
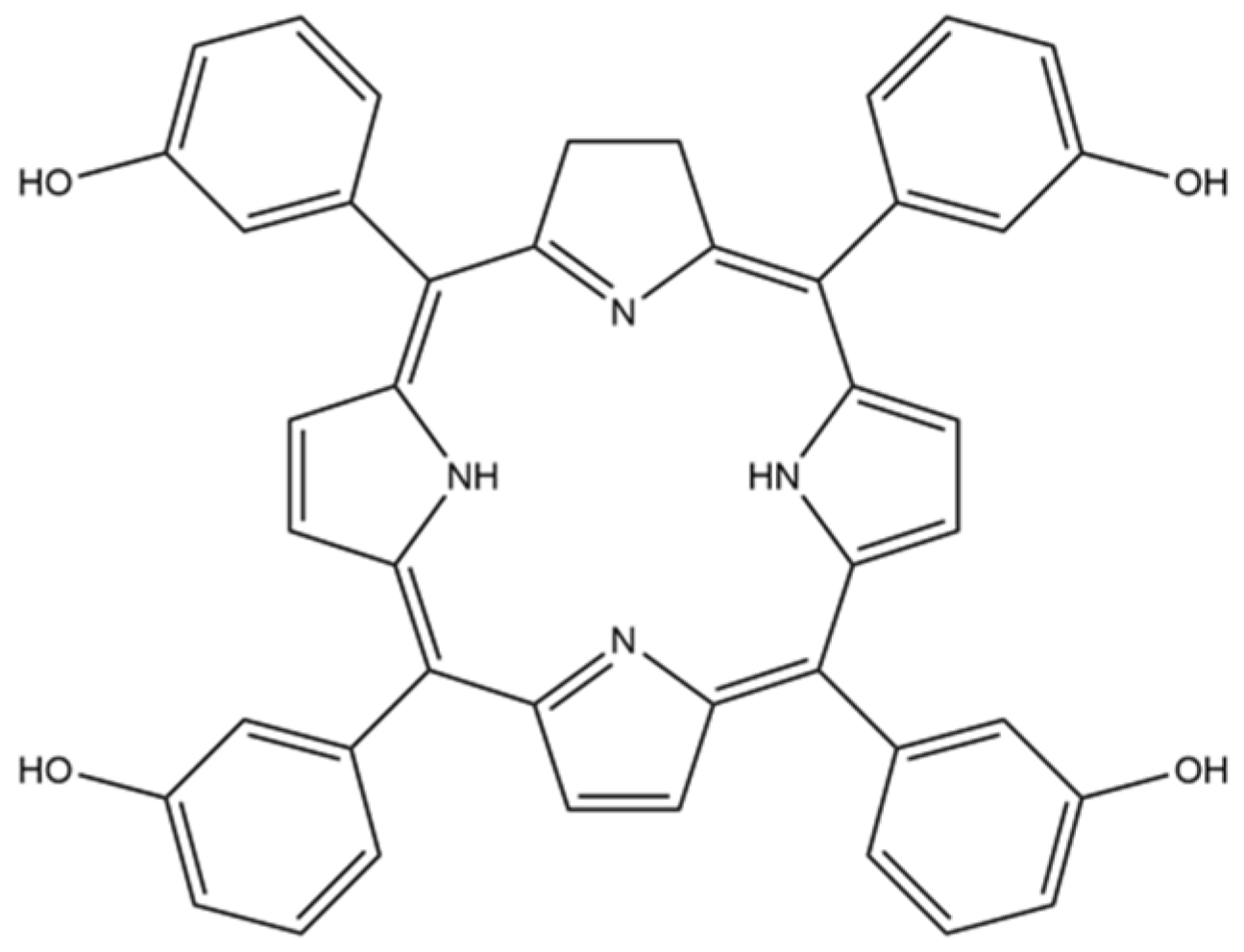
Carrying Temoporfin with Human Serum Albumin: A New Perspective for Photodynamic Application in Head and Neck Cancer

 , , , , , , ,
, , , , , , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Characterization of the mTHPC@HSA Complex
2.1.1. Materials
2.1.2. Synthesis and Purification of mTHPC@HSA Complex
2.1.3. Characterization of mTHPC@HSA Complex
2.1.4. Detection of Singlet Oxygen Generation
2.2. Computational Investigation of the mTHPC@HSA Complex
2.2.1. Ensemble Docking
2.2.2. MD Simulations
2.2.3. Molecular Mechanics/Generalized Born Surface Area (MM/GBSA) Analysis
2.3. Cytotoxicity and Phototoxicity of mTHPC@HSA Complex in HNSCC Cells
2.3.1. Cell Lines
2.3.2. Cell Viability
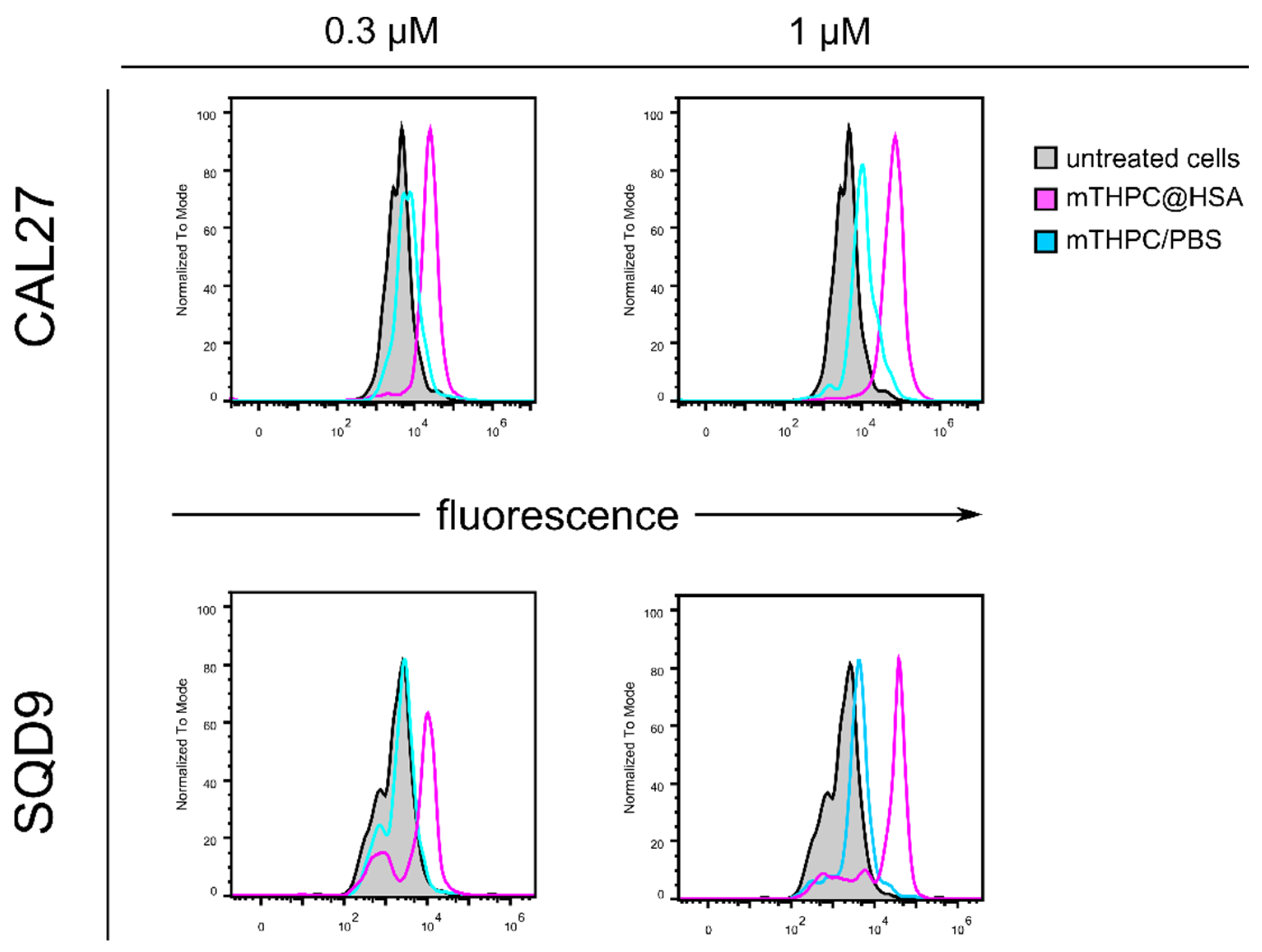
2.3.3. Analysis of Intracellular Uptake
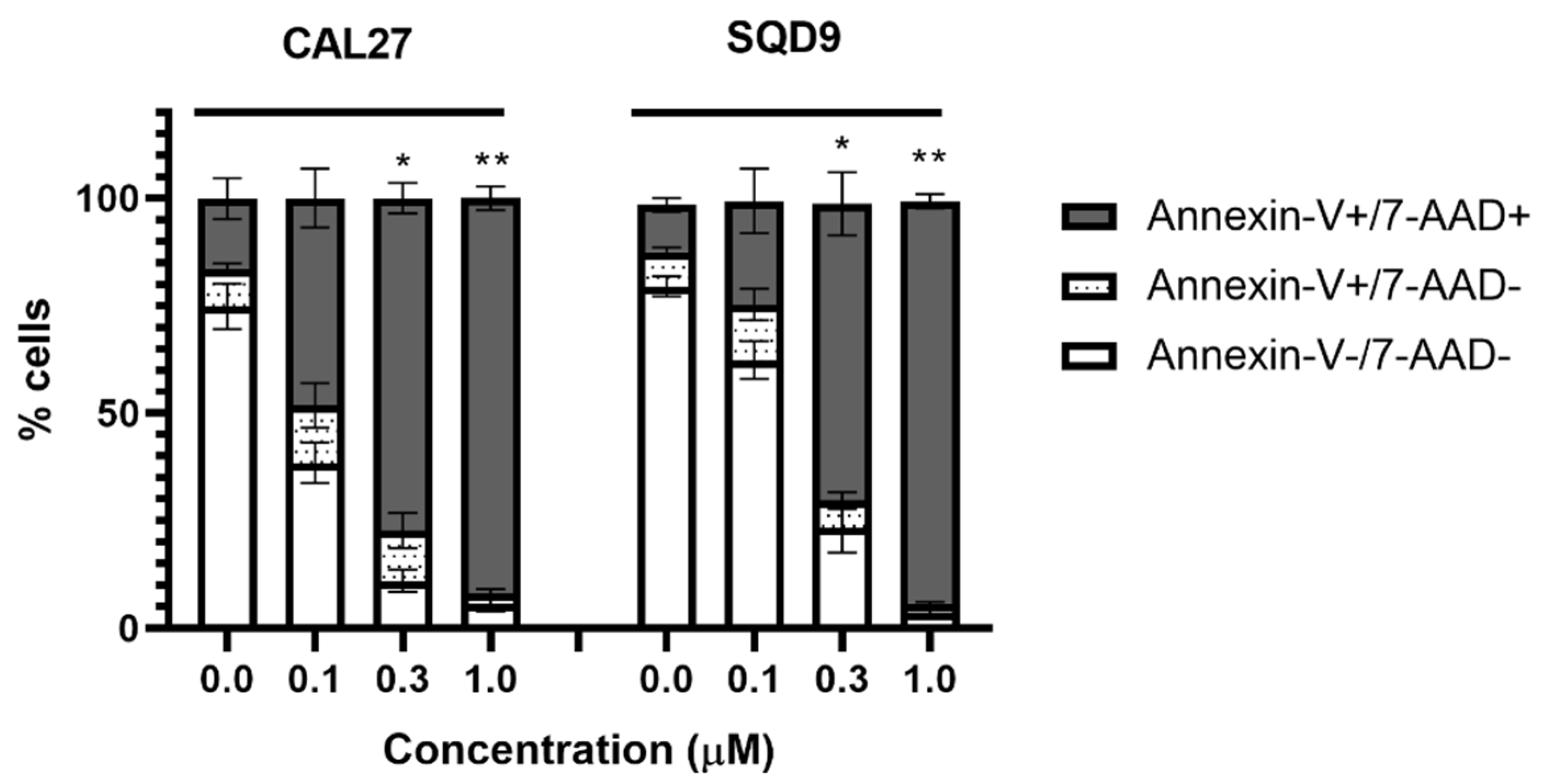
2.3.4. Analysis of Cell Death Mechanisms
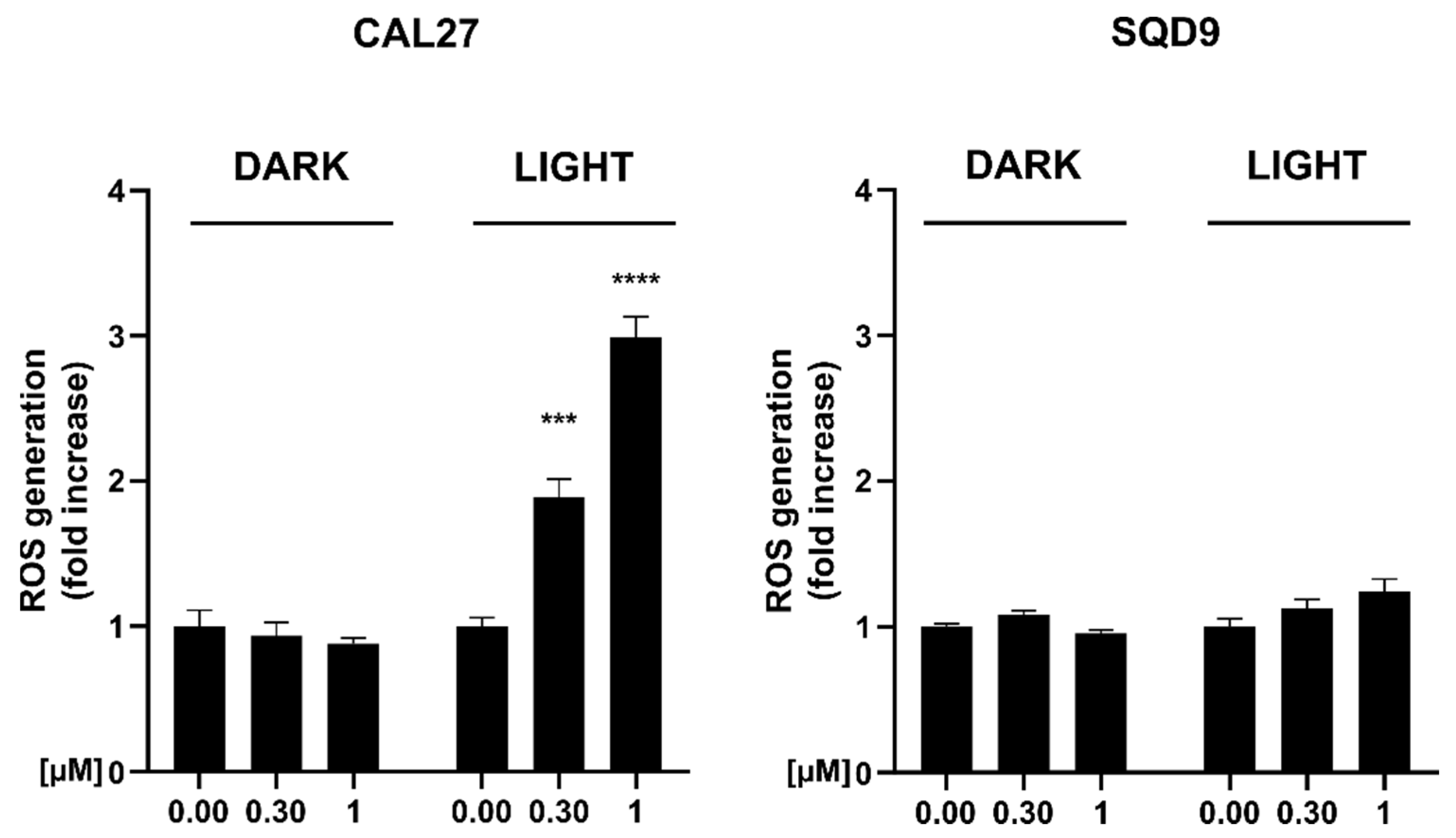
2.3.5. Intracellular ROS Generation
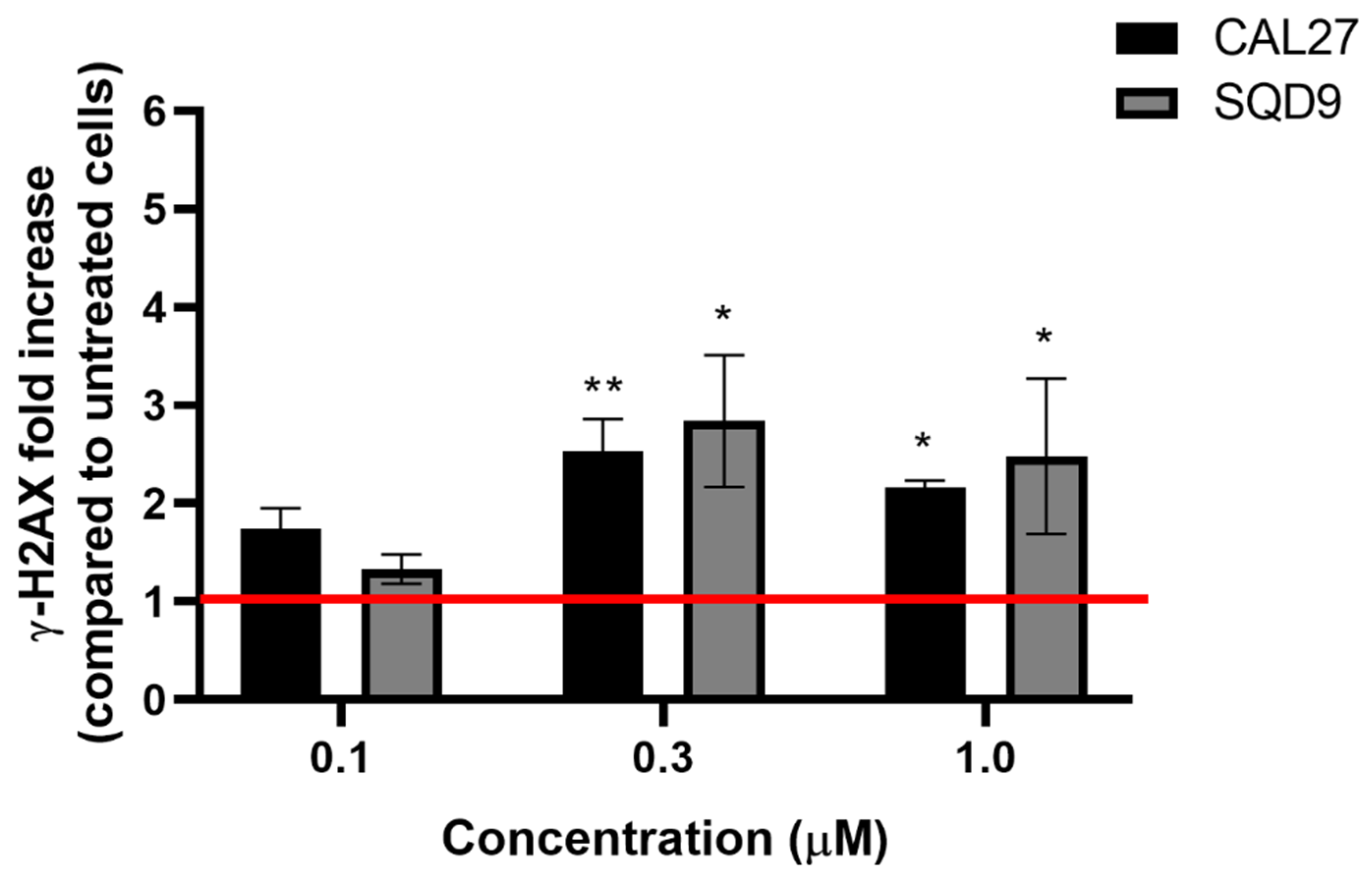
2.3.6. H2AX Phosphorylation Assay
2.3.7. Statistical Analysis
3. Results and Discussion
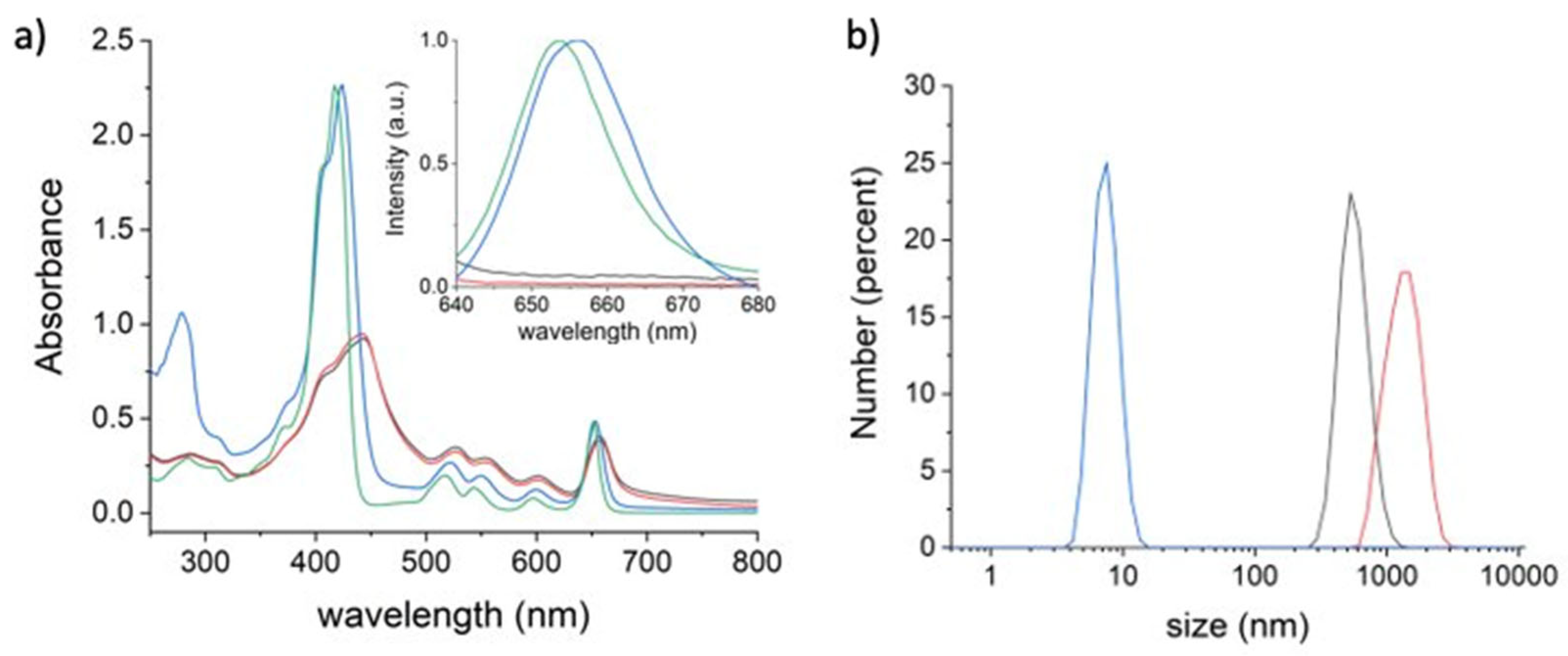
3.1. Synthesis and Characterization of THPC@HSA Complex
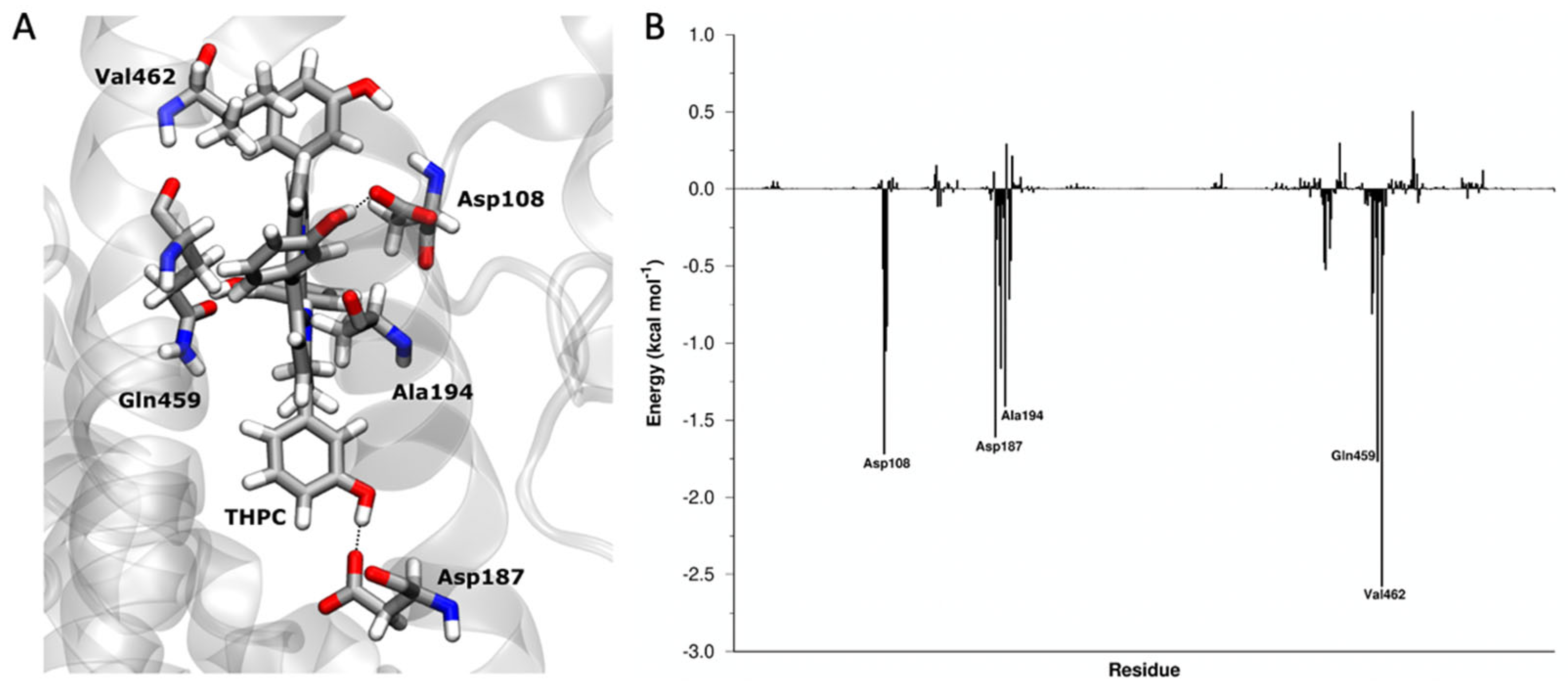
3.2. Atomistic Details of the mTHPC@HSA Complex
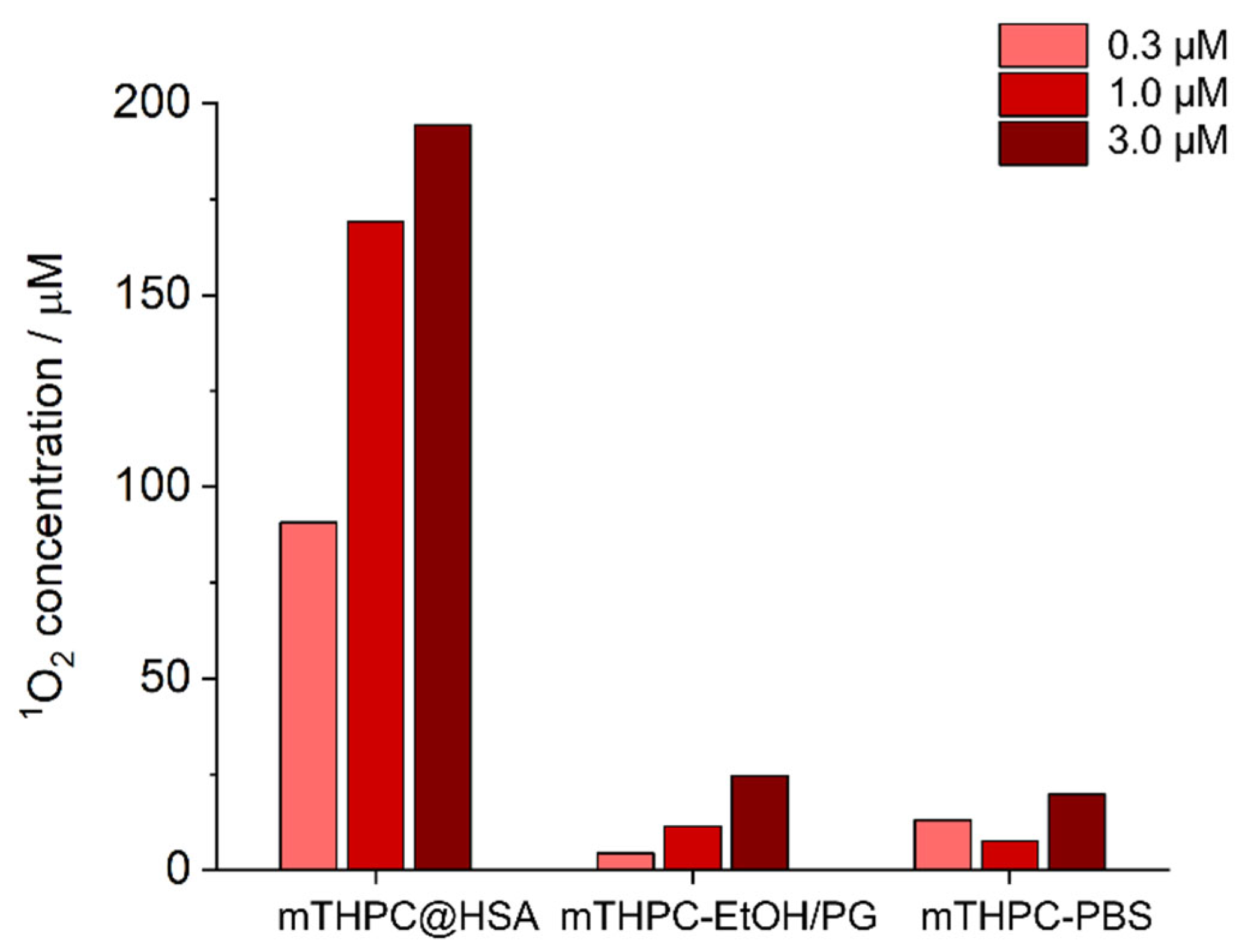
3.3. mTHPC@HSA Generates Singlet Oxygen in PBS
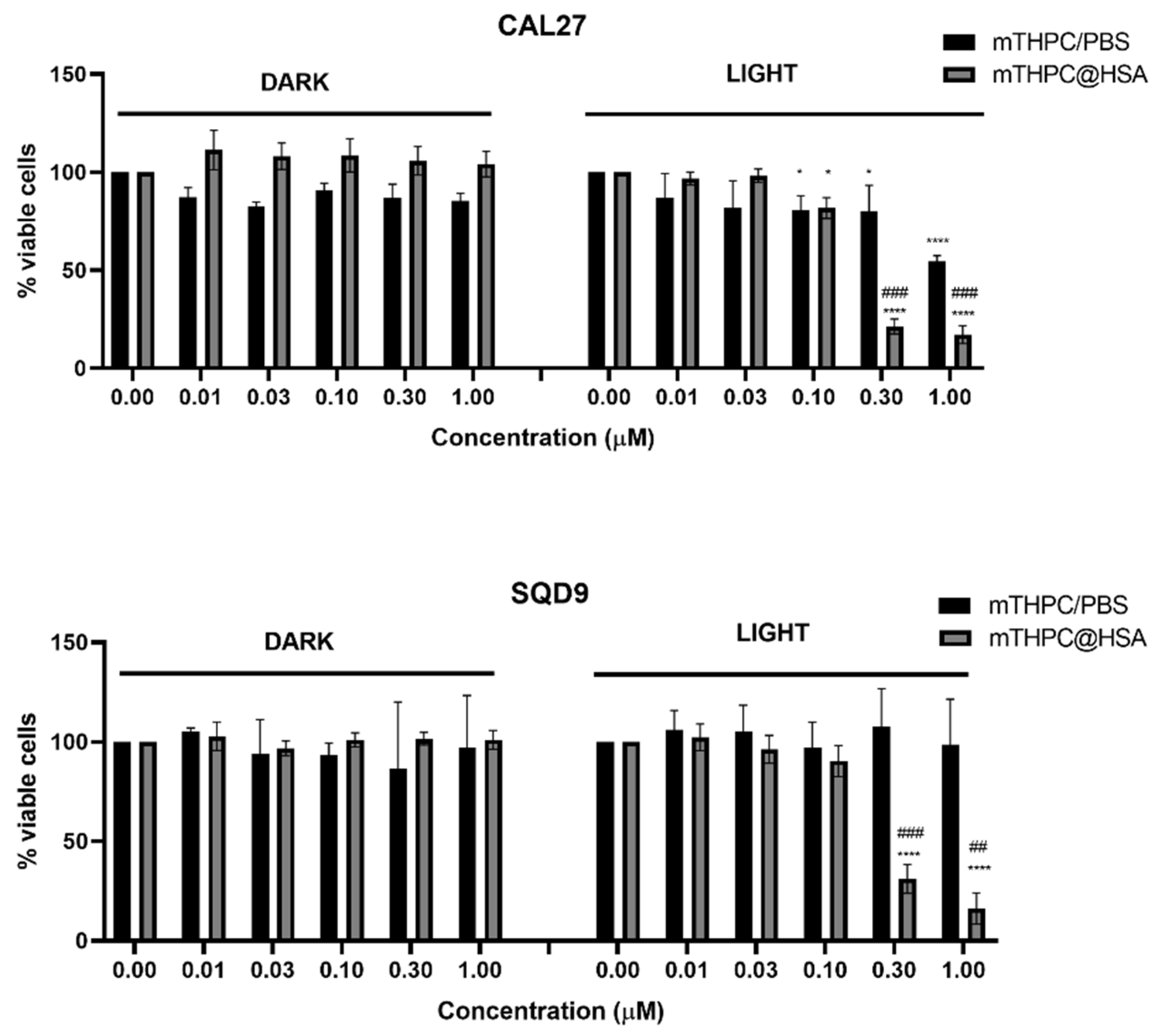
3.4. mTHPC@HSA Undergoes Intracellular Uptake and Decreases HNSCC Cells Viability upon PDT Treatment
3.5. mTHPC@HSA Increases Intracellular ROS Generation upon PDT Treatment
3.6. mTHPC@HSA Induces γ-H2AX upon PDT Treatment
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ibarra, A.M.C.; Cecatto, R.B.; Motta, L.J.; Dos Santos Franco, A.L.; de Fátima Teixeira da Silva, D.; Nunes, F.D.; Hamblin, M.R.; Rodrigues, M.F.S.D. Photodynamic Therapy for Squamous Cell Carcinoma of the Head and Neck: Narrative Review Focusing on Photosensitizers. Lasers Med. Sci. 2022, 37, 1441–1470. [Google Scholar] [CrossRef] [PubMed]
- Global Cancer Observatory; International Agency for Research on Cancer; World Health Organization. Available online: https://gco.iarc.fr/ (accessed on 15 October 2022).
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and Neck Squamous Cell Carcinoma. Nat. Rev. Dis. Primer 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Biel, M.A. Photodynamic Therapy of Head and Neck Cancers. Methods Mol. Biol. 2010, 635, 281–293. [Google Scholar]
- Brown, S.B.; Brown, E.A.; Walker, I. The Present and Future Role of Photodynamic Therapy in Cancer Treatment. Lancet Oncol. 2004, 5, 497–508. [Google Scholar] [CrossRef]
- Senge, M.O.; Brandt, J.C. Temoporfin (Foscan®, 5,10,15,20-Tetra(m-Hydroxyphenyl)Chlorin)—A Second-Generation Photosensitizer. Photochem. Photobiol. 2011, 87, 1240–1296. [Google Scholar] [CrossRef] [PubMed]
- EMA. Foscan. Eur. Med. Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/foscan (accessed on 15 October 2022).
- Yakavets, I.; Millard, M.; Zorin, V.; Lassalle, H.-P.; Bezdetnaya, L. Current State of the Nanoscale Delivery Systems for Temoporfin-Based Photodynamic Therapy: Advanced Delivery Strategies. J. Control. Release 2019, 304, 268–287. [Google Scholar] [CrossRef]
- Kratz, F. A Clinical Update of Using Albumin as a Drug Vehicle—A Commentary. J. Control. Release 2014, 190, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Hoogenboezem, E.N.; Duvall, C.L. Harnessing Albumin as a Carrier for Cancer Therapies. Adv. Drug Deliv. Rev. 2018, 130, 73–89. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, Z. Albumin Carriers for Cancer Theranostics: A Conventional Platform with New Promise. Adv. Mater. Deerfield Beach Fla 2016, 28, 10557–10566. [Google Scholar] [CrossRef]
- Rubio-Camacho, M.; Encinar, J.A.; Martínez-Tomé, M.J.; Esquembre, R.; Mateo, C.R. The Interaction of Temozolomide with Blood Components Suggests the Potential Use of Human Serum Albumin as a Biomimetic Carrier for the Drug. Biomolecules 2020, 10, 1015. [Google Scholar] [CrossRef]
- Comas-Barceló, J.; Rodríguez-Amigo, B.; Abbruzzetti, S.; del Rey-Puech, P.; Agut, M.; Nonell, S.; Viappiani, C. A Self-Assembled Nanostructured Material with Photosensitising Properties. RSC Adv. 2013, 3, 17874–17879. [Google Scholar] [CrossRef]
- Soldà, A.; Cantelli, A.; Giosia, M.D.; Montalti, M.; Zerbetto, F.; Rapino, S.; Calvaresi, M. C60@lysozyme: A New Photosensitizing Agent for Photodynamic Therapy. J. Mater. Chem. B 2017, 5, 6608–6615. [Google Scholar] [CrossRef] [PubMed]
- Giosia, M.D.; Bomans, P.H.H.; Bottoni, A.; Cantelli, A.; Falini, G.; Franchi, P.; Guarracino, G.; Friedrich, H.; Lucarini, M.; Paolucci, F.; et al. Proteins as Supramolecular Hosts for C60: A True Solution of C60 in Water. Nanoscale 2018, 10, 9908–9916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giosia, M.; Valle, F.; Cantelli, A.; Bottoni, A.; Zerbetto, F.; Calvaresi, M. C60 Bioconjugation with Proteins: Towards a Palette of Carriers for All PH Ranges. Materials 2018, 11, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giosia, M.; Genovese, D.; Cantelli, A.; Cingolani, M.; Rampazzo, E.; Strever, G.; Tavoni, M.; Zaccheroni, N.; Calvaresi, M.; Prodi, L. Synthesis and Characterization of a Reconstituted Myoglobin-Chlorin E6 Adduct for Theranostic Applications. J. Porphyr. Phthalocyanines 2020, 24, 887–893. [Google Scholar] [CrossRef]
- Di Giosia, M.; Soldà, A.; Seeger, M.; Cantelli, A.; Arnesano, F.; Nardella, M.I.; Mangini, V.; Valle, F.; Montalti, M.; Zerbetto, F.; et al. A Bio-Conjugated Fullerene as a Subcellular-Targeted and Multifaceted Phototheranostic Agent. Adv. Funct. Mater. 2021, 31, 2101527. [Google Scholar] [CrossRef]
- Delcanale, P.; Montali, C.; Rodríguez-Amigo, B.; Abbruzzetti, S.; Bruno, S.; Bianchini, P.; Diaspro, A.; Agut, M.; Nonell, S.; Viappiani, C. Zinc-Substituted Myoglobin Is a Naturally Occurring Photo-Antimicrobial Agent with Potential Applications in Food Decontamination. J. Agric. Food Chem. 2016, 64, 8633–8639. [Google Scholar] [CrossRef]
- Delcanale, P.; Rodríguez-Amigo, B.; Juárez-Jiménez, J.; Luque, F.J.; Abbruzzetti, S.; Agut, M.; Nonell, S.; Viappiani, C. Tuning the Local Solvent Composition at a Drug Carrier Surface: The Effect of Dimethyl Sulfoxide/Water Mixture on the Photofunctional Properties of Hypericin–β-Lactoglobulin Complexes. J. Mater. Chem. B 2017, 5, 1633–1641. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, M.; Delcanale, P.; Montali, C.; Tognolini, M.; Giorgio, C.; Corrado, M.; Cavanna, L.; Bianchini, P.; Diaspro, A.; Abbruzzetti, S.; et al. Enhanced Photosensitizing Properties of Protein Bound Curcumin. Life Sci. 2019, 233, 116710. [Google Scholar] [CrossRef]
- Rodríguez-Amigo, B.; Hally, C.; Roig-Yanovsky, N.; Delcanale, P.; Abbruzzetti, S.; Agut, M.; Viappiani, C.; Nonell, S. A Double Payload Complex between Hypericin and All-Trans Retinoic Acid in the β-Lactoglobulin Protein. Antibiotics 2022, 11, 282. [Google Scholar] [CrossRef]
- Zhang, P.; Huang, H.; Banerjee, S.; Clarkson, G.J.; Ge, C.; Imberti, C.; Sadler, P.J. Nucleus-Targeted Organoiridium–Albumin Conjugate for Photodynamic Cancer Therapy. Angew. Chem. Int. Ed. 2019, 58, 2350–2354. [Google Scholar] [CrossRef]
- Jeong, H.; Huh, M.; Lee, S.J.; Koo, H.; Kwon, I.C.; Jeong, S.Y.; Kim, K. Photosensitizer-Conjugated Human Serum Albumin Nanoparticles for Effective Photodynamic Therapy. Theranostics 2011, 1, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantelli, A.; Malferrari, M.; Soldà, A.; Simonetti, G.; Forni, S.; Toscanella, E.; Mattioli, E.J.; Zerbetto, F.; Zanelli, A.; Di Giosia, M.; et al. Human Serum Albumin–Oligothiophene Bioconjugate: A Phototheranostic Platform for Localized Killing of Cancer Cells by Precise Light Activation. JACS Au 2021, 1, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Cantelli, A.; Malferrari, M.; Mattioli, E.J.; Marconi, A.; Mirra, G.; Soldà, A.; Marforio, T.D.; Zerbetto, F.; Rapino, S.; Di Giosia, M.; et al. Enhanced Uptake and Phototoxicity of C60@albumin Hybrids by Folate Bioconjugation. Nanomaterials 2022, 12, 3501. [Google Scholar] [CrossRef] [PubMed]
- Rapozzi, V.; Moret, F.; Menilli, L.; Guerrini, A.; Tedesco, D.; Naldi, M.; Bartolini, M.; Gani, M.; Zorzet, S.; Columbaro, M.; et al. HSA-Binding Prodrugs-Based Nanoparticles Endowed with Chemo and Photo-Toxicity against Breast Cancer. Cancers 2022, 14, 877. [Google Scholar] [CrossRef]
- Hu, D.; Sheng, Z.; Gao, G.; Siu, F.; Liu, C.; Wan, Q.; Gong, P.; Zheng, H.; Ma, Y.; Cai, L. Activatable Albumin-Photosensitizer Nanoassemblies for Triple-Modal Imaging and Thermal-Modulated Photodynamic Therapy of Cancer. Biomaterials 2016, 93, 10–19. [Google Scholar] [CrossRef]
- Krumkacheva, O.A.; Timofeev, I.O.; Politanskaya, L.V.; Polienko, Y.F.; Tretyakov, E.V.; Rogozhnikova, O.Y.; Trukhin, D.V.; Tormyshev, V.M.; Chubarov, A.S.; Bagryanskaya, E.G.; et al. Triplet Fullerenes as Prospective Spin Labels for Nanoscale Distance Measurements by Pulsed Dipolar EPR Spectroscopy. Angew. Chem. Int. Ed. 2019, 58, 13271–13275. [Google Scholar] [CrossRef]
- Parodi, A.; Miao, J.; Soond, S.M.; Rudzińska, M.; Zamyatnin, A.A. Albumin Nanovectors in Cancer Therapy and Imaging. Biomolecules 2019, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.A.; Berman, M.C. Kinetics and Mechanism of the Interaction between Human Serum Albumin and Monomeric Haemin. Biochem. J. 1980, 191, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for Rigid and Symmetric Docking. Nucleic Acids Res. 2005, 33, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. Ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from Ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Case, D.A.; Betz, R.M.; Bottello-Smith, W.; Cerutti, D.S.; Cheatham, I.; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; et al. Amber 16; University of California: San Francisco, CA, USA, 2016. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Miller, B.R.I.; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.Py: An Efficient Program for End-State Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Chen, J.Y.; Mak, N.K.; Yow, C.M.N.; Fung, M.C.; Chiu, L.C.; Leung, W.N.; Cheung, N.H. The Binding Characteristics and Intracellular Localization of Temoporfin (MTHPC) in Myeloid Leukemia Cells: Phototoxicity and Mitochondrial Damage. Photochem. Photobiol. 2000, 72, 541–547. [Google Scholar] [CrossRef]
- Stein, N.C.; Mulac, D.; Fabian, J.; Herrmann, F.C.; Langer, K. Nanoparticle Albumin-Bound MTHPC for Photodynamic Therapy: Preparation and Comprehensive Characterization of a Promising Drug Delivery System. Int. J. Pharm. 2020, 582, 119347. [Google Scholar] [CrossRef]
- An, W.; Jiao, Y.; Dong, C.; Yang, C.; Inoue, Y.; Shuang, S. Spectroscopic and Molecular Modeling of the Binding of Meso-Tetrakis(4-Hydroxyphenyl)Porphyrin to Human Serum Albumin. Dyes Pigments 2009, 81, 1–9. [Google Scholar] [CrossRef]
- Synak, A.; Ziǒłek, M.; Organero, J.A.; Douhal, A. Femtosecond Dynamics of a Porphyrin Derivative Confined by the Human Serum Albumin Protein. J. Phys. Chem. B 2010, 114, 16567–16573. [Google Scholar] [CrossRef]
- Sannikova, N.E.; Timofeev, I.O.; Chubarov, A.S.; Lebedeva, N.S.; Semeikin, A.S.; Kirilyuk, I.A.; Tsentalovich, Y.P.; Fedin, M.V.; Bagryanskaya, E.G.; Krumkacheva, O.A. Application of EPR to Porphyrin-Protein Agents for Photodynamic Therapy. J. Photochem. Photobiol. B 2020, 211, 112008. [Google Scholar] [CrossRef]
- Li, X.; Li, D.; Han, M.; Chen, Z.; Zou, G. Neutral Porphyrin J-Aggregates in Premicellar SDS Solution. Colloids Surf. Physicochem. Eng. Asp. 2005, 256, 151–156. [Google Scholar] [CrossRef]
- Ma, L.; Moan, J.; Berg, K. Evaluation of a New Photosensitizer, Meso-Tetra-Hydroxyphenyl-Chlorin, for Use in Photodynamic Therapy: A Comparison of Its Photobiological Properties with Those of Two Other Photosensitizers. Int. J. Cancer 1994, 57, 883–888. [Google Scholar] [CrossRef]
- Belitchenko, I.; Melnikova, V.; Bezdetnaya, L.; Rezzoug, H.; Merlin, J.L.; Potapenko, A.; Guillemin, F. Characterization of Photodegradation of Meta-Tetra (Hydroxyphenyl)Chlorin (MTHPC) in Solution: Biological Consequences in Human Tumor Cells. Photochem. Photobiol. 1998, 67, 584–590. [Google Scholar] [CrossRef]
- Li, X.; Yu, S.; Lee, Y.; Guo, T.; Kwon, N.; Lee, D.; Yeom, S.C.; Cho, Y.; Kim, G.; Huang, J.-D.; et al. In Vivo Albumin Traps Photosensitizer Monomers from Self-Assembled Phthalocyanine Nanovesicles: A Facile and Switchable Theranostic Approach. J. Am. Chem. Soc. 2019, 141, 1366–1372. [Google Scholar] [CrossRef]
- Li, X.; Jeong, K.; Lee, Y.; Guo, T.; Lee, D.; Park, J.; Kwon, N.; Na, J.-H.; Hong, S.K.; Cha, S.-S.; et al. Water-Soluble Phthalocyanines Selectively Bind to Albumin Dimers: A Green Approach Toward Enhancing Tumor-Targeted Photodynamic Therapy. Theranostics 2019, 9, 6412–6423. [Google Scholar] [CrossRef]
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal Structure of Human Serum Albumin at 2.5 Å Resolution. Protein Eng. Des. Sel. 1999, 12, 439–446. [Google Scholar] [CrossRef]
- Di Giosia, M.; Zerbetto, F.; Calvaresi, M. Incorporation of Molecular Nanoparticles Inside Proteins: The Trojan Horse Approach in Theranostics. Acc. Mater. Res. 2021, 2, 594–605. [Google Scholar] [CrossRef]
- Cantelli, A.; Piro, F.; Pecchini, P.; Di Giosia, M.; Danielli, A.; Calvaresi, M. Concanavalin A-Rose Bengal Bioconjugate for Targeted Gram-Negative Antimicrobial Photodynamic Therapy. J. Photochem. Photobiol. B 2020, 206, 111852. [Google Scholar] [CrossRef]
- Yogo, T.; Urano, Y.; Mizushima, A.; Sunahara, H.; Inoue, T.; Hirose, K.; Iino, M.; Kikuchi, K.; Nagano, T. Selective Photoinactivation of Protein Function through Environment-Sensitive Switching of Singlet Oxygen Generation by Photosensitizer. Proc. Natl. Acad. Sci. USA 2008, 105, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Malferrari, M.; Turina, P.; Francia, F.; Mezzetti, A.; Leibl, W.; Venturoli, G. Dehydration Affects the Electronic Structure of the Primary Electron Donor in Bacterial Photosynthetic Reaction Centers: Evidence from Visible-NIR and Light-Induced Difference FTIR Spectroscopy. Photochem. Photobiol. Sci. 2015, 14, 238–251. [Google Scholar] [CrossRef] [Green Version]
- Lange, C.; Lehmann, C.; Mahler, M.; Bednarski, P.J. Comparison of Cellular Death Pathways after MTHPC-Mediated Photodynamic Therapy (PDT) in Five Human Cancer Cell Lines. Cancers 2019, 11, 702. [Google Scholar] [CrossRef] [Green Version]
- Tan, I.B.; Dolivet, G.; Ceruse, P.; Poorten, V.V.; Roest, G.; Rauschning, W. Temoporfin-Mediated Photodynamic Therapy in Patients with Advanced, Incurable Head and Neck Cancer: A Multicenter Study. Head Neck 2010, 32, 1597–1604. [Google Scholar] [CrossRef]
- Gamcsik, M.P.; Kasibhatla, M.S.; Teeter, S.D.; Colvin, O.M. Glutathione Levels in Human Tumors. Biomarkers 2012, 17, 671–691. [Google Scholar] [CrossRef] [Green Version]
- Sobhakumari, A.; Love-Homan, L.; Fletcher, E.V.M.; Martin, S.M.; Parsons, A.D.; Spitz, D.R.; Knudson, C.M.; Simons, A.L. Susceptibility of Human Head and Neck Cancer Cells to Combined Inhibition of Glutathione and Thioredoxin Metabolism. PloS One 2012, 7, e48175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firsanov, D.V.; Solovjeva, L.V.; Svetlova, M.P. H2AX Phosphorylation at the Sites of DNA Double-Strand Breaks in Cultivated Mammalian Cells and Tissues. Clin. Epigenetics 2011, 2, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yow, C.M.; Mak, N.K.; Szeto, S.; Chen, J.Y.; Lee, Y.L.; Cheung, N.H.; Huang, D.P.; Leung, A.W.N. Photocytotoxic and DNA Damaging Effect of Temoporfin (MTHPC) and Merocyanine 540 (MC540) on Nasopharyngeal Carcinoma Cell. Toxicol. Lett. 2000, 115, 53–61. [Google Scholar] [CrossRef]
- Rousset, N.; Kerninon, E.; Eléouet, S.; Le Néel, T.; Auget, J.-L.; Vonarx, V.; Carré, J.; Lajat, Y.; Patrice, T. Use of Alkaline Comet Assay to Assess DNA Repair after M-THPC-PDT. J. Photochem. Photobiol. B 2000, 56, 118–131. [Google Scholar] [CrossRef]
- Cadet, J.; Mouret, S.; Ravanat, J.-L.; Douki, T. Photoinduced Damage to Cellular DNA: Direct and Photosensitized Reactions. Photochem. Photobiol. 2012, 88, 1048–1065. [Google Scholar] [CrossRef]
- McNair, F.I.; Marples, B.; West, C.M.L.; Moore, J.V. A Comet Assay of DNA Damage and Repair in K562 Cells after Photodynamic Therapy Using Haematoporphyrin Derivative, Methylene Blue and Meso-Tetrahydroxyphenylchlorin. Br. J. Cancer 1997, 75, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Parsons, J.L. The Radiobiology of HPV-Positive and HPV-Negative Head and Neck Squamous Cell Carcinoma. Expert Rev. Mol. Med. 2020, 22, e3. [Google Scholar] [CrossRef]
- Park, C.R.; Jo, J.H.; Song, M.G.; Park, J.Y.; Kim, Y.-H.; Youn, H.; Paek, S.H.; Chung, J.K.; Jeong, J.M.; Lee, Y.-S.; et al. Secreted Protein Acidic and Rich in Cysteine Mediates Active Targeting of Human Serum Albumin in U87MG Xenograft Mouse Models. Theranostics 2019, 9, 7447–7457. [Google Scholar] [CrossRef]
- Yang, Y.; Niu, X.; Liu, W.; Xu, H. Expression and Significance of Secreted Protein Acidic and Rich in Cysteine in Human Osteosarcoma. Oncol. Lett. 2017, 14, 5491–5496. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, D.; Pan, J.; Xia, C.; Fan, L.; Pu, Y.; Zhang, Q.; hong Ni, Y.; Wang, J.; Hu, Q. A near Infrared Light-Triggered Human Serum Albumin Drug Delivery System with Coordination Bonding of Indocyanine Green and Cisplatin for Targeting Photochemistry Therapy against Oral Squamous Cell Cancer. Biomater. Sci. 2019, 7, 5270–5282. [Google Scholar] [CrossRef]
- Desai, N.; Trieu, V.; Damascelli, B.; Soon-Shiong, P. SPARC Expression Correlates with Tumor Response to Albumin-Bound Paclitaxel in Head and Neck Cancer Patients. Transl. Oncol. 2009, 2, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Rodriguez, E.M.; Naccache, R.; Forgione, P.; Lamoureux, G.; Sanz-Rodriguez, F.; Scheglmann, D.; Capobianco, J.A. Chemical Modification of Temoporfin—A Second Generation Photosensitizer Activated Using Upconverting Nanoparticles for Singlet Oxygen Generation. Chem. Commun. 2014, 50, 12150–12153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharman, W.M.; van Lier, J.E.; Allen, C.M. Targeted Photodynamic Therapy via Receptor Mediated Delivery Systems. Adv. Drug Deliv. Rev. 2004, 56, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Butzbach, K.; Rasse-Suriani, F.A.O.; Gonzalez, M.M.; Cabrerizo, F.M.; Epe, B. Albumin-Folate Conjugates for Drug-Targeting in Photodynamic Therapy. Photochem. Photobiol. 2016, 92, 611–619. [Google Scholar] [CrossRef]
- Ulfo, L.; Costantini, P.E.; Di Giosia, M.; Danielli, A.; Calvaresi, M. EGFR-Targeted Photodynamic Therapy. Pharmaceutics 2022, 14, 241. [Google Scholar] [CrossRef]
- Bortot, B.; Apollonio, M.; Baj, G.; Andolfi, L.; Zupin, L.; Crovella, S.; di Giosia, M.; Cantelli, A.; Saporetti, R.; Ulfo, L.; et al. Advanced Photodynamic Therapy with an Engineered M13 Phage Targeting EGFR: Mitochondrial Localization and Autophagy Induction in Ovarian Cancer Cell Lines. Free Radic. Biol. Med. 2022, 179, 242–251. [Google Scholar] [CrossRef]
- Ulfo, L.; Cantelli, A.; Petrosino, A.; Emidio Costantini, P.; Nigro, M.; Starinieri, F.; Turrini, E.; Khan Zadran, S.; Zuccheri, G.; Saporetti, R.; et al. Orthogonal Nanoarchitectonics of M13 Phage for Receptor Targeted Anticancer Photodynamic Therapy. Nanoscale 2022, 14, 632–641. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioli, E.J.; Ulfo, L.; Marconi, A.; Pellicioni, V.; Costantini, P.E.; Marforio, T.D.; Di Giosia, M.; Danielli, A.; Fimognari, C.; Turrini, E.; et al. Carrying Temoporfin with Human Serum Albumin: A New Perspective for Photodynamic Application in Head and Neck Cancer. Biomolecules 2023, 13, 68. https://doi.org/10.3390/biom13010068
Mattioli EJ, Ulfo L, Marconi A, Pellicioni V, Costantini PE, Marforio TD, Di Giosia M, Danielli A, Fimognari C, Turrini E, et al. Carrying Temoporfin with Human Serum Albumin: A New Perspective for Photodynamic Application in Head and Neck Cancer. Biomolecules. 2023; 13(1):68. https://doi.org/10.3390/biom13010068
Chicago/Turabian StyleMattioli, Edoardo Jun, Luca Ulfo, Alessia Marconi, Valentina Pellicioni, Paolo Emidio Costantini, Tainah Dorina Marforio, Matteo Di Giosia, Alberto Danielli, Carmela Fimognari, Eleonora Turrini, and et al. 2023. "Carrying Temoporfin with Human Serum Albumin: A New Perspective for Photodynamic Application in Head and Neck Cancer" Biomolecules 13, no. 1: 68. https://doi.org/10.3390/biom13010068