
Feasibility of Transport of 26 Biologically Active Ultrashort Peptides via LAT and PEPT Family Transporters

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Modeling and Preparation of Receptors and Ligands
2.2. Virtual Screening of Ligands
3. Results
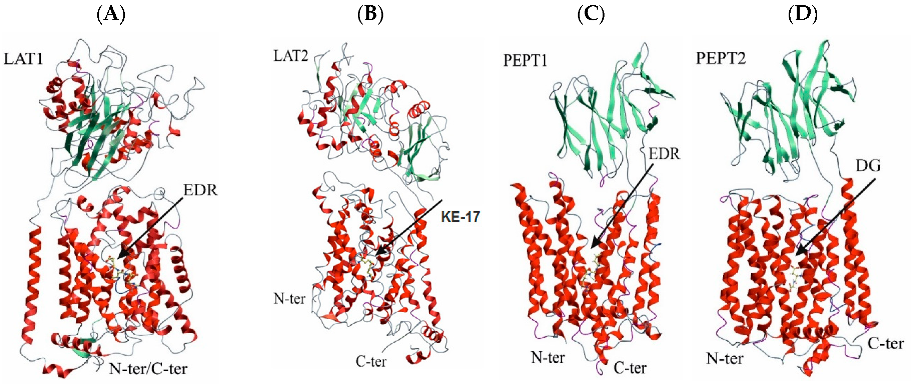
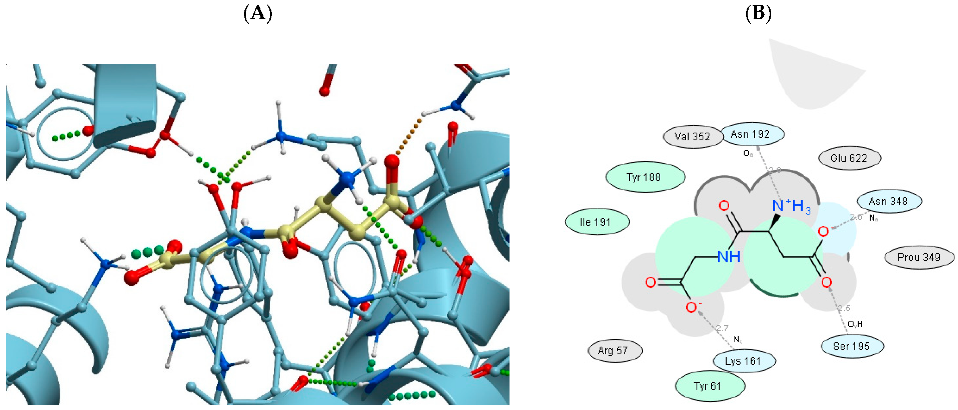
3.1. Spatial Structures of UPS Complexes with LAT1, LAT2, PEPT1, and PEPT2 Transporters
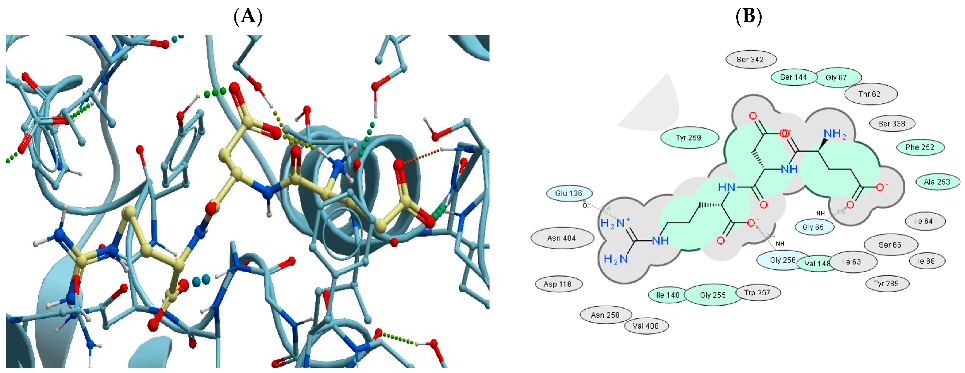
3.2. Binding of USPs to the LAT1 Amino Acid Transporter


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Two-Dimensional Structure of the Peptide/Peptoid | Amino Acid Sequence or Peptide/Peptoid Name | Receptor in the PDB | ICM-Score * |
|---|---|---|---|---|
| 1 |  | EDR | 7DSK | −45.32 |
| 2 |  | EDG | 7DSK | −43.40 |
| 3 |  | AEDR | 7DSN | −35.35 |
| 4 |  | EDL | 7DSK | −35.03 |
| 5 |  | ED | 6IRS | −34.55 |
| 6 |  | KEDG | 6IRT | −34.05 |
| 7 |  | AEDG | 6IRT | −32.93 |
| 8 |  | AB-17 | 7DSL | −32.76 |
| 9 |  | EDP | 7DSK | −32.60 |
| 10 |  | DS | 6IRT | −30.49 |
| 11 |  | KEDA | 6IRT | −28.83 |
| 12 |  | EW | 7DSK | −28.72 |
| 13 |  | KEDW-NH2 | 6IRT | −28.40 |
| 14 |  | AED | 6IRS | −28.14 |
| 15 |  | DR | 6IRT | −28.01 |
| 16 |  | DS7 | 7DSL | −27.38 |
| 17 |  | DA | 6IRS | −27.25 |
| 18 |  | DG | 6IRT | −26.49 |
| 19 |  | DW | 7DSN | −25.18 |
| 20 |  | AEDL | 6IRS | −25.11 |
| 21 |  | KED | 6IRS | −24.79 |
| 22 |  | KEDP | 6IRS | −22.79 |
| 23 |  | AEDP | 7DSK | −20.95 |
| 24 |  | KE | 7DSK | −19.02 |
| 25 |  | DL | 7DSK | −16.70 |
| 26 |  | AE | 7DSL | −16.59 |
| No. | Two-Dimensional Structure of the Inhibitor | Name of the Inhibitor | Receptor in the PDB | ICM-Score | IC50 µM | Reference |
|---|---|---|---|---|---|---|
| 1 |  | (2S)-2-amino-3-[4-[(5-amino-2-phenyl-1,3-benzox (JPH203) | 7DSK | −31.50 | 0.35 | [29] |
| 2 |  | 2-Amino-2-norbornanecarboxylic acid (BCH) | 6IRS | −17.37 | 78.0 | [29] |
| 3 |  | 5-(benzyloxy)-tryptophan | 7DSK | −25.25 | 1.48 * | [10] |
| 4 |  | l-2-amino-4-(3,5-dichlorophenyl)-butanoic acid | 7DSK | −19.84 | 0.62 * | [10] |
| 5 |  | l-Leucine | 7DSN | −23.96 | 9.0 | [29] |
| 6 |  | S-(3-bromo-4-methoxybenzyl)-l-cysteine | 7DSK | −23.44 | 33.2 | [30] |
| 7 |  | Tryptophan | 7DSN | −23.17 | 20.2 | [30] |
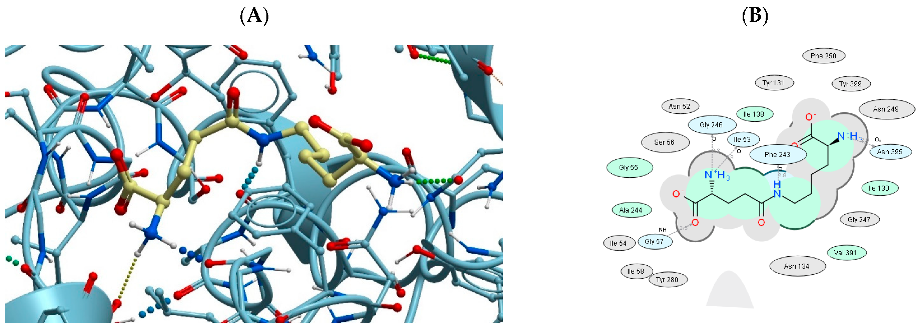
3.3. Binding of USPs to the LAT2 Amino Acid Transporter
| No. | Amino Acid Sequence or Peptide/Peptoid Name | Receptor in the PDB | ICM-Score |
|---|---|---|---|
| 1 | AB-17 | 7B00 | −39.60 |
| 2 | EDR | 7CMH | −32.04 |
| 3 | ED | 7B00 | −31.35 |
| 4 | DS7 | 7CMI | −29.29 |
| 5 | DS | 7CMI | −29.17 |
| 6 | DR | 7CMH | −27.98 |
| 7 | AEDR | 7CMI | −27.54 |
| 8 | EDG | 7B00 | −26.82 |
| 9 | KEDG | 7CMI | −26.57 |
| 10 | KE | 7CMH | −23.94 |
| 11 | DA | 7B00 | −23.73 |
| 12 | AED | 7CMH | −23.64 |
| 13 | AEDG | 7CMH | −23.62 |
| 14 | AEDP | 7CMI | −22.15 |
| 15 | DW | 7CMH | −22.08 |
| 16 | KEDW-NH2 | 7B00 | −21.78 |
| 17 | KEDA | 7CMH | −21.45 |
| 18 | EW | 7CMI | −20.12 |
| 19 | KED | 7CMH | −20.09 |
| 20 | DG | 7CMI | −20.08 |
| 21 | DL | 7CMH | −19.05 |
| 22 | AEDL | 7CMH | −18.88 |
| 23 | EDP | 7CMI | −18.75 |
| 24 | EDL | 7CMH | −17.37 |
| 25 | AE | 7CMH | −16.87 |
| 26 | KEDP | 7B00 | −11.68 |
| No. | Two-Dimensional Structure of the Inhibitor | Inhibitor Name | Receptor in the the PDB | ICM-Score | IC50 µM |
|---|---|---|---|---|---|
| 1 |  | (2S)-2-amino-3-[4-[(5-amino-2-phenyl-1,3-benzox (JPH203) | 7B00 | −34.69 | 1.7 |
| 2 |  | 2-Amino-2-norbornanecarboxylic acid (BCH) | 7B00 | −15.15 | 184.0 |
| 3 |  | L-Leucine | 7CMH | −27.78 | 39.0 |
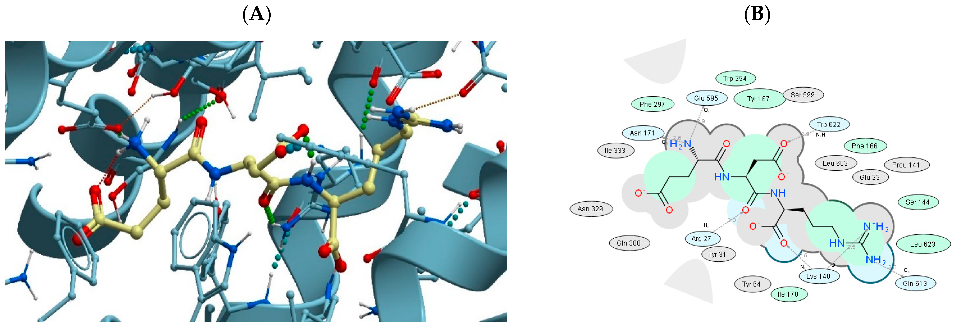
3.4. Binding of USPs to the Di- and Tri-Peptide Transporter PEPT1

| No. | Amino Acid Sequence or Peptide/Peptoid Name | Receptor in the PDB | ICM-Score |
|---|---|---|---|
| 1 | EDR | 7PMX | −37.59 |
| 2 | KEDP | 7PMX | −33.42 |
| 3 | AEDR | 7PN1 | −27.50 |
| 4 | DS | 7PN1 | −26.17 |
| 5 | DR | 7PMX | −25.74 |
| 6 | AEDL | 7PN1 | −25.35 |
| 7 | KEDG | 7PN1 | −22.72 |
| 8 | DG | 7PMX | −22.22 |
| 9 | KEDW-NH2 | 7PN1 | −20.57 |
| 10 | EDL | 7PN1 | −20.48 |
| 11 | AE | 7PMX | −20.05 |
| 12 | EW | 7PN1 | −19.00 |
| 13 | AED | 7PN1 | −18.95 |
| 14 | DA | 7PN1 | −18.26 |
| 15 | EDG | 7PN1 | −18.04 |
| 16 | AEDG | 7PN1 | −18.00 |
| 17 | DS7 | 7PN1 | −17.57 |
| 18 | EDP | 7PN1 | −17.29 |
| 19 | KE | 7PN1 | −17.02 |
| 20 | KEDA | 7PN1 | −16.70 |
| 21 | KED | 7PN1 | −16.42 |
| 22 | ED | 7PN1 | −15.76 |
| 23 | AB17 | 7PMW | −15.25 |
| 24 | DL | 7PMX | −13.88 |
| 25 | AEDP | 7PN1 | −12.65 |
| 26 | DW | 7PMW | −10.66 |
| No. | Two-Dimensional Structure of the Inhibitor | Inhibitor Name | Receptor in the PDB | ICM-Score | IC50 µM |
|---|---|---|---|---|---|
| 1 |  | Val-[3-(hydroxymethyl)-phenyl]guanidine | 7PN1 | −27.74 | 650 |
| 2 |  | Guanidin-oseltamivir-carboxylate-L-Val-up | 7PMX | −12.75 | 190 * |
| 3 |  | Guanidin-oseltamivir-carboxylate-L-Val-down | 7PMX | −11.64 | 190 * |
| 4 |  | Val-Zanamivir | 7PN1 | −7.90 | 1190 |
3.5. Binding of USPs to the Di- and Tri-Peptide Transporter PEPT2

| No. | Amino Acid Sequence or Peptide/Peptoid Name | ICM-Score |
|---|---|---|
| 1 | DG | −27.16 |
| 2 | DA | −25.92 |
| 3 | ED | −18.22 |
| 4 | DS | −13.39 |
| 5 | DS7 | −11.20 |
| 6 | AE | −10.70 |
| 7 | AED | −7.97 |
| 8 | KEDP | −6.36 |
| 9 | EDP | −5.40 |
| 10 | EDG | −5.20 |
| 11 | AB17 | −4.19 |
| 12 | DR | −3.80 |
| 13 | EW | −0.78 |
| 14 | AEDG | 1.46 |
| 15 | KE | 6.40 |
| 16 | KEDW-NH2 | 6.90 |
| 17 | EDR | 8.99 |
| 18 | DL | 9.17 |
| 19 | DW | 10.03 |
| 20 | KED | 11.14 |
| 21 | KEDA | 11.60 |
| 22 | AEDL | 12.13 |
| 23 | KEDG | 12.26 |
| 24 | AEDR | 13.15 |
| 25 | EDL | 13.48 |
| 26 | AEDP | 17.28 |
| No. | Two-Dimensional Structure of the Inhibitor | Inhibitor Name | Receptor in the PDB | ICM-Score | IC50 µM |
|---|---|---|---|---|---|
| 1 |  | Carnosine | 7PMY | −20.63 | 66 |
| 2 |  | Bestatin | 7PMY | −2.94 | 18 |
| 3 |  | Alanine-4-nitroanilide | 7PMY | −2.50 | 29 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khavinson, V.K.; Popovich, I.G.; Linkova, N.S.; Mironova, E.S.; Ilina, A.R. Peptide Regulation of Gene Expression: A Systematic Review. Molecules 2021, 26, 7053. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Dong, X.; Yang, F.; Zhou, S.; Xiang, M.; Lou, L.; Yao, S.Q.; Gao, L. Engineered Cell-Penetrating Peptides for Mito-chondrion-Targeted Drug Delivery in Cancer Therapy. Chem. A Eur. J. 2021, 27, 14721–14729. [Google Scholar] [CrossRef] [PubMed]
- Allolio, C.; Magarkar, A.; Jurkiewicz, P.; Baxová, K.; Javanainen, M.; Mason, P.E.; Šachl, R.; Cebecauer, M.; Hof, M.; Horinek, D.; et al. Arginine-rich cell-penetrating peptides induce membrane multilamellarity and subsequently enter via formation of a fusion pore. Proc. Natl. Acad. Sci. USA 2018, 115, 11923–11928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khavinson, V.; Linkova, N.; Kozhevnikova, E.; Dyatlova, A.; Petukhov, M. Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers. Int. J. Mol. Sci. 2022, 23, 7733. [Google Scholar] [CrossRef] [PubMed]
- Campos-Bedolla, P.; Walter, F.R.; Veszelka, S.; Deli, M.A. Role of the Blood–Brain Barrier in the Nutrition of the Central Nervous System. Arch. Med. Res. 2014, 45, 610–638. [Google Scholar] [CrossRef]
- Li, S.; Whorton, A.R. Identification of stereoselective transporters for S-nitroso-L-cysteine: Role of LAT1 and LAT2 in biological activity of S-nitrosothiols. J. Biol. Chem. 2005, 280, 20102–20110. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Nishimura, T.; Higuchi, K.; Noguchi, S.; Tega, Y.; Kurosawa, T.; Deguchi, Y.; Tomi, M. Transport of Pregabalin Via L-Type Amino Acid Transporter 1 (SLC7A5) in Human Brain Capillary Endothelial Cell Line. Pharm. Res. 2018, 35, 246. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, G.J.; Lam, H.Y.; Begleiter, A. Active carrier-mediated transport of melphalan by two separate amino acid transport systems in LPC-1 plasmacytoma cells in vitro. J. Biol. Chem. 1979, 254, 1057–1064. [Google Scholar] [CrossRef]
- Widdows, K.L.; Panitchob, N.; Crocker, I.P.; Please, C.P.; Hanson, M.A.; Sibley, C.P.; Johnstone, E.D.; Sengers, B.G.; Lewis, R.M.; Glazier, J.D. Integration of computational modeling with membrane transport studies reveals new insights into amino acid exchange transport mechanisms. FASEB J. 2015, 29, 2583–2594. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Ecker, G.F. Insights into the Structure, Function, and Ligand Discovery of the Large Neutral Amino Acid Transporter 1, LAT1. Int. J. Mol. Sci. 2018, 19, 1278. [Google Scholar] [CrossRef] [Green Version]
- Meier, C.; Ristic, Z.; Klauser, S.; Verrey, F. Activation of system L heterodimeric amino acid exchangers by intracellular substrates. EMBO J. 2002, 21, 580–589. [Google Scholar] [CrossRef]
- Okano, N.; Hana, K.; Naruge, D.; Kawai, K.; Kobayashi, T.; Nagashima, F.; Endou, H.; Furuse, J. Biomarker Analyses in Patients with Advanced Solid Tumors Treated with the LAT1 Inhibitor JPH203. In Vivo 2020, 34, 2595–2606. [Google Scholar] [CrossRef]
- Del Amo, E.M.; Urtti, A.; Yliperttula, M. Pharmacokinetic role of L-type amino acid transporters LAT1 and LAT2. Eur. J. Pharm. Sci. 2008, 35, 161–174. [Google Scholar] [CrossRef]
- Jeckelmann, J.M.; Lemmin, T.; Schlapschy, M.; Skerra, A.; Fotiadis, D. Structure of the human heterodimeric transporter 4F2hc-LAT2 in complex with Anticalin, an alternative binding protein for applications in single-particle cryo-EM. Sci. Rep. 2022, 12, 18269. [Google Scholar] [CrossRef]
- Pineda, M.; Fernández, E.; Torrents, D.; Estévez, R.; López, C.; Camps, M.; Lloberas, J.; Zorzano, A.; Palacín, M. Identification of a membrane protein, LAT-2, that co-expresses with 4F2 heavy chain, an L-type amino acid transport activity with broad specificity for small and large zwitterionic amino acids. J. Biol. Chem. 1999, 274, 19738–19744. [Google Scholar] [CrossRef] [Green Version]
- Zevenbergen, C.; Meima, M.E.; Lima de Souza, E.C.; Peeters, R.P.; Kinne, A.; Krause, G.; Visser, W.E.; Visser, T.J. Transport of Iodothyronines by Human L-Type Amino Acid Transporters. Endocrinology 2015, 156, 4345–4355. [Google Scholar] [CrossRef] [Green Version]
- Pinto, V.; Pinho, M.J.; Soares-da-Silva, P. Renal amino acid transport systems and essential hypertension. FASEB J. 2013, 27, 2927–2938. [Google Scholar] [CrossRef]
- Barollo, S.; Bertazza, L.; Watutantrige-Fernando, S.; Censi, S.; Cavedon, E.; Galuppini, F.; Pennelli, G.; Fassina, A.; Citton, M.; Rubin, B. Overexpression of L-type amino acid transporter 1 (LAT1) and 2 (LAT2): Novel markers of neuroendocrine tumors. PLoS ONE 2016, 11, e0156044. [Google Scholar] [CrossRef] [Green Version]
- Chiu, M.; Sabino, C.; Taurino, G.; Bianchi, M.G.; Andreoli, R.; Giuliani, N.; Bussolati, O. GPNA inhibits the sodium-independent transport system L for neutral amino acids. Amino Acids 2017, 49, 1365–1372. [Google Scholar] [CrossRef]
- Wang, Q.; Holst, J. L-type amino acid transport and cancer: Targeting the mTORC1 pathway to inhibit neoplasia. Am. J. Cancer Res. 2015, 5, 1281–1294. [Google Scholar]
- Terada, T.; Sawada, K.; Irie, M.; Saito, H.; Hashimoto, Y.; Inui, K.-I. Structural requirements for determining the substrate affinity of peptide transporters PEPT1 and PEPT2. Pflüg. Arch. 2000, 440, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.A.; Keep, R.F.; Smith, D.E. Role and Relevance of PEPT2 in Drug Disposition, Dynamics, and Toxicity. Drug Metab. Pharmacokinet. 2008, 23, 236–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abagyan, R.; Totrov, M. Biased probability Monte Carlo conformational searches and electrostatic calculations for peptides and proteins. J. Mol. Biol. 1994, 235, 983–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abagyan, R.; Orry, A.; Raush, E.; Totrov, M. What constitutes a good docking score? In ICM-Pro User Guide v.3.8; MolSoft LLC: San Diego, CA, USA, 2018. [Google Scholar]
- Kolchina, N.; Khavinson, V.; Linkova, N.; Yakimov, A.; Baitin, D.; Afanasyeva, A.; Petukhov, M. Systematic search for structural motifs of peptide binding to double-stranded DNA. Nucleic Acids Res. 2019, 47, 10553–10563. [Google Scholar] [CrossRef]
- Ilina, A.; Khavinson, V.; Linkova, N.; Petukhov, M. Neuroepigenetic Mechanisms of Action of Ultrashort Peptides in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 4259. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Linkova, N.S.; Umnov, R.S. Peptide KED: Molecular-Genetic Aspects of Neurogenesis Regulation in Alzheimer’s Disease. Bull. Exp. Biol. Med. 2021, 171, 190–193. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Pronyaeva, V.E.; Linkova, N.S.; Trofimova, S.V. Peptidergic Regulation of Differentiation of Embrionic Cells. Bull. Exp. Biol. Med. 2013, 155, 172–174. [Google Scholar] [CrossRef]
- Kantipudi, S.; Fotiadis, D. Yeast Cell-Based Transport Assay for the Functional Characterization of Human 4F2hc-LAT1 and-LAT2, and LAT1 and LAT2 Substrates and Inhibitors. Front. Mol. Biosci. 2021, 8, 456–467. [Google Scholar] [CrossRef]
- Yan, R.; Zhao, X.; Lei, J.; Zhou, Q. Structure of the human LAT1-4F2hc heteromeric amino acid transporter complex. Nature 2019, 568, 127–130. [Google Scholar] [CrossRef]
- Yan, R.; Zhou, J.; Li, Y.; Lei, J.; Zhou, Q. Structural insight into the substrate recognition and transport mechanism of the human LAT2–4F2hc complex. Cell Discov. 2020, 6, 82–85. [Google Scholar] [CrossRef]
- Saaby, L.; Nielsen, C.U.; Steffansen, B.; Larsen, S.B.; Brodin, B. Current status of rational design of prodrugs targeting the intestinal di/tri-peptide transporter hPEPT1 (SLC15A1). J. Drug Deliv. Sci. Technol. 2013, 23, 307–314. [Google Scholar] [CrossRef]
- Kaira, K.; Sunose, Y.; Ohshima, Y.; Ishioka, N.S.; Arakawa, K.; Ogawa, T.; Sunaga, N.; Shimizu, K.; Tominaga, H.; Oriuchi, N.; et al. Clinical significance of L-type amino acid transporter 1 expression as a prognostic marker and potential of new targeting therapy in biliary tract cancer. BMC Cancer 2013, 13, 482. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, Y.; Kaira, K.; Yamaguchi, A.; Oriuchi, N.; Tominaga, H.; Nagamori, S.; Kanai, Y.; Yokobori, T.; Miyazaki, T.; Asao, T.; et al. Efficacy of system l amino acid transporter 1 inhibition as a therapeutic target in esophageal squamous cell carcinoma. Cancer Sci. 2016, 107, 1499–1505. [Google Scholar] [CrossRef]
- Vinogradova, I.A.; Bukalev, A.V.; Zabezhinski, M.A.; Semenchenko, A.V.; Khavinson, V.K.; Anisimov, V.N. Geroprotective effect of Ala-Glu-Asp-Gly peptide in male rats exposed to different illumination regimens. Bull. Exp. Biol. Med. 2008, 145, 472–477. [Google Scholar] [CrossRef]
- Kossoy, G.; Zandbank, J.; Tendler, E.; Anisimov, V.; Khavinson, V.; Popovich, I.; Zabezhinski, M.; Zusman, I.; Ben-Hur, H. Epitalon and colon carcinogenesis in rats: Proliferative activity and apoptosis in colon tumors and mucosa. Int. J. Mol. Med. 2003, 12, 473–477. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Khavinson, V.K.; Alimova, I.N.; Semchenko, A.V.; Yashin, A.I. Epithalon Decelerates Aging and Suppresses Development of Breast Adenocarcinomas in Transgenic HER-2/neu Mice. Bull. Exp. Biol. Med. 2002, 1349, 187–190. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Khavinson, V.K.; Mikhalski, A.I.; Yashin, A.I. Effect of synthetic thymic and pineal peptides on biomarkers of ageing, survival and spontaneous tumour incidence in female CBA mice. Mech. Ageing Dev. 2001, 122, 41–68. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khavinson, V.K.; Linkova, N.S.; Rudskoy, A.I.; Petukhov, M.G. Feasibility of Transport of 26 Biologically Active Ultrashort Peptides via LAT and PEPT Family Transporters. Biomolecules 2023, 13, 552. https://doi.org/10.3390/biom13030552
Khavinson VK, Linkova NS, Rudskoy AI, Petukhov MG. Feasibility of Transport of 26 Biologically Active Ultrashort Peptides via LAT and PEPT Family Transporters. Biomolecules. 2023; 13(3):552. https://doi.org/10.3390/biom13030552
Chicago/Turabian StyleKhavinson, Vladimir Khatskelevich, Natalia Sergeevna Linkova, Andrey Ivanovich Rudskoy, and Michael Gennadievich Petukhov. 2023. "Feasibility of Transport of 26 Biologically Active Ultrashort Peptides via LAT and PEPT Family Transporters" Biomolecules 13, no. 3: 552. https://doi.org/10.3390/biom13030552