
HSP70 Family in Cancer: Signaling Mechanisms and Therapeutic Advances

Abstract
:1. Introduction
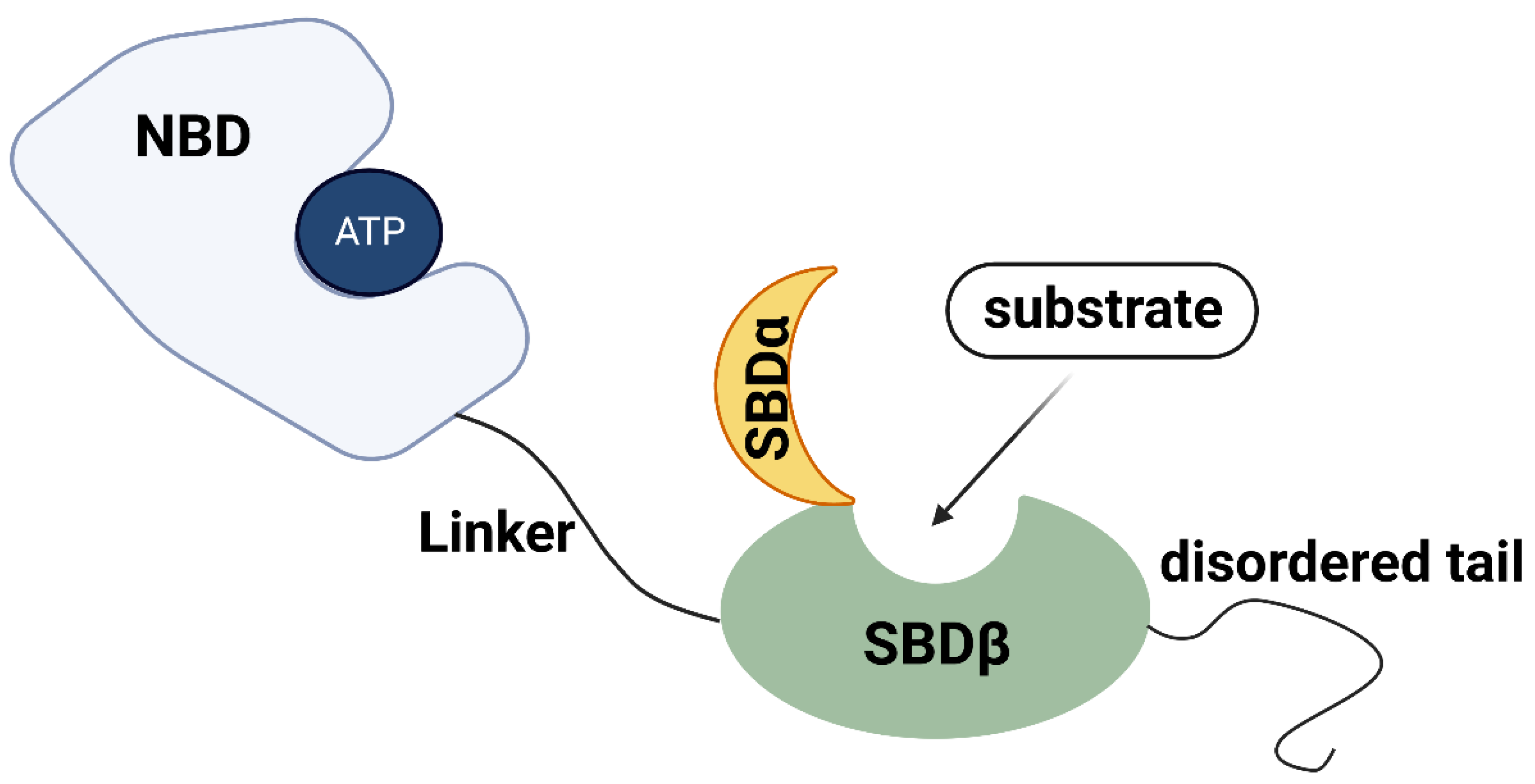
2. HSP70 Family: Family Members, Common Structure, and Basic Function
3. HSP70 Regulates Multiple Cancer Related Signaling Pathways
3.1. RTKs-RAS-RAF-MEK-ERK Pathway
3.2. PI3K/AKT/mTOR Pathway
3.3. Effect of HSP70 on Key Proteins of Other Signaling Pathways
4. Targeting HSP70 in Cancer Therapy
4.1. HSP70 as an Inhibitory Target in Cancer Therapies
4.1.1. HSP70 Inhibitors in Monotherapy Mode
NBD-Binding Inhibitors
SBD-Binding Inhibitors
C-Terminal EEVD-Binding Inhibitors
Other Inhibitors
4.1.2. HSP70 Inhibitors in Combination Therapy Mode
4.2. HSP70 as an Adjuvant in Cancer Vaccine Therapies
4.3. HSP70-Based Cancer Therapies in Clinical Trials
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in drosophila. Cell. Mol. Life Sci. 1962, 18, 57–573. [Google Scholar] [CrossRef]
- Jäättelä, M.; Saksela, K.; Saksela, E. Heat shock protects WEHI-164 target cells from the cytolysis by tumor necrosis factors alpha and beta. Eur. J. Immunol. 1989, 19, 1413–1417. [Google Scholar] [CrossRef] [PubMed]
- Petersen, N.S.; Mitchell, H.K. Recovery of protein synthesis after heat shock: Prior heat treatment affects the ability of cells to translate mRNA. Proc. Natl. Acad. Sci. USA 1981, 78, 1708–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapareto, S.A.; Hopwood, L.E.; Dewey, W.C.; Raju, M.R.; Gray, J.W. Effects of hyperthermia on survival and progression of Chinese hamster ovary cells. Cancer Res. 1978, 38, 393–400. [Google Scholar]
- Henle, K.J.; Karamuz, J.E.; Leeper, D.B. Induction of thermotolerance in Chinese hamster ovary cells by high (45 degrees) or low (40 degrees) hyperthermia. Cancer Res. 1978, 38, 570–574. [Google Scholar]
- Gerner, E.W.; Boone, R.; Connor, W.G.; Hicks, J.A.; Boone, M.L. A transient thermotolerant survival response produced by single thermal doses in HeLa cells. Cancer Res. 1976, 36, 1035–1040. [Google Scholar]
- Ritossa, F. Discovery of the heat shock response. Cell Stress Chaperones 1996, 1, 97–98. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Reviews. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Albakova, Z.; Armeev, G.A.; Kanevskiy, L.M.; Kovalenko, E.I.; Sapozhnikov, A.M. HSP70 Multi-Functionality in Cancer. Cells 2020, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, M.Y.; Gabai, V.L. Hsp70 in cancer: Back to the future. Oncogene 2014, 34, 4153–4161. [Google Scholar] [CrossRef] [Green Version]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Khaleque, M.A.; Sawyer, D.B.; Ciocca, D.R. Heat shock proteins in cancer: Chaperones of tumorigenesis. Trends Biochem. Sci. 2006, 31, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Brondani Da Rocha, A.; Regner, A.; Grivicich, I.; Pretto Schunemann, D.; Diel, C.; Kovaleski, G.; Brunetto De Farias, C.; Mondadori, E.; Almeida, L.; Braga Filho, A.; et al. Radioresistance is associated to increased Hsp70 content in human glioblastoma cell lines. Int. J. Oncol. 2004, 25, 777–785. [Google Scholar] [CrossRef]
- Yang, Z.; Zhuang, L.; Szatmary, P.; Wen, L.; Sun, H.; Lu, Y.; Xu, Q.; Chen, X. Upregulation of heat shock proteins (HSPA12A, HSP90B1, HSPA4, HSPA5 and HSPA6) in tumour tissues is associated with poor outcomes from HBV-related early-stage hepatocellular carcinoma. Int. J. Med. Sci. 2015, 12, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Ji, M.; Chen, L.; Liu, Q.; Che, S.; Xu, M.; Lin, Z. The clinicopathological significance of Mortalin overexpression in invasive ductal carcinoma of breast. J. Exp. Clin. Cancer Res. CR 2016, 35, 42. [Google Scholar] [CrossRef] [Green Version]
- Radons, J. The human HSP70 family of chaperones: Where do we stand? Cell Stress Chaperones 2016, 21, 379–404. [Google Scholar] [CrossRef] [Green Version]
- Hunt, C.; Morimoto, R.I. Conserved features of eukaryotic hsp70 genes revealed by comparison with the nucleotide sequence of human hsp70. Proc. Natl. Acad. Sci. USA 1985, 82, 6455–6459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.S.; Singh, B. Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr. Biol. CB 1994, 4, 1104–1114. [Google Scholar] [CrossRef]
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat-shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, A.; Handa, N.; Shirouzu, M.; Yokoyama, S. Biochemical and structural studies on the high affinity of Hsp70 for ADP. Protein Sci. A Publ. Protein Soc. 2011, 20, 1367–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, M.; Mayer, M.P.; Bukau, B. Allosteric regulation of Hsp70 chaperones involves a conserved interdomain linker. J. Biol. Chem. 2006, 281, 38705–38711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhao, X.; Burkholder, W.F.; Gragerov, A.; Ogata, C.M.; Gottesman, M.E.; Hendrickson, W.A. Structural analysis of substrate binding by the molecular chaperone DnaK. Science 1996, 272, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Kityk, R.; Vogel, M.; Schlecht, R.; Bukau, B.; Mayer, M.P. Pathways of allosteric regulation in Hsp70 chaperones. Nat. Commun. 2015, 6, 8308. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, A.; Clerico, E.M.; Gierasch, L.M. An interdomain energetic tug-of-war creates the allosterically active state in Hsp70 molecular chaperones. Cell 2012, 151, 1296–1307. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Schröder, H.; Rüdiger, S.; Paal, K.; Laufen, T.; Bukau, B. Multistep mechanism of substrate binding determines chaperone activity of Hsp70. Nat. Struct. Biol. 2000, 7, 586–593. [Google Scholar] [CrossRef]
- Zuiderweg, E.R.; Hightower, L.E.; Gestwicki, J.E. The remarkable multivalency of the Hsp70 chaperones. Cell Stress Chaperones 2017, 22, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Scheufler, C.; Brinker, A.; Bourenkov, G.; Pegoraro, S.; Moroder, L.; Bartunik, H.; Hartl, F.U.; Moarefi, I. Structure of TPR domain-peptide complexes: Critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 2000, 101, 199–210. [Google Scholar] [CrossRef]
- Odunuga, O.O.; Hornby, J.A.; Bies, C.; Zimmermann, R.; Pugh, D.J.; Blatch, G.L. Tetratricopeptide repeat motif-mediated Hsc70-mSTI1 interaction. Molecular characterization of the critical contacts for successful binding and specificity. J. Biol. Chem. 2003, 278, 6896–6904. [Google Scholar] [CrossRef] [Green Version]
- Laufen, T.; Mayer, M.P.; Beisel, C.; Klostermeier, D.; Mogk, A.; Reinstein, J.; Bukau, B. Mechanism of regulation of hsp70 chaperones by DnaJ cochaperones. Proc. Natl. Acad. Sci. USA 1999, 96, 5452–5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. CMLS 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Evdonin, A.; Kinev, A.; Tsupkina, N.; Guerriero, V.; Raynes, D.A.; Medvedeva, N. Extracellular HspBP1 and Hsp72 synergistically activate epidermal growth factor receptor. Biol. Cell 2009, 101, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yanjiao, G.; Wubin, H.; Yue, W.; Jianhua, H.; Huachuan, Z.; Rongjian, S.; Zhidong, L. Secreted GRP78 activates EGFR-SRC-STAT3 signaling and confers the resistance to sorafeinib in HCC cells. Oncotarget 2017, 8, 19354–19364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Chen, C.; Chen, J.; Zhan, R.; Zhang, Q.; Xu, X.; Li, D.; Li, M. Cell surface GRP78 facilitates hepatoma cells proliferation and migration by activating IGF-IR. Cell. Signal. 2017, 35, 154–162. [Google Scholar] [CrossRef]
- Bonvini, P.; Zorzi, E.; Basso, G.; Rosolen, A. Bortezomib-mediated 26S proteasome inhibition causes cell-cycle arrest and induces apoptosis in CD-30+ anaplastic large cell lymphoma. Leukemia 2007, 21, 838–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonvini, P.; Zorzi, E.; Mussolin, L.; Pillon, M.; Romualdi, C.; Peron, M.; d’Amore, E.S.; Lamant, L.; Rosolen, A. Consequences of heat shock protein 72 (Hsp72) expression and activity on stress-induced apoptosis in CD30+ NPM-ALK+ anaplastic large-cell lymphomas. Leukemia 2012, 26, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Hunt, C.; Yaglom, J.A.; Gabai, V.L.; Sherman, M.Y. Heat shock protein Hsp72 plays an essential role in Her2-induced mammary tumorigenesis. Oncogene 2011, 30, 2836–2845. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Ha, D.P.; Zhu, G.; Rangel, D.F.; Kobielak, A.; Gill, P.S.; Groshen, S.; Dubeau, L.; Lee, A.S. GRP78 haploinsufficiency suppresses acinar-to-ductal metaplasia, signaling, and mutant Kras-driven pancreatic tumorigenesis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, E4020–E4029. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.K.; Hong, S.K.; Starenki, D.; Oshima, K.; Shao, H.; Gestwicki, J.E.; Tsai, S.; Park, J.I. Mortalin/HSPA9 targeting selectively induces KRAS tumor cell death by perturbing mitochondrial membrane permeability. Oncogene 2020, 39, 4257–4270. [Google Scholar] [CrossRef]
- Rangel, D.F.; Dubeau, L.; Park, R.; Chan, P.; Ha, D.P.; Pulido, M.A.; Mullen, D.J.; Vorobyova, I.; Zhou, B.; Borok, Z.; et al. Endoplasmic reticulum chaperone GRP78/BiP is critical for mutant Kras-driven lung tumorigenesis. Oncogene 2021, 40, 3624–3632. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.P.; Huang, B.; Wang, H.; Rangel, D.F.; Van Krieken, R.; Liu, Z.; Samanta, S.; Neamati, N.; Lee, A.S. Targeting GRP78 suppresses oncogenic KRAS protein expression and reduces viability of cancer cells bearing various KRAS mutations. Neoplasia 2022, 33, 100837. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Takeda, M.; Morimoto, R.I. Bag1-Hsp70 mediates a physiological stress signalling pathway that regulates Raf-1/ERK and cell growth. Nat. Cell Biol. 2001, 3, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.K.; Hong, S.K.; Park, J.I. Steady-State Levels of Phosphorylated Mitogen-Activated Protein Kinase Kinase 1/2 Determined by Mortalin/HSPA9 and Protein Phosphatase 1 Alpha in KRAS and BRAF Tumor Cells. Mol. Cell. Biol. 2017, 37, 17–69. [Google Scholar] [CrossRef] [Green Version]
- Kelber, J.A.; Panopoulos, A.D.; Shani, G.; Booker, E.C.; Belmonte, J.C.; Vale, W.W.; Gray, P.C. Blockade of Cripto binding to cell surface GRP78 inhibits oncogenic Cripto signaling via MAPK/PI3K and Smad2/3 pathways. Oncogene 2009, 28, 2324–2336. [Google Scholar] [CrossRef] [Green Version]
- Misra, U.K.; Pizzo, S.V. Ligation of cell surface GRP78 with antibody directed against the COOH-terminal domain of GRP78 suppresses Ras/MAPK and PI 3-kinase/AKT signaling while promoting caspase activation in human prostate cancer cells. Cancer Biol. Ther. 2010, 9, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.C.; Lai, N.S.; Yin, W.Y.; Yu, H.C.; Huang, H.B.; Tung, C.H.; Huang, K.Y.; Yu, C.L. Anti-citrullinated protein antibodies activated ERK1/2 and JNK mitogen-activated protein kinases via binding to surface-expressed citrullinated GRP78 on mononuclear cells. J. Clin. Immunol. 2013, 33, 558–566. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, L.; Yang, Y.; Han, Y.; Wang, Y.; Liu, W.; Zuo, J. Oncogenic role of mortalin contributes to ovarian tumorigenesis by activating the MAPK-ERK pathway. J. Cell. Mol. Med. 2016, 20, 2111–2121. [Google Scholar] [CrossRef]
- Somensi, N.; Brum, P.O.; de Miranda Ramos, V.; Gasparotto, J.; Zanotto-Filho, A.; Rostirolla, D.C.; da Silva Morrone, M.; Moreira, J.C.F.; Pens Gelain, D. Extracellular HSP70 Activates ERK1/2, NF-kB and Pro-Inflammatory Gene Transcription Through Binding with RAGE in A549 Human Lung Cancer Cells. Cell. Physiol. Biochem. 2017, 42, 2507–2522. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Y.; Zhou, L.; Li, Y.; Xu, P.; Liu, X.; Lv, Q.; Li, J.; Guo, H.; Cai, H.; et al. GRP78 Promotes Neural Stem Cell Antiapoptosis and Survival in Response to Oxygen-Glucose Deprivation (OGD)/Reoxygenation through PI3K/Akt, ERK1/2, and NF-κB/p65 Pathways. Oxidative Med. Cell. Longev. 2018, 2018, 3541807. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Yuan, X.; Bao, F.; Lv, W.; He, Z.; Tang, J.; Han, J.; Hu, J. Downregulation of HSPA2 inhibits proliferation via ERK1/2 pathway and endoplasmic reticular stress in lung adenocarcinoma. Ann. Transl. Med. 2019, 7, 540. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.K.; Hong, S.K.; Park, J.I. Mortalin depletion induces MEK/ERK-dependent and ANT/CypD-mediated death in vemurafenib-resistant B-Raf(V600E) melanoma cells. Cancer Lett. 2021, 502, 25–33. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Liu, X.; Yuan, S.; Qiao, T. HSPA12B overexpression induces cisplatin resistance in non-small-cell lung cancer by regulating the PI3K/Akt/NF-κB signaling pathway. Oncol. Lett. 2018, 15, 3883–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Yang, P.; Wu, H.L.; Li, Z.W.; Li, Z.Y. GRP78 secreted by colon cancer cells facilitates cell proliferation via PI3K/Akt signaling. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 7245–7249. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, Z.; Li, Q.; Jia, C.; Zhang, N.; Qu, Y.; Hu, D. HSP70 inhibition suppressed glioma cell viability during hypoxia/reoxygenation by inhibiting the ERK1/2 and PI3K/AKT signaling pathways. J. Bioenerg. Biomembr. 2021, 53, 405–413. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Gao, W.; Zhou, Y.; Wey, S.; Mitra, S.K.; Krasnoperov, V.; Dong, D.; Liu, S.; Li, D.; et al. Monoclonal antibody against cell surface GRP78 as a novel agent in suppressing PI3K/AKT signaling, tumor growth, and metastasis. Clin. Cancer Res. 2013, 19, 6802–6811. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tseng, C.C.; Tsai, Y.L.; Fu, X.; Schiff, R.; Lee, A.S. Cancer cells resistant to therapy promote cell surface relocalization of GRP78 which complexes with PI3K and enhances PI(3,4,5)P3 production. PloS ONE 2013, 8, e80071. [Google Scholar] [CrossRef]
- Xu, F.; Lin, D.; Jiang, W.; Meng, L.; Xu, Y.; Wang, C.; Wang, X.; He, H.; Xu, D.; Zhu, Y. HSP70 inhibitor VER155008 suppresses pheochromocytoma cell and xenograft growth by inhibition of PI3K/AKT/mTOR and MEK/ERK pathways. Int. J. Clin. Exp. Pathol. 2019, 12, 2585–2594. [Google Scholar]
- Fu, Y.; Wey, S.; Wang, M.; Ye, R.; Liao, C.P.; Roy-Burman, P.; Lee, A.S. Pten null prostate tumorigenesis and AKT activation are blocked by targeted knockout of ER chaperone GRP78/BiP in prostate epithelium. Proc. Natl. Acad. Sci. USA 2008, 105, 19444–19449. [Google Scholar] [CrossRef] [Green Version]
- Wey, S.; Luo, B.; Tseng, C.C.; Ni, M.; Zhou, H.; Fu, Y.; Bhojwani, D.; Carroll, W.L.; Lee, A.S. Inducible knockout of GRP78/BiP in the hematopoietic system suppresses Pten-null leukemogenesis and AKT oncogenic signaling. Blood 2012, 119, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.T.; Zhu, G.; Pfaffenbach, K.; Kanel, G.; Stiles, B.; Lee, A.S. GRP78 as a regulator of liver steatosis and cancer progression mediated by loss of the tumor suppressor PTEN. Oncogene 2014, 33, 4997–5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Guo, W.; Zhang, Q.; Li, H.; Liu, X.; Yang, Y.; Zuo, J.; Liu, W. Crosstalk between Raf/MEK/ERK and PI3K/AKT in suppression of Bax conformational change by Grp75 under glucose deprivation conditions. J. Mol. Biol. 2011, 414, 654–666. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, S.Y.; Kim, D.H.; Park, J.S.; Jeong, S.H.; Choi, Y.W.; Kim, C.H. Crosstalk between HSPA5 arginylation and sequential ubiquitination leads to AKT degradation through autophagy flux. Autophagy 2021, 17, 961–979. [Google Scholar] [CrossRef]
- Chen, S.; Wu, J.; Jiao, K.; Wu, Q.; Ma, J.; Chen, D.; Kang, J.; Zhao, G.; Shi, Y.; Fan, D.; et al. MicroRNA-495-3p inhibits multidrug resistance by modulating autophagy through GRP78/mTOR axis in gastric cancer. Cell Death Dis. 2018, 9, 1070. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.H.; Ha, S.H. HSP70 interacts with Rheb, inhibiting mTORC1 signaling. Biochem. Biophys. Res. Commun. 2020, 533, 1198–1203. [Google Scholar] [CrossRef]
- Hansen, S.; Hupp, T.R.; Lane, D.P. Allosteric regulation of the thermostability and DNA binding activity of human p53 by specific interacting proteins. CRC Cell Transformation Group. J. Biol. Chem. 1996, 271, 3917–3924. [Google Scholar] [CrossRef] [Green Version]
- Hupp, T.R.; Meek, D.W.; Midgley, C.A.; Lane, D.P. Regulation of the specific DNA binding function of p53. Cell 1992, 71, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Walerych, D.; Olszewski, M.B.; Gutkowska, M.; Helwak, A.; Zylicz, M.; Zylicz, A. Hsp70 molecular chaperones are required to support p53 tumor suppressor activity under stress conditions. Oncogene 2009, 28, 4284–4294. [Google Scholar] [CrossRef] [Green Version]
- Wiech, M.; Olszewski, M.B.; Tracz-Gaszewska, Z.; Wawrzynow, B.; Zylicz, M.; Zylicz, A. Molecular mechanism of mutant p53 stabilization: The role of HSP70 and MDM2. PloS ONE 2012, 7, e51426. [Google Scholar] [CrossRef] [Green Version]
- Akakura, S.; Yoshida, M.; Yoneda, Y.; Horinouchi, S. A role for Hsc70 in regulating nucleocytoplasmic transport of a temperature-sensitive p53 (p53Val-135). J. Biol. Chem. 2001, 276, 14649–14657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, M.; Daugaard, M.; Jensen, M.H.; Helin, K.; Nylandsted, J.; Jäättelä, M. Members of the heat-shock protein 70 family promote cancer cell growth by distinct mechanisms. Genes Dev. 2005, 19, 570–582. [Google Scholar] [CrossRef] [Green Version]
- Boysen, M.; Kityk, R.; Mayer, M.P. Hsp70- and Hsp90-Mediated Regulation of the Conformation of p53 DNA Binding Domain and p53 Cancer Variants. Mol. Cell 2019, 74, 831–843.e4. [Google Scholar] [CrossRef]
- Dahiya, V.; Agam, G.; Lawatscheck, J.; Rutz, D.A.; Lamb, D.C.; Buchner, J. Coordinated Conformational Processing of the Tumor Suppressor Protein p53 by the Hsp70 and Hsp90 Chaperone Machineries. Mol. Cell 2019, 74, 816–830.e7. [Google Scholar] [CrossRef]
- Wang, N.; Wang, Z.; Peng, C.; You, J.; Shen, J.; Han, S.; Chen, J. Dietary compound isoliquiritigenin targets GRP78 to chemosensitize breast cancer stem cells via β-catenin/ABCG2 signaling. Carcinogenesis 2014, 35, 2544–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, Y.; Wu, H.; Zhang, L.; Yang, P.; Li, Z. GRP78 enhances the glutamine metabolism to support cell survival from glucose deficiency by modulating the β-catenin signaling. Oncotarget 2014, 5, 5369–5380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Cao, J.; Tian, J.H.; Yu, C.Y.; Huang, Q.; Yu, J.J.; Ma, R.; Wang, J.; Xu, F.; Wang, L.B. Mortalin maintains breast cancer stem cells stemness via activation of Wnt/GSK3β/β-catenin signaling pathway. Am. J. Cancer Res. 2021, 11, 2696–2716. [Google Scholar] [PubMed]
- Zhang, W.; Xue, D.; Yin, H.; Wang, S.; Li, C.; Chen, E.; Hu, D.; Tao, Y.; Yu, J.; Zheng, Q.; et al. Overexpression of HSPA1A enhances the osteogenic differentiation of bone marrow mesenchymal stem cells via activation of the Wnt/β-catenin signaling pathway. Sci. Rep. 2016, 6, 27622. [Google Scholar] [CrossRef] [Green Version]
- Ikezaki, M.; Higashimoto, N.; Matsumura, K.; Ihara, Y. Hsc70 facilitates TGF-β-induced activation of Smad2/3 in fibroblastic NRK-49F cells. Biochem. Biophys. Res. Commun. 2016, 477, 448–453. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Fan, Y.; Li, H.; Li, Z.; Li, Y. Overexpressed GRP78 affects EMT and cell-matrix adhesion via autocrine TGF-β/Smad2/3 signaling. Int. J. Biochem. Cell Biol. 2015, 64, 202–211. [Google Scholar] [CrossRef]
- Li, Y.; Kang, X.; Wang, Q. HSP70 decreases receptor-dependent phosphorylation of Smad2 and blocks TGF-β-induced epithelial-mesenchymal transition. J. Genet. Genom.=Yi Chuan Xue Bao 2011, 38, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bao, J.; Hao, J.; Peng, Y.; Hong, F. HSP70 inhibits high glucose-induced Smad3 activation and attenuates epithelial-to-mesenchymal transition of peritoneal mesothelial cells. Mol. Med. Rep. 2014, 10, 1089–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Mao, H.; Li, S.; Cao, S.; Li, Z.; Zhuang, S.; Fan, J.; Dong, X.; Borkan, S.C.; Wang, Y.; et al. HSP72 inhibits Smad3 activation and nuclear translocation in renal epithelial-to-mesenchymal transition. J. Am. Soc. Nephrol. JASN 2010, 21, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Boudesco, C.; Cause, S.; Jego, G.; Garrido, C. Hsp70: A Cancer Target Inside and Outside the Cell. Methods Mol. Biol. (Clifton N.J.) 2018, 1709, 371–396. [Google Scholar] [CrossRef]
- Sheppard, P.W.; Sun, X.; Khammash, M.; Giffard, R.G. Overexpression of heat shock protein 72 attenuates NF-κB activation using a combination of regulatory mechanisms in microglia. PLoS Comput. Biol. 2014, 10, e1003471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Gee, H.Y.; Baek, K.H.; Ko, S.K.; Park, J.M.; Lee, H.; Kim, N.D.; Lee, M.G.; Shin, I. A small molecule that binds to an ATPase domain of Hsc70 promotes membrane trafficking of mutant cystic fibrosis transmembrane conductance regulator. J. Am. Chem. Soc. 2011, 133, 20267–20276. [Google Scholar] [CrossRef]
- Macias, A.T.; Williamson, D.S.; Allen, N.; Borgognoni, J.; Clay, A.; Daniels, Z.; Dokurno, P.; Drysdale, M.J.; Francis, G.L.; Graham, C.J.; et al. Adenosine-derived inhibitors of 78 kDa glucose regulated protein (Grp78) ATPase: Insights into isoform selectivity. J. Med. Chem. 2011, 54, 4034–4041. [Google Scholar] [CrossRef]
- Guerrero-Juarez, C.F.; Lee, G.H.; Liu, Y.; Wang, S.; Karikomi, M.; Sha, Y.; Chow, R.Y.; Nguyen, T.T.L.; Iglesias, V.S.; Aasi, S.; et al. Single-cell analysis of human basal cell carcinoma reveals novel regulators of tumor growth and the tumor microenvironment. Sci. Adv. 2022, 8, eabm7981. [Google Scholar] [CrossRef]
- Sakai, K.; Inoue, M.; Mikami, S.; Nishimura, H.; Kuwabara, Y.; Kojima, A.; Toda, M.; Ogawa-Kobayashi, Y.; Kikuchi, S.; Hirata, Y.; et al. Functional inhibition of heat shock protein 70 by VER-155008 suppresses pleural mesothelioma cell proliferation via an autophagy mechanism. Thorac. Cancer 2021, 12, 491–503. [Google Scholar] [CrossRef]
- Brünnert, D.; Langer, C.; Zimmermann, L.; Bargou, R.C.; Burchardt, M.; Chatterjee, M.; Stope, M.B. The heat shock protein 70 inhibitor VER155008 suppresses the expression of HSP27, HOP and HSP90β and the androgen receptor, induces apoptosis, and attenuates prostate cancer cell growth. J. Cell. Biochem. 2020, 121, 407–417. [Google Scholar] [CrossRef]
- Dong, J.; Wu, Z.; Wang, D.; Pascal, L.E.; Nelson, J.B.; Wipf, P.; Wang, Z. Hsp70 Binds to the Androgen Receptor N-terminal Domain and Modulates the Receptor Function in Prostate Cancer Cells. Mol. Cancer Ther. 2019, 18, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kita, K.; Shiota, M.; Tanaka, M.; Otsuka, A.; Matsumoto, M.; Kato, M.; Tamada, S.; Iwao, H.; Miura, K.; Nakatani, T.; et al. Heat shock protein 70 inhibitors suppress androgen receptor expression in LNCaP95 prostate cancer cells. Cancer Sci. 2017, 108, 1820–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. The hsp70 inhibitor VER155008 induces paraptosis requiring de novo protein synthesis in anaplastic thyroid carcinoma cells. Biochem. Biophys. Res. Commun. 2014, 454, 36–41. [Google Scholar] [CrossRef]
- Massey, A.J.; Williamson, D.S.; Browne, H.; Murray, J.B.; Dokurno, P.; Shaw, T.; Macias, A.T.; Daniels, Z.; Geoffroy, S.; Dopson, M.; et al. A novel, small molecule inhibitor of Hsc70/Hsp70 potentiates Hsp90 inhibitor induced apoptosis in HCT116 colon carcinoma cells. Cancer Chemother. Pharmacol. 2010, 66, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.X.; Zhang, J.; Yang, S.S.; Wu, J.; Su, T.; Wang, W.M. Heat Shock Proteins 70 Regulate Cell Motility and Invadopodia-Associated Proteins Expression in Oral Squamous Cell Carcinoma. Front. Endocrinol. 2022, 13, 890218. [Google Scholar] [CrossRef]
- Ko, S.K.; Kim, J.; Na, D.C.; Park, S.; Park, S.H.; Hyun, J.Y.; Baek, K.H.; Kim, N.D.; Kim, N.K.; Park, Y.N.; et al. A small molecule inhibitor of ATPase activity of HSP70 induces apoptosis and has antitumor activities. Chem. Biol. 2015, 22, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Baek, K.H.; Shin, I.; Shin, I. Subcellular Hsp70 Inhibitors Promote Cancer Cell Death via Different Mechanisms. Cell Chem. Biol. 2018, 25, 1242–1254.e8. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.L.; Kim, S.A.; Choi, H.S.; Yoon, J.H.; Ahn, S.G. Epigallocatechin-3-gallate suppresses the expression of HSP70 and HSP90 and exhibits anti-tumor activity in vitro and in vivo. BMC Cancer 2010, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Ermakova, S.P.; Kang, B.S.; Choi, B.Y.; Choi, H.S.; Schuster, T.F.; Ma, W.Y.; Bode, A.M.; Dong, Z. (-)-Epigallocatechin gallate overcomes resistance to etoposide-induced cell death by targeting the molecular chaperone glucose-regulated protein 78. Cancer Res. 2006, 66, 9260–9269. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Cao, R.; Zhang, T.; Li, S.; Zhong, W. Design and synthesis of piperidine derivatives as novel human heat shock protein 70 inhibitors for the treatment of drug-resistant tumors. Eur. J. Med. Chem. 2015, 97, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Fiengo, L.; Lauro, G.; Bellone, M.L.; Bifulco, G.; Dal Piaz, F.; De Tommasi, N. The plant diterpene epoxysiderol targets Hsp70 in cancer cells, affecting its ATPase activity and reducing its translocation to plasma membrane. Int. J. Biol. Macromol. 2021, 189, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Rérole, A.L.; Gobbo, J.; De Thonel, A.; Schmitt, E.; Pais de Barros, J.P.; Hammann, A.; Lanneau, D.; Fourmaux, E.; Demidov, O.N.; Micheau, O.; et al. Peptides and aptamers targeting HSP70: A novel approach for anticancer chemotherapy. Cancer Res. 2011, 71, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Rodina, A.; Patel, P.D.; Kang, Y.; Patel, Y.; Baaklini, I.; Wong, M.J.; Taldone, T.; Yan, P.; Yang, C.; Maharaj, R.; et al. Identification of an allosteric pocket on human hsp70 reveals a mode of inhibition of this therapeutically important protein. Chem. Biol. 2013, 20, 1469–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.; Guo, Y.; Xue, Z.; Guo, Z.; Wang, Z.; Lin, D.; Zhang, H.; Pan, H.; Zhang, X.; Yin, F.; et al. Small-molecule inhibitor targeting the Hsp70-Bim protein-protein interaction in CML cells overcomes BCR-ABL-independent TKI resistance. Leukemia 2021, 35, 2862–2874. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.; Wang, Z.; Laura Bonnette, U.; Guo, Y.; Wang, H.; Gao, Q.; Zhang, Z. Bcr-Abl drives the formation of Hsp70/Bim PPI to stabilize oncogenic clients and prevent cells from undergoing apoptosis. Biochem. Pharmacol. 2022, 198, 114964. [Google Scholar] [CrossRef]
- Wang, Z.; Song, T.; Guo, Z.; Uwituze, L.B.; Guo, Y.; Zhang, H.; Wang, H.; Zhang, X.; Pan, H.; Ji, T.; et al. A novel Hsp70 inhibitor specifically targeting the cancer-related Hsp70-Bim protein-protein interaction. Eur. J. Med. Chem. 2021, 220, 113452. [Google Scholar] [CrossRef] [PubMed]
- Petit, T.; Izbicka, E.; Lawrence, R.A.; Nalin, C.; Weitman, S.D.; Von Hoff, D.D. Activity of MKT 077, a rhodacyanine dye, against human tumor colony-forming units. Anti-Cancer Drugs 1999, 10, 309–315. [Google Scholar] [CrossRef]
- Wadhwa, R.; Sugihara, T.; Yoshida, A.; Nomura, H.; Reddel, R.R.; Simpson, R.; Maruta, H.; Kaul, S.C. Selective toxicity of MKT-077 to cancer cells is mediated by its binding to the hsp70 family protein mot-2 and reactivation of p53 function. Cancer Res. 2000, 60, 6818–6821. [Google Scholar]
- Rousaki, A.; Miyata, Y.; Jinwal, U.K.; Dickey, C.A.; Gestwicki, J.E.; Zuiderweg, E.R. Allosteric drugs: The interaction of antitumor compound MKT-077 with human Hsp70 chaperones. J. Mol. Biol. 2011, 411, 614–632. [Google Scholar] [CrossRef] [Green Version]
- Deocaris, C.C.; Widodo, N.; Shrestha, B.G.; Kaur, K.; Ohtaka, M.; Yamasaki, K.; Kaul, S.C.; Wadhwa, R. Mortalin sensitizes human cancer cells to MKT-077-induced senescence. Cancer Lett. 2007, 252, 259–269. [Google Scholar] [CrossRef]
- Colvin, T.A.; Gabai, V.L.; Gong, J.; Calderwood, S.K.; Li, H.; Gummuluru, S.; Matchuk, O.N.; Smirnova, S.G.; Orlova, N.V.; Zamulaeva, I.A.; et al. Hsp70-Bag3 interactions regulate cancer-related signaling networks. Cancer Res. 2014, 74, 4731–4740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.M.; Miyata, Y.; Klinedinst, S.; Peng, H.M.; Chua, J.P.; Komiyama, T.; Li, X.; Morishima, Y.; Merry, D.E.; Pratt, W.B.; et al. Activation of Hsp70 reduces neurotoxicity by promoting polyglutamine protein degradation. Nat. Chem. Biol. 2013, 9, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, Y.; Li, X.; Lee, H.F.; Jinwal, U.K.; Srinivasan, S.R.; Seguin, S.P.; Young, Z.T.; Brodsky, J.L.; Dickey, C.A.; Sun, D.; et al. Synthesis and initial evaluation of YM-08, a blood-brain barrier permeable derivative of the heat shock protein 70 (Hsp70) inhibitor MKT-077, which reduces tau levels. ACS Chem. Neurosci. 2013, 4, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.K.; Hong, S.K.; Chen, W.; Becker, A.E.; Gundry, R.L.; Lin, C.W.; Shao, H.; Gestwicki, J.E.; Park, J.I. Mortalin (HSPA9) facilitates BRAF-mutant tumor cell survival by suppressing ANT3-mediated mitochondrial membrane permeability. Sci. Signal. 2020, 13, 1478–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Srinivasan, S.R.; Connarn, J.; Ahmad, A.; Young, Z.T.; Kabza, A.M.; Zuiderweg, E.R.; Sun, D.; Gestwicki, J.E. Analogs of the Allosteric Heat Shock Protein 70 (Hsp70) Inhibitor, MKT-077, as Anti-Cancer Agents. ACS Med. Chem. Lett. 2013, 4, 1042–1047. [Google Scholar] [CrossRef]
- Li, X.; Colvin, T.; Rauch, J.N.; Acosta-Alvear, D.; Kampmann, M.; Dunyak, B.; Hann, B.; Aftab, B.T.; Murnane, M.; Cho, M.; et al. Validation of the Hsp70-Bag3 protein-protein interaction as a potential therapeutic target in cancer. Mol. Cancer Ther. 2015, 14, 642–648. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Gestwicki, J.E. Neutral analogs of the heat shock protein 70 (Hsp70) inhibitor, JG-98. Bioorganic Med. Chem. Lett. 2020, 30, 126954. [Google Scholar] [CrossRef]
- Shao, H.; Li, X.; Moses, M.A.; Gilbert, L.A.; Kalyanaraman, C.; Young, Z.T.; Chernova, M.; Journey, S.N.; Weissman, J.S.; Hann, B.; et al. Exploration of Benzothiazole Rhodacyanines as Allosteric Inhibitors of Protein-Protein Interactions with Heat Shock Protein 70 (Hsp70). J. Med. Chem. 2018, 61, 6163–6177. [Google Scholar] [CrossRef]
- Howe, M.K.; Bodoor, K.; Carlson, D.A.; Hughes, P.F.; Alwarawrah, Y.; Loiselle, D.R.; Jaeger, A.M.; Darr, D.B.; Jordan, J.L.; Hunter, L.M.; et al. Identification of an allosteric small-molecule inhibitor selective for the inducible form of heat shock protein 70. Chem. Biol. 2014, 21, 1648–1659. [Google Scholar] [CrossRef] [Green Version]
- Adam, C.; Baeurle, A.; Brodsky, J.L.; Wipf, P.; Schrama, D.; Becker, J.C.; Houben, R. The HSP70 modulator MAL3-101 inhibits Merkel cell carcinoma. PloS ONE 2014, 9, e92041. [Google Scholar] [CrossRef] [Green Version]
- Braunstein, M.J.; Scott, S.S.; Scott, C.M.; Behrman, S.; Walter, P.; Wipf, P.; Coplan, J.D.; Chrico, W.; Joseph, D.; Brodsky, J.L.; et al. Antimyeloma Effects of the Heat Shock Protein 70 Molecular Chaperone Inhibitor MAL3-101. J. Oncol. 2011, 2011, 232037. [Google Scholar] [CrossRef] [Green Version]
- Fewell, S.W.; Smith, C.M.; Lyon, M.A.; Dumitrescu, T.P.; Wipf, P.; Day, B.W.; Brodsky, J.L. Small molecule modulators of endogenous and co-chaperone-stimulated Hsp70 ATPase activity. J. Biol. Chem. 2004, 279, 51131–51140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huryn, D.M.; Brodsky, J.L.; Brummond, K.M.; Chambers, P.G.; Eyer, B.; Ireland, A.W.; Kawasumi, M.; Laporte, M.G.; Lloyd, K.; Manteau, B.; et al. Chemical methodology as a source of small-molecule checkpoint inhibitors and heat shock protein 70 (Hsp70) modulators. Proc. Natl. Acad. Sci. USA 2011, 108, 6757–6762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Ma, J.; Zhang, J.; He, L.; Gong, J.; Long, C. Pifithrin-μ is efficacious against non-small cell lung cancer via inhibition of heat shock protein 70. Oncol. Rep. 2017, 37, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, J.I.; Pimkina, J.; Pandey, P.; Murphy, M.E.; George, D.L. HSP70 inhibition by the small-molecule 2-phenylethynesulfonamide impairs protein clearance pathways in tumor cells. Molecular Cancer Res. MCR 2011, 9, 936–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, M.; Kühnl, A.; Reins, J.; Fischer, S.; Ortiz-Tanchez, J.; Schlee, C.; Mochmann, L.H.; Heesch, S.; Benlasfer, O.; Hofmann, W.K.; et al. Antileukemic activity of the HSP70 inhibitor pifithrin-μ in acute leukemia. Blood Cancer J. 2011, 1, e28. [Google Scholar] [CrossRef] [Green Version]
- Leu, J.I.; Pimkina, J.; Frank, A.; Murphy, M.E.; George, D.L. A small molecule inhibitor of inducible heat shock protein 70. Mol. Cell 2009, 36, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Leu, J.I.; Zhang, P.; Murphy, M.E.; Marmorstein, R.; George, D.L. Structural basis for the inhibition of HSP70 and DnaK chaperones by small-molecule targeting of a C-terminal allosteric pocket. ACS Chem. Biol. 2014, 9, 2508–2516. [Google Scholar] [CrossRef] [Green Version]
- Balaburski, G.M.; Leu, J.I.; Beeharry, N.; Hayik, S.; Andrake, M.D.; Zhang, G.; Herlyn, M.; Villanueva, J.; Dunbrack, R.L., Jr.; Yen, T.; et al. A modified HSP70 inhibitor shows broad activity as an anticancer agent. Mol. Cancer Res. MCR 2013, 11, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.K.; Budina-Kolomets, A.; Murphy, M.E.; Nefedova, Y. Efficacy of the HSP70 inhibitor PET-16 in multiple myeloma. Cancer Biol. Ther. 2015, 16, 1422–1426. [Google Scholar] [CrossRef]
- Barnoud, T.; Leung, J.C.; Leu, J.I.; Basu, S.; Poli, A.N.R.; Parris, J.L.D.; Indeglia, A.; Martynyuk, T.; Good, M.; Gnanapradeepan, K.; et al. A Novel Inhibitor of HSP70 Induces Mitochondrial Toxicity and Immune Cell Recruitment in Tumors. Cancer Res. 2020, 80, 5270–5281. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.Q.; Kirby, C.A.; Zhou, W.; Schuhmann, T.; Kityk, R.; Kipp, D.R.; Baird, J.; Chen, J.; Chen, Y.; Chung, F.; et al. The novolactone natural product disrupts the allosteric regulation of Hsp70. Chem. Biol. 2015, 22, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Ambrose, A.J.; Zerio, C.J.; Sivinski, J.; Schmidlin, C.J.; Shi, T.; Ross, A.B.; Widrick, K.J.; Johnson, S.M.; Zhang, D.D.; Chapman, E. A high throughput substrate binding assay reveals hexachlorophene as an inhibitor of the ER-resident HSP70 chaperone GRP78. Bioorganic Med. Chem. Lett. 2019, 29, 1689–1693. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Maingret, L.; Puig, P.E.; Rerole, A.L.; Ghiringhelli, F.; Hammann, A.; Solary, E.; Kroemer, G.; Garrido, C. Heat shock protein 70 neutralization exerts potent antitumor effects in animal models of colon cancer and melanoma. Cancer Res. 2006, 66, 4191–4197. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, E.; Parcellier, A.; Gurbuxani, S.; Cande, C.; Hammann, A.; Morales, M.C.; Hunt, C.R.; Dix, D.J.; Kroemer, R.T.; Giordanetto, F.; et al. Chemosensitization by a non-apoptogenic heat shock protein 70-binding apoptosis-inducing factor mutant. Cancer Res. 2003, 63, 8233–8240. [Google Scholar] [PubMed]
- Ernst, K.; Liebscher, M.; Mathea, S.; Granzhan, A.; Schmid, J.; Popoff, M.R.; Ihmels, H.; Barth, H.; Schiene-Fischer, C. A novel Hsp70 inhibitor prevents cell intoxication with the actin ADP-ribosylating Clostridium perfringens iota toxin. Sci. Rep. 2016, 6, 20301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadler, S.G.; Dischino, D.D.; Malacko, A.R.; Cleaveland, J.S.; Fujihara, S.M.; Marquardt, H. Identification of a binding site on Hsc70 for the immunosuppressant 15-deoxyspergualin. Biochem. Biophys. Res. Commun. 1998, 253, 176–180. [Google Scholar] [CrossRef]
- Kawada, M.; Someno, T.; Inuma, H.; Masuda, T.; Ishizuka, M.; Takeuchi, T. The long-lasting antiproliferative effect of 15-deoxyspergualin through its spermidine moiety. J. Antibiot. 2000, 53, 705–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawada, M.; Masuda, T.; Ishizuka, M.; Takeuchi, T. 15-Deoxyspergualin inhibits Akt kinase activation and phosphatidylcholine synthesis. J. Biol. Chem. 2002, 277, 27765–27771. [Google Scholar] [CrossRef] [Green Version]
- Ramya, T.N.; Surolia, N.; Surolia, A. 15-Deoxyspergualin modulates Plasmodium falciparum heat shock protein function. Biochem. Biophys. Res. Commun. 2006, 348, 585–592. [Google Scholar] [CrossRef]
- Cerezo, M.; Lehraiki, A.; Millet, A.; Rouaud, F.; Plaisant, M.; Jaune, E.; Botton, T.; Ronco, C.; Abbe, P.; Amdouni, H.; et al. Compounds Triggering ER Stress Exert Anti-Melanoma Effects and Overcome BRAF Inhibitor Resistance. Cancer Cell 2016, 29, 805–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrose, A.J.; Santos, E.A.; Jimenez, P.C.; Rocha, D.D.; Wilke, D.V.; Beuzer, P.; Axelrod, J.; Kumar Kanduluru, A.; Fuchs, P.L.; Cang, H.; et al. Ritterostatin G(N) 1(N), a Cephalostatin-Ritterazine Bis-steroidal Pyrazine Hybrid, Selectively Targets GRP78. Chembiochem A Eur. J. Chem. Biol. 2017, 18, 506–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikeladze, M.A.; Dutysheva, E.A.; Kartsev, V.G.; Margulis, B.A.; Guzhova, I.V.; Lazarev, V.F. Disruption of the Complex between GAPDH and Hsp70 Sensitizes C6 Glioblastoma Cells to Hypoxic Stress. Int. J. Mol. Sci. 2021, 22, 1520. [Google Scholar] [CrossRef] [PubMed]
- Sverchinsky, D.V.; Nikotina, A.D.; Komarova, E.Y.; Mikhaylova, E.R.; Aksenov, N.D.; Lazarev, V.F.; Mitkevich, V.A.; Suezov, R.; Druzhilovskiy, D.S.; Poroikov, V.V.; et al. Etoposide-Induced Apoptosis in Cancer Cells Can Be Reinforced by an Uncoupled Link between Hsp70 and Caspase-3. Int. J. Mol. Sci. 2018, 19, 2519. [Google Scholar] [CrossRef] [Green Version]
- Phillips, P.A.; Dudeja, V.; McCarroll, J.A.; Borja-Cacho, D.; Dawra, R.K.; Grizzle, W.E.; Vickers, S.M.; Saluja, A.K. Triptolide induces pancreatic cancer cell death via inhibition of heat shock protein 70. Cancer Res. 2007, 67, 9407–9416. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.; Beyer, G.; Chugh, R.; Skube, S.J.; Majumder, K.; Banerjee, S.; Sangwan, V.; Li, L.; Dawra, R.; Subramanian, S.; et al. Triptolide abrogates growth of colon cancer and induces cell cycle arrest by inhibiting transcriptional activation of E2F. Lab. Investig. A J. Tech. Methods Pathol. 2015, 95, 648–659. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Thayanithy, V.; Sangwan, V.; Mackenzie, T.N.; Saluja, A.K.; Subramanian, S. Minnelide reduces tumor burden in preclinical models of osteosarcoma. Cancer Lett. 2013, 335, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Antonoff, M.B.; Chugh, R.; Skube, S.J.; Dudeja, V.; Borja-Cacho, D.; Clawson, K.A.; Vickers, S.M.; Saluja, A.K. Role of Hsp-70 in triptolide-mediated cell death of neuroblastoma. J. Surg. Res. 2010, 163, 72–78. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, T.N.; Mujumdar, N.; Banerjee, S.; Sangwan, V.; Sarver, A.; Vickers, S.; Subramanian, S.; Saluja, A.K. Triptolide induces the expression of miR-142-3p: A negative regulator of heat shock protein 70 and pancreatic cancer cell proliferation. Mol. Cancer Ther. 2013, 12, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, B.A.; Chen, E.Z.; Tang, S.; Belgum, H.S.; McCauley, J.A.; Evenson, K.A.; Etchison, R.G.; Jay-Dixon, J.; Patel, M.R.; Raza, A.; et al. Triptolide and its prodrug minnelide suppress Hsp70 and inhibit in vivo growth in a xenograft model of mesothelioma. Genes Cancer 2015, 6, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Rousalova, I.; Banerjee, S.; Sangwan, V.; Evenson, K.; McCauley, J.A.; Kratzke, R.; Vickers, S.M.; Saluja, A.; D’Cunha, J. Minnelide: A novel therapeutic that promotes apoptosis in non-small cell lung carcinoma in vivo. PloS ONE 2013, 8, e77411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chugh, R.; Sangwan, V.; Patil, S.P.; Dudeja, V.; Dawra, R.K.; Banerjee, S.; Schumacher, R.J.; Blazar, B.R.; Georg, G.I.; Vickers, S.M.; et al. A preclinical evaluation of Minnelide as a therapeutic agent against pancreatic cancer. Sci. Transl. Med. 2012, 4, 156ra139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.H.; Weng, J.R.; Lin, H.W.; Lu, M.T.; Liu, Y.C.; Chu, P.C. Targeting Triple Negative Breast Cancer Stem Cells by Heat Shock Protein 70 Inhibitors. Cancers 2022, 14, 4898. [Google Scholar] [CrossRef]
- Chang, C.S.; Kumar, V.; Lee, D.Y.; Chen, Y.; Wu, Y.C.; Gao, J.Y.; Chu, P.C. Development of Novel Rhodacyanine-Based Heat Shock Protein 70 Inhibitors. Curr. Med. Chem. 2021, 28, 5431–5446. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Song, T.; Wang, Z.; Lin, D.; Cao, K.; Liu, P.; Feng, Y.; Zhang, X.; Wang, P.; Yin, F.; et al. The chaperone Hsp70 is a BH3 receptor activated by the pro-apoptotic Bim to stabilize anti-apoptotic clients. J. Biol. Chem. 2020, 295, 12900–12909. [Google Scholar] [CrossRef]
- Sannino, S.; Yates, M.E.; Schurdak, M.E.; Oesterreich, S.; Lee, A.V.; Wipf, P.; Brodsky, J.L. Unique integrated stress response sensors regulate cancer cell susceptibility when Hsp70 activity is compromised. eLife 2021, 10, 64977–65023. [Google Scholar] [CrossRef]
- Sannino, S.; Guerriero, C.J.; Sabnis, A.J.; Stolz, D.B.; Wallace, C.T.; Wipf, P.; Watkins, S.C.; Bivona, T.G.; Brodsky, J.L. Compensatory increases of select proteostasis networks after Hsp70 inhibition in cancer cells. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.C.; Myers, M.P.; Schumacher, R.; Morimoto, R.I. Identification of a regulatory motif in Hsp70 that affects ATPase activity, substrate binding and interaction with HDJ-1. EMBO J. 1995, 14, 2281–2292. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Sverchinsky, D.V.; Mikhaylova, E.R.; Semenyuk, P.I.; Komarova, E.Y.; Niskanen, S.A.; Nikotina, A.D.; Burakov, A.V.; Kartsev, V.G.; Guzhova, I.V.; et al. Sensitizing tumor cells to conventional drugs: HSP70 chaperone inhibitors, their selection and application in cancer models. Cell Death Dis. 2018, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Guo, Z.; Guo, Y.; Wang, Z.; Tang, Y.; Song, T.; Zhang, Z. Bim transfer between Bcl-2-like protein and Hsp70 underlines Bcl-2/Hsp70 crosstalk to regulate apoptosis. Biochem. Pharmacol. 2021, 190, 114660. [Google Scholar] [CrossRef]
- Moses, M.A.; Kim, Y.S.; Rivera-Marquez, G.M.; Oshima, N.; Watson, M.J.; Beebe, K.E.; Wells, C.; Lee, S.; Zuehlke, A.D.; Shao, H.; et al. Targeting the Hsp40/Hsp70 Chaperone Axis as a Novel Strategy to Treat Castration-Resistant Prostate Cancer. Cancer Res. 2018, 78, 4022–4035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Kim, W.J.; Li, H.; Seo, W.; Park, S.H.; Kim, H.; Shin, S.C.; Zuiderweg, E.R.P.; Kim, E.E.; Sim, T.; et al. Anti-leukemia activity of a Hsp70 inhibitor and its hybrid molecules. Sci. Rep. 2017, 7, 3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanaugh, A.; Juengst, B.; Sheridan, K.; Danella, J.F.; Williams, H. Combined inhibition of heat shock proteins 90 and 70 leads to simultaneous degradation of the oncogenic signaling proteins involved in muscle invasive bladder cancer. Oncotarget 2015, 6, 39821–39838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyagawa, T.; Saito, H.; Minamiya, Y.; Mitobe, K.; Takashima, S.; Takahashi, N.; Ito, A.; Imai, K.; Motoyama, S.; Ogawa, J. Inhibition of Hsp90 and 70 sensitizes melanoma cells to hyperthermia using ferromagnetic particles with a low Curie temperature. Int. J. Clin. Oncol. 2014, 19, 722–730. [Google Scholar] [CrossRef]
- Ma, L.; Sato, F.; Sato, R.; Matsubara, T.; Hirai, K.; Yamasaki, M.; Shin, T.; Shimada, T.; Nomura, T.; Mori, K.; et al. Dual targeting of heat shock proteins 90 and 70 promotes cell death and enhances the anticancer effect of chemotherapeutic agents in bladder cancer. Oncol. Rep. 2014, 31, 2482–2492. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. Hsp70 inhibition potentiates radicicol-induced cell death in anaplastic thyroid carcinoma cells. AntiCancer Res. 2014, 34, 4829–4837. [Google Scholar]
- Reikvam, H.; Nepstad, I.; Sulen, A.; Gjertsen, B.T.; Hatfield, K.J.; Bruserud, Ø. Increased antileukemic effects in human acute myeloid leukemia by combining HSP70 and HSP90 inhibitors. Expert Opin. Investig. Drugs 2013, 22, 551–563. [Google Scholar] [CrossRef]
- McKeon, A.M.; Egan, A.; Chandanshive, J.; McMahon, H.; Griffith, D.M. Novel Improved Synthesis of HSP70 Inhibitor, Pifithrin-μ. In Vitro Synergy Quantification of Pifithrin-μ Combined with Pt Drugs in Prostate and Colorectal Cancer Cells. Molecules 2016, 21, 949. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, J.; Guo, S.Y.; Liu, H.L.; Li, S.Z. HSP70 inhibitor combined with cisplatin suppresses the cervical cancer proliferation in vitro and transplanted tumor growth: An experimental study. Asian Pac. J. Trop. Med. 2017, 10, 184–188. [Google Scholar] [CrossRef]
- Liu, J.; Liu, J.; Li, S.Z.; Zheng, Y.A.; Guo, S.Y.; Wang, X. [Inhibiting HSP70 expression enhances cisplatin sensitivity of cervical cancer cells. Nan Fang Yi Ke Da Xue Xue Bao=J. South. Med. Univ. 2016, 37, 475–481. [Google Scholar] [CrossRef]
- Alsaied, O.A.; Sangwan, V.; Banerjee, S.; Krosch, T.C.; Chugh, R.; Saluja, A.; Vickers, S.M.; Jensen, E.H. Sorafenib and triptolide as combination therapy for hepatocellular carcinoma. Surgery 2014, 156, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Budina-Kolomets, A.; Webster, M.R.; Leu, J.I.; Jennis, M.; Krepler, C.; Guerrini, A.; Kossenkov, A.V.; Xu, W.; Karakousis, G.; Schuchter, L.; et al. HSP70 Inhibition Limits FAK-Dependent Invasion and Enhances the Response to Melanoma Treatment with BRAF Inhibitors. Cancer Res. 2016, 76, 2720–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Wang, Y.; Bai, J.; Yang, Y.; Wang, F.; Feng, Y.; Zhang, R.; Li, F.; Zhang, P.; Lv, N.; et al. Blockade of HSP70 by VER-155008 synergistically enhances bortezomib-induced cytotoxicity in multiple myeloma. Cell Stress Chaperones 2020, 25, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, A.; Okur, E.; Eker, S.; Erin, N. Combined effects of the proteasome inhibitor bortezomib and Hsp70 inhibitors on the B16F10 melanoma cell line. Mol. Med. Rep. 2010, 3, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, K.; Zhang, X.; Qiu, Y.; Huang, D.; Li, W.; Xiao, X.; Tian, Y. HSP70: A promising target for laryngeal carcinoma radiaotherapy by inhibiting cleavage and degradation of nucleolin. J. Exp. Clin. Cancer Res. CR 2010, 29, 106. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stokes, J., 3rd; Singh, U.P.; Scissum Gunn, K.; Acharya, A.; Manne, U.; Mishra, M. Targeting Hsp70: A possible therapy for cancer. Cancer Lett. 2016, 374, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, B.; Lv, W.; Lai, C.; Chen, Z.; Wang, R.; Long, X.; Feng, X. Triptolide inhibits cell growth and GRP78 protein expression but induces cell apoptosis in original and radioresistant NPC cells. Oncotarget 2016, 7, 49588–49596. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Li, C.; Cao, J.; Chen, Z.; Wang, J.; Wu, Y.; Zhang, X. Liposome-templated gold nanoparticles for precisely temperature-controlled photothermal therapy based on heat shock protein expression. Colloids Surfaces. B Biointerfaces 2022, 217, 112686. [Google Scholar] [CrossRef]
- Liu, C.; Qin, H.; Kang, L.; Chen, Z.; Wang, H.; Qiu, H.; Ren, J.; Qu, X. Graphitic carbon nitride nanosheets as a multifunctional nanoplatform for photochemical internalization-enhanced photodynamic therapy. J. Mater. Chem. B 2018, 6, 7908–7915. [Google Scholar] [CrossRef]
- Yang, W.; Cui, M.; Lee, J.; Gong, W.; Wang, S.; Fu, J.; Wu, G.; Yan, K. Heat shock protein inhibitor, quercetin, as a novel adjuvant agent to improve radiofrequency ablation-induced tumor destruction and its molecular mechanism. Chin. J. Cancer Res.=Chung-Kuo Yen Cheng Yen Chiu 2016, 28, 19–28. [Google Scholar] [CrossRef]
- Zhong, Y.; Zou, Y.; Liu, L.; Li, R.; Xue, F.; Yi, T. pH-responsive Ag(2)S nanodots loaded with heat shock protein 70 inhibitor for photoacoustic imaging-guided photothermal cancer therapy. Acta Biomater. 2020, 115, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Song, H.B. Possible involvement of HSP70 in pancreatic cancer cell proliferation after heat exposure and impact on RFA postoperative patient prognosis. Biochem. Biophys. Rep. 2019, 20, 100700. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tseng, Y.T.; Suo, G.; Chen, L.; Yu, J.; Chiu, W.J.; Huang, C.C.; Lin, C.H. Photothermal therapeutic response of cancer cells to aptamer-gold nanoparticle-hybridized graphene oxide under NIR illumination. ACS Appl. Mater. Interfaces 2015, 7, 5097–5106. [Google Scholar] [CrossRef] [PubMed]
- Garrod, T.; Grubor-Bauk, B.; Yu, S.; Gargett, T.; Gowans, E.J. Encoded novel forms of HSP70 or a cytolytic protein increase DNA vaccine potency. Hum. Vaccines Immunother. 2014, 10, 2679–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xing, Y.; Zhou, Z.; Yao, W.; Cao, R.; Li, T.; Xu, M.; Wu, J. Microbial HSP70 peptide epitope 407-426 as adjuvant in tumor-derived autophagosome vaccine therapy of mouse lung cancer. Tumour Biol 2016, 37, 15097–15105. [Google Scholar] [CrossRef]
- Han, L.; Wang, W.; Lu, J.; Kong, F.; Ma, G.; Zhu, Y.; Zhao, D.; Zhu, J.; Shuai, W.; Zhou, Q.; et al. AAV-sBTLA facilitates HSP70 vaccine-triggered prophylactic antitumor immunity against a murine melanoma pulmonary metastasis model in vivo. Cancer Lett. 2014, 354, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, W.; Fang, Y.; Feng, Z.; Liao, S.; Li, W.; Li, Y.; Li, C.; Maitituoheti, M.; Dong, H.; et al. Soluble B and T lymphocyte attenuator possesses antitumor effects and facilitates heat shock protein 70 vaccine-triggered antitumor immunity against a murine TC-1 cervical cancer model in vivo. J. Immunol. 2009, 183, 7842–7850. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wan, T.; Zhou, X.; Wang, B.; Yang, F.; Li, N.; Chen, G.; Dai, S.; Liu, S.; Zhang, M.; et al. Hsp70-like protein 1 fusion protein enhances induction of carcinoembryonic antigen-specific CD8+ CTL response by dendritic cell vaccine. Cancer Res. 2005, 65, 4947–4954. [Google Scholar] [CrossRef] [Green Version]
- Ge, C.; Xing, Y.; Wang, Q.; Xiao, W.; Lu, Y.; Hu, X.; Gao, Z.; Xu, M.; Ma, Y.; Cao, R.; et al. Improved efficacy of therapeutic vaccination with dendritic cells pulsed with tumor cell lysate against hepatocellular carcinoma by introduction of 2 tandem repeats of microbial HSP70 peptide epitope 407-426 and OK-432. Int. Immunopharmacol. 2011, 11, 2200–2207. [Google Scholar] [CrossRef] [PubMed]
- Soleimanjahi, H.; Razavinikoo, H.; Fotouhi, F.; Ardebili, A. Antitumor Response to a Codon-Optimized HPV-16 E7/HSP70 Fusion Antigen DNA Vaccine. Iran. J. Immunol. IJI 2017, 14, 180–191. [Google Scholar]
- Zong, J.; Wang, C.; Wang, Q.; Peng, Q.; Xu, Y.; Xie, X.; Xu, X. HSP70 and modified HPV 16 E7 fusion gene without the addition of a signal peptide gene sequence as a candidate therapeutic tumor vaccine. Oncol. Rep. 2013, 30, 3020–3026. [Google Scholar] [CrossRef] [Green Version]
- Zong, J.; Wang, C.; Liu, B.; Liu, M.; Cao, Y.; Sun, X.; Yao, Y.; Sun, G. Human hsp70 and HPV16 oE7 fusion protein vaccine induces an effective antitumor efficacy. Oncol. Rep. 2013, 30, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzanehpour, M.; Soleimanjahi, H.; Hassan, Z.M.; Amanzadeh, A.; Ghaemi, A.; Fazeli, M. HSP70 modified response against HPV based tumor. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 228–234. [Google Scholar] [PubMed]
- Zong, J.; Peng, Q.; Wang, Q.; Zhang, T.; Fan, D.; Xu, X. Human HSP70 and modified HPV16 E7 fusion DNA vaccine induces enhanced specific CD8+ T cell responses and anti-tumor effects. Oncol. Rep. 2009, 22, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ou, X.; Xiong, J. Modified HPV16 E7/HSP70 DNA vaccine with high safety and enhanced cellular immunity represses murine lung metastatic tumors with downregulated expression of MHC class I molecules. Gynecol. Oncol. 2007, 104, 564–571. [Google Scholar] [CrossRef]
- Hauser, H.; Chen, S.Y. Augmentation of DNA vaccine potency through secretory heat shock protein-mediated antigen targeting. Methods 2003, 31, 225–231. [Google Scholar] [CrossRef]
- Chen, C.H.; Wang, T.L.; Hung, C.F.; Yang, Y.; Young, R.A.; Pardoll, D.M.; Wu, T.C. Enhancement of DNA vaccine potency by linkage of antigen gene to an HSP70 gene. Cancer Res. 2000, 60, 1035–1042. [Google Scholar] [PubMed]
- Zhang, X.; Huang, Y.; Li, X.; Wang, Y.; Yuan, Y.; Li, M. Preparation of a new combination nanoemulsion-encapsulated MAGE1-MAGE3-MAGEn/HSP70 vaccine and study of its immunotherapeutic effect. Pathol. Res. Pract. 2020, 216, 152954. [Google Scholar] [CrossRef]
- Long, P.; Zhang, Q.; Xue, M.; Cao, G.; Li, C.; Chen, W.; Jin, F.; Li, Z.; Li, R.; Wang, X.; et al. Tomato lectin-modified nanoemulsion-encapsulated MAGE1-HSP70/SEA complex protein vaccine: Targeting intestinal M cells following peroral administration. Biomed. Pharmacother. 2019, 115, 108886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Xie, D.; Zhang, W.; Xiao, G.; Wen, J. Fusion of Hsp70 to Mage-a1 enhances the potency of vaccine-specific immune responses. J. Transl. Med. 2013, 11, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, W.; Li, Y.; Li, Z.S.; Zhang, S.H.; Sun, Y.J.; Hu, P.Z.; Wang, X.M.; Huang, Y.; Si, S.Y.; Zhang, X.M.; et al. The antitumor immune responses induced by nanoemulsion-encapsulated MAGE1-HSP70/SEA complex protein vaccine following peroral administration route. Cancer Immunol. Immunother. CII 2009, 58, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Wu, Y.; Niu, T. Human DKK1 and human HSP70 fusion DNA vaccine induces an effective anti-tumor efficacy in murine multiple myeloma. Oncotarget 2018, 9, 178–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.H.; Li, Y.G.; Liang, Z.W.; Chen, M.; Peng, M.L.; Tang, L.; Hu, H.D.; Ren, H. A DNA vaccine against chimeric AFP enhanced by HSP70 suppresses growth of hepatocellular carcinoma. Cancer Immunol. Immunother. CII 2007, 56, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.H.; Li, Y.G.; Chen, M.; Tang, L.; Ren, H. [Immunotherapy with a chimeric AFP and HSP70 gene DNA vaccine targeting on a murine hepatocellular carcinoma. Zhonghua Gan Zang Bing Za Zhi=Zhonghua Ganzangbing Zazhi=Chin. J. Hepatol. 2006, 14, 510–513. [Google Scholar]
- Dong, L.; Zhang, X.; Ren, J.; Wu, S.; Yu, T.; Hou, L.; Fu, L.; Yi, S.; Yu, C. Human prostate stem cell antigen and HSP70 fusion protein vaccine inhibits prostate stem cell antigen-expressing tumor growth in mice. Cancer Biother. Radiopharm. 2013, 28, 391–397. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, C.; Zhao, J.; Fu, L.; Yi, S.; Liu, S.; Yu, T.; Chen, W. Vaccination with a DNA vaccine based on human PSCA and HSP70 adjuvant enhances the antigen-specific CD8+ T-cell response and inhibits the PSCA+ tumors growth in mice. J. Gene Med. 2007, 9, 715–726. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, W. Fusion proteins of Hsp70 with tumor-associated antigen acting as a potent tumor vaccine and the C-terminal peptide-binding domain of Hsp70 being essential in inducing antigen-independent anti-tumor response in vivo. Cell Stress Chaperones 2006, 11, 216–226. [Google Scholar] [CrossRef]
- Yamaoka, A.; Guan, X.; Takemoto, S.; Nishikawa, M.; Takakura, Y. Development of a novel Hsp70-based DNA vaccine as a multifunctional antigen delivery system. J. Control. Release 2010, 142, 411–415. [Google Scholar] [CrossRef]
- Karyampudi, L.; Ghosh, S.K. Mycobacterial HSP70 as an adjuvant in the design of an idiotype vaccine against a murine lymphoma. Cell. Immunol. 2008, 254, 74–80. [Google Scholar] [CrossRef]
- Kim, J.H.; Majumder, N.; Lin, H.; Watkins, S.; Falo, L.D., Jr.; You, Z. Induction of therapeutic antitumor immunity by in vivo administration of a lentiviral vaccine. Hum. Gene Ther. 2005, 16, 1255–1266. [Google Scholar] [CrossRef]
- Xu, M.; Zhou, L.; Zhang, Y.; Xie, Z.; Zhang, J.; Guo, L.; Wang, C.; Yang, X. A Fixed Human Umbilical Vein Endothelial Cell Vaccine With 2 Tandem Repeats of Microbial HSP70 Peptide Epitope 407-426 As Adjuvant for Therapy of Hepatoma in Mice. J. Immunother. 2015, 38, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Xing, Y.; Liu, B.; Lu, L.; Huang, X.; Ge, C.Y.; Yao, W.J.; Xu, M.L.; Gao, Z.Q.; Cao, R.Y.; et al. Protective antitumor immunity induced by tumor cell lysates conjugated with diphtheria toxin and adjuvant epitope in mouse breast tumor models. Chin. J. Cancer 2012, 31, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.H.; Wang, T.L.; Hung, C.F.; Pardoll, D.M.; Wu, T.C. Boosting with recombinant vaccinia increases HPV-16 E7-specific T cell precursor frequencies of HPV-16 E7-expressing DNA vaccines. Vaccine 2000, 18, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, J.A.; Xu, X.; Kast, W.M. The efficacy of a DNA vaccine containing inserted and replicated regions of the E7 gene for treatment of HPV-16 induced tumors. Vaccine 2007, 25, 3437–3444. [Google Scholar] [CrossRef] [Green Version]
- Propper, D.J.; Braybrooke, J.P.; Taylor, D.J.; Lodi, R.; Styles, P.; Cramer, J.A.; Collins, W.C.; Levitt, N.C.; Talbot, D.C.; Ganesan, T.S.; et al. Phase I trial of the selective mitochondrial toxin MKT077 in chemo-resistant solid tumours. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 1999, 10, 923–927. [Google Scholar] [CrossRef]
- Matsui, H.M.; Hazama, S.; Nakajima, M.; Xu, M.; Matsukuma, S.; Tokumitsu, Y.; Shindo, Y.; Tomochika, S.; Yoshida, S.; Iida, M.; et al. Novel adjuvant dendritic cell therapy with transfection of heat-shock protein 70 messenger RNA for patients with hepatocellular carcinoma: A phase I/II prospective randomized controlled clinical trial. Cancer Immunol. Immunother. CII 2021, 70, 945–957. [Google Scholar] [CrossRef]
- Nakajima, M.; Hazama, S.; Tamada, K.; Udaka, K.; Kouki, Y.; Uematsu, T.; Arima, H.; Saito, A.; Doi, S.; Matsui, H.; et al. A phase I study of multi-HLA-binding peptides derived from heat shock protein 70/glypican-3 and a novel combination adjuvant of hLAG-3Ig and Poly-ICLC for patients with metastatic gastrointestinal cancers: YNP01 trial. Cancer Immunol. Immunother. CII 2020, 69, 1651–1662. [Google Scholar] [CrossRef] [Green Version]
- Krause, S.W.; Gastpar, R.; Andreesen, R.; Gross, C.; Ullrich, H.; Thonigs, G.; Pfister, K.; Multhoff, G. Treatment of colon and lung cancer patients with ex vivo heat shock protein 70-peptide-activated, autologous natural killer cells: A clinical phase i trial. Clin. Cancer Res. 2004, 10, 3699–3707. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Qiao, Y.; Liu, B.; Laska, E.J.; Chakravarthi, P.; Kulko, J.M.; Bona, R.D.; Fang, M.; Hegde, U.; Moyo, V.; et al. Combination of imatinib mesylate with autologous leukocyte-derived heat shock protein and chronic myelogenous leukemia. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 4460–4468. [Google Scholar] [CrossRef] [Green Version]
- Skorupan, N.; Ahmad, M.I.; Steinberg, S.M.; Trepel, J.B.; Cridebring, D.; Han, H.; Von Hoff, D.D.; Alewine, C. A phase II trial of the super-enhancer inhibitor Minnelide™ in advanced refractory adenosquamous carcinoma of the pancreas. Future Oncol. 2022, 18, 2475–2481. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Binding Sites | Order | Inhibitor Name | Chemical Structures | Effects on HSP70 Activity | Cancer Types | Effects on Cancers | Mechanisms | Refs |
|---|---|---|---|---|---|---|---|---|
| NBD | 1 | VER-155008 |  | Inhibits ATPase activity of HSP70 (all isoforms) through interacting with the ATP binding site | Vismodegib-resistant basal cell carcinoma, colon cancer, mesothelioma, pheochromocytoma, castration-resistant prostate cancer, anaplastic thyroid carcinoma, breast cancer | Inhibits tumor cell proliferation, invasion, and migration in vitro; induces cell cycle arrest, apoptosis, and autophagy in vitro; inhibits tumor growth in xenograft models | Decreases the expression of the downstream Hedgehog target gene Gli1; down-regulates phosphorylation of the PI3K/AKT/mTOR and MEK/ERK signaling pathways | [59,86,87,88,89,90,91,92,93,94] |
| 2 | Apoptozole |  | Inhibits ATPase activity of HSP70 through interacting with the ATP binding site | A broad spectrum of cancer cells | Inhibits tumor cell proliferation and migration in vitro; induces cell apoptosis in vitro; inhibits tumor growth in xenograft models | Induces caspase-dependent apoptosis by blocking interaction of HSP70 with APAF-1; promotes lysosome-mediated apoptosis and impairs autophagy | [95,96,97] | |
| 3 | EGCG |  | Inhibits ATPase activity of HSP70 (HSPA5) through interacting with the ATP binding site | Breast cancer, colon cancer | Inhibits tumor cell proliferation and induce apoptosis in vitro; inhibits tumor growth in xenograft models | Induces caspase-dependent apoptosis by blocking interaction of HSPA5 with caspase-7 | [98,99] | |
| 4 | HSP70-36 |  | Inhibits ATPase activity of HSP70 through interacting with the ATP binding site | lapatinib-resistant breast cancer | Inhibits tumor cell proliferation in vitro | [100] | ||
| 5 | Epoxysiderol |  | Inhibits ATPase activity of HSP70 (HSP70A1A) through interacting with the ATP binding site | Cervical cancer | Induces cell cycle arrest and apoptosis in vitro | Decreases the expression of pAkt, pERK2, JNK1 and p38 | [101] | |
| 6 | Synthetic peptide P17 | Inhibits ATPase activity of HSP70 through interacting with the ATP binding site | Melanoma | Inhibits tumor growth in vivo | Triggers an antitumor immune response | [102] | ||
| 7 | YK5 |  | Inhibits ATPase activity of HSP70 (HSPA1A/B, A9) through interacting with a site outside the ATP binding domain | Breast cancer | Induces tumor cell apoptosis in vitro | Induces the degradation of HER2, Raf1, and Akt kinases | [103] | |
| 8 | S1g-2 |  | Inhibits ATPase activity of HSP70 through disrupting HSP70-Bim PPI | TKI-resistant chronic myeloid leukemia | Inhibits tumor cell proliferation and induces apoptosis in vitro; inhibits tumor growth in vivo | Down-regulates the level of key oncoclient proteins including AKT, Raf-1, eIF4E, and RPS16 | [104,105] | |
| 9 | S1g-6 | Inhibits ATPase activity of HSP70 through disrupting HSP70-Bim PPI | Chronic myeloid leukemia | Induces tumor cell apoptosis in vitro | Decreases the expression and phosphorylation levels of AKT and Raf-1 | [106] | ||
| 10 | MKT-077 |  | Inhibits HSP70 (HSPA8, A9)-NEFs complex | Breast cancer, ovarian cancer, endometrial cancer, colon cancer, non-small cell lung cancer, cervical cancer, osteosarcoma, and melanoma | Inhibits tumor cell proliferation and induces senescence in vitro | Releases wild-type p53 from HSP70-p53 complex; reduces tau levels | [107,108,109,110] | |
| 11 | YM-01 (MKT-077 derivative) |  | Inhibits HSP70-NEFs complex | Breast cancer, cervical cancer, melanoma | Inhibits tumor growth in xenograft models | Affects the activity of several transcription factors NF-κB, FoxM1, and Hif1α, translation regulator HuR, and cell cycle regulators p21 and survivin | [111,112] | |
| 12 | YM-08 (MKT-077 derivative) |  | Inhibits HSP70 (HSPA8, A9)-NEFs complex | Breast cancer, cervical cancer | Has a low cytotoxicity to tumor cell viability in vitro | BBB penetrant; reduces tau levels | [113] | |
| 13 | JG-98 (MKT-077 derivative) |  | Inhibits HSP70 (HSPA1A, A5, A8, A9)-NEFs complex | B-RafV600E melanoma, breast cancer, cervical cancer, skin cancer, ovarian cancer, bone marrow cancer | Inhibits tumor cell proliferation and induce apoptosis in vitro; inhibits tumor growth in xenograft models | Destabilizes FoxM1 and relieved suppression of downstream effectors, p21 and p27; destabilizes Akt1 and Raf1 | [114,115,116] | |
| 14 | JG2-38 (MKT-077 derivative) |  | Inhibits HSP70 (HSPA1A)-NEFs complex | Breast cancer, prostate cancer | Inhibits tumor cell proliferation in vitro | [117] | ||
| 15 | JG-231 (MKT-077 derivative) |  | Inhibits HSP70 (HSPA9)-NEFs complex | B-RafV600E melanoma, breast cancer | Inhibits tumor growth in xenograft models | Increases mitochondrial permeability through deregulating MEK-ERK activity | [114,118] | |
| 16 | HS-72 |  | Inhibits HSP70 (HSPA1A/B)-NEFs complex | Breast cancer, prostate cancer | Inhibits tumor growth in vitro and in vivo | [119] | ||
| 17 | MAL3-101 |  | Inhibits HSP70-JDP complex | Merkel cell carcinoma, multiple myeloma | Inhibits tumor cell proliferation and induce apoptosis in vitro; inhibits tumor growth in xenograft models | [120,121,122] | ||
| 18 | DMT3132 (MAL3-101 analog) |  | Inhibits HSP70-JDP complex | Breast cancer | Inhibits tumor cell proliferation in vitro | [123] | ||
| SBD | 19 | PES |  | Inhibits HSP70 (HSPA1A/B)-client interaction | NSCLC, breast cancer, Osteosarcoma, pancreatic cancer, Myc-Induced Lymphoma, acute myeloid leukemia, acute lymphoblastic leukemia | Inhibits tumor cell proliferation and migration in vitro; induces apoptosis and cell cycle arrest in vitro; inhibits tumor growth in xenograft models; | Down-regulates phosphorylation of AKT and ERK; causes dysfunctional autophagy and altered lysosomal function | [124,125,126,127,128] |
| 20 | PES-Cl (PES derivative) |  | Inhibits HSP70-client interaction | B-cell lymphoma, melanoma | inhibits tumor cell proliferation and autophagy in vitro; induces cell cycle arrest and genomic instability in cancer cells; inhibits tumor growth in xenograft models | [129] | ||
| 21 | PET-16 (PES derivative) |  | Inhibits HSP70-client interaction | Multiple myeloma, melanoma | Inhibits tumor cell proliferation and induces apoptosis in vitro | [128,130] | ||
| 22 | AP-4-139B (PET-16 derivative) |  | Inhibits HSP70-client interaction | Colorectal cancer | Inhibits tumor growth in vitro and in vivo | Releases DAMPs through disruption of mitochondrial function and increases immune cell recruitment into tumors | [131] | |
| 23 | Novolactone |  | Inhibits JDP-dependent ATPase activity and the allosteric control mechanism between the SBD and the NBD of HSP70 (HSPA1A/B, A5, A8) | Lung cancer | Destabilizes HER2 and EGFR in cancer cells | [132] | ||
| 24 | Hexachlorophene |  | Inhibits HSP70 (HSPA5)-client interaction | Colon cancer | Induces tumor cell apoptosis and inhibits autophagy in vitro | [133] | ||
| 25 | ADD70 | Unknown | Colorectal cancer, melanoma, breast cancer, cervical cancer, leukemia | Increases sensitivity of tumor cells to apoptosis in vitro; inhibits tumor growth in vivo | Improves cytotoxic activity of CD8+ tumor-infiltrating T cells | [134,135] | ||
| 26 | Acridizinium derivative 1 |  | Inhibits HSP70 relocalization after heat shock | Cervical cancer | Induces tumor cell apoptosis in vitro | [136] | ||
| C-terminal EEVD motif D | 27 | DSG |  | Unknown | Lymphoma | Inhibits protein synthesis and induces apoptosis in vitro; inhibits tumor growth in vivo | Inhibits activation of p70S6K and Akt | [137,138,139,140] |
| Unknown | 28 | HA15 |  | Inhibits ATPase activity of HSP70 (HSPA5) | Melanoma, prostate cancer, breast cancer, colon cancer, pancreatic cancer, glioma, cervical cancer | Induces tumor cell apoptosis and inhibits autophagy in vitro; inhibits tumor growth in xenograft models | Induces ER stress | [141] |
| 29 | Ritterostatin GN1N |  | Inhibits HSP70 (HSPA5) activity | Melanoma, colon cancer | Inhibits tumor cell proliferation in vitro | Induces ER stress and UPR | [142] | |
| 30 | AEAC | Disrupts HSP70-oxidized GAPDH complex | Glioblastoma | Inhibits tumor growth in vitro and in vivo | Promotes the aggregation of oxidized GAPDH and increases sensitivity of cells to hypoxic stress | [143] | ||
| 31 | BT-44 |  | Disrupts HSP70-Caspase-3 complex | Colon cancer, lymphoma | Increases sensitivity of tumor cells to apoptosis in vitro | [144] | ||
| 32 | Triptolide |  | Reduces the expression level of HSP70 | Pancreatic cancer, colon cancer, neuroblastoma, osteosarcoma, malignant mesothelioma, non-small cell lung cancer | Inhibits tumor cell proliferation in vitro; induces apoptosis and cell cycle arrest in vitro; inhibits tumor growth and decreases local-regional tumor spread in vivo | Down-regulates E2F activity by modulating events downstream of DNA binding; downregulates the levels of pro-survival proteins such as cMYC and survivin and targets NF-κB pathway | [145,146,147,148,149,150,151] | |
| 33 | Minnelide (Triptolide derivative) |  | Reduces the expression level of HSP70 | Colon cancer, osteosarcoma, malignant mesothelioma, non-small cell lung cancer | Inhibits the tumor growth in the xenograft and metastasis model | Downregulates the levels of pro-survival proteins such as cMYC and survivin and targets NF-κB pathway | [146,147,149,150,151,152] | |
| 34 | Quercetin |  | Reduces the expression level of HSP70 | Prostate cancer | Inhibits tumor cell proliferation and induces apoptosis in vitro | [92] | ||
| 35 | Rhodacyanine-based compound 1 |  | Inhibits the chaperone activity of HSP70 | Triple negative breast cancer | Inhibits tumor growth in vitro and in vivo | Down-regulates β-catenin | [153,154] | |
| 36 | Rhodacyanine-based compound 6 |  | Inhibits the chaperone activity of HSP70 | Triple negative breast cancer | Inhibits tumor growth in vitro and in vivo | Down-regulates β-catenin | [153,154] |
| Order | Vaccines | Vaccine Types | Cancer Types | Administration Methods | Immunotherapy Effects | Immune Mechanisms | Refs |
|---|---|---|---|---|---|---|---|
| 1 | E7-HSP70 (optimizedE7/mHSP70, modifiedE7/mtHSP70, optimizedE7/hHSP70, sigmE7/mtHSP70, modifiedE7/HSP70) | DNA vaccine, protein vaccine | Cervical cancer, lung metastatic melanoma | Subcutaneous injection, intramuscular injection | Prophylactic and therapeutic antitumor effects | Cellular immune response | [190,191,192,193,194,195,196,197] |
| 2 | MAGE-HSP70 (MAGE1-MAGE3-MAGEn/HSP70, TL-MAGE1-HSP70/SEA, MAGE1/HSP70, MAGE1-HSP70/SEA) | Nanoemulsion-encapsulated protein vaccine, recombinant protein vaccine | Hepatocellular carcinoma, melanoma, | Intraperitoneal injection, oral, subcutaneous injection | Prophylactic and therapeutic antitumor effects | Humoral and cellular immune responses | [198,199,200,201] |
| 3 | hDKK1-hHSP70 | DNA vaccine | Multiple myeloma | Intramuscular injection | Prophylactic and therapeutic antitumor effects | Humoral and cellular immune responses | [202] |
| 4 | AFP-HSP70 | DNA vaccine, protein vaccine | Hepatocellular carcinoma | Intramuscular injection | Therapeutic antitumor effect | Cellular immune response | [203,204] |
| 5 | PSCA-HSP70 | Protein vaccine, DNA vacine | Prostate cancer | Subcutaneous injection, intramuscular injection | Therapeutic antitumor effect | Humoral and cellular immune responses | [205,206] |
| 6 | Mela-HSP70 | Protein vaccine | Melanoma | Subcutaneous injection | Prophylactic antitumor effect | Cellular immune response | [207] |
| 7 | OVA257–264-HSP70 | DNA vaccine | Lymphoma | Intradermal injection | Prophylactic antitumor effect | Cellular immune response | [208] |
| 8 | A20Id-mycHSP70 | Protein vaccine | Lymphoma | Intraperitoneal injection | Prophylactic antitumor effect | Humoral and cellular immune responses | [209] |
| 9 | LV-TRP2-HSP70 | Lentiviral vector vaccine | Melanoma, breast cancer, glioblastoma | Subcutaneous injection | Therapeutic antitumor effect | Cellular immune response | [210] |
| 10 | DRibble vaccine+mtHSP70407–426 (M2) | Conjugated vaccine | Lung cancer | Subcutaneous injection | Therapeutic antitumor effect | Cellular immune response | [185] |
| 11 | HUVEC vaccine+mtHSP70407–426 (M2) | Conjugated vaccine | Hepatocellular carcinoma | Subcutaneous injection | Prophylactic and therapeutic antitumor effects | Humoral and cellular immune responses | [211] |
| 12 | BTLA vaccine+HSP70 vaccine (AAV-sBTLA vaccine+HSP70 vaccine, psBTLA vaccine+HSP70 vaccine) | Recombinant adenovirus vector vaccine+protein vaccine, DNA vaccine+protein vaccine | Lung metastatic melanoma, cervical cancer | Tail vein injection, intramuscular injection | Prophylactic and therapeutic antitumor effects | Cellular immune response | [186,187] |
| 13 | DT-TCL-mtHSP70407–426 (M2) | Tumor cell lysate vaccine | Breast cancer | Subcutaneous injection | Prophylactic antitumor effect | Humoral and cellular immune responses | [212] |
| 14 | DCs pulsed with CEA576–669-HSP70L1 | DC vaccine | Colon cancer | Intraperitoneal injection | Therapeutic antitumor effect | Cellular immune response | [188] |
| 15 | DCs pulsed with tumor cell lysate pulsed with mtHSP70407–426 (M2) and OK-432 | DC vaccine | Hepatocellular carcinoma | Subcutaneous injection | Therapeutic antitumor effect | Cellular immune response | [189] |
| Therapy Type | NCT Number | Drug Name or Vaccine Component | Inclusion Criteria | Study Type | Clinical Phase | Recruitment Status | Publication |
|---|---|---|---|---|---|---|---|
| HSP70 inhibitors | NCT01927965 | Minnelide | Advanced gastrointestinal tumors | Non-randomized/one-arm | Phase I | Completed | |
| NCT04896073 | Minnelide | Advanced refractory adenosquamous carcinoma of the pancreas | Non-randomized/one-arm | Phase II | Recruiting | [220] | |
| HSP70-based vaccines | NCT00027144 | Autologous tumor-derived HSP70 protein vaccine | Chronic myeloid leukemia | Non-randomized/one-arm | Phase I | Completed | |
| NCT00030303 | Autologous PMNCs-derived HSP70 protein vaccine | Chronic myeloid leukemia | Non-randomized/one-arm | Phase I | Completed | [219] | |
| NCT00005633 | Tyrosinase and gp100 peptides fused with OVA BiP peptide and recombinant HSP70 protein | Stage III or stage IV melanoma | Non-randomized/one-arm | Phase I | Completed | ||
| NCT05059821 | Autologous activated monocytes with autologous tumor-derived HSP70 protein vaccine | Hepatocellular carcinoma who developed recurrence after surgical resection and refractory to the available institutional standard of care lines of treatment | Non-randomized/one-arm | Phase I | Recruiting |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, K.; Zhou, G.; Liu, Y.; Zhang, J.; Chen, Y.; Liu, L.; Zhang, G. HSP70 Family in Cancer: Signaling Mechanisms and Therapeutic Advances. Biomolecules 2023, 13, 601. https://doi.org/10.3390/biom13040601
Zhao K, Zhou G, Liu Y, Zhang J, Chen Y, Liu L, Zhang G. HSP70 Family in Cancer: Signaling Mechanisms and Therapeutic Advances. Biomolecules. 2023; 13(4):601. https://doi.org/10.3390/biom13040601
Chicago/Turabian StyleZhao, Kejia, Guanyu Zhou, Yu Liu, Jian Zhang, Yaohui Chen, Lunxu Liu, and Gao Zhang. 2023. "HSP70 Family in Cancer: Signaling Mechanisms and Therapeutic Advances" Biomolecules 13, no. 4: 601. https://doi.org/10.3390/biom13040601