
Oligodendrocytes Prune Axons Containing α-Synuclein Aggregates In Vivo: Lewy Neurites as Precursors of Glial Cytoplasmic Inclusions in Multiple System Atrophy?
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. α-Syn Expression
2.2. α-Syn Purification
2.3. α-Syn Fibrillization
2.4. Sonication
2.5. In Vivo α-Syn Pathology and Histology
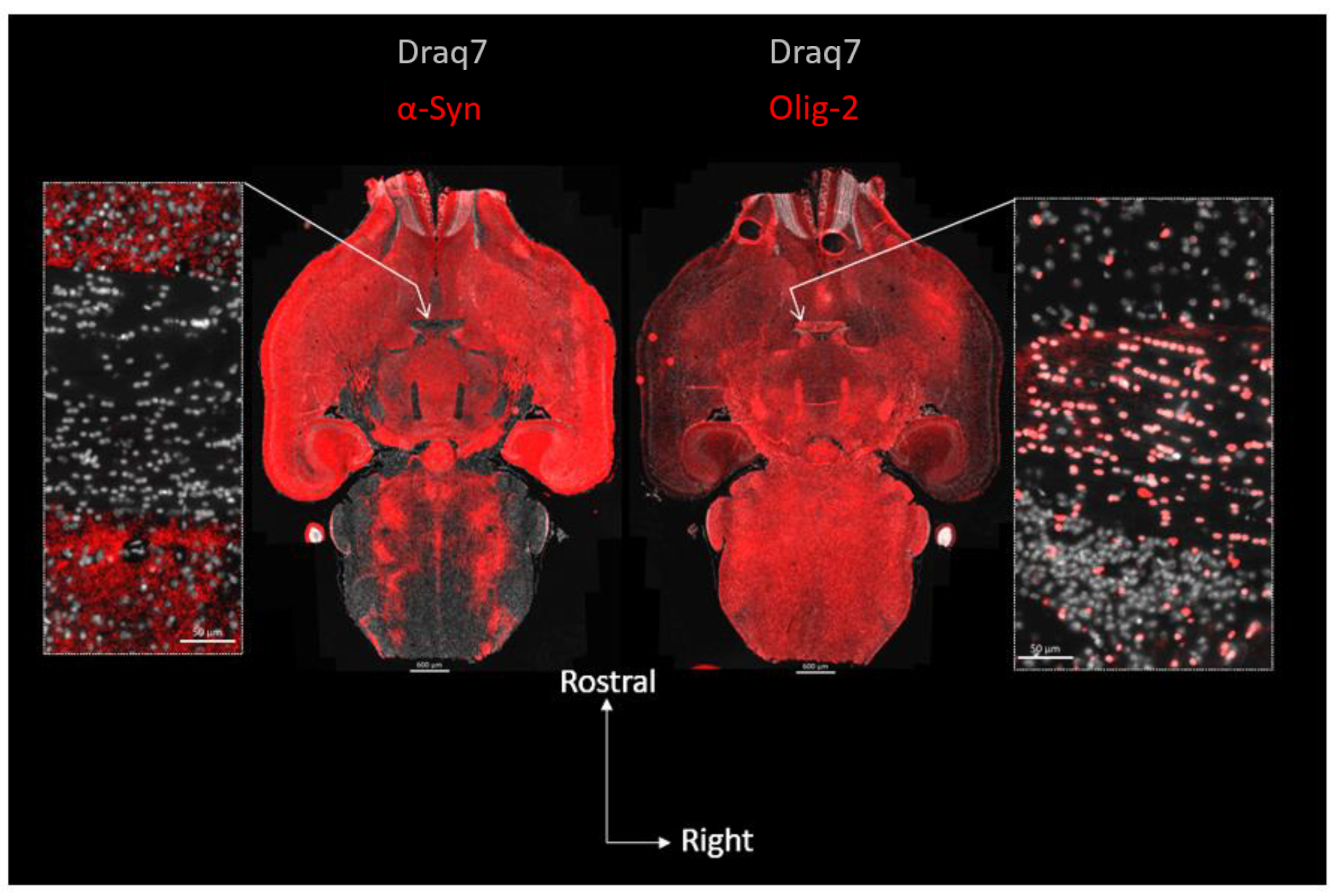
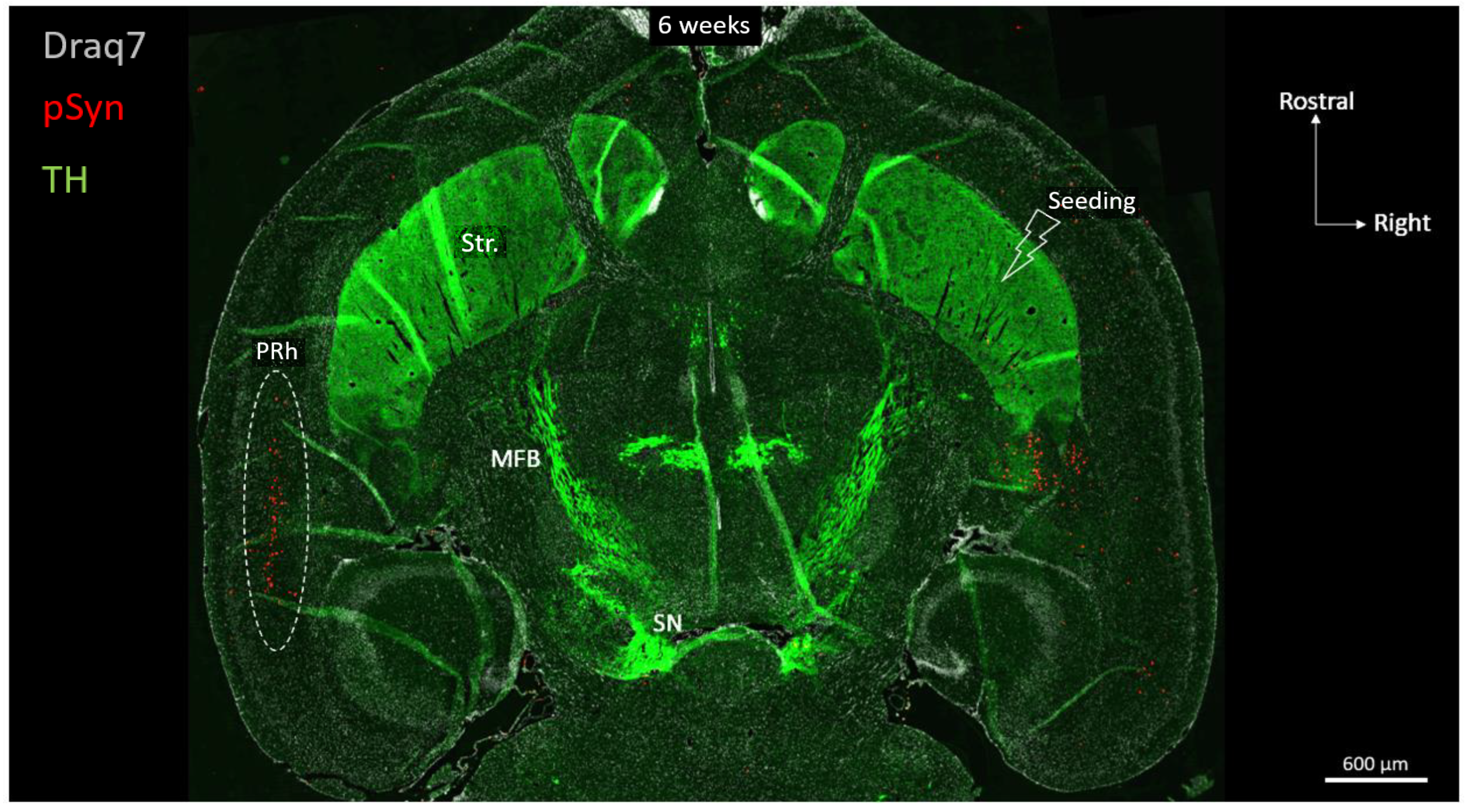
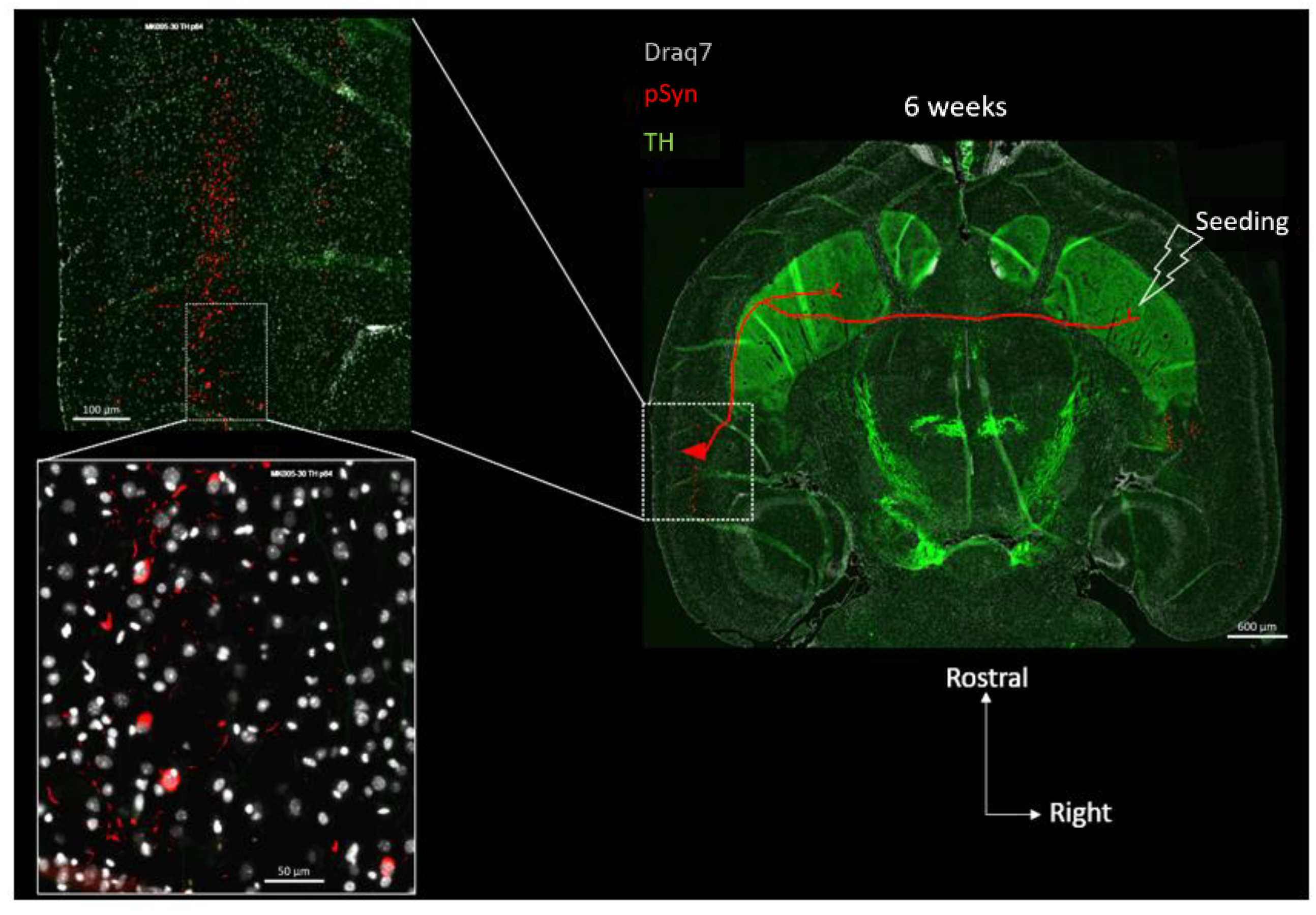
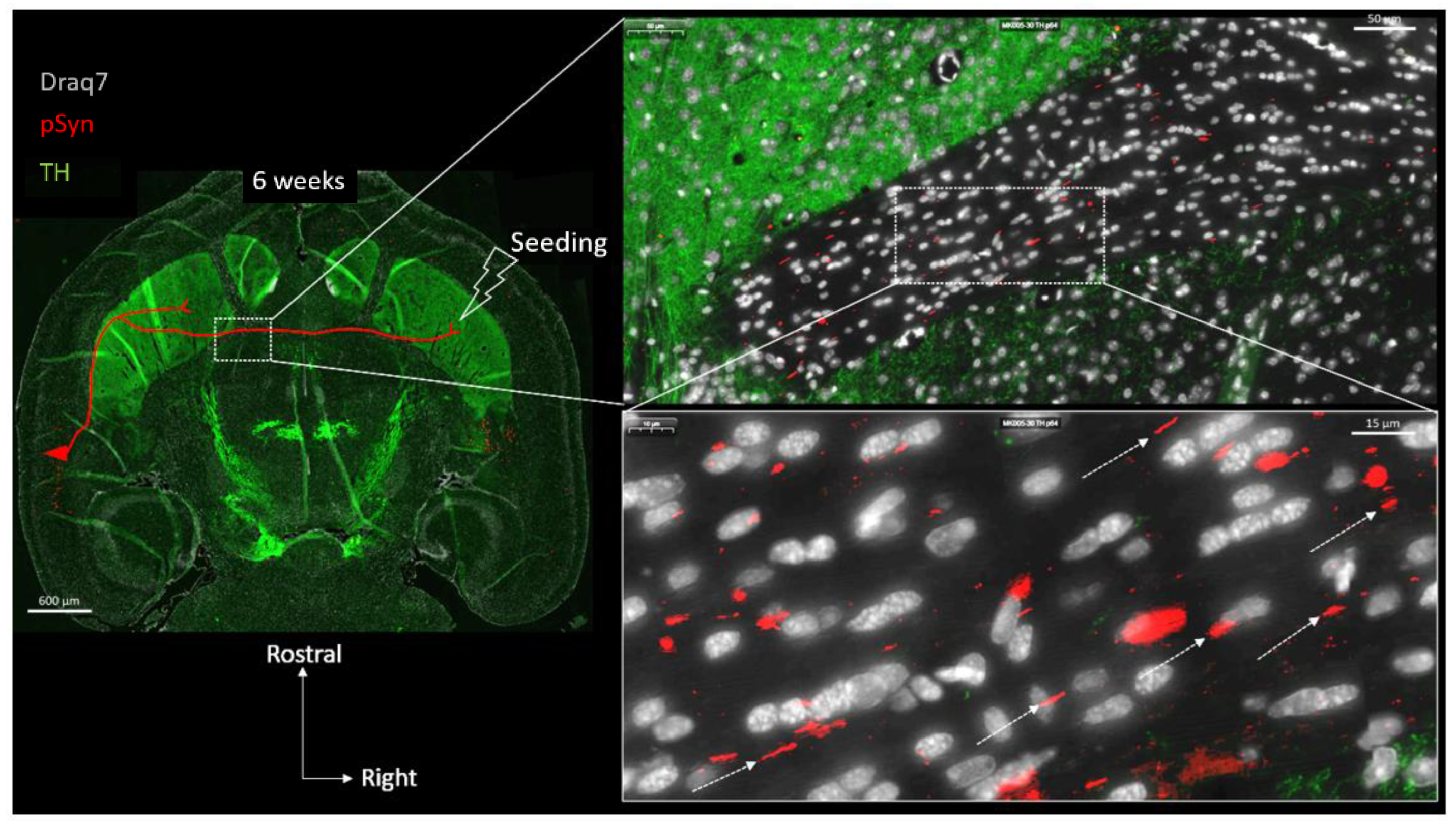
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Giorgi, F.; Uversky, V.N.; Ichas, F. α-Synuclein Fibrils as Penrose Machines: A Chameleon in the Gear. Biomolecules 2022, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.H.; Galvin, J.E.; Baba, M.; Giasson, B.; Tomita, T.; Leight, S.; Nakajo, S.; Iwatsubo, T.; Trojanowski, J.Q.; Lee, V.M.Y. Glial Cytoplasmic Inclusions in White Matter Oligodendrocytes of Multiple System Atrophy Brains Contain Insoluble α-Synuclein. Ann. Neurol. 1998, 44, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Lippa, C.F.; Duda, J.E.; Grossman, M.; Hurtig, H.I.; Aarsland, D.; Boeve, B.F.; Brooks, D.J.; Dickson, D.W.; Dubois, B.; Emre, M.; et al. DLB and PDD Boundary Issues: Diagnosis, Treatment, Molecular Pathology, and Biomarkers. Neurology 2007, 68, 812–819. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Goedert, M. The α-Synucleinopathies: Parkinson’s Disease, Dementia with Lewy Bodies, and Multiple System Atrophy. Ann. N. Y. Acad. Sci. 2006, 920, 16–27. [Google Scholar] [CrossRef] [PubMed]
- McKeith, I.G.; Dickson, D.W.; Lowe, J.; Emre, M.; O’Brien, J.T.; Feldman, H.; Cummings, J.; Duda, J.E.; Lippa, C.; Perry, E.K.; et al. Diagnosis and Management of Dementia with Lewy Bodies: Third Report of the DLB Consortium. Neurology 2005, 65, 1863–1872. [Google Scholar] [CrossRef] [PubMed]
- Krismer, F.; Wenning, G.K. Multiple System Atrophy: Insights into a Rare and Debilitating Movement Disorder. Nat. Rev. Neurol. 2017, 13, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.L.; Delucia, M.W.; Dickson, D.W. α-Synuclein Immunoreactivity in Neuronal Nuclear Inclusions and Neurites in Multiple System Atrophy. Neurosci. Lett. 2004, 354, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Nishie, M.; Mori, F.; Yoshimoto, M.; Takahashi, H.; Wakabayashi, K. A Quantitative Investigation of Neuronal Cytoplasmic and Intranuclear Inclusions in the Pontine and Inferior Olivary Nuclei in Multiple System Atrophy. Neuropathol. Appl. Neurobiol. 2004, 30, 546–554. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Takahashi, H. Cellular pathology in multiple system atrophy. Neuropathology 2006, 26, 338–345. [Google Scholar] [CrossRef]
- Ebrahimi-Fakhari, D.; Wahlster, L.; McLean, P.J. Protein Degradation Pathways in Parkinson’s Disease: Curse or Blessing. Acta Neuropathol. 2012, 124, 153–172. [Google Scholar] [CrossRef] [Green Version]
- Lee, V.M.Y.; Trojanowski, J.Q. Mechanisms of Parkinson’s Disease Linked to Pathological α-Synuclein: New Targets for Drug Discovery. Neuron 2006, 52, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Kahle, P.J.; Neumann, M.; Ozmen, L.; Müller, V.; Jacobsen, H.; Spooren, W.; Fuss, B.; Mallon, B.; Macklin, W.B.; Fujiwara, H.; et al. Hyperphosphorylation and Insolubility of α-Synuclein in Transgenic Mouse Oligodendrocytes. EMBO Rep. 2002, 3, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Shults, C.W.; Rockenstein, E.; Crews, L.; Adame, A.; Mante, M.; Larrea, G.; Hashimoto, M.; Song, D.; Iwatsubo, T.; Tsuboi, K.; et al. Neurological and Neurodegenerative Alterations in a Transgenic Mouse Model Expressing Human α-Synuclein under Oligodendrocyte Promoter: Implications for Multiple System Atrophy. J. Neurosci. 2005, 25, 10689–10699. [Google Scholar] [CrossRef]
- Yazawa, I.; Giasson, B.I.; Sasaki, R.; Zhang, B.; Joyce, S.; Uryu, K.; Trojanowski, J.Q.; Lee, V.M.Y. Mouse Model of Multiple System Atrophy α-Synuclein Expression in Oligodendrocytes Causes Glial and Neuronal Degeneration. Neuron 2005, 45, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Bleasel, J.M.; Halliday, G.M.; Kim, W.S. Animal Modeling an Oligodendrogliopathy–Multiple System Atrophy. Acta Neuropathol. Commun. 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Laferrière, F.; He, X.; Zinghirino, F.; Doudnikoff, E.; Faggiani, E.; Meissner, W.G.; Bezard, E.; de Giorgi, F.; Ichas, F. Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy. Cells 2020, 9, 2371. [Google Scholar] [CrossRef] [PubMed]
- Asi, Y.T.; Simpson, J.E.; Heath, P.R.; Wharton, S.B.; Lees, A.J.; Revesz, T.; Houlden, H.; Holton, J.L. Alpha-Synuclein MRNA Expression in Oligodendrocytes in MSA. Glia 2014, 62, 964–970. [Google Scholar] [CrossRef]
- Miller, D.W.; Johnson, J.M.; Solano, S.M.; Hollingsworth, Z.R.; Standaert, D.G.; Young, A.B. Absence of α-Synuclein mRNA Expression in Normal and Multiple System Atrophy Oligodendroglia. J. Neural Transm. 2005, 112, 1613–1624. [Google Scholar] [CrossRef]
- Langerveld, A.J.; Mihalko, D.; DeLong, C.; Walburn, J.; Ide, C.F. Gene Expression Changes in Postmortem Tissue from the Rostral Pons of Multiple System Atrophy Patients. Mov. Disord. 2007, 22, 766–777. [Google Scholar] [CrossRef]
- de Giorgi, F.; Laferrière, F.; Zinghirino, F.; Faggiani, E.; Lends, A.; Bertoni, M.; Yu, X.; Grélard, A.; Morvan, E.; Habenstein, B.; et al. Novel Self-Replicating α-Synuclein Polymorphs That Escape ThT Monitoring Can Spontaneously Emerge and Acutely Spread in Neurons. Sci. Adv. 2020, 6, eabc4364. [Google Scholar] [CrossRef]
- Trojanowski, J.; Walkenstein, N.; Lee, V. Expression of Neurofilament Subunits in Neurons of the Central and Peripheral Nervous System: An Immunohistochemical Study with Monoclonal Antibodies. J. Neurosci. 1986, 6, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Bin, J.M.; Harris, S.N.; Kennedy, T.E. The Oligodendrocyte-Specific Antibody “CC1” Binds Quaking 7. J. Neurochem. 2016, 139, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Kanaan, N.M.; Grabinski, T. Neuronal and Glial Distribution of Tau Protein in the Adult Rat and Monkey. Front. Mol. Neurosci. 2021, 14, 607303. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Misonou, H.; Matsuyama, M.; Nomori, A.; Wada-Kakuda, S.; Takashima, A.; Kawata, M.; Murayama, S.; Ihara, Y.; Miyasaka, T. Distribution of Endogenous Normal Tau in the Mouse Brain. J. Comp. Neurol. 2019, 527, 985–998. [Google Scholar] [CrossRef] [PubMed]
- de Chaumont, F.; Dallongeville, S.; Chenouard, N.; Hervé, N.; Pop, S.; Provoost, T.; Meas-Yedid, V.; Pankajakshan, P.; Lecomte, T.; le Montagner, Y.; et al. Icy: An Open Bioimage Informatics Platform for Extended Reproducible Research. Nat. Methods 2012, 9, 690–696. [Google Scholar] [CrossRef]
- Peelaerts, W.; Bousset, L.; van der Perren, A.; Moskalyuk, A.; Pulizzi, R.; Giugliano, M.; van den Haute, C.; Melki, R.; Baekelandt, V. α-Synuclein Strains Cause Distinct Synucleinopathies after Local and Systemic Administration. Nature 2015, 522, 340–344. [Google Scholar] [CrossRef]
- Reddy, K.; Dieriks, B.V. Multiple System Atrophy: α-Synuclein Strains at the Neuron-Oligodendrocyte Crossroad. Mol. Neurodegener. 2022, 17, 77. [Google Scholar] [CrossRef]
- Djelloul, M.; Holmqvist, S.; Boza-Serrano, A.; Azevedo, C.; Yeung, M.S.; Goldwurm, S.; Frisén, J.; Deierborg, T.; Roybon, L. Alpha-Synuclein Expression in the Oligodendrocyte Lineage: An in Vitro and in Vivo Study Using Rodent and Human Models. Stem Cell Rep. 2015, 5, 174–184. [Google Scholar] [CrossRef]
- Laferrière, F.; Claverol, S.; Bezard, E.; de Giorgi, F.; Ichas, F. Similar Neuronal Imprint and No Cross-Seeded Fibrils in α-Synuclein Aggregates from MSA and Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 10. [Google Scholar] [CrossRef]
- Uemura, N.; Uemura, M.T.; Lo, A.; Bassil, F.; Zhang, B.; Luk, K.C.; Lee, V.M.Y.; Takahashi, R.; Trojanowski, J.Q. Slow Progressive Accumulation of Oligodendroglial Alpha-Synuclein (a-Syn) Pathology in Synthetic a-Syn Fibril-Induced Mouse Models of Synucleinopathy. J. Neuropathol. Exp. Neurol. 2019, 78, 877–890. [Google Scholar] [CrossRef]
- Luk, K.C.; Covell, D.J.; Kehm, V.M.; Zhang, B.; Song, I.Y.; Byrne, M.D.; Pitkin, R.M.; Decker, S.C.; Trojanowski, J.Q.; Lee, V.M.Y. Molecular and Biological Compatibility with Host Alpha-Synuclein Influences Fibril Pathogenicity. Cell Rep. 2016, 16, 3373–3387. [Google Scholar] [CrossRef] [PubMed]
- Bousset, L.; Pieri, L.; Ruiz-Arlandis, G.; Gath, J.; Jensen, P.H.; Habenstein, B.; Madiona, K.; Olieric, V.; Böckmann, A.; Meier, B.H.; et al. Structural and Functional Characterization of Two Alpha-Synuclein Strains. Nat. Commun. 2013, 4, 2575. [Google Scholar] [CrossRef] [PubMed]
- Shahnawaz, M.; Mukherjee, A.; Pritzkow, S.; Mendez, N.; Rabadia, P.; Liu, X.; Hu, B.; Schmeichel, A.; Singer, W.; Wu, G.; et al. Discriminating α-Synuclein Strains in Parkinson’s Disease and Multiple System Atrophy. Nature 2020, 578, 273–277. [Google Scholar] [CrossRef] [PubMed]
- de Giorgi, F.; Abdul-Shukkoor, M.B.; Kashyrina, M.; Largitte, L.A.; de Nuccio, F.; Kauffmann, B.; Lends, A.; Laferrière, F.; Bonhommeau, S.; Lofrumento, D.D.; et al. Neurons with Cat’s Eyes: A Synthetic Strain of α-Synuclein Fibrils Seeding Neuronal Intranuclear Inclusions. Biomolecules 2022, 12, 436. [Google Scholar] [CrossRef]
- Woulfe, J.M. Abnormalities of the Nucleus and Nuclear Inclusions in Neurodegenerative Disease: A Work in Progress. Neuropathol. Appl. Neurobiol. 2007, 33, 2–42. [Google Scholar] [CrossRef]
- Covell, D.J.; Robinson, J.L.; Akhtar, R.S.; Grossman, M.; Weintraub, D.; Bucklin, H.M.; Pitkin, R.M.; Riddle, D.; Yousef, A.; Trojanowski, J.Q.; et al. Novel Conformation-Selective Alpha-Synuclein Antibodies Raised against Different in Vitro Fibril Forms Show Distinct Patterns of Lewy Pathology in Parkinson’s Disease. Neuropathol. Appl. Neurobiol. 2017, 43, 604–620. [Google Scholar] [CrossRef]
- Luk, K.C.; Kehm, V.; Carroll, J.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M.Y. Pathological α-Synuclein Transmission Initiates Parkinson-like Neurodegeneration in Nontransgenic Mice. Science 2012, 338, 949–953. [Google Scholar] [CrossRef]
- Masuda-Suzukake, M.; Nonaka, T.; Hosokawa, M.; Oikawa, T.; Arai, T.; Akiyama, H.; Mann, D.M.A.; Hasegawa, M. Prion-like Spreading of Pathological α-Synuclein in Brain. Brain 2013, 136, 1128–1138. [Google Scholar] [CrossRef]
- McGeorge, A.J.; Faull, R.L.M. The Organization of the Projection from the Cerebral Cortex to the Striatum in the Rat. Neuroscience 1989, 29, 503–537. [Google Scholar] [CrossRef]
- Mathai, A.; Smith, Y. The Corticostriatal and Corticosubthalamic Pathways: Two Entries, One Target. So What? Front. Syst. Neurosci. 2011, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Suárez, R.; Paolino, A.; Fenlon, L.R.; Morcom, L.R.; Kozulin, P.; Kurniawan, N.D.; Richards, L.J. A Pan-Mammalian Map of Interhemispheric Brain Connections Predates the Evolution of the Corpus Callosum. Proc. Natl. Acad. Sci. USA 2018, 115, 9622–9627. [Google Scholar] [CrossRef] [PubMed]
- Hintiryan, H.; Foster, N.N.; Bowman, I.; Bay, M.; Song, M.Y.; Gou, L.; Yamashita, S.; Bienkowski, M.S.; Zingg, B.; Zhu, M.; et al. The Mouse Cortico-Striatal Projectome. Nat. Neurosci. 2016, 19, 1100–1114. [Google Scholar] [CrossRef] [PubMed]
- Brog, J.S.; Salyapongse, A.; Deutch, A.Y.; Zahm, D.S. The Patterns of Afferent Innervation of the Core and Shell in the “Accumbens” Part of the Rat Ventral Striatum: Immunohistochemical Detection of Retrogradely Transported Fluoro-gold. J. Comp. Neurol. 1993, 338, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Awa, S.; Suzuki, G.; Masuda-Suzukake, M.; Nonaka, T.; Saito, M.; Hasegawa, M. Phosphorylation of Endogenous α-Synuclein Induced by Extracellular Seeds Initiates at the Pre-Synaptic Region and Spreads to the Cell Body. Sci. Rep. 2022, 12, 1163. [Google Scholar] [CrossRef]
- Rahayel, S.; Mišić, B.; Zheng, Y.Q.; Liu, Z.Q.; Abdelgawad, A.; Abbasi, N.; Caputo, A.; Zhang, B.; Lo, A.; Kehm, V.; et al. Differentially Targeted Seeding Reveals Unique Pathological Alpha-Synuclein Propagation Patterns. Brain 2022, 145, 1743–1756. [Google Scholar] [CrossRef]
- Hijaz, B.A.; Volpicelli-Daley, L.A. Initiation and Propagation of α-Synuclein Aggregation in the Nervous System. Mol. Neurodegener. 2020, 15, 19. [Google Scholar] [CrossRef]
- Sturrock, R.R. Age-Related Changes in the Number of Myelinated Axons and Glial Cells in the Anterior and Posterior Limbs of the Mouse Anterior Commissure. J. Anat. 1987, 150, 111. [Google Scholar]
- Bonetto, G.; Belin, D.; Káradóttir, R.T. Myelin: A Gatekeeper of Activity-Dependent Circuit Plasticity? Science 2021, 374, eaba6905. [Google Scholar] [CrossRef]
- Lopresti, P.; Szuchet, S.; Papasozomenos, S.C.; Zinkowski, R.P.; Binder, L.I. Functional Implications for the Microtubule-Associated Protein Tau: Localization in Oligodendrocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 10369–10373. [Google Scholar] [CrossRef]
- Ximerakis, M.; Lipnick, S.L.; Innes, B.T.; Simmons, S.K.; Adiconis, X.; Dionne, D.; Mayweather, B.A.; Nguyen, L.; Niziolek, Z.; Ozek, C.; et al. Single-Cell Transcriptomic Profiling of the Aging Mouse Brain. Nat. Neurosci. 2019, 22, 1696–1708. [Google Scholar] [CrossRef]
- Papp, M.I.; Lantos, P.L. Accumulation of Tubular Structures in Oligodendroglial and Neuronal Cells as the Basic Alteration in Multiple System Atrophy. J. Neurol. Sci. 1992, 107, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Du, J.L.; He, C. Developmental Pruning of Early-Stage Myelin Segments during CNS Myelination in Vivo. Cell Res. 2013, 23, 962–964. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Petrucco, L.; Hoodless, L.J.; Portugues, R.; Czopka, T. Oligodendrocyte Precursor Cells Sculpt the Visual System by Regulating Axonal Remodeling. Nat. Neurosci. 2022, 25, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, J.; Elabbady, L.; Collman, F.; Jorstad, N.L.; Bakken, T.E.; Ott, C.; Glatzer, J.; Bleckert, A.A.; Bodor, A.L.; Brittain, D.; et al. Oligodendrocyte Precursor Cells Ingest Axons in the Mouse Neocortex. Proc. Natl. Acad. Sci. USA 2022, 119, e2202580119. [Google Scholar] [CrossRef]
- Hass, E.W.; Sorrentino, Z.A.; Lloyd, G.M.; McFarland, N.R.; Prokop, S.; Giasson, B.I. Robust α-Synuclein Pathology in Select Brainstem Neuronal Populations Is a Potential Instigator of Multiple System Atrophy. Acta Neuropathol. Commun. 2021, 9, 80. [Google Scholar] [CrossRef]
- Salvesen, L.; Ullerup, B.H.; Sunay, F.B.; Brudek, T.; Løkkegaard, A.; Agander, T.K.; Winge, K.; Pakkenberg, B. Changes in Total Cell Numbers of the Basal Ganglia in Patients with Multiple System Atrophy—A Stereological Study. Neurobiol. Dis. 2015, 74, 104–113. [Google Scholar] [CrossRef]
- Papp, M.I.; Kahn, J.E.; Lantos, P.L. Glial Cytoplasmic Inclusions in the CNS of Patients with Multiple System Atrophy (Striatonigral Degeneration, Olivopontocerebellar Atrophy and Shy-Drager Syndrome). J. Neurol. Sci. 1989, 94, 79–100. [Google Scholar] [CrossRef]
- Prodoehl, J.; Li, H.; Planetta, P.J.; Goetz, C.G.; Shannon, K.M.; Tangonan, R.; Comella, C.L.; Simuni, T.; Zhou, X.J.; Leurgans, S.; et al. Diffusion Tensor Imaging of Parkinson’s Disease, Atypical Parkinsonism, and Essential Tremor. Mov. Disord. 2013, 28, 1816–1822. [Google Scholar] [CrossRef]
- Brooks, D.J.; Seppi, K. Proposed Neuroimaging Criteria for the Diagnosis of Multiple System Atrophy. Mov. Disord. 2009, 24, 949–964. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
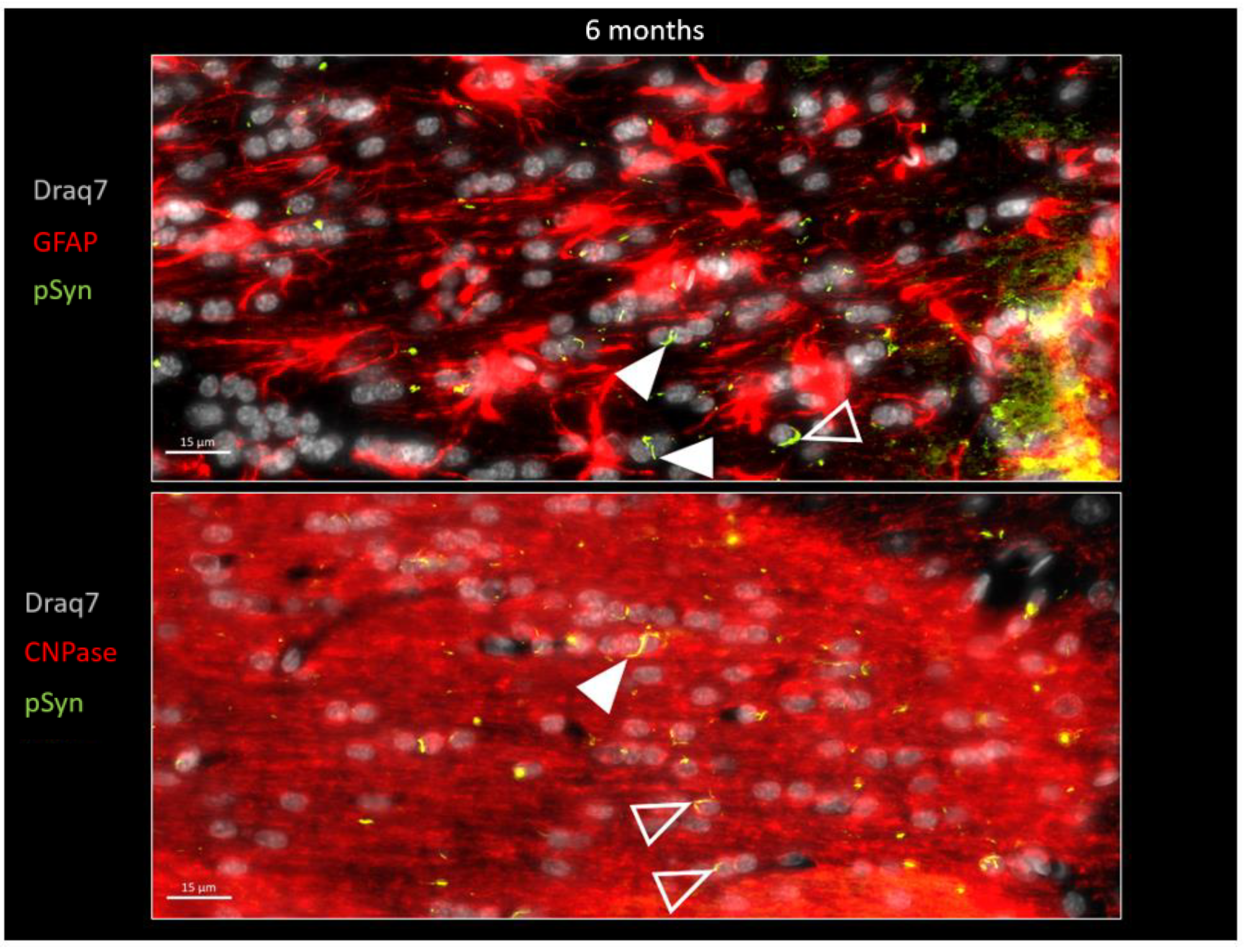{kind=link}
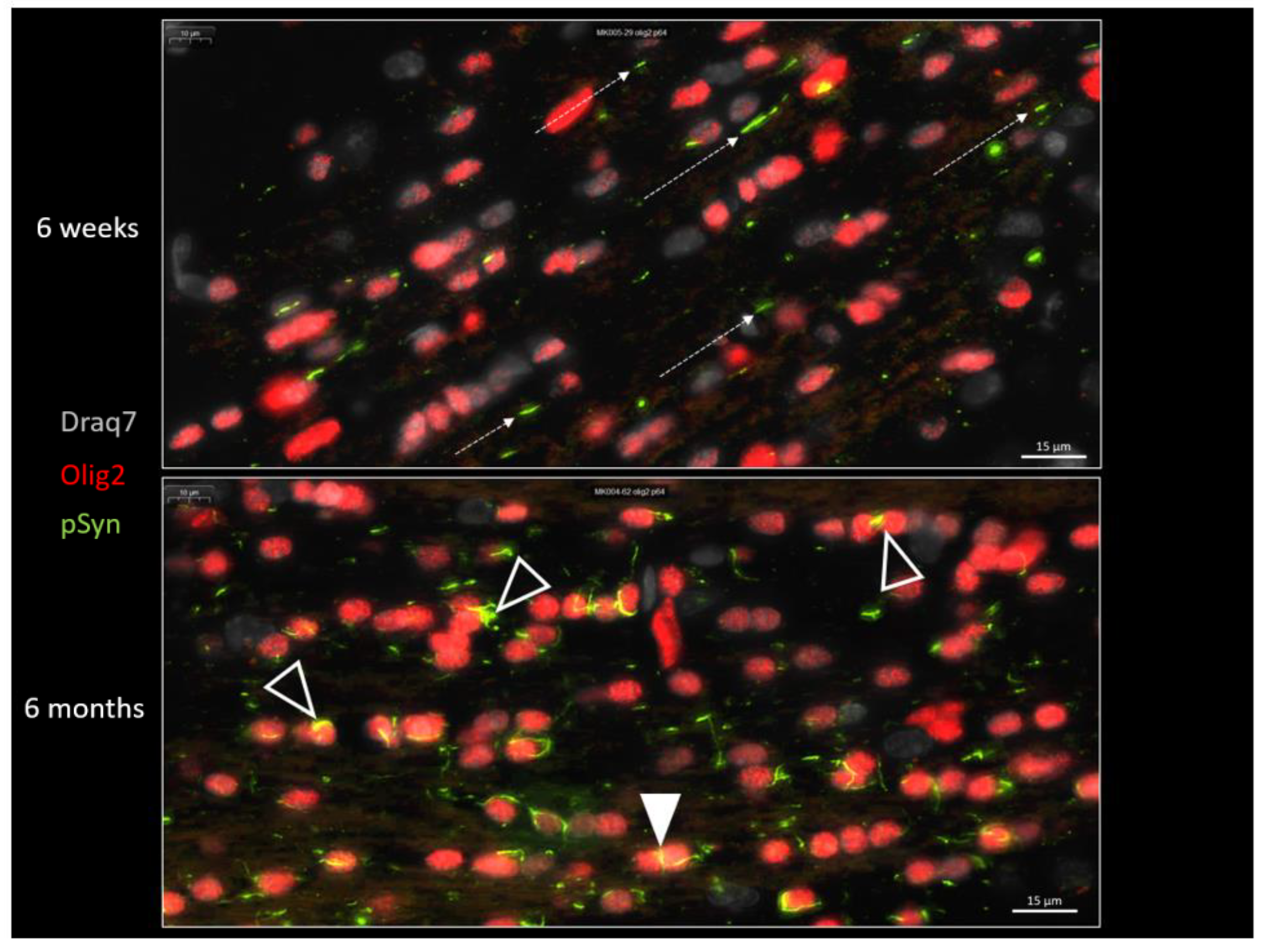{kind=link}
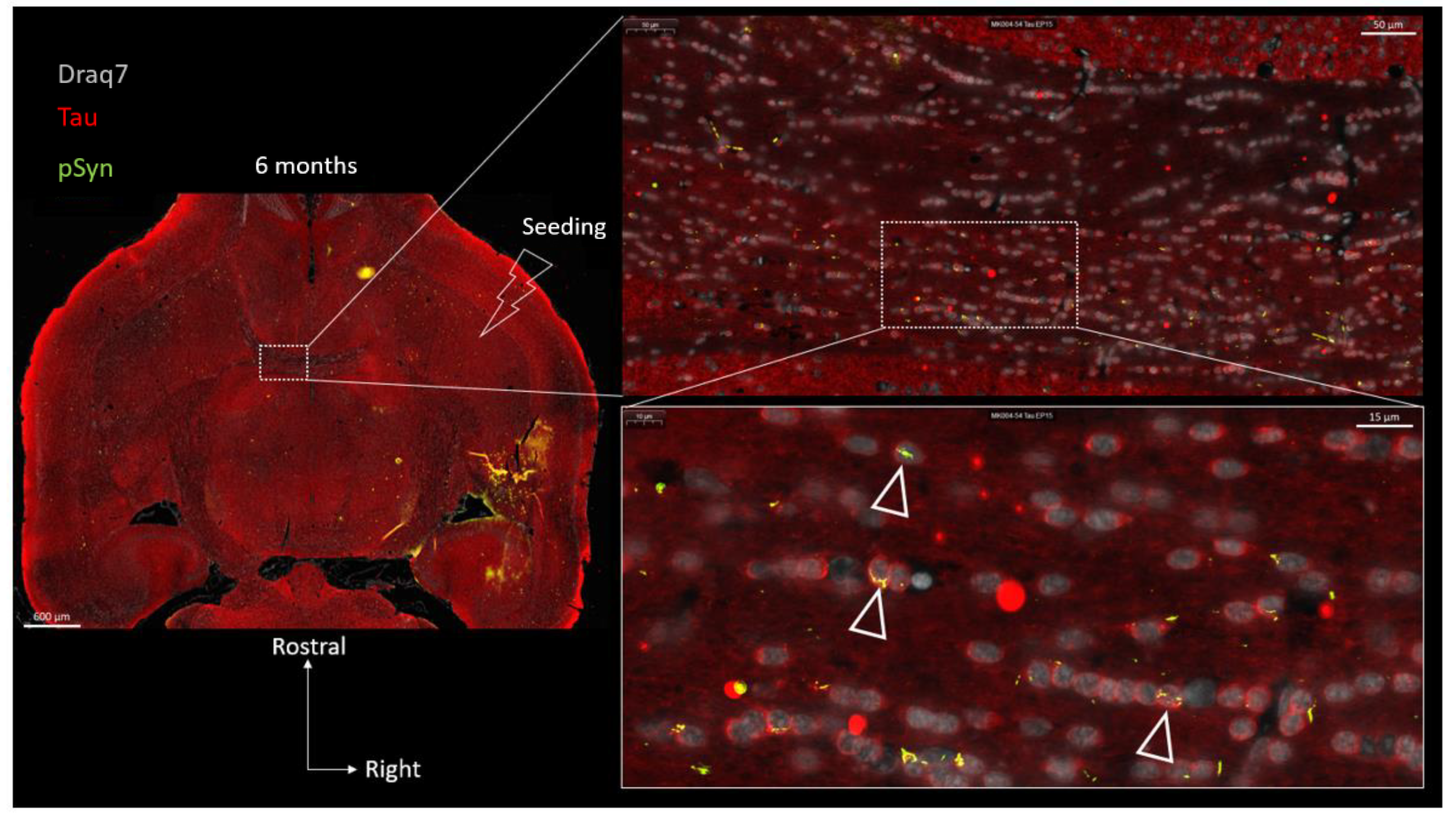{kind=link}
{kind=link}
{kind=link}
| DOI | ThT Value for Equal Amounts of α-Syn Fibrils (Variable A.U.) | Fold Difference | |
|---|---|---|---|
| mouse α-Syn fibrils | 10.1016/j.celrep.2016.08.053 | 40 | 3.75 |
| human α-Syn fibrils | 150 | ||
| human α-Syn ribbons | 10.1038/ncomms3575 | 4 | 8.25 |
| human α-Syn fibrils (type 2) | 33 | ||
| MSA-templated α-Syn fibrils (BH) | 10.1038/s41586-020-1984-7 | 480 | 12.7 |
| PD-templated α-Syn fibrils (BH) | 6100 | ||
| humanα-Syn fibrils 1B | 10.3390/biom12030436 | 2437 | 10.3 |
| human α-Syn fibrils (type 2) | 25100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Nuccio, F.; Kashyrina, M.; Serinelli, F.; Laferrière, F.; Lofrumento, D.D.; De Giorgi, F.; Ichas, F. Oligodendrocytes Prune Axons Containing α-Synuclein Aggregates In Vivo: Lewy Neurites as Precursors of Glial Cytoplasmic Inclusions in Multiple System Atrophy? Biomolecules 2023, 13, 269. https://doi.org/10.3390/biom13020269
De Nuccio F, Kashyrina M, Serinelli F, Laferrière F, Lofrumento DD, De Giorgi F, Ichas F. Oligodendrocytes Prune Axons Containing α-Synuclein Aggregates In Vivo: Lewy Neurites as Precursors of Glial Cytoplasmic Inclusions in Multiple System Atrophy? Biomolecules. 2023; 13(2):269. https://doi.org/10.3390/biom13020269
Chicago/Turabian StyleDe Nuccio, Francesco, Marianna Kashyrina, Francesca Serinelli, Florent Laferrière, Dario Domenico Lofrumento, Francesca De Giorgi, and François Ichas. 2023. "Oligodendrocytes Prune Axons Containing α-Synuclein Aggregates In Vivo: Lewy Neurites as Precursors of Glial Cytoplasmic Inclusions in Multiple System Atrophy?" Biomolecules 13, no. 2: 269. https://doi.org/10.3390/biom13020269