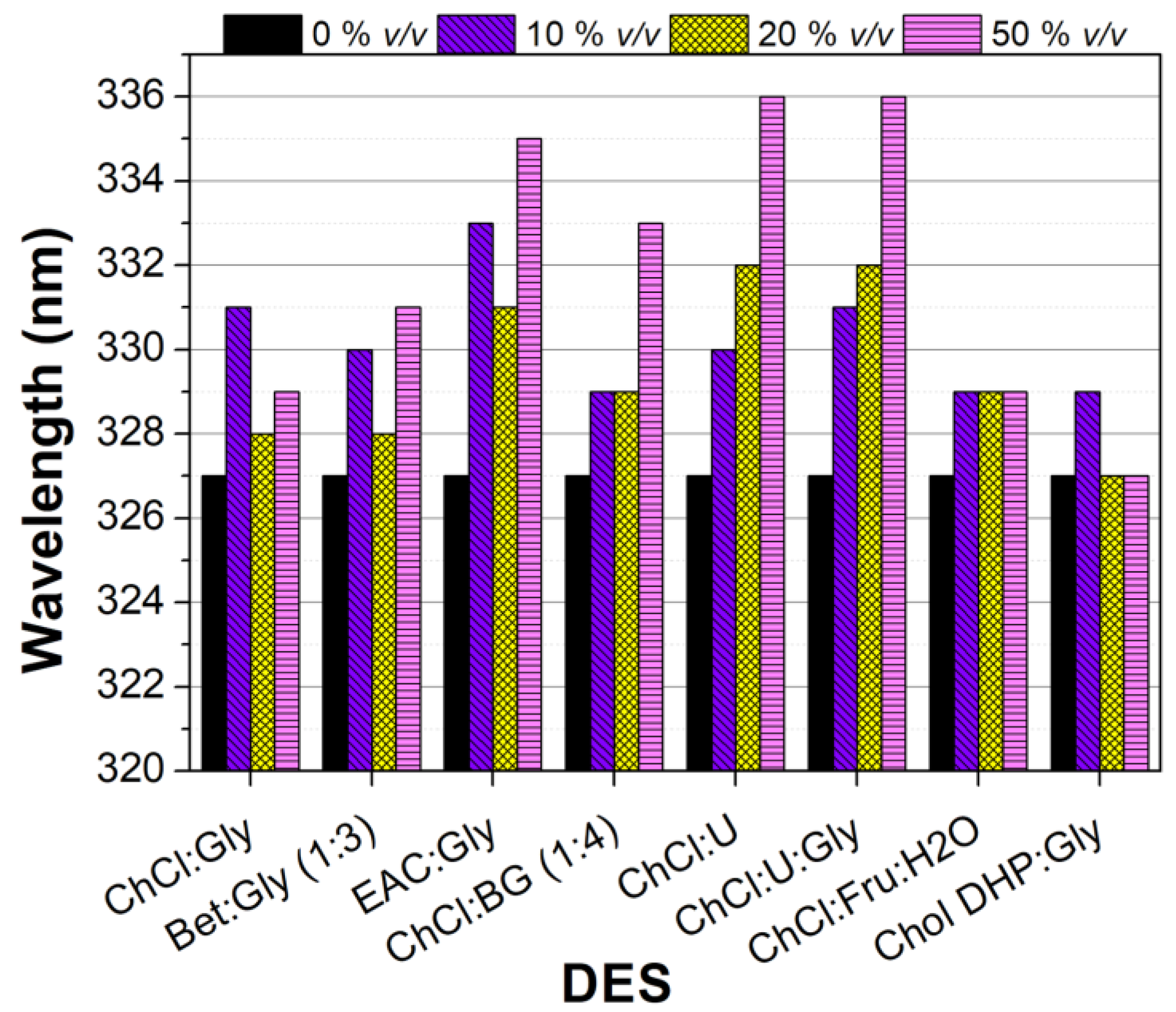
3.3.1. Fluorescence of Free Tyrosinase in DES-Buffer Solutions
3.3.2. Circular Dichroism of Tyrosinase in the Presence of DESs
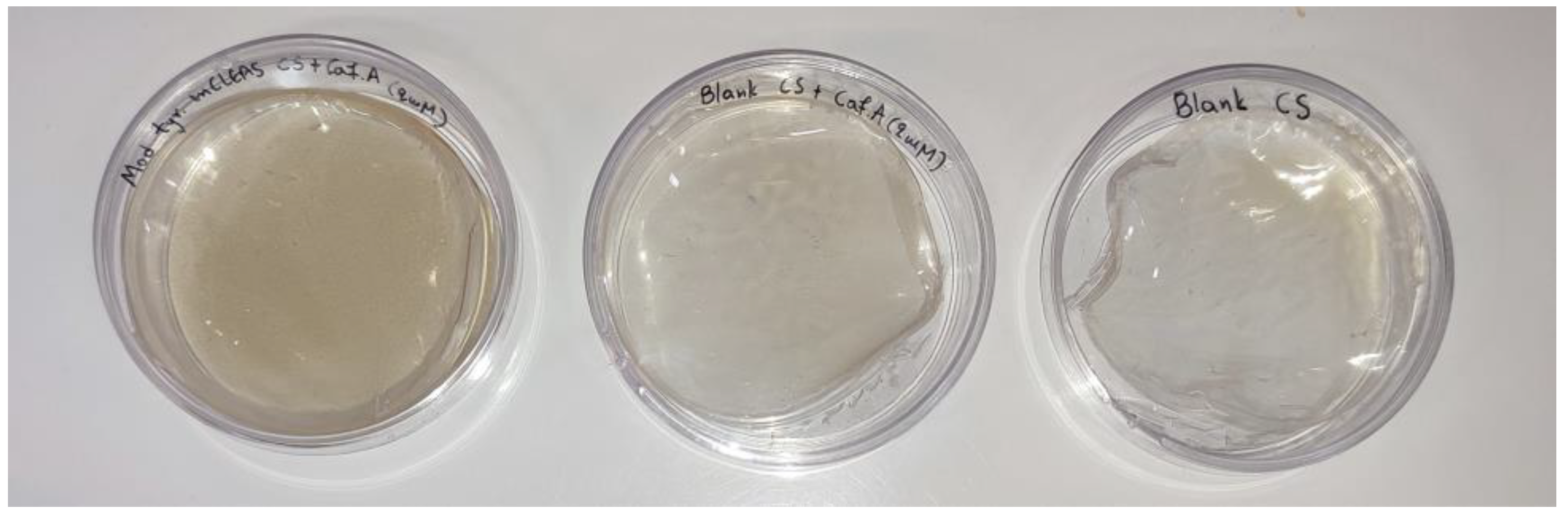
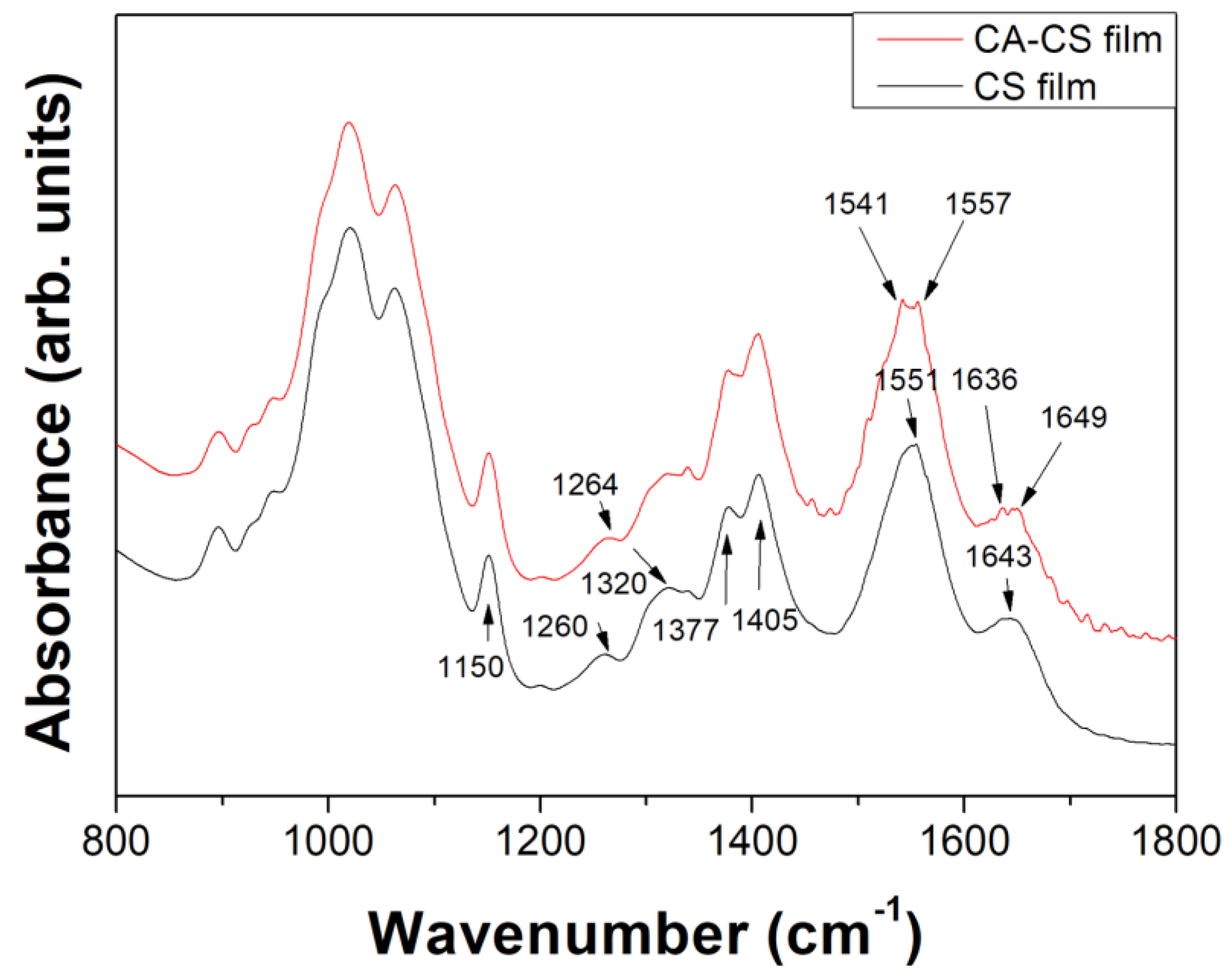
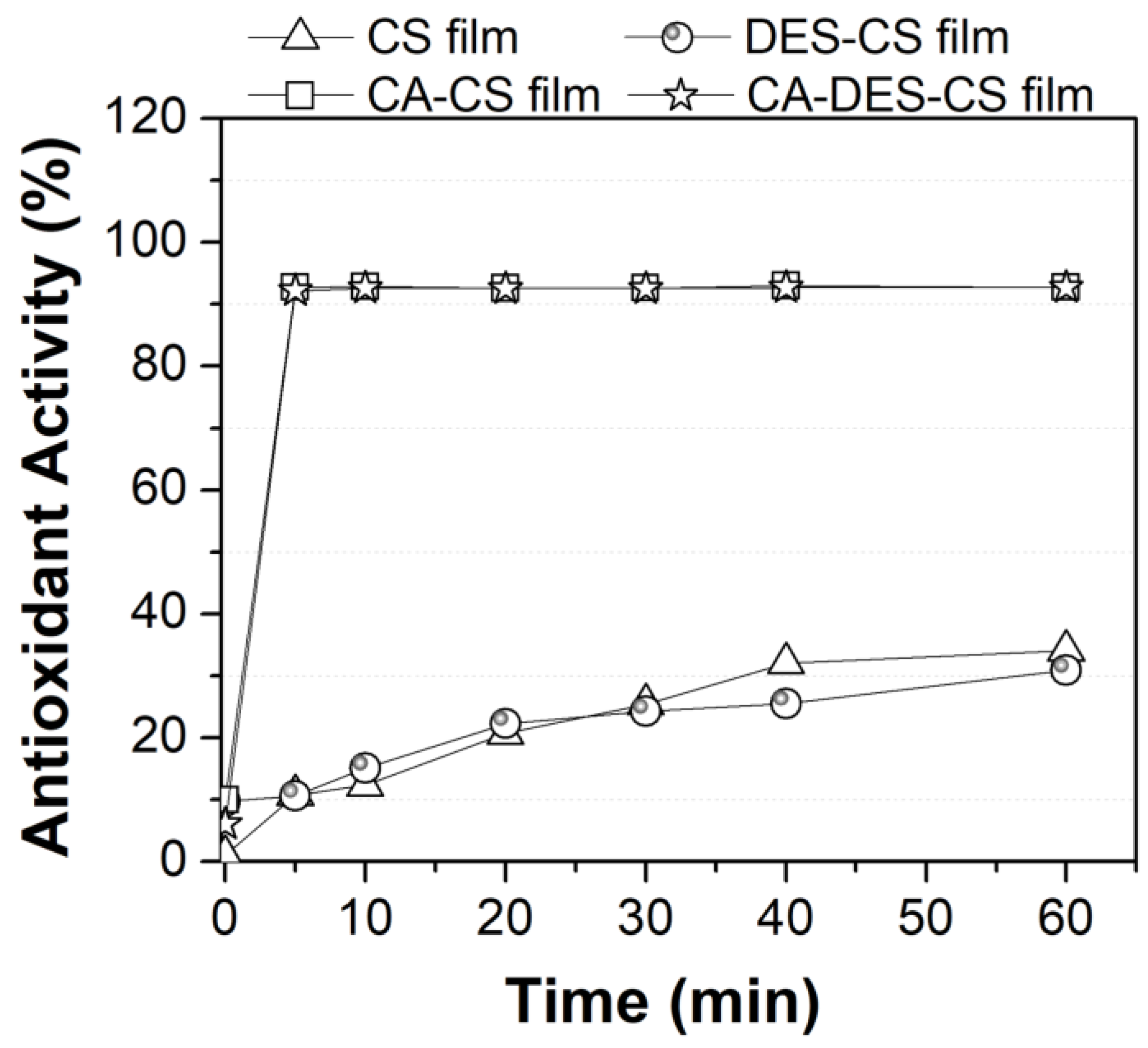
3.4. Functionalization of Chitosan with Caffeic Acid in the Presence of DES
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sheldon, R.A.; Brady, D.; Bode, M.L. The Hitchhiker’s Guide to Biocatalysis: Recent Advances in the Use of Enzymes in Organic Synthesis. Chem. Sci. 2020, 11, 2587–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.-M.; Chen, J.; Shi, Y.-P. Advances on Methods and Easy Separated Support Materials for Enzymes Immobilization. TrAC Trends Anal. Chem. 2018, 102, 332–342. [Google Scholar] [CrossRef]
- Gkantzou, E.; Chatzikonstantinou, A.V.; Fotiadou, R.; Giannakopoulou, A.; Patila, M.; Stamatis, H. Trends in the Development of Innovative Nanobiocatalysts and Their Application in Biocatalytic Transformations. Biotechnol. Adv. 2021, 51, 107738. [Google Scholar] [CrossRef]
- De María, P.D.; Hollmann, F. On the (Un)Greenness of Biocatalysis: Some Challenging Figures and Some Promising Options. Front. Microbiol. 2015, 6, 6–10. [Google Scholar] [CrossRef] [Green Version]
- De María, P.D.; Guajardo, N.; Kara, S. Enzyme Catalysis: In DES, with DES, and in the Presence of DES. In Deep Eutectic Solvents: Synthesis, Properties, and Applications; Wiley: Hoboken, NJ, USA, 2019; pp. 257–271. ISBN 9783527818471. [Google Scholar]
- De María, P.D. Biocatalysis, Sustainability, and Industrial Applications: Show Me the Metrics. Curr. Opin. Green Sustain. Chem. 2021, 31, 100514. [Google Scholar] [CrossRef]
- Sekharan, T.R.; Chandira, R.M.; Tamilvanan, S.; Rajesh, S.C.; Venkateswarlu, B.S. Deep Eutectic Solvents as an Alternate to Other Harmful Solvents. Biointerface Res. Appl. Chem. 2021, 12, 847–860. [Google Scholar] [CrossRef]
- Hessel, V.; Tran, N.N.; Asrami, M.R.; Tran, Q.D.; Van Duc Long, N.; Escribà-Gelonch, M.; Tejada, J.O.; Linke, S.; Sundmacher, K. Sustainability of Green Solvents—Review and Perspective. Green Chem. 2022, 24, 410–437. [Google Scholar] [CrossRef]
- Bryant, S.J.; Christofferson, A.J.; Greaves, T.L.; McConville, C.F.; Bryant, G.; Elbourne, A. Bulk and Interfacial Nanostructure and Properties in Deep Eutectic Solvents: Current Perspectives and Future Directions. J. Colloid. Interface Sci. 2022, 608, 2430–2454. [Google Scholar] [CrossRef] [PubMed]
- Pätzold, M.; Siebenhaller, S.; Kara, S.; Liese, A.; Syldatk, C.; Holtmann, D. Deep Eutectic Solvents as Efficient Solvents in Biocatalysis. Trends Biotechnol. 2019, 37, 943–959. [Google Scholar] [CrossRef]
- Huber, V.; Häckl, K.; Touraud, D.; Kunz, W. Natural Deep Eutectic Solvents: From Simple Systems to Complex Colloidal Mixtures. Adv. Bot. Res. 2021, 97, 17–40. [Google Scholar]
- Guajardo, N.; Ahumada, K.; de María, P.D. Immobilized Lipase-CLEA Aggregates Encapsulated in Lentikats® as Robust Biocatalysts for Continuous Processes in Deep Eutectic Solvents. J. Biotechnol. 2020, 310, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Panić, M.; Cvjetko Bubalo, M.; Radojčić Redovniković, I. Designing a Biocatalytic Process Involving Deep Eutectic Solvents. J. Chem. Technol. Biotechnol. 2021, 96, 14–30. [Google Scholar] [CrossRef]
- Guajardo, N.; De María, P.D.; Canales, R. Integrating Biocatalysis with Viscous Deep Eutectic Solvents in Lab-On-A-Chip Microreactors. ChemSusChem 2022, 15, e202102674. [Google Scholar] [CrossRef] [PubMed]
- Bellou, M.G.; Gkantzou, E.; Skonta, A.; Moschovas, D.; Spyrou, K.; Avgeropoulos, A.; Gournis, D.; Stamatis, H. Development of 3D Printed Enzymatic Microreactors for Lipase-Catalyzed Reactions in Deep Eutectic Solvent-Based Media. Micromachines 2022, 13, 1954. [Google Scholar] [CrossRef]
- Hümmer, M.; Kara, S.; Liese, A.; Huth, I.; Schrader, J.; Holtmann, D. Synthesis of (-)-Menthol Fatty Acid Esters in and from (-)-Menthol and Fatty Acids—Novel Concept for Lipase Catalyzed Esterification Based on Eutectic Solvents. Mol. Catal. 2018, 458, 67–72. [Google Scholar] [CrossRef]
- Guajardo, N.; Ahumada, K.; De María, P.D. Immobilization of Pseudomonas Stutzeri Lipase through Cross-Linking Aggregates (CLEA) for Reactions in Deep Eutectic Solvents. J. Biotechnol. 2021, 337, 18–23. [Google Scholar] [CrossRef]
- Guajardo, N.; Ahumada, K.; De María, P.D.; Schrebler, R.A. Remarkable Stability of Candida Antarctica Lipase B Immobilized via Cross-Linking Aggregates (CLEA) in Deep Eutectic Solvents. Biocatal. Biotransformation 2019, 37, 106–114. [Google Scholar] [CrossRef]
- Guajardo, N.; De María, P.D. Continuous Biocatalysis in Environmentally-Friendly Media: A Triple Synergy for Future Sustainable Processes. ChemCatChem 2019, 11, 3128–3137. [Google Scholar] [CrossRef]
- Imam, H.T.; Krasňan, V.; Rebroš, M.; Marr, A.C. Applications of Ionic Liquids in Whole-Cell and Isolated Enzyme Biocatalysis. Molecules 2021, 26, 4791. [Google Scholar] [CrossRef]
- Erol, Ö.; Hollmann, F. Natural Deep Eutectic Solvents as Performance Additives for Biocatalysis. Adv. Bot. Res. 2021, 97, 95–132. [Google Scholar]
- Xu, P.; Zheng, G.-W.; Zong, M.-H.; Li, N.; Lou, W.-Y. Recent Progress on Deep Eutectic Solvents in Biocatalysis. Bioresour. Bioprocess. 2017, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, P.; Singh, M.; Singh, J.; Singh, R.P. Microbial Tyrosinases: A Novel Enzyme, Structural Features, and Applications. In Applied Microbiology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 3–19. [Google Scholar]
- Aljawish, A.; Chevalot, I.; Jasniewski, J.; Scher, J.; Muniglia, L. Enzymatic Synthesis of Chitosan Derivatives and Their Potential Applications. J. Mol. Catal. B Enzym. 2015, 112, 25–39. [Google Scholar] [CrossRef]
- Liu, D.-M.; Chen, J.; Shi, Y.-P. Tyrosinase Immobilization on Aminated Magnetic Nanoparticles by Physical Adsorption Combined with Covalent Crosslinking with Improved Catalytic Activity, Reusability and Storage Stability. Anal. Chim. Acta 2018, 1006, 90–98. [Google Scholar] [CrossRef]
- Wei, C.; Feng, C.; Li, S.; Zou, Y.; Yang, Z. Mushroom Tyrosinase Immobilized in Metal–Organic Frameworks as an Excellent Catalyst for Both Catecholic Product Synthesis and Phenolic Wastewater Treatment. J. Chem. Technol. Biotechnol. 2022, 97, 962–972. [Google Scholar] [CrossRef]
- Wu, L.; Rathi, B.; Chen, Y.; Wu, X.; Liu, H.; Li, J.; Ming, A.; Han, G. Characterization of Immobilized Tyrosinase—An Enzyme That Is Stable in Organic Solvent at 100 °C. RSC Adv. 2018, 8, 39529–39535. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.X.; Wei, C.M.; Li, S.; Zou, Y.; Yang, Z. Novel Biocatalyst for Efficient Synthesis of Catecholic Products. ACS Sustain. Chem. Eng. 2020, 8, 12277–12285. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, Z.; Duan, Y.; Zhu, Y.; Ou, M.; Xu, X. Immobilization of Tyrosinase on Polyacrylonitrile Beads: Biodegradation of Phenol from Aqueous Solution and the Relevant Cytotoxicity Assessment. RSC Adv. 2017, 7, 28114–28123. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Kiyota, Y.; Miyazaki, M. Techniques for Preparation of Cross-Linked Enzyme Aggregates and Their Applications in Bioconversions. Catalysts 2018, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Doraiswamy, N.; Sarathi, M.; Pennathur, G. Improvement in Biochemical Characteristics of Cross-Linked Enzyme Aggregates (CLEAs) with Magnetic Nanoparticles as Support Matrix. In Nanoarmoring of Enzymes with Carbon Nanotubes and Magnetic Nanoparticles; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 630, pp. 133–158. [Google Scholar]
- Noori, R.; Perwez, M.; Mazumder, J.A.; Ali, J.; Sardar, M. Bio-Imprinted Magnetic Cross-Linked Polyphenol Oxidase Aggregates for Enhanced Synthesis of L-Dopa, a Neurodegenerative Therapeutic Drug. Int. J. Biol. Macromol. 2023, 227, 974–985. [Google Scholar] [CrossRef]
- Aytar, B.S.; Bakir, U. Preparation of Cross-Linked Tyrosinase Aggregates. Process Biochem. 2008, 43, 125–131. [Google Scholar] [CrossRef]
- Xu, D.-Y.; Yang, Z. Cross-Linked Tyrosinase Aggregates for Elimination of Phenolic Compounds from Wastewater. Chemosphere 2013, 92, 391–398. [Google Scholar] [CrossRef]
- Xu, D.-Y.; Chen, J.-Y.; Yang, Z. Use of Cross-Linked Tyrosinase Aggregates as Catalyst for Synthesis of l-DOPA. Biochem. Eng. J. 2012, 63, 88–94. [Google Scholar] [CrossRef]
- Yang, Z.; Yue, Y.-J.; Huang, W.-C.; Zhuang, X.-M.; Chen, Z.-T.; Xing, M. Importance of the Ionic Nature of Ionic Liquids in Affecting Enzyme Performance. J. Biochem. 2009, 145, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-Q.; Li, Z.; Lü, Y.-H.; Yang, Z. Specific Ion Effects of Ionic Liquids on Enzyme Activity and Stability. Green Chem. 2011, 13, 1860. [Google Scholar] [CrossRef]
- Xu, D.-Y.; Yang, Y.; Yang, Z. Activity and Stability of Cross-Linked Tyrosinase Aggregates in Aqueous and Nonaqueous Media. J. Biotechnol. 2011, 152, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Goldfeder, M.; Egozy, M.; Shuster Ben-Yosef, V.; Adir, N.; Fishman, A. Changes in Tyrosinase Specificity by Ionic Liquids and Sodium Dodecyl Sulfate. Appl. Microbiol. Biotechnol. 2013, 97, 1953–1961. [Google Scholar] [CrossRef]
- Heitz, M.P.; Rupp, J.W.; Horn, K.W. Biocatalytic Activity of Mushroom Tyrosinase in Ionic Liquids: Specific Ion Effects and the Hofmeister Series. Insights Enzym. Res. 2018, 2, 1–9. [Google Scholar] [CrossRef]
- Cheng, H.; Zou, Y.; Luo, X.; Song, X.-H.; Yang, Z. Enzymatic Synthesis of Catechol-Functionalized Polyphenols with Excellent Selectivity and Productivity. Process. Biochem. 2018, 70, 90–97. [Google Scholar] [CrossRef]
- Xie, W.; Xu, P.; Liu, Q. Antioxidant Activity of Water-Soluble Chitosan Derivatives. Bioorg. Med. Chem. Lett. 2001, 11, 1699–1701. [Google Scholar] [CrossRef]
- Mendoza, S.M.; Arfaoui, I.; Zanarini, S.; Paolucci, F.; Rudolf, P. Improvements in the Characterization of the Crystalline Structure of Acid-Terminated Alkanethiol Self-Assembled Monolayers on Au(111). Langmuir 2007, 23, 582–588. [Google Scholar] [CrossRef] [Green Version]
- Delorme, A.E.; Andanson, J.-M.; Verney, V. Improving Laccase Thermostability with Aqueous Natural Deep Eutectic Solvents. Int. J. Biol. Macromol. 2020, 163, 919–926. [Google Scholar] [CrossRef]
- Winder, A.J.; Harris, H. New Assays for the Tyrosine Hydroxylase and Dopa Oxidase Activities of Tyrosinase. Eur. J. Biochem. 1991, 198, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.J.; Ramalli, S.G.; Wallace, B.A. DichroWeb, a Website for Calculating Protein Secondary Structure from Circular Dichroism Spectroscopic Data. Protein. Sci. 2022, 31, 37–46. [Google Scholar] [CrossRef]
- Božič, M.; Gorgieva, S.; Kokol, V. Laccase-Mediated Functionalization of Chitosan by Caffeic and Gallic Acids for Modulating Antioxidant and Antimicrobial Properties. Carbohydr. Polym. 2012, 87, 2388–2398. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, M.; Li, W.; Wang, L.; Zheng, J.; Gan, W.; Xu, J. Large-Scale Fabrication and Application of Magnetite Coated Ag NW-Core Water-Dispersible Hybrid Nanomaterials. Dalton. Trans. 2015, 44, 7803–7810. [Google Scholar] [CrossRef] [PubMed]
- Malega, F.; Indrayana, I.P.T.; Suharyadi, E. Synthesis and Characterization of the Microstructure and Functional Group Bond of Fe3O4 Nanoparticles from Natural Iron Sand in Tobelo North Halmahera. J. Ilm. Pendidik. Fis. Al-Biruni 2018, 7, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Nandiyanto, A.B.D.; Oktiani, R.; Ragadhita, R. How to Read and Interpret FTIR Spectroscope of Organic Material. Indones. J. Sci. Technol. 2019, 4, 97. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Zhang, W.; Yang, Z.; Yang, X.; Wang, N.; Yu, X. Novel Magnetic Cross-Linked Cellulase Aggregates with a Potential Application in Lignocellulosic Biomass Bioconversion. Molecules 2017, 22, 269. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, Z.; Han, S.; Li, C.; Lei, B.; Stewart, M.P.; Tour, J.M.; Zhou, C. Magnetite (Fe3O4) Core−Shell Nanowires: Synthesis and Magnetoresistance. Nano Lett. 2004, 4, 2151–2155. [Google Scholar] [CrossRef]
- Daou, T.J.; Pourroy, G.; Bégin-Colin, S.; Grenèche, J.M.; Ulhaq-Bouillet, C.; Legaré, P.; Bernhardt, P.; Leuvrey, C.; Rogez, G. Hydrothermal Synthesis of Monodisperse Magnetite Nanoparticles. Chem. Mater. 2006, 18, 4399–4404. [Google Scholar] [CrossRef]
- Radu, T.; Iacovita, C.; Benea, D.; Turcu, R. X-Ray Photoelectron Spectroscopic Characterization of Iron Oxide Nanoparticles. Appl. Surf. Sci. 2017, 405, 337–343. [Google Scholar] [CrossRef]
- Khodaverdian, S.; Dabirmanesh, B.; Heydari, A.; Dashtban-moghadam, E.; Khajeh, K.; Ghazi, F. Activity, Stability and Structure of Laccase in Betaine Based Natural Deep Eutectic Solvents. Int. J. Biol. Macromol. 2018, 107, 2574–2579. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-D.; Kim, S.; Lyou, Y.-J.; Lee, J.-Y.; Yang, J.-M. A New Type of Uncompetitive Inhibition of Tyrosinase Induced by Cl− Binding. Biochimie 2005, 87, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Toledo, M.L.; Pereira, M.M.; Freire, M.G.; Silva, J.P.A.; Coutinho, J.A.P.; Tavares, A.P.M. Laccase Activation in Deep Eutectic Solvents. ACS Sustain. Chem. Eng. 2019, 7, 11806–11814. [Google Scholar] [CrossRef]
- Zhang, Q.; De Oliveira Vigier, K.; Royer, S.; Jérôme, F. Deep Eutectic Solvents: Syntheses, Properties and Applications. Chem. Soc. Rev. 2012, 41, 7108. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.A.; Efstathiadou, E.; Patila, M.; Polydera, A.C.; Stamatis, H. Deep Eutectic Solvents as Media for Peroxidation Reactions Catalyzed by Heme-Dependent Biocatalysts. Ind. Eng. Chem. Res. 2016, 55, 5145–5151. [Google Scholar] [CrossRef]
- Chan, J.C.; Zhang, B.; Martinez, M.; Kuruba, B.; Brozik, J.; Kang, C.; Zhang, X. Structural Studies of Myceliophthora Thermophila Laccase in the Presence of Deep Eutectic Solvents. Enzym. Microb. Technol. 2021, 150, 109890. [Google Scholar] [CrossRef]
- Sun, H.; Xin, R.; Qu, D.; Yao, F. Mechanism of Deep Eutectic Solvents Enhancing Catalytic Function of Cytochrome P450 Enzymes in Biosynthesis and Organic Synthesis. J. Biotechnol. 2020, 323, 264–273. [Google Scholar] [CrossRef]
- de Gonzalo, G.; Martin, C.; Fraaije, M.W. Positive Impact of Natural Deep Eutectic Solvents on the Biocatalytic Performance of 5-Hydroxymethyl-Furfural Oxidase. Catalysts 2020, 10, 447. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.-J.; Huang, Y.-K.; Li, F.; Wang, D.-D.; Yin, M.-N.; Wang, M.; Xia, Z.-N. Improving β-Glucosidase Biocatalysis with Deep Eutectic Solvents Based on Choline Chloride. Biochem. Eng. J. 2018, 138, 37–46. [Google Scholar] [CrossRef]
- Fotiadou, R.; Bellou, M.G.; Spyrou, K.; Yan, F.; Rudolf, P.; Gournis, D.; Stamatis, H. Effect of Deep Eutectic Solvents on the Biocatalytic Properties of β-Glucosidase@ZnOFe Nano-Biocatalyst. Sustain. Chem. Pharm. 2022, 30, 100886. [Google Scholar] [CrossRef]
- Deshayes, S.; Divita, G. Fluorescence Technologies for Monitoring Interactions Between Biological Molecules In Vitro. Prog. Mol. Biol. Transl. Sci. 2013, 113, 109–143. [Google Scholar] [PubMed]
- Song, X.; Hu, X.; Zhang, Y.; Pan, J.; Gong, D.; Zhang, G. Inhibitory Mechanism of Epicatechin Gallate on Tyrosinase: Inhibitory Interaction, Conformational Change and Computational Simulation. Food Funct. 2020, 11, 4892–4902. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Ma, J.; Ahmed, I.; Lv, L.; Li, Z.; Lin, H. Effect of Tyrosinase-Catalyzed Crosslinking on the Structure and Allergenicity of Turbot Parvalbumin Mediated by Caffeic Acid. J. Sci. Food Agric. 2019, 99, 3501–3508. [Google Scholar] [CrossRef]
- Hridya, H.; Amrita, A.; Sankari, M.; George Priya Doss, C.; Gopalakrishnan, M.; Gopalakrishnan, C.; Siva, R. Inhibitory Effect of Brazilein on Tyrosinase and Melanin Synthesis: Kinetics and in Silico Approach. Int. J. Biol. Macromol. 2015, 81, 228–234. [Google Scholar] [CrossRef]
- Biswas, R.; Chanda, J.; Kar, A.; Mukherjee, P.K. Tyrosinase Inhibitory Mechanism of Betulinic Acid from Dillenia Indica. Food Chem. 2017, 232, 689–696. [Google Scholar] [CrossRef]
- Hridya, H.; Amrita, A.; Mohan, S.; Gopalakrishnan, M.; Kumar, D.T.; Doss, G.P.; Siva, R. Functionality Study of Santalin as Tyrosinase Inhibitor: A Potential Depigmentation Agent. Int. J. Biol. Macromol. 2016, 86, 383–389. [Google Scholar] [CrossRef]
- Galvis-Sánchez, A.C.; Castro, M.C.R.; Biernacki, K.; Gonçalves, M.P.; Souza, H.K.S. Natural Deep Eutectic Solvents as Green Plasticizers for Chitosan Thermoplastic Production with Controlled/Desired Mechanical and Barrier Properties. Food Hydrocoll. 2018, 82, 478–489. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, J.; Gao, P.; Jiang, Q.; Xia, W. Sustainable Chitosan Films Containing a Betaine-Based Deep Eutectic Solvent and Lignin: Physicochemical, Antioxidant, and Antimicrobial Properties. Food Hydrocoll. 2022, 129, 107656. [Google Scholar] [CrossRef]
- Jakubowska, E.; Gierszewska, M.; Nowaczyk, J.; Olewnik-Kruszkowska, E. The Role of a Deep Eutectic Solvent in Changes of Physicochemical and Antioxidative Properties of Chitosan-Based Films. Carbohydr. Polym. 2021, 255, 117527. [Google Scholar] [CrossRef]
- Jakubowska, E.; Gierszewska, M.; Nowaczyk, J.; Olewnik-Kruszkowska, E. Physicochemical and Storage Properties of Chitosan-Based Films Plasticized with Deep Eutectic Solvent. Food Hydrocoll. 2020, 108, 106007. [Google Scholar] [CrossRef]
- Sokolova, M.P.; Smirnov, M.A.; Samarov, A.A.; Bobrova, N.V.; Vorobiov, V.K.; Popova, E.N.; Filippova, E.; Geydt, P.; Lahderanta, E.; Toikka, A.M. Plasticizing of Chitosan Films with Deep Eutectic Mixture of Malonic Acid and Choline Chloride. Carbohydr. Polym. 2018, 197, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, M.A.; Nikolaeva, A.L.; Vorobiov, V.K.; Bobrova, N.V.; Abalov, I.V.; Smirnov, A.V.; Sokolova, M.P. Ionic Conductivity and Structure of Chitosan Films Modified with Lactic Acid-Choline Chloride NADES. Polymers 2020, 12, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, B.; Javvaji, V.; Kim, E.; Lee, M.E.; Raghavan, S.R.; Wang, Q.; Payne, G.F. Tyrosinase-Mediated Grafting and Crosslinking of Natural Phenols Confers Functional Properties to Chitosan. Biochem. Eng. J. 2014, 89, 21–27. [Google Scholar] [CrossRef]
- Aljawish, A.; Muniglia, L.; Klouj, A.; Jasniewski, J.; Scher, J.; Desobry, S. Characterization of Films Based on Enzymatically Modified Chitosan Derivatives with Phenol Compounds. Food Hydrocoll. 2016, 60, 551–558. [Google Scholar] [CrossRef]
- Ferreira, A.S.; Nunes, C.; Castro, A.; Ferreira, P.; Coimbra, M.A. Influence of Grape Pomace Extract Incorporation on Chitosan Films Properties. Carbohydr. Polym. 2014, 113, 490–499. [Google Scholar] [CrossRef]
- Zeng, J.; Ren, X.; Zhu, S.; Gao, Y. Fabrication and Characterization of an Economical Active Packaging Film Based on Chitosan Incorporated with Pomegranate Peel. Int. J. Biol. Macromol. 2021, 192, 1160–1168. [Google Scholar] [CrossRef]
- Mohammed-Ziegler, I.; Billes, F. Vibrational Spectroscopic Calculations on Pyrogallol and Gallic Acid. J. Mol. Struct. THEOCHEM 2002, 618, 259–265. [Google Scholar] [CrossRef]
- Sampaio, S.; Taddei, P.; Monti, P.; Buchert, J.; Freddi, G. Enzymatic Grafting of Chitosan onto Bombyx Mori Silk Fibroin: Kinetic and IR Vibrational Studies. J. Biotechnol. 2005, 116, 21–33. [Google Scholar] [CrossRef]
- Elderderi, S.; Leman-Loubière, C.; Wils, L.; Henry, S.; Bertrand, D.; Byrne, H.J.; Chourpa, I.; Enguehard-Gueiffier, C.; Munnier, E.; Elbashir, A.A.; et al. ATR-IR Spectroscopy for Rapid Quantification of Water Content in Deep Eutectic Solvents. J. Mol. Liq. 2020, 311, 113361. [Google Scholar] [CrossRef]
- Wang, L.; Dong, Y.; Men, H.; Tong, J.; Zhou, J. Preparation and Characterization of Active Films Based on Chitosan Incorporated Tea Polyphenols. Food Hydrocoll. 2013, 32, 35–41. [Google Scholar] [CrossRef]
- Zhou, X.; Iqbal, A.; Li, J.; Liu, C.; Murtaza, A.; Xu, X.; Pan, S.; Hu, W. Changes in Browning Degree and Reducibility of Polyphenols during Autoxidation and Enzymatic Oxidation. Antioxidants 2021, 10, 1809. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DES | Abbreviation | Molar Ratio |
|---|---|---|
| Choline Chloride:Urea | ChCl:U | 1:2 |
| Choline Chloride:Glycerol | ChCl:Gly | 1:2 |
| Choline Chloride:Ethylene Glycol | ChCl:EG | 1:2 |
| Choline Chloride:Propylene Glycol | ChCl:PG | 1:2 |
| Choline Chloride:Butylene Glycol | ChCl:BG | 1:2 |
| Choline Chloride:Butylene Glycol | ChCl:BG | 1:4 |
| Choline Chloride:Fructose:H2O | ChCl:Fru:H2O | 5:2:5 |
| Choline Chloride: Urea:Glycerol | ChCl:U:Gly | 1:1:1 |
| Choline Chloride:Urea:Ethylene Glycol | ChCl:U:EG | 1:1:1 |
| Choline Chloride:Glycerol:Ethylene Glycol | ChCl:Gly:EG | 1:1:1 |
| Ethylammonium Chloride:Urea | EAC:U | 1:1.5 |
| Ethylammonium Chloride:Glycerol | EAC:Gly | 1:1.5 |
| Ethylammonium Chloride:Ethylene Glycol | EAC:EG | 1:1.5 |
| Betaine:Glycerol | Bet:Gly | 1:2 |
| Betaine:Glycerol | Bet:Gly | 1:3 |
| Betaine:Ethylene Glycol | Bet:EG | 1:3 |
| Choline Dihydrogen Phosphate:Glycerol | Chol DHP:Gly | 1:3 |
| Choline Dihydrogen Phosphate:Ethylene Glycol | Chol DHP:EG | 1:2 |
| DES | Kmapp [mM] | Vmaxapp [μΜ min−1] |
|---|---|---|
| No DES | 9.7 ± 1.4 | 26.0 ± 1.6 |
| ChCl:Gly | 12.5 ± 1.9 | 20.1 ± 1.5 |
| EAC:Gly | 11.0 ± 1.6 | 13.6 ± 0.9 |
| Bet:Gly (1:3) | 9.0 ± 2.0 | 15.5 ± 1.4 |
| Chol DHP:Gly | 27.6 ± 5.2 | 31.9 ± 3.9 |
| DES | Kmapp [mM] | Vmaxapp [μΜ min−1] |
|---|---|---|
| No DES | 9.7 ± 1.4 | 26.0 ± 1.6 |
| ChCl:Gly | 12.5 ± 1.9 | 20.1 ± 1.5 |
| ChCl:U | 15.2 ± 2.7 | 16.4 ± 1.5 |
| ChCl:U:Gly | 12.1 ± 1.8 | 15.0 ± 1.1 |
| ChCl:BG (1:4) | 22.2 ± 4.2 | 14.2 ± 1.6 |
| ChCl:Fru:H2O | 9.9 ± 0.9 | 13.0 ± 0.5 |
| DES Concentration | α-Helix (%) | β-Sheet (%) | Random Coil (%) |
|---|---|---|---|
| DES-free | 31 | 16 | 54 |
| 10% v/v Chol DHP:EG | 31 | 12 | 57 |
| 10% v/v ChCl:Fru:H2O | 29 | 13 | 57 |
| 10% v/v ChCl:Gly:EG | 28 | 15 | 56 |
| 10% v/v ChCl:EG | 26 | 18 | 56 |
| 10% v/v ChCl:Gly | 26 | 20 | 54 |
| 10% v/v ChCl:U | 25 | 19 | 56 |
| 10% v/v ChCl:PG | 24 | 25 | 51 |
| 10% v/v EAC:EG | 22 | 23 | 55 |
| 10% v/v ChCl:U:Gly | 21 | 26 | 53 |
| DES Concentration | α-Helix (%) | β-Sheet (%) | Random Coil (%) |
|---|---|---|---|
| DES-free | 17 | 31 | 52 |
| 10% v/v ChCl:PG | 9 | 43 | 48 |
| 10% v/v ChCl:EG | 9 | 43 | 48 |
| 10% v/v ChCl:U:EG | 8 | 44 | 48 |
| 10% v/v EAC:EG | 5 | 48 | 48 |
| 10% v/v ChCl:Fru:H2O | 4 | 48 | 48 |
| 10% v/v Chol DHP:EG | 4 | 48 | 48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellou, M.G.; Patila, M.; Fotiadou, R.; Spyrou, K.; Yan, F.; Rudolf, P.; Gournis, D.P.; Stamatis, H. Tyrosinase Magnetic Cross-Linked Enzyme Aggregates: Biocatalytic Study in Deep Eutectic Solvent Aqueous Solutions. Biomolecules 2023, 13, 643. https://doi.org/10.3390/biom13040643
Bellou MG, Patila M, Fotiadou R, Spyrou K, Yan F, Rudolf P, Gournis DP, Stamatis H. Tyrosinase Magnetic Cross-Linked Enzyme Aggregates: Biocatalytic Study in Deep Eutectic Solvent Aqueous Solutions. Biomolecules. 2023; 13(4):643. https://doi.org/10.3390/biom13040643
Chicago/Turabian StyleBellou, Myrto G., Michaela Patila, Renia Fotiadou, Konstantinos Spyrou, Feng Yan, Petra Rudolf, Dimitrios P. Gournis, and Haralambos Stamatis. 2023. "Tyrosinase Magnetic Cross-Linked Enzyme Aggregates: Biocatalytic Study in Deep Eutectic Solvent Aqueous Solutions" Biomolecules 13, no. 4: 643. https://doi.org/10.3390/biom13040643