
HSP70 and Primary Arterial Hypertension
 and
and
Abstract
:1. Introduction
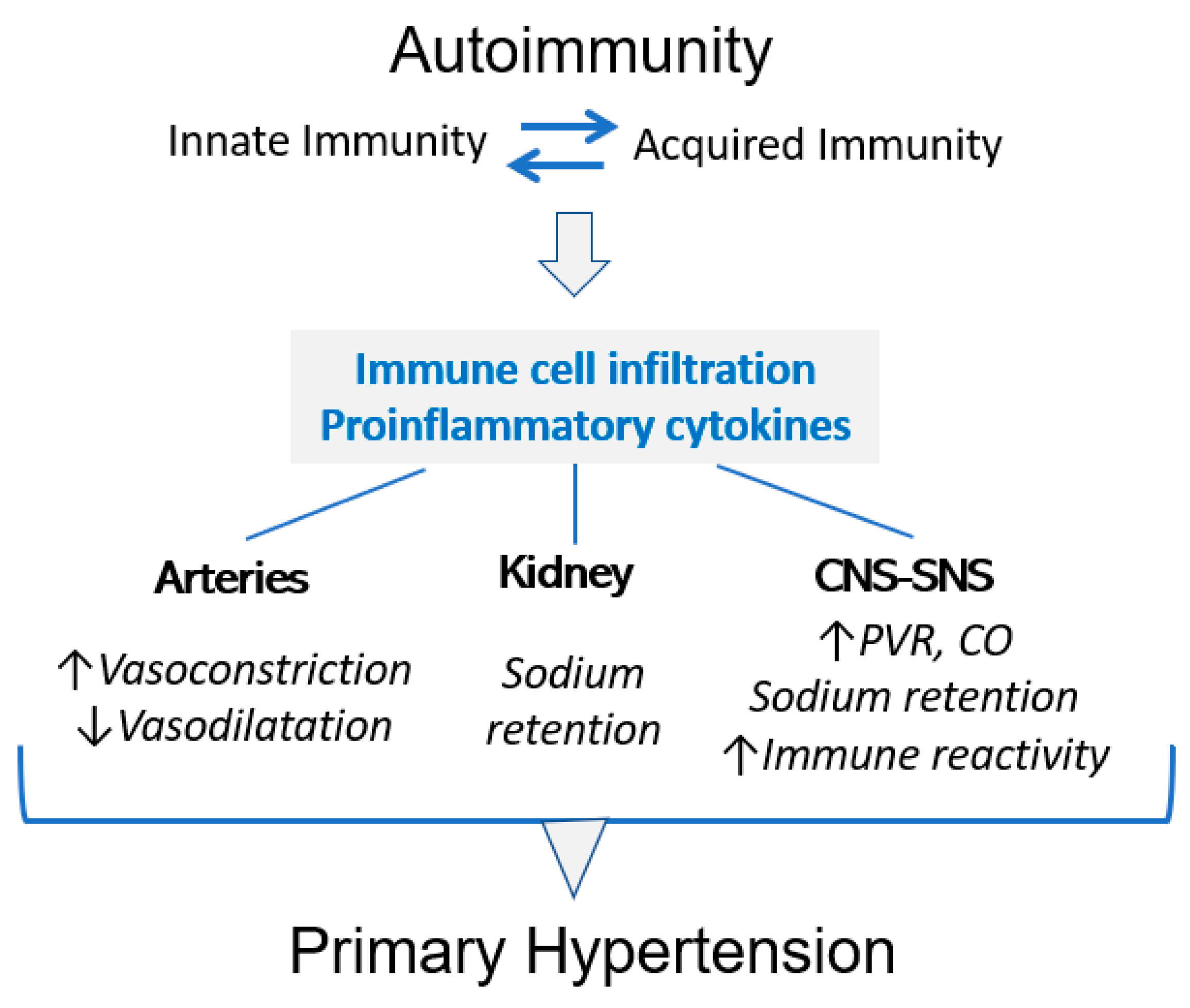
2. HSP70, Inflammation and Immune Reactivity
3. HSP70 and High Blood Pressure
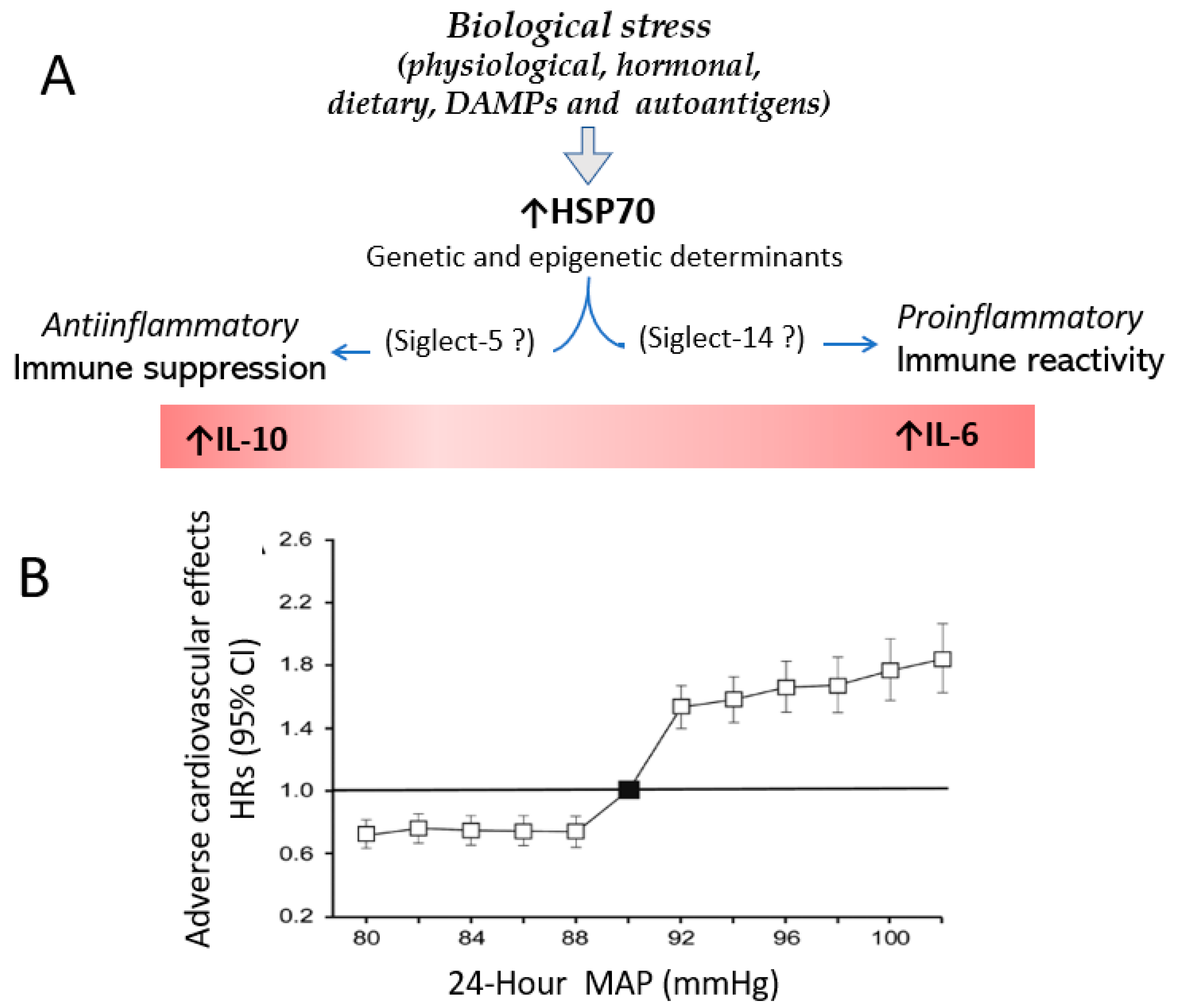
4. Stress, HSP70 and Hypertension
5. Anti-HSP70 Antibodies in Primary Hypertension
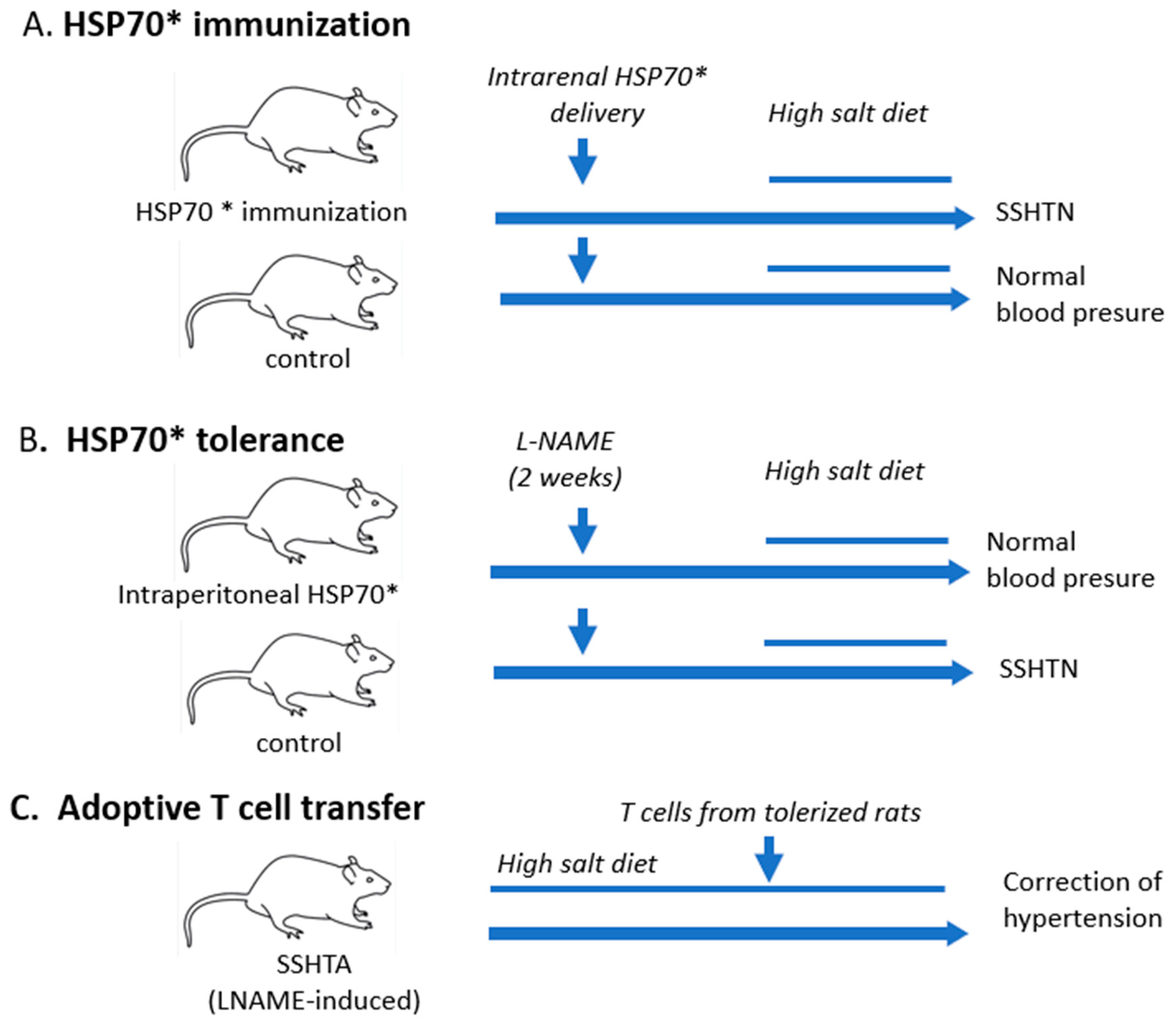
6. Hypertension Is Induced by Immune Reactivity to HSP70
7. Immune Tolerance to HSP70 Corrects Hypertension
8. HSP70-Induced Immune Reactivity Modulates Inflammation in Hypertension
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welch, W.J. Mammalian stress response: Cell physiology, structure/function of stress proteins, and implications for medicine and disease. Physiol. Rev. 1992, 72, 1063–1081. [Google Scholar] [CrossRef]
- Kurop, M.K.; Huyen, C.M.; Kelly, J.H.; Blagg, B.S. The heat shock response and small molecule regulators. Eur. J. Med. Chem. 2021, 226, 113846. [Google Scholar] [CrossRef] [PubMed]
- Beere, H.M. The stress of dying’: The role of heat shock proteins in the regulation of apoptosis. J. Cell Sci. 2004, 117, 1641–1651. [Google Scholar] [CrossRef]
- Lang, B.J.; Guerrero, M.E.; Prince, T.L.; Okusha, Y.; Bonorino, C.; Calderwood, S.K. The functions and regulation of heat shock proteins; key orchestrators of proteostasis and the heat shock response. Arch. Toxicol. 2021, 95, 1943–1970. [Google Scholar] [CrossRef] [PubMed]
- Bolhassani, A.; Agi, E. Heat shock proteins in infection. Clin. Chim. Acta 2019, 498, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Redlich, K.; Xu, Q.; Bizan, P.; Gröger, M.; Tohidast-Akrad, M.; Kiener, H.; Smolen, J.; Steiner, G. Enhanced expression of heat shock protein 70 (hsp70) and heat shock factor 1 (HSF1) activation in rheumatoid arthritis synovial tissue. Differential regulation of hsp70 expression and hsf1 activation in synovial fibroblasts by proinflammatory cytokines, shear stress, and antiinflammatory drugs. J. Clin. Investig. 1998, 102, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Bertorelli, G.; Bocchino, V.; Zhuo, X.; Chetta, A.; Donno, D.; Foresi, A.; Testi, R.; Olivieri, D. Heat shock protein 70 upregulation is related to HLA-DR expression in bronchial asthma. Effects of inhaled glucocorticoids. Clin. Exp. Allergy 1998, 28, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, T.; Rios, Z.; Mei, Q.; Lin, X.; Cao, S. Heat Shock Proteins and Cancer. Trends Pharmacol. Sci. 2017, 38, 226–256. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Radons, J. The human HSP70 family of chaperones: Where do we stand? Cell Stress Chaperones 2016, 21, 379–404. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liang, C.; Zhou, L. Structural and functional analysis of the Hsp70/Hsp40 chaperone system. Protein Sci. 2020, 29, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Luengo, T.M.; Mayer, M.P.; Rüdiger, S.G. The Hsp70–Hsp90 Chaperone Cascade in Protein Folding. Trends Cell Biol. 2019, 29, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Radli, M.; Rüdoger, S.G.D. Dancing with the Diva: Hsp90–Client Interactions. J. Mol Biol. 2018, 430, 3029–3040. [Google Scholar] [CrossRef]
- Chen, Y.; Voegeli, T.S.; Liu, P.P.; Noble, E.G.; Currie, R.W. Heat Shock Paradox and a New Role of Heat Shock Proteins and their Receptors as Anti-Inflammation Targets. Inflamm. Allergy-Drug Targets 2007, 6, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Martine, P.; Rébé, C. Heat Shock Proteins and Inflammasomes. Int. J. Mol. Sci. 2019, 20, 4508. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Gong, J.; Murshid, A. Extracellular HSPs: The Complicated Roles of Extracellular HSPs in Immunity. Front. Immunol. 2016, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Stocki, P.; Wang, X.N.; Dickinson, A.M. Inducible Heat Shock Protein 70 Reduces T Cell Responses and Stimulatory Capacity of Monocyte-derived Dendritic Cells. J. Biol. Chem. 2012, 287, 12387–12394. [Google Scholar] [CrossRef]
- Martine, P.; Chevriaux, A.; Derangère, V.; Apetoh, L.; Garrido, C.; Ghiringhelli, F.; Rébé, C. HSP70 is a negative regulator of NLRP3 inflammasome activation. Cell Death Dis. 2019, 10, 256. [Google Scholar] [CrossRef]
- Borges, T.J.; Wieten, L.; van Herwijnen, M.J.C.; Broere, F.; van der Zee, R.; Bonorino, C.; van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef]
- Borges, T.J.; Porto, B.N.; Teixeira, C.A.; Rodrigues, M.; Machado, F.D.; Ornaghi, A.P.; de Souza, A.P.D.; Maito, F.; Pavanelli, W.R.; Silva, J.S.; et al. Prolonged Survival of Allografts Induced by Mycobacterial Hsp70 Is Dependent on CD4+CD25+ Regulatory T Cells. PLoS ONE 2010, 5, e14264. [Google Scholar] [CrossRef] [Green Version]
- Van Eden, W.; Tholet, J.E.R.; Van Der Zee, R.; Noordzij, A.; Van Embden, J.D.A.; Hensen, E.J.; Cohen, I.R. Cloning of the mycobacterial epitope recognized by T lymphocytes in adjuvant arthritis. Nature 1988, 331, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Anderton, S.M.; van der Zee, R.; Noordzij, A.; van Eden, W. Differential mycobacterial 65-kDa heat shock protein T cell epitope recognition after adjuvant arthritis-inducing or protective immunization protocols. J. Immunol. 1994, 152, 3656–3664. [Google Scholar] [CrossRef] [PubMed]
- van Eden, W.; van der Zee, R.; Prakken, B. Heat-shock proteins induce T-cell regulation of chronic inflammation. Nat. Rev. Immunol. 2005, 5, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Asea, A.; Kraeft, S.-K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Gao, B.; Tsan, M.-F. Endotoxin Contamination in Recombinant Human Heat Shock Protein 70 (Hsp70) Preparation Is Responsible for the Induction of Tumor Necrosis Factor α Release by Murine Macrophages. J. Biol. Chem. 2003, 278, 174–179. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, B.; Lanaspa, M.A.; Johnson, R.J. The role of autoimmune reactivity induced by heat shock protein 70 in the pathogenesis of essential hypertension. Br. J. Pharmacol. 2019, 176, 1829–1838. [Google Scholar] [CrossRef]
- Millar, D.; Garza, K.M.; Odermatt, B.; Elford, A.R.; Ono, N.; Li, Z.; Ohashi, P.S. Hsp70 promotes antigen-presenting cell function and converts T-cell tolerance to autoimmunity in vivo. Nat. Med. 2003, 9, 1469–1476. [Google Scholar] [CrossRef]
- Kolb, H.; Burkart, V. Chaperones may cause the focus of diabetes autoimmunity on distinct (pro)insulin peptides. J. Autoimmun. 2019, 105, 102304. [Google Scholar] [CrossRef]
- Androvitsanea, A.; Stylianou, K.; Drosataki, E.; Petrakis, I. The Pathophysiological Role of Heat Shock Response in Autoimmunity: A Literature Review. Cells 2021, 10, 2626. [Google Scholar] [CrossRef]
- Fong, J.J.; Sreedhara, K.; Deng, L.; Varki, N.M.; Angata, T.; Liu, Q.; Nizet, V.; Varki, A. Immunomodulatory activity of extracellular Hsp70 mediated via paired receptors Siglec-5 and Siglec-14. EMBO J. 2015, 34, 2775–2788. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, B.; Pons, H.; Johnson, R.J. Role of the Immune System in Hypertension. Physiol. Rev. 2017, 97, 1127–1164. [Google Scholar] [CrossRef]
- Trott, D.W.; Thabet, S.R.; Kirabo, A.; Saleh, M.A.; Itani, H.; Norlander, A.E.; Wu, J.; Goldstein, A.; Arendshorst, W.J.; Madhur, M.S.; et al. Oligoclonal cd8+ t cells play a critical role in the development of hypertension. Hypertension 2014, 64, 1108–1115. [Google Scholar] [CrossRef]
- Abais-Battad, J.M.; Rudemiller, N.P.; Mattson, D. L Hypertension and immunity: Mechanisms of T cell activation and pathways of hypertension. Curr. Opin. Nephrol. Hypertens. 2015, 24, 470–474. [Google Scholar] [CrossRef]
- Mikolajczyk, T.P.; Szczepaniak, P.; Vidler, F.; Maffia, P.; Graham, G.J.; Guzik, T.J. Role of inflammatory chemokines in hypertension. Pharmacol. Ther. 2021, 223, 107799. [Google Scholar] [CrossRef]
- Guzik, T.J.; Hoch, N.E.; Brown, K.A.; McCann, L.A.; Rahman, A.; Dikalov, S.; Goronzy, J.; Weyand, C.; Harrison, D.G. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J. Exp. Med. 2007, 204, 2449–2460. [Google Scholar] [CrossRef]
- Kirabo, A.; Fontana, V.; de Faria, A.P.; Loperena, R.; Galindo, C.L.; Wu, J.; Bikineyeva, A.T.; Dikalov, S.; Xiao, L.; Chen, W.; et al. DC isoketal-modified proteins activate T cells and promote hypertension. J. Clin. Investig. 2014, 124, 4642–4656. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Iturbe, B.; Pons, H.; Quiroz, Y.; Johnson, R.J. The immunological basis of hypertension. Am. J. Hypertens. 2019, 32, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Rose, G. Seasonal Variation in Blood Pressure in Man. Nature 1961, 189, 235. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Ogihara, T.; Maruyama, A.; Mikami, H.; Nakamaru, M.; Naka, T.; Kumahara, Y.; Nugent, C.A. The Seasonal Variation of Blood Pressure in Patients with Essential Hypertension. Clin. Exp. Hypertens. A 1982, 4, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Kunes, J.; Tremblay, J.; Bellavance, F.; Hamet, P. Influence of Environmental Temperature on the Blood Pressure of Hypertensive Patients in Montreal. Am. J. Hypertens. 1991, 4, 422–426. [Google Scholar] [CrossRef] [PubMed]
- McMurtry, J.P.; Wexler, B.C. Hypersensitivity of Spontaneously Hypertensive Rats to Heat and Ether before the Onset of High Blood Pressure. Endocrinology 1983, 112, 166–171. [Google Scholar] [CrossRef]
- Hamet, P.; Malo, D.; Tremblay, J. Increased transcription of a major stress gene in spontaneously hypertensive mice. Hypertension 1990, 15, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Kunes, J.; Poirier, M.; Tremblay, J.; Hamet, P. Expression of hsp70 gene in lymphocytes from normotensive and hypertensive Humans. Acta Physiol. Scand. 1992, 146, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Hamet, P.; Kong, D.; Pravenec, M.; Kunes, J.; Kren, V.; Klir, P.; Sun, Y.L.; Tremblay, J. Restriction fragment length polymorphism of hsp70 gene, localized in the RT1 complex, is associated with hypertension in spontaneously hypertensive rats. Hypertension 1992, 19, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ross, B.M.; Currie, R.W. Heat shock treatment protects against angiotensin II–induced hypertension and inflammation in aorta. Cell Stress Chaperones 2004, 9, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, D.-G.; Holbrook, N.J.; Udelsman, R. Acute Hypertension Induces Heat-Shock Protein 70 Gene Expression in Rat Aorta. Circulation 1995, 92, 1223–1229. [Google Scholar] [CrossRef]
- Kvetnansky, R.; Mccarty, R.; Thoa, N.B.; Lake, C.R.; Kopin, I.J. Sympatho-adrenal responses of spontaneously hypertensive rats to immobilization stress. Am. J. Physiol. 1979, 236, H457–H462. [Google Scholar] [CrossRef]
- Malo, D.; Pang, S.C.; Schlager, G.; Tremblay, J.; Hamet, P. Decrease of Blood Pressure in Spontaneously Hypertensive Mice by Heat Treatment. Am. J. Hypertens. 1990, 3, 400–404. [Google Scholar] [CrossRef]
- Bravo, J.; Quiroz, Y.; Pons, H.; Parra, G.; Herrera-Acosta, J.; Johnson, R.J.; Rodríguez-Iturbe, B. Vimentin and heat shock protein expression are induced in the kidney by angiotensin and by nitric oxide inhibition. Kidney Int. 2003, 86, S46–S51. [Google Scholar] [CrossRef]
- Wu, T.; Ma, J.; Chen, S.; Sun, Y.; Xiao, C.; Gao, Y.; Wang, R.; Poudrier, J.; Dargis, M.; Currie, W.; et al. Association of plasma antibodies against the inducible Hsp70 with hypertension and harsh working conditions. Cell Stress Chaperones 2001, 6, 394. [Google Scholar] [CrossRef]
- Srivastava, K.; Narang, R.; Bhatia, J.; Saluja, D. Expression of Heat Shock Protein 70 Gene and Its Correlation with Inflammatory Markers in Essential Hypertension. PLoS ONE 2016, 11, e0151060. [Google Scholar] [CrossRef]
- Pockley, A.; De Faire, U.; Kiessling, R.; Lemne, C.; Thulin, T.; Frostegard, J. Circulating heat shock protein and heat shock protein antibody levels in established hypertension. J. Hypertens. 2002, 20, 1815–1820. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, B.; Johnson, R.J. The role of renal microvascular disease and interstitial inflammation in salt-sensitive hypertension. Hypertens. Res. 2010, 33, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Tang, B.-P.; Sun, H.-P.; Feng, M.; Cheng, Z.-H.; Niu, W.-Q. Interacting contribution of the five polymorphisms in three genes of Hsp70 family to essential hypertension in Uygur ethnicity. Cell Stress Chaperones 2009, 14, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Ping, J.P.; Lyu., Y.; Li, R.; Tian, F.-J.; Mu, J.-B. Occup. Environ. Med. Epub ahead of print. [CrossRef]
- Barhoumi, T.; Kasal, D.A.; Li, M.W.; Shbat, L.; Laurant, P.; Neves, M.F.; Paradis, P.; Schiffrin, E.L. T Regulatory Lymphocytes Prevent Angiotensin II–Induced Hypertension and Vascular Injury. Hypertension 2011, 57, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Bueno, M.; Ellison, D.H. Blood pressure effects of sodium transport along the distal nephron. Kidney Int. 2022, 102, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, D.; Pallante, F.; Fardella, V.; Fardella, S.; Iacobucci, R.; Federici, M.; Cifelli, G.; De Lucia, M.; Lembo, G. The Angiogenic Factor PlGF Mediates a Neuroimmune Interaction in the Spleen to Allow the Onset of Hypertension. Immunity 2014, 41, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Pons, H.; Ferrebuz, A.; Quiroz, Y.; Romero-Vasquez, F.; Parra, G.; Johnson, R.J.; Rodriguez-Iturbe, B. Immune reactivity to heat shock protein 70 expressed in the kidney is cause of salt-sensitive hypertension. Am. J. Physiol. Physiol. 2013, 304, F289–F299. [Google Scholar] [CrossRef]
- Quiroz, Y.; Pons, H.; Gordon, K.L.; Rincón, J.; Chávez, M.; Parra, G.; Herrera-Acosta, J.; Gómez-Garre, D.; Largo, R.; Egido, J.; et al. Mycophenolate mofetil prevents salt-sensitive hypertension resulting from nitric oxide synthesis inhibition. Am. J. Physiol. Physiol. 2001, 281, F38–F47. [Google Scholar] [CrossRef] [Green Version]
- Wendling, U.; Paul, L.; van der Zee, R.; Prakken, B.; Singh, M.; van Eden, W. A Conserved Mycobacterial Heat Shock Protein (hsp) 70 Sequence Prevents Adjuvant Arthritis upon Nasal Administration and Induces IL-10-Producing T Cells That Cross-React with the Mammalian Self-hsp70 Homologue. J. Immunol. 2000, 164, 2711–2717. [Google Scholar] [CrossRef]
- Prakken, B.J.; Wendling, U.; van der Zee, R.; Rutten, V.P.; Kuis, W.; Van Eden, W. Induction of IL-10 and Inhibition of Experimental Arthritis Are Specific Features of Microbial Heat Shock Proteins That Are Absent for Other Evolutionarily Conserved Immunodominant Proteins. J. Immunol. 2001, 167, 4147–4153. [Google Scholar] [CrossRef]
- Oldham, P.; Pickering, G.; Roberts, J.F.; Sowry, G. The nature of essential hypertension. Lancet 1960, 1, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Pater, C. The Blood Pressure “Uncertainty Range”—A pragmatic approach to overcome current diagnostic uncertainties (II). Curr. Control. Trials Cardiovasc. Med. 2005, 6, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Ostchega, Y.; Fryar, C.D.; Nwankwo, T.; Nguyen, D.T. Hypertension Prevalence among Adults Aged 18 and Over: United States, 2017–2018; NCHS Data Brief, No. 364; National Center for Health Statistics: Hyattsville, MD, USA, 2020. Available online: https://www.cdc.gov/nchs/products/in,dex.htm (accessed on 16 December 2022).
- Melgarejo, J.D.; Yang, W.Y.; Thijs, L.; Li, Y.; Asayama, K.; Hansen, T.W.; Wei, F.-F.; Kikuya, M.; Ohkubo, T.; Dolan, E.; et al. For the International Database on Ambulatory Blood Pressure in Relation to Cardiovascular Outcome Investigators. Association of Fatal and Nonfatal Cardiovascular Outcomes with 24-Hour Mean Arterial Pressure. Hypertension 2021, 77, 39–48. [Google Scholar] [CrossRef]
- Ban, H.S.; Han, T.-S.; Hur, K.; Cho, H.-S. Epigenetic Alterations of Heat Shock Proteins (HSPs) in Cancer. Int. J. Mol. Sci. 2019, 20, 4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Experimental/Clinical | Results | References |
|---|---|---|---|
| HSP70 expression | Salt-sensitive hypertension | Overexpression in proximal tubular cells of the kidney | [49] |
| HSP70 expression in relation to development of hypertension | Spontaneously hypertensive rats | HSP72 overexpression precedes hypertension | [31] |
| Working conditions and anti-HSP antibodies | Harsh working conditions | Anti-HSP70 antibodies in 40.2% of hypertensives and 18.6% of normotensive individuals | [50] |
| Heat-induced HSP70 mRNA in lymphocytes | 15 min at 42 °C in peripheral blood lymphocytes | Increased HSP70 mRNA in lymphocytes from hypertensive patients | [43] |
| Constitutive HSP70 mRNA and expression and inflammation | 132 hypertensives and 132 normotensive individuals | 6-fold increase in HSP70 mRNA in hypertensives. Positive correlation with IL6 and TNFα levels | [51] |
| Constitutive anti-HSP antibodies | Serum samples from European Lacidipine Study on Atherosclerosis (ELSA) and from Swedish population-screening programme | Anti-HSP70 and anti-HSP65 levels increased in hypertensive patients | [52,53] |
| Peripheral blood lymphocytes challenged with a mycobacterial HSP70 peptide sequence | 10 patients with primary hypertension, 12 normotensive patients | Proliferative response (x2 proliferation index) in patients with primary hypertension | [59] |
| Genetic HSP70 polymorphisms | Uygur ethnicity | 5 polymorphisms of HSP70 associated with increased risk of hypertension | [54] |
| HSP70 polymorphisms and working conditions | Coke oven workers | Increased hypertension risk: HSP70-1 GC, CC and C+CCgenotype Reduced hypertension risk: HSP70-2 AG, GG and AG+GGgenotype | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Iturbe, B.; Johnson, R.J.; Sanchez-Lozada, L.G.; Pons, H. HSP70 and Primary Arterial Hypertension. Biomolecules 2023, 13, 272. https://doi.org/10.3390/biom13020272
Rodriguez-Iturbe B, Johnson RJ, Sanchez-Lozada LG, Pons H. HSP70 and Primary Arterial Hypertension. Biomolecules. 2023; 13(2):272. https://doi.org/10.3390/biom13020272
Chicago/Turabian StyleRodriguez-Iturbe, Bernardo, Richard J. Johnson, Laura Gabriela Sanchez-Lozada, and Hector Pons. 2023. "HSP70 and Primary Arterial Hypertension" Biomolecules 13, no. 2: 272. https://doi.org/10.3390/biom13020272