
Specific Human Milk Oligosaccharides Differentially Promote Th1 and Regulatory Responses in a CpG-Activated Epithelial/Immune Cell Coculture

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Human Peripheral Blood Mononuclear Cells
2.2. Preparation of HMOS
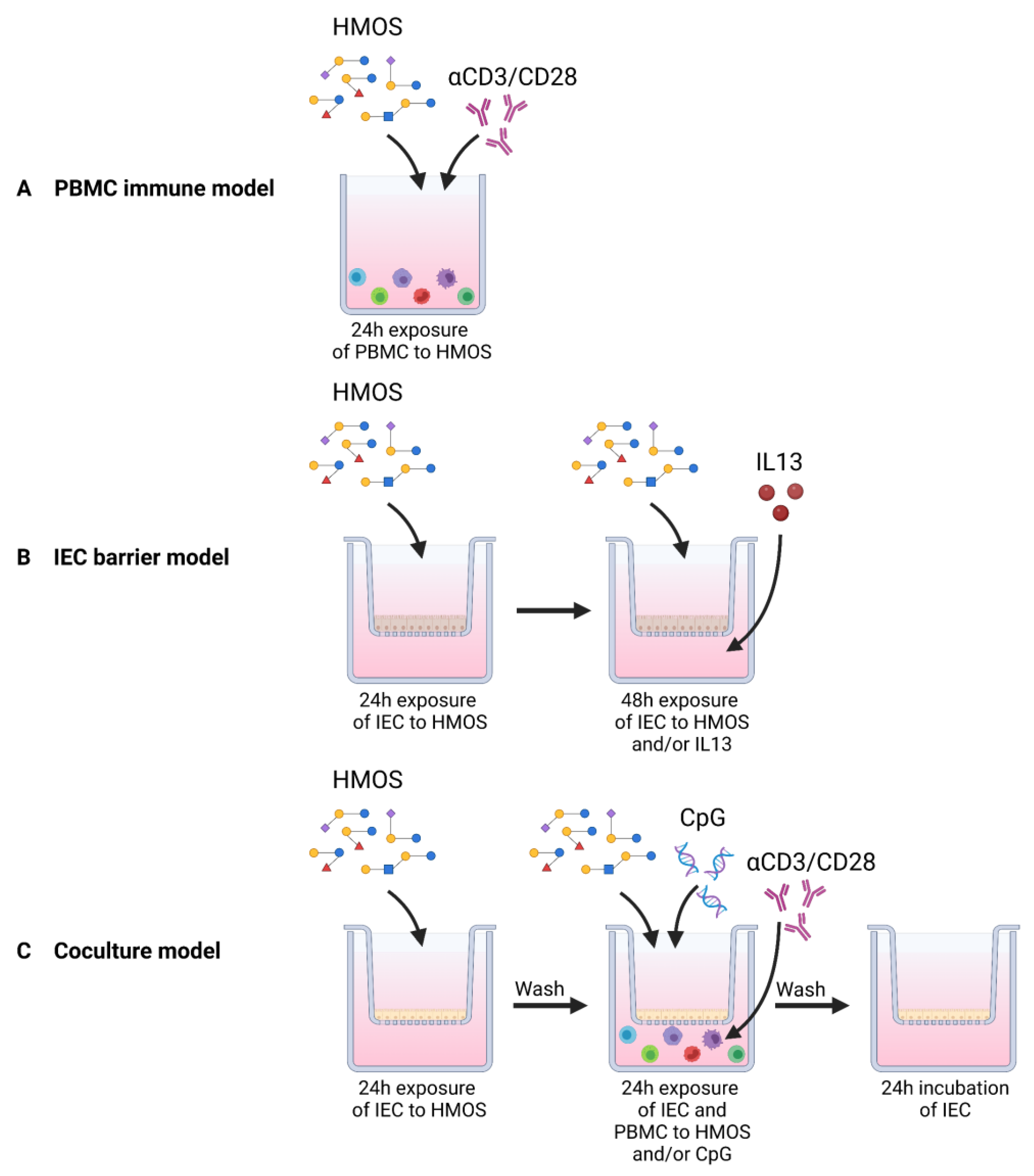
2.3. PBMC Immune Model
2.4. IEC Barrier Model
2.5. IEC/PBMC Coculture Model
2.6. Enzyme-Linked Immunosorbent Assay
2.7. Flow Cytometry Analysis
2.8. Statistical Analysis
3. Results
3.1. Direct Immunomodulatory Effects of Specific HMOS, Leading to Th1 Type and Regulatory Cytokine Responses from Activated PBMCs
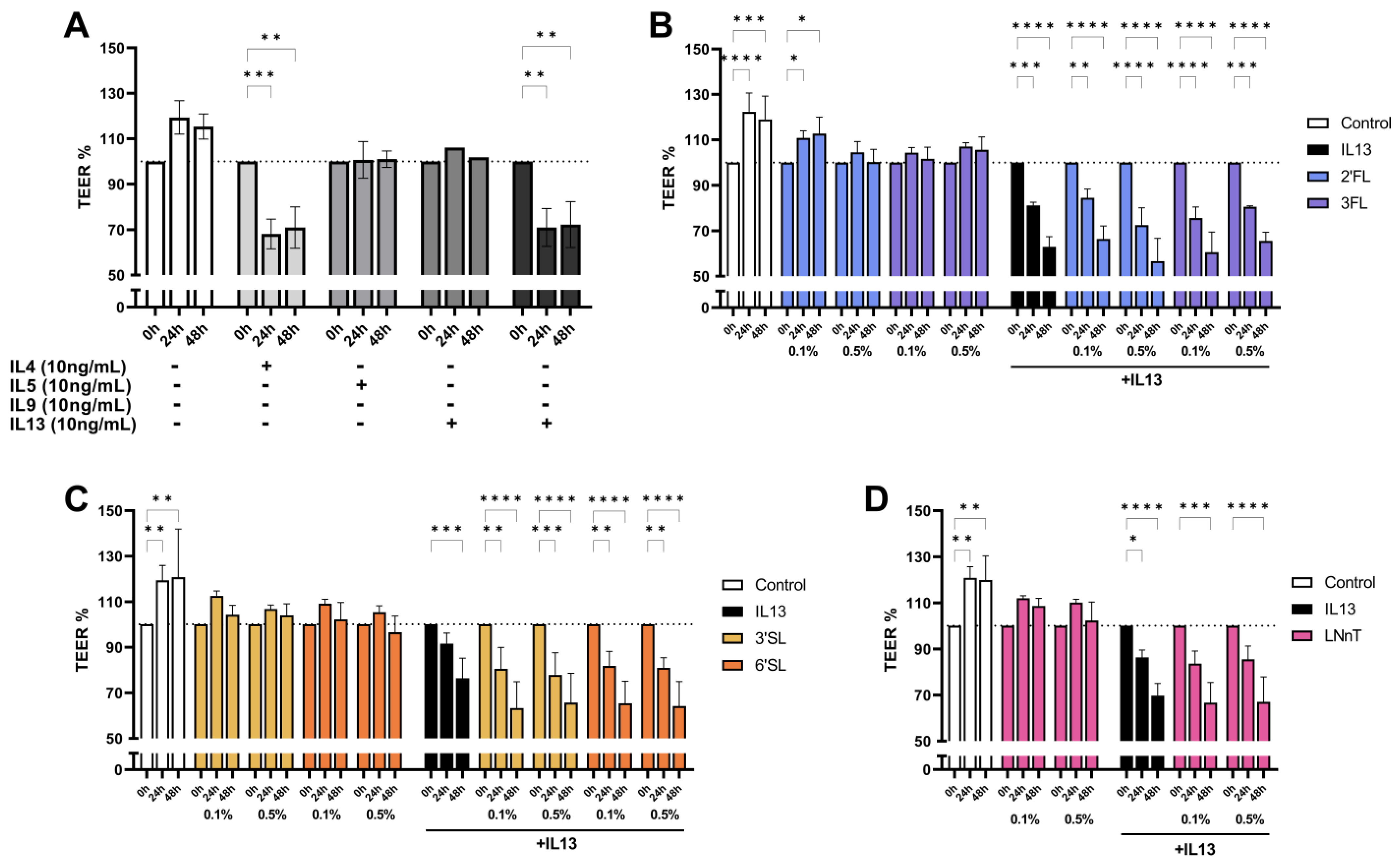
3.2. IL13 Induced Barrier Disruption in T84 Cells Is Not Prevented by HMOS Pre-Exposure
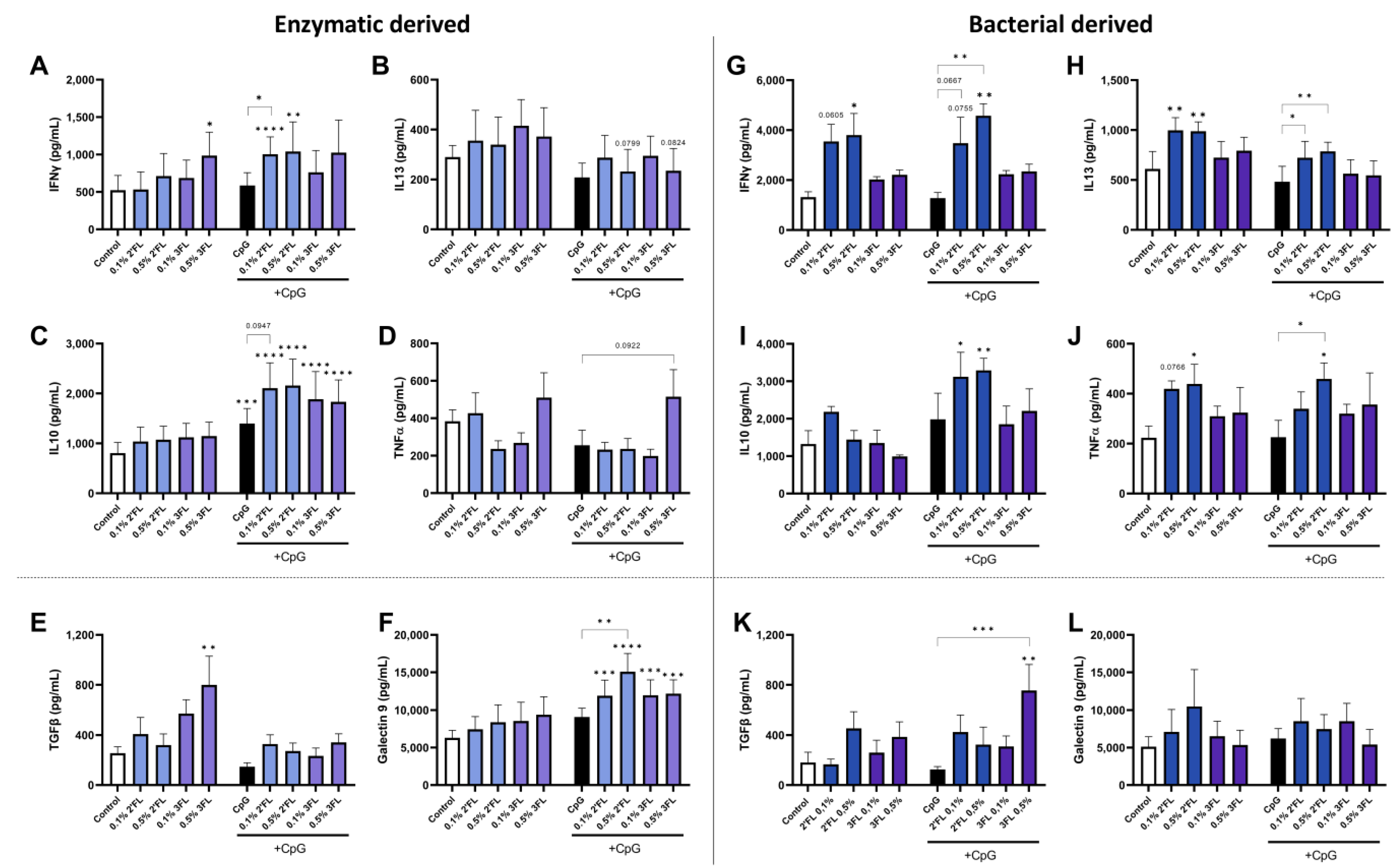
3.3. Crosstalk between IEC and PBMC Is Differentially Affected by 2′FL and 3FL
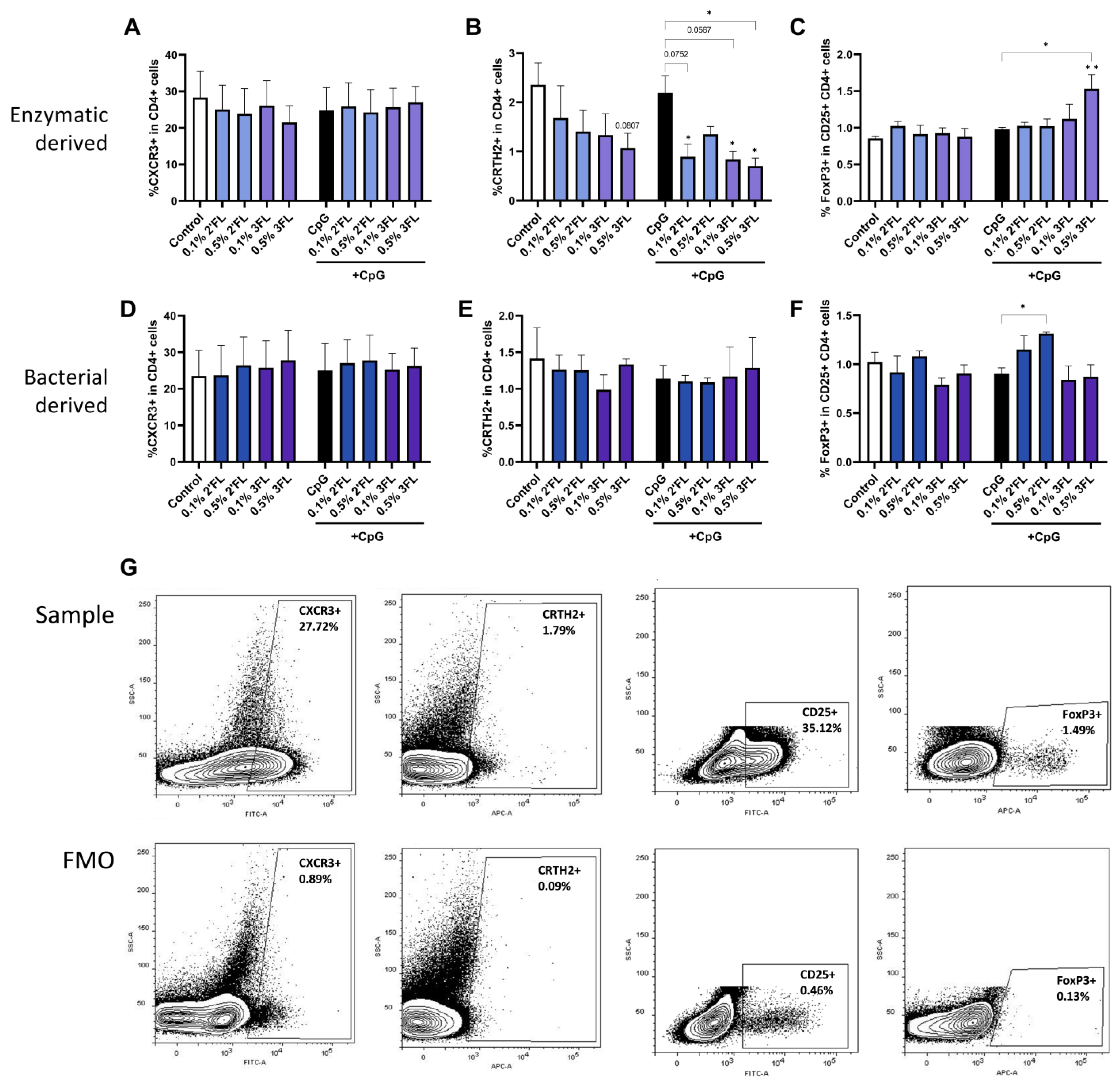
3.4. Enzymatic 3FL Decreases Th2 Type Development and Enhances the Polarization of Tregs
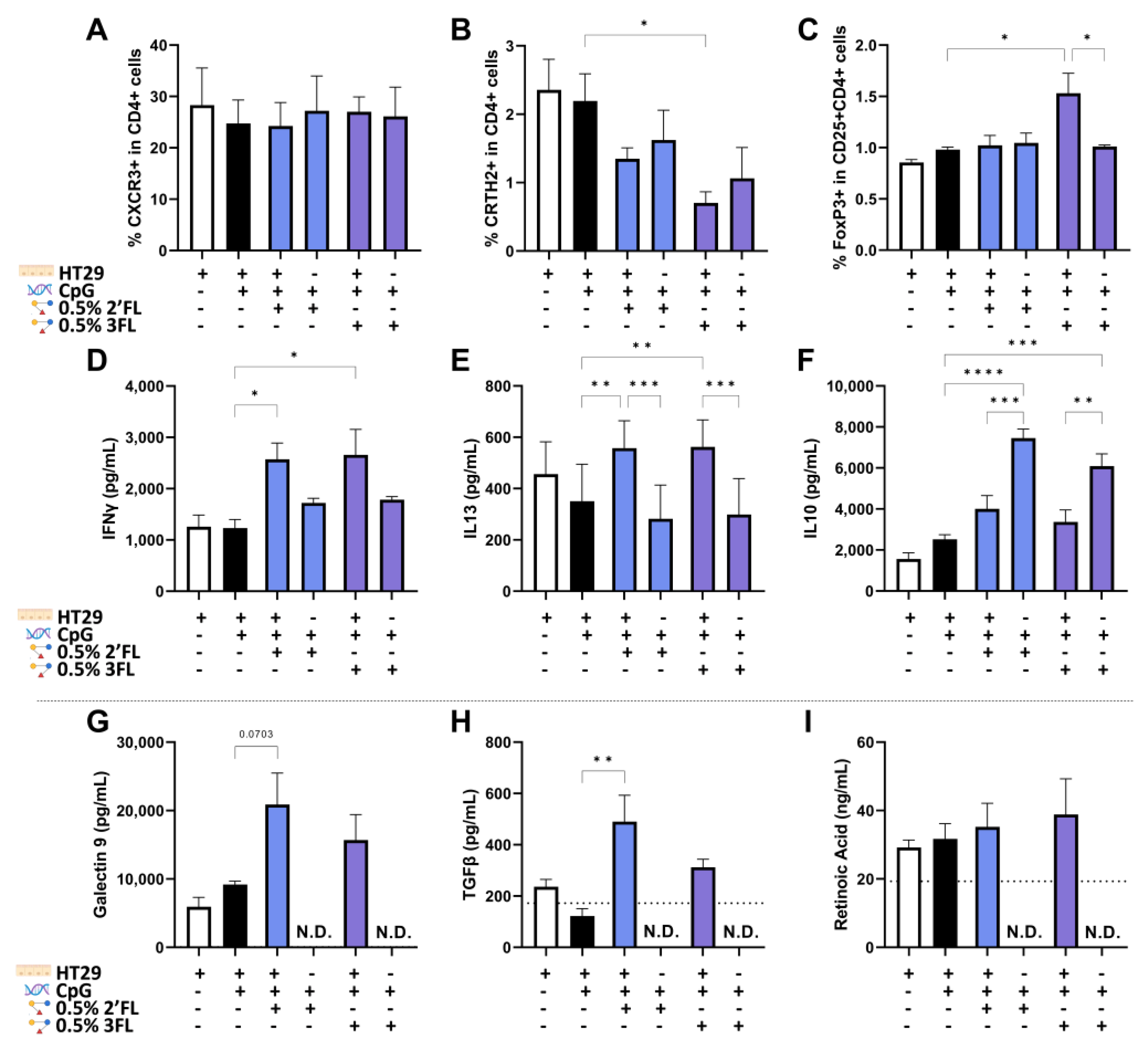
3.5. IEC Are Required for 3FL Mediated Treg Polarization and IEC Enhance 2′FL and 3FL Mediated Th1 Type and Th2 Type Cytokine Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thurl, S.; Munzert, M.; Boehm, G.; Matthews, C.; Stahl, B. Systematic review of the concentrations of oligosaccharides in human milk. Nutr. Rev. 2017, 75, 920–933. [Google Scholar] [CrossRef] [Green Version]
- Triantis, V.; Bode, L.; van Neerven, R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef] [Green Version]
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; van’t Land, B. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front. Pediatr. 2018, 6, 239. [Google Scholar] [CrossRef] [Green Version]
- Zivkovic, A.M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Human milk glycobiome and its impact on the infant gastrointestinal microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 4653–4658. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, S.; Schols, H.A.; Van Den Heuvel, E.G.H.M.; Voragen, A.G.J.; Gruppen, H. Occurrence of oligosaccharides in feces of breast-fed babies in their first six months of life and the corresponding breast milk. Carbohydr. Res. 2011, 346, 2540–2550. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Stroble, C.; Underwood, M.A.; Lebrilla, C.B. Detection of milk oligosaccharides in plasma of infants. Anal. Bioanal. Chem. 2014, 406, 5775–5784. [Google Scholar] [CrossRef] [Green Version]
- Goehring, K.C.; Kennedy, A.D.; Prieto, P.A.; Buck, R.H. Direct evidence for the presence of human milk oligosaccharides in the circulation of breastfed infants. PLoS ONE 2014, 9, e101692. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.R.; Chow, J.M.; Buck, R.H. Multifunctional Benefits of Prevalent HMOs: Implications for Infant Health. Nutrients 2021, 13, 3364. [Google Scholar] [CrossRef]
- Brugman, S.; Perdijk, O.; van Neerven, R.J.J.; Savelkoul, H.F.J. Mucosal Immune Development in Early Life: Setting the Stage. Arch. Immunol. Ther. Exp. 2015, 63, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Estorninos, E.; Lawenko, R.B.; Palestroque, E.; Sprenger, N.; Benyacoub, J.; Kortman, G.A.M.; Boekhorst, J.; Bettler, J.; Cercamondi, C.I.; Berger, B. Term infant formula supplemented with milk-derived oligosaccharides shifts the gut microbiota closer to that of human milk-fed infants and improves intestinal immune defense: A randomized controlled trial. Am. J. Clin. Nutr. 2022, 115, 142–153. [Google Scholar] [CrossRef]
- Groer, M.W.; Davis, M.W. Cytokines, infections, stress, and dysphoric moods in breastfeeders and formula feeders. JOGNN—J. Obstet. Gynecol. Neonatal Nurs. 2006, 35, 599–607. [Google Scholar] [CrossRef]
- Cheng, Y.; Yeung, C. Recent advance in infant nutrition: Human milk oligosaccharides. Pediatr. Neonatol. 2021, 62, 347–353. [Google Scholar] [CrossRef]
- Yu, J.; Shin, J.; Park, M.; Seydametova, E.; Jung, S.M.; Seo, J.H.; Kweon, D.H. Engineering of α-1,3-fucosyltransferases for production of 3-fucosyllactose in Escherichia coli. Metab. Eng. 2018, 48, 269–278. [Google Scholar] [CrossRef]
- Liu, J.J.; Kwak, S.; Pathanibul, P.; Lee, J.W.; Yu, S.; Yun, E.J.; Lim, H.; Kim, K.H.; Jin, Y.S. Biosynthesis of a Functional Human Milk Oligosaccharide, 2′-Fucosyllactose, and l -Fucose Using Engineered Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 2529–2536. [Google Scholar] [CrossRef]
- Jung, S.M.; Park, Y.C.; Seo, J.H. Production of 3-Fucosyllactose in Engineered Escherichia coli with α-1,3-Fucosyltransferase from Helicobacter pylori. Biotechnol. J. 2019, 14, 1800498. [Google Scholar] [CrossRef]
- Guo, Y.; Jers, C.; Meyer, A.S.; Li, H.; Kirpekar, F.; Mikkelsen, J.D. Modulating the regioselectivity of a Pasteurella multocida sialyltransferase for biocatalytic production of 3’- and 6’-sialyllactose. Enzym. Microb. Technol. 2015, 78, 54–62. [Google Scholar] [CrossRef]
- Biel-Nielsen, T.L.; Li, K.; Sørensen, S.O.; Sejberg, J.J.P.; Meyer, A.S.; Holck, J. Utilization of industrial citrus pectin side streams for enzymatic production of human milk oligosaccharides. Carbohydr. Res. 2022, 519, 108627. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Xue, M.; Liu, X.W.; Li, Y.; Chen, X.; Wang, P.G.; Wang, F.; Cao, H. Sequential one-pot multienzyme (OPME) synthesis of lacto-N-neotetraose and its sialyl and fucosyl derivatives. Chem. Commun. 2015, 51, 7689–7692. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Siziba, L.P.; Mank, M.; Stahl, B.; Gonsalves, J.; Blijenberg, B.; Rothenbacher, D.; Genuneit, J. Human Milk Oligosaccharide Profiles over 12 Months of Lactation: The Ulm SPATZ Health Study. Nutrients 2021, 13, 1973. [Google Scholar] [CrossRef]
- Shams-Ud-Doha, K.; Kitova, E.N.; Kitov, P.I.; St-Pierre, Y.; Klassen, J.S. Human Milk Oligosaccharide Specificities of Human Galectins. Comparison of Electrospray Ionization Mass Spectrometry and Glycan Microarray Screening Results. Anal. Chem. 2017, 89, 4914–4921. [Google Scholar] [CrossRef]
- El-Hawiet, A.; Chen, Y.; Shams-Ud-Doha, K.; Kitova, E.N.; St-Pierre, Y.; Klassen, J.S. High-Throughput Label- and Immobilization-Free Screening of Human Milk Oligosaccharides Against Lectins. Anal. Chem. 2017, 89, 8713–8722. [Google Scholar] [CrossRef]
- McFadden, J.P.; Thyssen, J.P.; Basketter, D.A.; Puangpet, P.; Kimber, I. T helper cell 2 immune skewing in pregnancy/early life: Chemical exposure and the development of atopic disease and allergy. Br. J. Dermatol. 2015, 172, 584–591. [Google Scholar] [CrossRef]
- Niewiem, M.; Grzybowska-Chlebowczyk, U. Intestinal Barrier Permeability in Allergic Diseases. Nutrients 2022, 14, 1893. [Google Scholar] [CrossRef]
- Wu, C.; Thalhamer, T.; Franca, R.F.; Xiao, S.; Wang, C.; Hotaa, C.; Zhu, C.; Hirashima, M.; Anderson, A.C.; Kuchroo, V.K. Galectin-9-CD44 interaction enhances stability and function of adaptive regulatory T cells. Immunity 2014, 41, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Robinson, B.S.; Saeedi, B.; Arthur, C.M.; Owens, J.; Naudin, C.; Ahmed, N.; Luo, L.; Jones, R.; Neish, A.; Stowell, S.R. Galectin-9 Is a Novel Regulator of Epithelial Restitution. Am. J. Pathol. 2020, 190, 1657–1666. [Google Scholar] [CrossRef]
- De Kivit, S.; Kraneveld, A.D.; Knippels, L.M.J.; Van Kooyk, Y.; Garssen, J.; Willemsen, L.E.M. Intestinal epithelium-derived galectin-9 is involved in the immunomodulating effects of nondigestible oligosaccharides. J. Innate Immun. 2013, 5, 625–638. [Google Scholar] [CrossRef]
- Ayechu-Muruzabal, V.; Overbeek, S.A.; Kostadinova, A.I.; Stahl, B.; Garssen, J.; van’t Land, B.; Willemsen, L.E.M. Exposure of intestinal epithelial cells to 2′-fucosyllactose and cpg enhances galectin release and instructs dendritic cells to drive th1 and regulatory-type immune development. Biomolecules 2020, 10, 784. [Google Scholar] [CrossRef]
- Rudloff, S.; Pohlentz, G.; Diekmann, L.; Egge, H.; Kunz, C. Urinary excretion of lactose and oligosaccharides in preterm infants fed human milk or infant formula. Acta Paediatr. Int. J. Paediatr. 1996, 85, 598–603. [Google Scholar] [CrossRef]
- Zuurveld, M.; van Witzenburg, N.P.; Garssen, J.; Folkerts, G.; Stahl, B.; van’t Land, B.; Willemsen, L.E.M. Immunomodulation by Human Milk Oligosaccharides: The Potential Role in Prevention of Allergic Diseases. Front. Immunol. 2020, 11, 801. [Google Scholar] [CrossRef]
- McCormick, S.M.; Heller, N.M. Commentary: IL-4 and IL-13 Receptors and Signaling. Cytokine 2015, 75, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Hayen, S.M.; Otten, H.G.; Overbeek, S.A.; Knulst, A.C.; Garssen, J.; Willemsen, L.E.M. Exposure of intestinal epithelial cells to short- and long-chain fructo-oligosaccharides and CpG oligodeoxynucleotides enhances peanut-specific T Helper 1 polarization. Front. Immunol. 2018, 9, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kivit, S.; van Hoffen, E.; Korthagen, N.; Garssen, J.; Willemsen, L.E.M. Apical TLR ligation of intestinal epithelial cells drives a Th1-polarized regulatory or inflammatory type effector response in vitro. Immunobiology 2011, 216, 518–527. [Google Scholar] [CrossRef]
- Xiao, S.; Jin, H.; Liu, S.; Korn, T.; Oukka, M.; Lim, B.; Kuchroo, V. Retinoic Acid Increases Foxp3+ Regulatory T Cells and Inhibits Development of Th17 Cells by Enhancing TGF-b-driven Smad3 Signaling and Inhibiting IL-6 and IL-23 Receptor Expression. Clin. Immunol. 2008, 127, S55. [Google Scholar] [CrossRef]
- Lv, K.; Zhang, Y.; Zhang, M.; Zhong, M.; Suo, Q. Galectin-9 promotes TGF-β1-dependent induction of regulatory T cells via the TGF-β/Smad signaling pathway. Mol. Med. Rep. 2013, 7, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Cummings, R.D. T cells are Smad’ly in Love with galectin-9. Immunity 2014, 41, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Boehm, G.; Stahl, B. Oligosaccharides from Milk. J. Nutr. 2007, 137, 847S–849S. [Google Scholar] [CrossRef] [Green Version]
- Toutounchi, N.S.; Braber, S.; Hogenkamp, A.; Varasteh, S.; Cai, Y.; Wehkamp, T.; Tims, S.; Leusink-Muis, T.; van Ark, I.; Wiertsema, S.; et al. Human milk oligosaccharide 3′-gl improves influenza-specific vaccination responsiveness and immunity after deoxynivalenol exposure in preclinical models. Nutrients 2021, 13, 3190. [Google Scholar] [CrossRef]
- Shim, J.O. Human milk oligosaccharides as immunonutrition key in early life. Clin. Exp. Pediatr. 2022, 65, 344–345. [Google Scholar] [CrossRef]
- Mavrogeni, M.E.; Asadpoor, M.; Henricks, P.A.J.; Keshavarzian, A.; Folkerts, G.; Braber, S. Direct action of non-digestible oligosaccharides against a leaky gut. Nutrients 2022, 14, 4699. [Google Scholar] [CrossRef]
- Xiao, L.; van De Worp, W.R.P.H.; Stassen, R.; van Maastrigt, C.; Kettelarij, N.; Stahl, B.; Blijenberg, B.; Overbeek, S.A.; Folkerts, G.; Garssen, J.; et al. Human milk oligosaccharides promote immune tolerance via direct interactions with human dendritic cells. Eur. J. Immunol. 2019, 49, 1001–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, F.; Sharma, A.K.; Gurung, M.; Casero, D.; Matazel, K.; Bode, L.; Simecka, C.; Elolimy, A.A.; Tripp, P.; Randolph, C.; et al. Human Milk Oligosaccharides Impact Cellular and Inflammatory Gene Expression and Immune Response. Front. Immunol. 2022, 13, 907529. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D.S.; Ko, J.S.; Leone, S.; Nanthakumar, N.N. Human Milk Oligosaccharides and Synthetic Galactosyloligosaccharides Contain 3′-, 4-, and 6′-Galactosyllactose and Attenuate Inflammation in Human T84, NCM-460, and H4 Cells and Intestinal Tissue Ex Vivo. J. Nutr. 2016, 146, 358–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Mowat, A.M.I. To respond or not to respond—A personal perspective of intestinal tolerance. Nat. Rev. Immunol. 2018, 18, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Moossavi, S.; Miliku, K.; Sepehri, S.; Khafipour, E.; Azad, M.B. The Prebiotic and Probiotic Properties of Human Milk: Implications for Infant Immune Development and Pediatric Asthma. Front. Pediatr. 2018, 6, 197. [Google Scholar] [CrossRef] [Green Version]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef]
- Sprenger, N.; Tytgat, H.L.P.; Binia, A.; Austin, S.; Singhal, A. Biology of human milk oligosaccharides: From basic science to clinical evidence. J. Hum. Nutr. Diet. 2022, 35, 280–299. [Google Scholar] [CrossRef]
- Liu, F.; Simpson, A.B.; D’Costa, E.; Bunn, F.S.; van Leeuwen, S.S. Sialic acid, the secret gift for the brain. Crit. Rev. Food Sci. Nutr. 2022, 1–20. [Google Scholar] [CrossRef]
- Holscher, H.D.; Davis, S.R.; Tappenden, K.A. Human Milk Oligosaccharides Influence Maturation of Human Intestinal Caco-2Bbe and HT-29 Cell Lines. J. Nutr. 2014, 144, 586–591. [Google Scholar] [CrossRef]
- Holscher, H.D.; Bode, L.; Tappenden, K.A. Human Milk Oligosaccharides Influence Intestinal Epithelial Cell Maturation in Vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Perdijk, O.; Joost van Neerven, R.J.; Van den Brink, E.; Savelkoul, H.F.J.; Brugman, S. The oligosaccharides 6’-sialyllactose, 2’-fucosyllactose or galactooligosaccharides do not directly modulate human dendritic cell differentiation or maturation. PLoS ONE 2018, 13, e0200356. [Google Scholar] [CrossRef] [PubMed]
- Perdijk, O.; Joost Van Neerven, R.J.; Meijer, B.; Savelkoul, H.F.J.; Brugman, S. Induction of human tolerogenic dendritic cells by 3′-sialyllactose via TLR4 is explained by LPS contamination. Glycobiology 2018, 28, 126–130. [Google Scholar] [CrossRef] [Green Version]
- De Kivit, S.; Saeland, E.; Kraneveld, A.D.; Van De Kant, H.J.G.; Schouten, B.; Van Esch, B.C.A.M.; Knol, J.; Sprikkelman, A.B.; Van Der Aa, L.B.; Knippels, L.M.J.; et al. Galectin-9 induced by dietary synbiotics is involved in suppression of allergic symptoms in mice and humans. Allergy Eur. J. Allergy Clin. Immunol. 2012, 67, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhu, Y.; Li, Z.; Zhang, W.; Mu, W. Recent Advances on Lacto- N-neotetraose, a Commercially Added Human Milk Oligosaccharide in Infant Formula. J. Agric. Food Chem. 2022, 70, 4534–4547. [Google Scholar] [CrossRef] [PubMed]
- Hester, S.N.; Donovan, S.M. Individual and Combined Effects of Nucleotides and Human Milk Oligosaccharides on Proliferation, Apoptosis and Necrosis in a Human Fetal Intestinal Cell Line. Food Nutr. Sci. 2012, 03, 1567–1576. [Google Scholar] [CrossRef] [Green Version]
- Natividad, J.M.; Rytz, A.; Keddani, S.; Bergonzelli, G.; Garcia-rodenas, C.L. Blends of human milk oligosaccharides confer intestinal epithelial barrier protection in vitro. Nutrients 2020, 12, 3047. [Google Scholar] [CrossRef]
- He, Y.Y.; Liu, S.B.; Kling, D.E.; Leone, S.; Lawlor, N.T.; Huang, Y.; Feinberg, S.B.; Hill, D.R.; Newburg, D.S. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016, 65, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Williams, J.; Washington, M.K.; Yang, Y.; Long, J.; Townsend, S.D.; Yan, F. 2’-Fucosyllactose Ameliorates Chemotherapy-Induced Intestinal Mucositis by Protecting Intestinal Epithelial Cells Against Apoptosis. CMGH 2022, 13, 441–457. [Google Scholar] [CrossRef]
- Cheng, L.; Kong, C.; Walvoort, M.T.C.; Faas, M.M.; Vos, P. De Human Milk Oligosaccharides Differently Modulate Goblet Cells Under Homeostatic, Proinflammatory Conditions and ER Stress. Mol. Nutr. Food Res. 2020, 64, 1900976. [Google Scholar] [CrossRef]
- Natividad, J.M.; Marsaux, B.; Rodenas, C.L.G.; Rytz, A.; Vandevijver, G.; Marzorati, M.; Van den Abbeele, P.; Calatayud, M.; Rochat, F. Human Milk Oligosaccharides and Lactose Differentially Affect Infant Gut Microbiota and Intestinal Barrier In Vitro. Nutrients 2022, 14, 2546. [Google Scholar] [CrossRef] [PubMed]
- Šuligoj, T.; Vigsnæs, L.K.; Van Den Abbeele, P.; Apostolou, A.; Karalis, K.; Savva, G.M.; Mcconnell, B.; Juge, N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients 2020, 12, 2808. [Google Scholar] [CrossRef] [PubMed]
- Ayechu-Muruzabal, V.; van de Kaa, M.; Mukherjee, R.; Garssen, J.; Stahl, B.; Pieters, R.J.; Van’T Land, B.; Kraneveld, A.D.; Willemsen, L.E.M. Modulation of the Epithelial-Immune Cell Crosstalk and Related Galectin Secretion by DP3-5 Galacto-Oligosaccharides and β-3′galactosyllactose. Biomolecules 2022, 12, 384. [Google Scholar] [CrossRef] [PubMed]
- De Kivit, S.; Kostadinova, A.I.; Kerperien, J.; Morgan, M.E.; Ayechu-Muruzabal, V.; Hofman, G.A.; Knippels, L.M.J.; Kraneveld, A.D.; Garssen, J.; Willemsen, L.E.M. Dietary, nondigestible oligosaccharides and Bifidobacterium breve M-16V suppress allergic inflammation in intestine via targeting dendritic cell maturation. J. Leukoc. Biol. 2017, 102, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, R.; Van De Kaa, M.; Garssen, J.; Pieters, R.J.; Kraneveld, A.D.; Willemsen, L.E.M. Lactulose synergizes with CpG-ODN to modulate epithelial and immune cells cross talk. Food Funct. 2019, 10, 33–37. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Origin | Purity | Endotoxin (EU/mg) |
|---|---|---|---|
| 2′FL | Enzymatic | >95% | 0.03 |
| 3FL | Enzymatic | 96.8% | 0.0626 |
| 2′FL | Bacterial | 92.7% | 0.0175 |
| 3FL | Bacterial | 94.1% | 0.025 |
| LNnT | Enzymatic | 99.9% | 0.0616 |
| 3′SL | Enzymatic | 95.8% | 0.0516 |
| 6′SL | Enzymatic | 98.1% | 1.023 |
| (pg/mL) | Control | 0.01% 2′FL | 0.05% 2′FL | 0.1% 2′FL | 0.01% 3FL | 0.05% 3FL | 0.1% 3FL | 0.01% 3′SL | 0.05% 3′SL | 0.1% 3′SL | 0.01% 6′SL | 0.05% 6′SL | 0.1% 6′SL | 0.01% LNnT | 0.05% LNnT | 0.1% LNnT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IFNγ | 5.7 ± 3.5 | 18.5 ± 11.8 | 41.3 ± 23.6 | 145.7 ± 108.3 | 40.7 ± 35.4 | 182.5 ± 87.9 1 | 193.5 ± 101.8 * | 77.2 ± 36.2 | 125.4 ± 45.7 | 63.5 ± 23.5 | 5.7 ± 3.6 | 144.2 ± 65.6 | 78.2 ± 37.8 | 19.8 ± 12.1 | 38.3 ± 15.3 | 164.6 ± 94.6 |
| TNFα | 1424 ± 802.8 | 1195 ± 746.0 | 1185 ± 476.4 | 703.8 ± 258.2 | 633.6 ± 260.2 | 699.4 ± 142.6 | 905.1 ± 257.6 | 1083 ± 455.4 | 826.5 ± 332.6 | 673.8 ± 248.5 | 2526 ± 1922 | 2175 ± 1129 | 2757 ± 1501 | 700.5 ± 282.1 | 908.7 ± 499.5 | 980.5 ± 528.1 |
| IL13 | 236.4 ± 91.4 | 128.2 ± 39.6 | 123.8 ± 50.5 | 167.7 ± 71.1 | 128.2 ± 56.4 | 181.6 ± 55.5 | 184.0 ± 57.0 | 101.0 ± 57.0 | 160.2 ± 93.4 | 82.9 ± 32.1 | 253.3 ± 100.4 | 129.4 ± 37.5 | 119.3 ± 24.4 | 158.1 ± 68. | 105.5 ± 33.6 | 139.3 ± 35.8 |
| IL9 | 8.5 ± 1.9 | 17.8 ± 7.0 | 33.2 ± 16.9 2 | 22.8 ± 4.7 | 23.9 ± 8.7 | 16.2 ± 4.7 | 16.3 ± 6.4 | 20.8 ± 6.3 | 24.8 ± 7.3 | 17.1 ± 5.2 | 15.4 ± 3.5 | 20.3 ± 4.1 | 26.2 ± 9.9 | 27.2 ± 7.2 | 21.4 ± 7.2 | 18.9 ± 6.5 |
| IL10 | 113.9 ± 63.7 | 134.4 ± 61.8 | 164.4 ± 92.0 | 398.7 ± 160.8 | 68.4 ± 46.2 | 164.1 ± 136.0 | 161.5 ± 90.4 | 296.4 ± 163.5 | 269.5 ± 142.6 | 250.2 ± 144.1 | 111.5 ± 41.0 | 442.7 ± 139.6 | 612.9 ± 127.4 * | 156.3 ± 53.5 | 58.1 ± 36.6 | 153.6 ± 111.8 |
| Gal-9 | 11755 ± 2229 | 5386 ± 1669 * | 3646 ± 1693 ** | 4740 ± 1435 * | 2221 ± 655.1 *** | 3041 ± 1131 ** | 2777 ± 924.6 *** | 1483 ± 332.8 *** | 1815 ± 575.6 *** | 4325 ± 2077 * | 9040 ± 1097 | 9219 ± 3542 | 4470 ± 1395 * | 2604 ± 1087 *** | 3828 ± 1111 *** | 3122± 1090 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuurveld, M.; Ayechu-Muruzabal, V.; Folkerts, G.; Garssen, J.; van‘t Land, B.; Willemsen, L.E.M. Specific Human Milk Oligosaccharides Differentially Promote Th1 and Regulatory Responses in a CpG-Activated Epithelial/Immune Cell Coculture. Biomolecules 2023, 13, 263. https://doi.org/10.3390/biom13020263
Zuurveld M, Ayechu-Muruzabal V, Folkerts G, Garssen J, van‘t Land B, Willemsen LEM. Specific Human Milk Oligosaccharides Differentially Promote Th1 and Regulatory Responses in a CpG-Activated Epithelial/Immune Cell Coculture. Biomolecules. 2023; 13(2):263. https://doi.org/10.3390/biom13020263
Chicago/Turabian StyleZuurveld, Marit, Veronica Ayechu-Muruzabal, Gert Folkerts, Johan Garssen, Belinda van‘t Land, and Linette E. M. Willemsen. 2023. "Specific Human Milk Oligosaccharides Differentially Promote Th1 and Regulatory Responses in a CpG-Activated Epithelial/Immune Cell Coculture" Biomolecules 13, no. 2: 263. https://doi.org/10.3390/biom13020263