
Cucurbitacins as Potent Chemo-Preventive Agents: Mechanistic Insight and Recent Trends

 , ,
, ,  , ,
, ,  , ,
, , Manoj_Kumar.jpg) ,
,  , and
, and 
Abstract
:1. Introduction
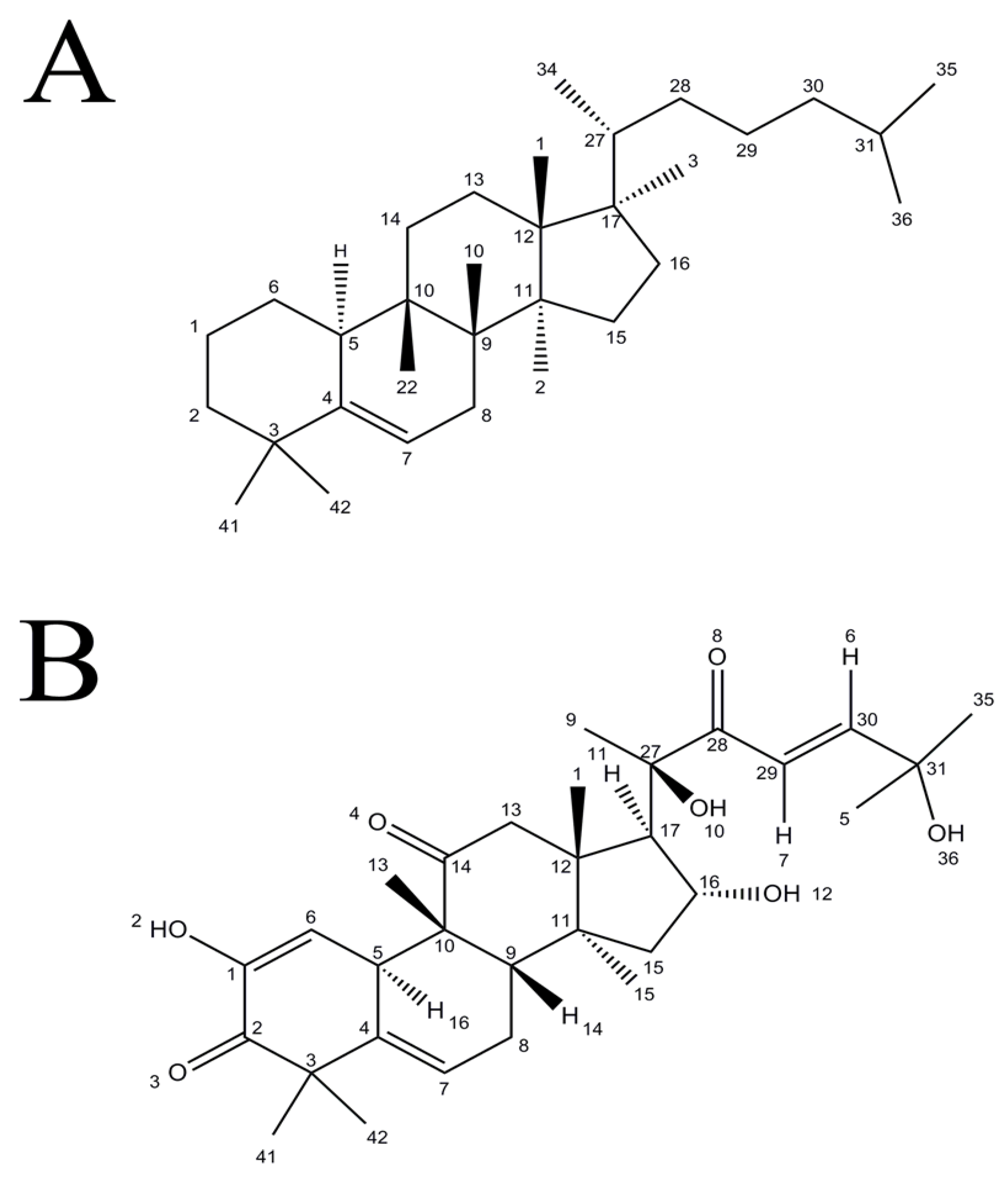
2. Chemistry of Cucurbitacins
3. Absorption and Metabolism of Cucurbitacins
4. Anti-Cancer Mechanisms of Cucurbitacins
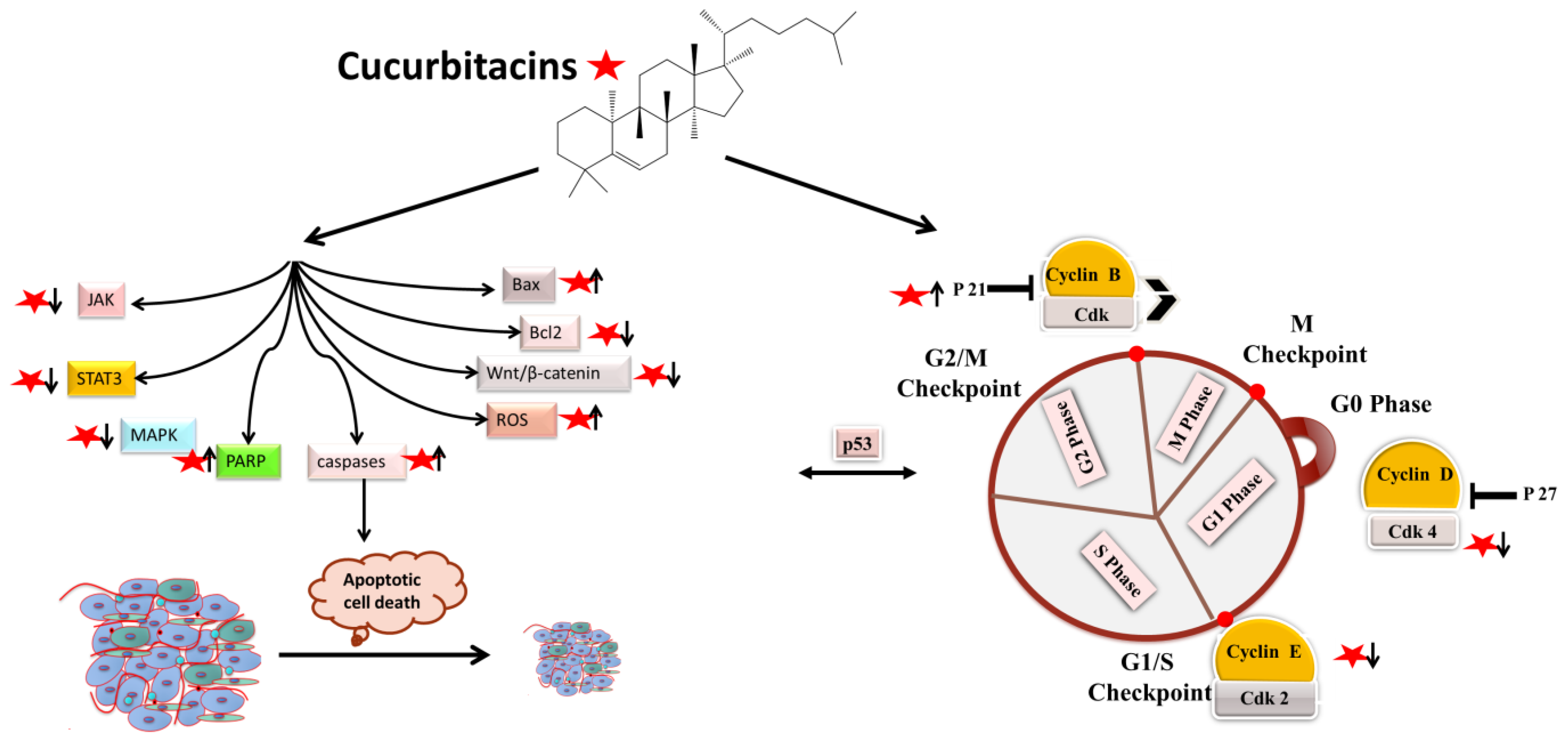
4.1. Apoptotic and Cell-Cycle Arrest
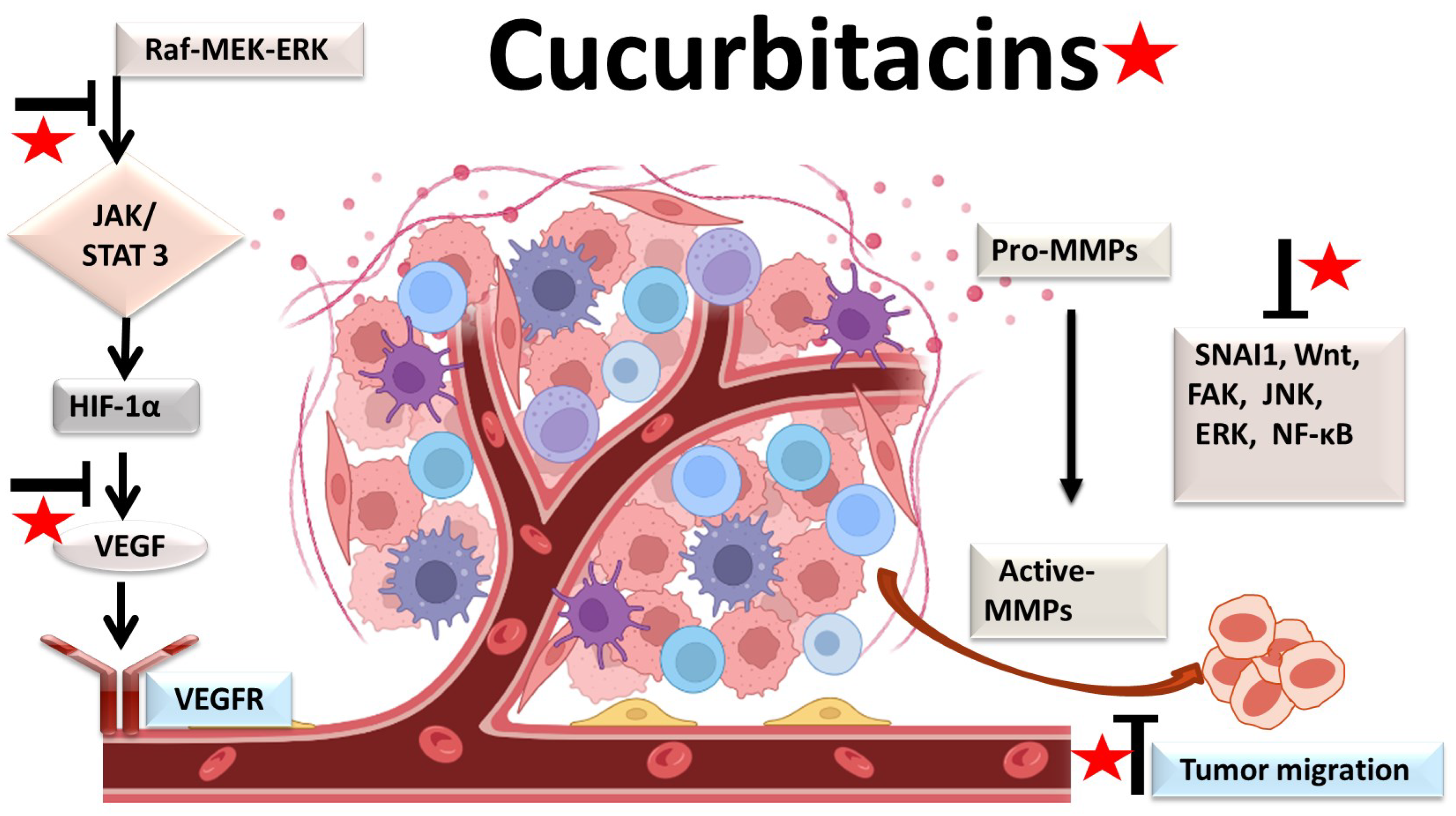
4.2. Antiangiogenic and Antimetastatic Mechanisms
4.3. Anti-Inflammatory Mechanisms
5. Synergistic Effects with Other Drugs
6. Safety Studies
7. Conclusions and Further Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Devlin, E.J.; Denson, L.A.; Whitford, H.S. Cancer Treatment Side Effects: A Meta-analysis of the Relationship Between Response Expectancies and Experience. J. Pain Symptom Manag. 2017, 54, 245–258.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, M.; Mellotte, G.; Ryan, B.; O’Connor, A. Gastrointestinal side effects of cancer treatments. Ther. Adv. Chronic Dis. 2020, 11, 2040622320970354. [Google Scholar] [CrossRef] [PubMed]
- Kirtonia, A.; Gala, K.; Fernandes, S.G.; Pandya, G.; Pandey, A.K.; Sethi, G.; Khattar, E.; Garg, M. Repurposing of drugs: An attractive pharmacological strategy for cancer therapeutics. Semin. Cancer Biol. 2021, 68, 258–278. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Sadaria, D.; Warrier, V.U.; Kirtonia, A.; Kant, R.; Awasthi, A.; Baligar, P.; Pal, J.K.; Yuba, E.; Sethi, G.; et al. Plant lectins and their usage in preparing targeted nanovaccines for cancer immunotherapy. Semin. Cancer Biol. 2022, 80, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Sadia, S.; Pan, K.; Ullah, I.; Mussarat, S.; Sun, F.; Abiodun, O.O.; Batbaatar, A.; Li, Z.; Song, D.; et al. A systematic review on ethnomedicines of anti-cancer plants. Phytother. Res. 2017, 31, 202–264. [Google Scholar] [CrossRef]
- Yang, S.F.; Weng, C.J.; Sethi, G.; Hu, D.N. Natural bioactives and phytochemicals serve in cancer treatment and prevention. Evid. Based. Complement. Alternat. Med. 2013, 2013, 698190. [Google Scholar] [CrossRef] [Green Version]
- Ren, B.; Kwah, M.X.Y.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.L.; Wang, L.; Ong, P.S.; et al. Resveratrol for cancer therapy: Challenges and future perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef]
- Sak, K. Anticancer action of plant products: Changing stereotyped attitudes. Explor. Target. Anti-Tumor Ther. 2022, 3, 423–427. [Google Scholar] [CrossRef]
- Liu, C.; Ho, P.C.L.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Dehshahri, A.; Ashrafizadeh, M.; Ghasemipour Afshar, E.; Pardakhty, A.; Mandegary, A.; Mohammadinejad, R.; Sethi, G. Topoisomerase inhibitors: Pharmacology and emerging nanoscale delivery systems. Pharmacol. Res. 2020, 151, 104551. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Que, R.; Liu, C.; Ji, W.; Sun, B.; Lin, X.; Zhang, Q.; Zhao, X.; Peng, Z.; Zhang, X.; et al. Survivin-targeted drug screening platform identifies a matrine derivative WM-127 as a potential therapeutics against hepatocellular carcinoma. Cancer Lett. 2018, 425, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, Z.; Lin, M.; Shang, Y.; Wang, F.; Zhou, J.Y.; Wang, F.; Zhang, X.; Luo, X.; Huang, W. In Vitro and In Vivo Antitumor Activity of Cucurbitacin C, a Novel Natural Product from Cucumber. Front. Pharmacol. 2019, 10, 1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Ma, B.; Lu, S.; Zeng, L.; Wang, H.; Shi, W.; Zhou, L.; Xia, Y.; Zhang, X.; Zhang, J.; et al. Cucumber-Derived Nanovesicles Containing Cucurbitacin B for Non-Small Cell Lung Cancer Therapy. Int. J. Nanomed. 2022, 17, 3583–3599. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Zhang, L.; Zhou, H.; Cao, L.; Shao, Y.; Li, T. Reactive Oxygen Species-Responsive Nanococktail With Self-Amplificated Drug Release for Efficient Co-Delivery of Paclitaxel/Cucurbitacin B and Synergistic Treatment of Gastric Cancer. Front. Chem. 2022, 10, 844426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, B.; Wei, H.Z.; Zeng, H.; Sheng, D.; Zhang, Y. Cucurbitacin B controls M2 macrophage polarization to suppresses metastasis via targeting JAK-2/STAT3 signalling pathway in colorectal cancer. J. Ethnopharmacol. 2022, 287, 114915. [Google Scholar] [CrossRef]
- Üremiş, N.; Üremiş, M.M.; Çiğremiş, Y.; Tosun, E.; Baysar, A.; Türköz, Y. Cucurbitacin I exhibits anticancer efficacy through induction of apoptosis and modulation of JAK/STAT3, MAPK/ERK, and AKT/mTOR signaling pathways in HepG2 cell line. J. Food Biochem. 2022, 46, e14333. [Google Scholar] [CrossRef]
- Xu, D.; Shen, H.; Tian, M.; Chen, W.; Zhang, X. Cucurbitacin I inhibits the proliferation of pancreatic cancer through the JAK2/STAT3 signalling pathway in vivo and in vitro. J. Cancer 2022, 13, 2050–2060. [Google Scholar] [CrossRef]
- Li, R.; Xiao, J.; Tang, S.; Lin, X.; Xu, H.; Han, B.; Yang, M.; Liu, F. Cucurbitacin I induces apoptosis in ovarian cancer cells through oxidative stress and the p190B-Rac1 signaling axis. Mol. Med. Rep. 2020, 22, 2545–2550. [Google Scholar] [CrossRef]
- Vidal-Gutiérrez, M.; Torres-Moreno, H.; Arenas-Luna, V.; Loredo-Mendoza, M.L.; Tejeda-Dominguez, F.; Velazquez, C.; Vilegas, W.; Hernández-Gutiérrez, S.; Robles-Zepeda, R.E. Standardized phytopreparations and cucurbitacin IIb from Ibervillea sonorae (S. Watson) greene induce apoptosis in cervical cancer cells by Nrf2 inhibition. J. Ethnopharmacol. 2022, 298, 115606. [Google Scholar] [CrossRef] [PubMed]
- Aiswarya, S.U.D.; Vikas, G.; Haritha, N.H.; Liju, V.B.; Shabna, A.; Swetha, M.; Rayginia, T.P.; Keerthana, C.K.; Nath, L.R.; Reshma, M.V.; et al. Cucurbitacin B, Purified and Characterized From the Rhizome of Corallocarpus epigaeus Exhibits Anti-Melanoma Potential. Front. Oncol. 2022, 12, 903832. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.T.B.; Font, R.; Gómez, P.; Río Celestino, M. Del Summer squash. Nutr. Compos. Antioxid. Prop. Fruits Veg. 2020, 239–254. [Google Scholar] [CrossRef]
- Kaushik, U.; Aeri, V.; Mir, S.R. Cucurbitacins–An insight into medicinal leads from nature. Pharmacogn. Rev. 2015, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, C.C.; Ming, H.C.; Rui, L.N.; Cordel, G.A.; Qiuz, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef]
- Chan, K.T.; Meng, F.Y.; Li, Q.; Ho, C.Y.; Lam, T.S.; To, Y.; Lee, W.H.; Li, M.; Chu, K.H.; Toh, M. Cucurbitacin B induces apoptosis and S phase cell cycle arrest in BEL-7402 human hepatocellular carcinoma cells and is effective via oral administration. Cancer Lett. 2010, 294, 118–124. [Google Scholar] [CrossRef]
- Ding, T.; Zhang, Y.; Chen, A.; Tang, Y.; Liu, M.; Wang, X. Effects of Cucurbitacin E, a Tetracyclic Triterpene Compound from Cucurbitaceae, on the Pharmacokinetics and Pharmacodynamics of Warfarin in Rats. Basic Clin. Pharmacol. Toxicol. 2015, 116, 385–389. [Google Scholar] [CrossRef]
- Hunsakunachai, N.; Nuengchamnong, N.; Jiratchariyakul, W.; Kummalue, T.; Khemawoot, P. Pharmacokinetics of cucurbitacin B from Trichosanthes cucumerina L. in rats. BMC Complement. Altern. Med. 2019, 19, 157. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Fu, L.; Zhu, Z.; Yang, Y.; Sun, B.; Shan, W.; Zhang, Z. Modified mixed nanomicelles with collagen peptides enhanced oral absorption of Cucurbitacin B: Preparation and evaluation. Drug Deliv. 2019, 25, 862–871. [Google Scholar] [CrossRef] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Li, F.; Chen, L.; Siveen, K.S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J. Mol. Med. 2014, 92, 267–276. [Google Scholar] [CrossRef]
- Sethi, G.; Kwang, S.A.; Sandur, S.K.; Lin, X.; Chaturvedi, M.M.; Aggarwal, B.B. Indirubin enhances tumor necrosis factor-induced apoptosis through modulation of nuclear factor-kappa B signaling pathway. J. Biol. Chem. 2006, 281, 23425–23435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.Y.; Um, J.Y.; Chinnathambi, A.; Govindasamy, C.; Sethi, G.; Ahn, K.S. Leelamine Modulates STAT5 Pathway Causing Both Autophagy and Apoptosis in Chronic Myelogenous Leukemia Cells. Biology 2022, 11, 366. [Google Scholar] [CrossRef] [PubMed]
- Chopra, P.; Sethi, G.; Dastidar, S.G.; Ray, A. Polo-like kinase inhibitors: An emerging opportunity for cancer therapeutics. Expert Opin. Investig. Drugs 2010, 19, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Li, J.; Hu, J.; Wang, L.; Huang, J.R.; Sethi, G.; Ma, Z. Circular RNAs in cell cycle regulation: Mechanisms to clinical significance. Cell Prolif. 2021, 54, 13143. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, A.; Sundarraj, K.; Arfuso, F.; Sethi, G.; Perumal, E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers 2018, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Xiang, X.; Li, S.; Xie, P.; Gong, Q.; Goh, B.C.; Wang, L. Targeting hypoxia-inducible factor-1, for cancer treatment: Recent advances in developing small-molecule inhibitors from natural compounds. Semin. Cancer Biol. 2022, 80, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Ong, T.H.; Chen, L.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Sharma, A.; Kumar, A.P.; et al. Suppression of signal transducer and activator of transcription 3 activation by butein inhibits growth of human hepatocellular carcinoma in vivo. Clin. Cancer Res. 2011, 17, 1425–1439. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Simvastatin potentiates TNF-alpha-induced apoptosis through the down-regulation of NF-kappaB-dependent antiapoptotic gene products: Role of IkappaBalpha kinase and TGF-beta-activated kinase-1. J. Immunol. 2007, 178, 2507–2516. [Google Scholar] [CrossRef] [Green Version]
- Mohan, C.D.; Rangappa, S.; Nayak, S.C.; Jadimurthy, R.; Wang, L.; Sethi, G.; Garg, M.; Rangappa, K.S. Bacteria as a treasure house of secondary metabolites with anticancer potential. Semin. Cancer Biol. 2021, 86, 998–1013. [Google Scholar] [CrossRef]
- Li, H.; Chen, H.; Li, R.; Xin, J.; Wu, S.; Lan, J.; Xue, K.; Li, X.; Zuo, C.; Jiang, W.; et al. Cucurbitacin I induces cancer cell death through the endoplasmic reticulum stress pathway. J. Cell. Biochem. 2018, 120, 2391–2403. [Google Scholar] [CrossRef]
- Mao, D.; Liu, A.H.; Wang, Z.P.; Zhang, X.W.; Lu, H. Cucurbitacin B inhibits cell proliferation and induces cell apoptosis in colorectal cancer by modulating methylation status of BTG3. Neoplasma 2019, 66, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Chen, J.; Zhou, Z.; Xia, H.; Qiu, M.H.; Chen, C. Cucurbitacin E induces cell cycle G2/M phase arrest and apoptosis in triple negative breast cancer. PLoS ONE 2014, 9, e103760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Lee, K.; Ku, J.M.; Choi, Y.J.; Mok, K.; Kim, D.; Cheon, C.; Ko, S.G. Cucurbitacin D Induces G2/M Phase Arrest and Apoptosis via the ROS/p38 Pathway in Capan-1 Pancreatic Cancer Cell Line. Evid. Based. Complement. Alternat. Med. 2020, 2020, 6571674. [Google Scholar] [CrossRef] [PubMed]
- Duangmano, S.; Sae-Lim, P.; Suksamrarn, A.; Patmasiriwat, P.; Domann, F.E. Cucurbitacin B Causes Increased Radiation Sensitivity of Human Breast Cancer Cells via G2/M Cell Cycle Arrest. J. Oncol. 2012, 2012, 601682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.W.; Yang, J.S.; Lin, M.W.; Chen, P.Y.; Chiou, S.M.; Chueh, F.S.; Lan, Y.H.; Pai, S.J.; Tsuzuki, M.; Ho, W.J.; et al. Cucurbitacin E Induces G(2)/M Phase Arrest through STAT3/p53/p21 Signaling and Provokes Apoptosis via Fas/CD95 and Mitochondria-Dependent Pathways in Human Bladder Cancer T24 Cells. Evid. Based. Complement. Alternat. Med. 2012, 2012, 952762. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, M.; Zhang, H.; Sun, C.; Deng, Y. Inhibitory effects of cucurbitacin B on laryngeal squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2008, 265, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Blaskovich, M.A.; Sun, J.; Cantor, A.; Turkson, J.; Jove, R.; Sebti, S.M. Discovery of JSI-124 (cucurbitacin I), a selective Janus kinase/signal transducer and activator of transcription 3 signaling pathway inhibitor with potent antitumor activity against human and murine cancer cells in mice-PubMed. Cancer Res. 2003, 63, 1270–1279. [Google Scholar] [PubMed]
- Liang, J.; Chen, D. Advances in research on the anticancer mechanism of the natural compound cucurbitacin from Cucurbitaceae plants: A review. Tradit. Chin. Med. 2019, 4, 68. [Google Scholar] [CrossRef]
- Dong, Y.; Lu, B.; Zhang, X.; Zhang, J.; Lai, L.; Li, D.; Wu, Y.; Song, Y.; Luo, J.; Pang, X.; et al. Cucurbitacin E, a tetracyclic triterpenes compound from Chinese medicine, inhibits tumor angiogenesis through VEGFR2-mediated Jak2-STAT3 signaling pathway. Carcinogenesis 2010, 31, 2097–2104. [Google Scholar] [CrossRef] [Green Version]
- Torres-Moreno, H.; Marcotullio, M.C.; Velázquez, C.; Ianni, F.; Garibay-Escobar, A.; Robles-Zepeda, R.E. Cucurbitacin IIb, a steroidal triterpene from Ibervillea sonorae induces antiproliferative and apoptotic effects on cervical and lung cancer cells. Steroids 2020, 157, 108597. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, T.; Ren, L.; Jing, S.; Li, Z.; Zuo, P.; Li, T.; Wang, Y.; Zhang, J.; Wei, Z. Cucurbitacin IIb induces apoptosis and cell cycle arrest through regulating EGFR/MAPK pathway. Environ. Toxicol. Pharmacol. 2021, 81, 103542. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Farooqi, A.A. Cucurbitacin mediated regulation of deregulated oncogenic signaling cascades and non-coding RNAs in different cancers: Spotlight on JAK/STAT, Wnt/β-catenin, mTOR, TRAIL-mediated pathways. Semin. Cancer Biol. 2021, 73, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Li, C.; Cao, L.; Zhang, C.H.; Zhang, Z.H. Cucurbitacin B regulates lung cancer cell proliferation and apoptosis via inhibiting the IL-6/STAT3 pathway through the lncRNA XIST/miR-let-7c axis. Pharm. Biol. 2022, 60, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Siveen, K.S.; Ahn, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Yap, W.N.; Kumar, A.P.; Fong, C.W.; Tergaonkar, V.; Hui, K.M.; et al. Y-tocotrienol inhibits angiogenesis-dependent growth of human hepatocellular carcinoma through abrogation of AKT/mTOR pathway in an orthotopic mouse model. Oncotarget 2014, 5, 1897–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; You, L.; Nepovimova, E.; Heger, Z.; Wu, W.; Kuca, K.; Adam, V. Hypoxia-inducible factors: Master regulators of hypoxic tumor immune escape. J. Hematol. Oncol. 2022, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Piao, X.M.; Gao, F.; Zhu, J.X.; Wang, L.J.; Zhao, X.; Li, X.; Sheng, M.M.; Zhang, Y. Cucurbitacin B inhibits tumor angiogenesis by triggering the mitochondrial signaling pathway in endothelial cells. Int. J. Mol. Med. 2018, 42, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, I.; Sharma, V.; Tuli, H.S.; Aggarwal, D.; Sankhyan, A.; Vyas, P.; Sharma, A.K.; Bishayee, A. Cancer Chemoprevention by Flavonoids, Dietary Polyphenols and Terpenoids. Biointerface Res. Appl. Chem. 2021, 11, 8502–8537. [Google Scholar] [CrossRef]
- Yin, B.; Fang, D.M.; Zhou, X.L.; Gao, F. Natural products as important tyrosine kinase inhibitors. Eur. J. Med. Chem. 2019, 182, 111664. [Google Scholar] [CrossRef]
- Sinha, S.; Khan, S.; Shukla, S.; Lakra, A.D.; Kumar, S.; Das, G.; Maurya, R.; Meeran, S.M. Cucurbitacin B inhibits breast cancer metastasis and angiogenesis through VEGF-mediated suppression of FAK/MMP-9 signaling axis. Int. J. Biochem. Cell Biol. 2016, 77, 41–56. [Google Scholar] [CrossRef]
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (review). Int. J. Oncol. 2018, 52, 19–37. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Guo, Q.; Liu, T.; Jiang, Y.; Zhao, M.; Zeng, K.; Tu, P. Cucurbitacin E Inhibits Huh7 Hepatoma Carcinoma Cell Proliferation and Metastasis via Suppressing MAPKs and JAK/STAT3 Pathways. Molecules 2020, 25, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, M.; Hasani, M.; Ramezani, F.; Abdolmaleki, M.K. Cucurbitacins: A focus on Cucurbitacin E as a natural product and their biological activities. Pharm. Sci. 2020, 27, 1–13. [Google Scholar] [CrossRef]
- Yun, W.; Dan, W.; Liu, J.; Guo, X.; Li, M.; He, Q. Investigation of the Mechanism of Traditional Chinese Medicines in Angiogenesis through Network Pharmacology and Data Mining. Evid.-Based Complement. Altern. Med. 2021, 2021, 5539970. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Kinghorn, A.D. Natural Product Triterpenoids and Their Semi-Synthetic Derivatives with Potential Anticancer Activity. Planta Med. 2019, 85, 802–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathya, T.N.; Mehta, V.A.; Senthil, D.; Navaneethakrishnan, K.; Murugan, S.; Kumaravel, T. Cytotoxicity evaluation of CELNORM, a nutritional health supplement, on MCF7 breast cancer cells. Indian J. Sci. Technol. 2020, 13, 3070–3075. [Google Scholar] [CrossRef]
- Ong, P.S.; Wang, L.Z.; Dai, X.; Tseng, S.H.; Loo, S.J.; Sethi, G. Judicious Toggling of mTOR Activity to Combat Insulin Resistance and Cancer: Current Evidence and Perspectives. Front. Pharmacol. 2016, 7, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Wang, Y.Y.; Xin, H.W.; Wang, L.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Wang, H.; Tang, F.R.; Warrier, S.; et al. The expanding roles of long non-coding RNAs in the regulation of cancer stem cells. Int. J. Biochem. Cell Biol. 2019, 108, 17–20. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Mostafavi, E.; Aref, A.R.; Sethi, G.; Wang, L.; Tergaonkar, V. Non-coding RNA-based regulation of inflammation. Semin. Immunol. 2022, 59, 101606. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.; Garg, M.; Tergaonkar, V.; Tan, S.Y.; Sethi, G. Pharmacological significance of the non-canonical NF-κB pathway in tumorigenesis. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188449. [Google Scholar] [CrossRef]
- Cheng, J.T.; Wang, L.; Wang, H.; Tang, F.R.; Cai, W.Q.; Sethi, G.; Xin, H.W.; Ma, Z. Insights into Biological Role of LncRNAs in Epithelial-Mesenchymal Transition. Cells 2019, 8, 1178. [Google Scholar] [CrossRef]
- Cai, W.; Chen, Z.X.; Rane, G.; Singh, S.S.; Choo, Z.; Wang, C.; Yuan, Y.; Tan, T.Z.; Arfuso, F.; Yap, C.T.; et al. Wanted DEAD/H or Alive: Helicases Winding Up in Cancers. J. Natl. Cancer Inst. 2017, 109, djw278. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.K.L.; Shanmugam, M.K.; Fan, L.; Fraser, S.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Bishayee, A. Focus on Formononetin: Anticancer Potential and Molecular Targets. Cancers 2019, 11, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, N.; Ghorai, S.M.; Kushwaha, P.K.; Shukla, R.; Aggarwal, C.; Bandichhor, R. Plausible mechanisms explaining the role of cucurbitacins as potential therapeutic drugs against coronavirus 2019. Inform. Med. Unlocked 2020, 21, 100484. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Tuo, X.; Zhang, F.; Bowen, L.; Zhao, W.; Xu, Y. Dietary cucurbitacin E reduces high-strength altitude training induced oxidative stress, inflammation and immunosuppression. An. Acad. Bras. Cienc. 2020, 92, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.; Liu, X.; Lv, S.; Wang, Z. Protective Effects of Cucurbitacin B on Acute Lung Injury Induced by Sepsis in Rats. Med. Sci. Monit. 2017, 23, 1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Ao, Q.; Zhong, Q.; Li, W.; Li, W. SIRT1/IGFBPrP1/TGF β1 axis involved in cucurbitacin B ameliorating concanavalin A-induced mice liver fibrosis. Basic Clin. Pharmacol. Toxicol. 2020, 127, 371–379. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, M.J.; Kwon, J.E.; Jung, K.A.; Jhun, J.Y.; Lee, S.Y.; Seo, H.B.; Ryu, J.Y.; Beak, J.A.; Choi, J.Y.; et al. Cucurbitacin E ameliorates acute graft-versus-host disease by modulating Th17 cell subsets and inhibiting STAT3 activation. Immunol. Lett. 2018, 203, 62–69. [Google Scholar] [CrossRef]
- Shang, J.; Liu, W.; Yin, C.; Chu, H.; Zhang, M. Cucurbitacin E ameliorates lipopolysaccharide-evoked injury, inflammation and MUC5AC expression in bronchial epithelial cells by restraining the HMGB1-TLR4-NF-κB signaling. Mol. Immunol. 2019, 114, 571–577. [Google Scholar] [CrossRef]
- Yuan, R.; Zhao, W.; Wang, Q.Q.; He, J.; Han, S.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits non-small cell lung cancer in vivo and in vitro by triggering TLR4/NLRP3/GSDMD-dependent pyroptosis. Pharmacol. Res. 2021, 170, 105748. [Google Scholar] [CrossRef]
- Wang, X.; Bai, Y.; Yan, X.; Li, J.; Lin, B.; Dai, L.; Xu, C.; Li, H.; Li, D.; Yang, T.; et al. Cucurbitacin B exhibits antitumor effects on CD133+ HepG2 liver cancer stem cells by inhibiting JAK2/STAT3 signaling pathway. Anticancer. Drugs 2021, 32, 548–557. [Google Scholar] [CrossRef]
- Sadzuka, Y.; Fujiki, S.; Itai, S. Enhancement of doxorubicin-induced antitumor activity and reduction of adverse reactions by cucurbitacin I. Food Res. Int. 2012, 47, 64–69. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.; Zhou, J.; Huang, Z.; Hu, H.; Qiao, M.; Zhao, X.; Chen, D. Synergistic effect of cucurbitacin B in combination with curcumin via enhancing apoptosis induction and reversing multidrug resistance in human hepatoma cells. Eur. J. Pharmacol. 2015, 768, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Eyol, E.; Tanrıverdi, Z.; Karakuş, F.; Yılmaz, K.; Ünüvar, S. Synergistic Anti-proliferative Effects of Cucurbitacin I and Irinotecan on Human Colorectal Cancer Cell Lines. J. Clin. Exp. Pharmacol. 2016, 6, 1000219. [Google Scholar] [CrossRef]
- Sikander, M.; Malik, S.; Hafeez, B.B.; Mandil, H.; Halaweish, F.T.; Jaggi, M.; Chauhan, S.C. Cucurbitacin D enhances the therapeutic efficacy of docetaxel via targeting cancer stem cells and miR-145. Cancer Res. 2018, 78, 2934. [Google Scholar] [CrossRef]
- Arjaibi, H.M.; Ahmed, M.S.; Halaweish, F.T. Mechanistic investigation of hepato-protective potential for cucurbitacins. Med. Chem. Res. 2017, 26, 1567–1573. [Google Scholar] [CrossRef]
- Aribi, A.; Gery, S.; Lee, D.H.; Thoennissen, N.H.; Thoennissen, G.B.; Alvarez, R.; Ho, Q.; Lee, K.; Doan, N.B.; Chan, K.T.; et al. The triterpenoid cucurbitacin B augments the antiproliferative activity of chemotherapy in human breast cancer. Int. J. Cancer 2013, 132, 2730–2737. [Google Scholar] [CrossRef] [Green Version]
- Bakar, F. Cucurbitacin B Enhances the Anticancer Effect of Imatinib Mesylate Through Inhibition of MMP-2 Expression in MCF-7 and SW480 Tumor Cell Lines. Anti-Cancer Agents Med. Chem. 2016, 8, 747–754. [Google Scholar] [CrossRef]
- Jin, Z.Q.; Hao, J.; Yang, X.; He, J.H.; Liang, J.; Yuan, J.W.; Mao, Y.; Liu, D.; Cao, R.; Wu, X.Z.; et al. Higenamine enhances the antitumor effects of cucurbitacin B in breast cancer by inhibiting the interaction of AKT and CDK2. Oncol. Rep. 2018, 40, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Jing, S.; Zou, H.; Wu, Z.; Ren, L.; Zhang, T.; Zhang, J.; Wei, Z. Cucurbitacins: Bioactivities and synergistic effect with small-molecule drugs. J. Funct. Foods 2020, 72, 104042. [Google Scholar] [CrossRef]
- Torres-Moreno, H.; Valenzuela-Chavira, I.; Marcotullio, M.C.; Vidal Gutiérrez, M.; Arrellín-Rosas, G.; Angulo-Molina, A.; Hernández Gutiérrez, S.; Robles Zepeda, R.E. In Silico Prediction Model of STAT3 Inhibition and in Vivo Antitumor Activity of Cucurbitacin IIb and Kinoin a from Ibervillea Sonorae. SSRN Electron. J. 2022, 1–42. [Google Scholar] [CrossRef]
- Omokhua-Uyi, A.G.; Van Staden, J. Phytomedicinal relevance of South African Cucurbitaceae species and their safety assessment: A review. J. Ethnopharmacol. 2020, 259, 112967. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yan, Q.; Peng, B.; Cai, Y.; Zeng, S.; Xu, Z.; Yan, Y.; Gong, Z. Use of cucurbitacins for lung cancer research and therapy. Cancer Chemother. Pharmacol. 2021, 88, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kausar, H.; Munagala, R.; Bansal, S.S.; Aqil, F.; Vadhanam, M.V.; Gupta, R.C. Cucurbitacin B potently suppresses non-small-cell lung cancer growth: Identification of intracellular thiols as critical targets. Cancer Lett. 2013, 332, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingston, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin Q: A selective STAT3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Liu, Y.; Liu, W.; Ma, F.; Zhou, Y.I.; Chen, M.; Chang, J.; Wang, Y.; Yang, G.; He, G. Cucurbitacin B inhibits growth and induces apoptosis through the JAK2/STAT3 and MAPK pathways in SH-SY5Y human neuroblastoma cells. Mol. Med. Rep. 2014, 10, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.R.; Gao, M.X.; Yang, K. Cucurbitacin B inhibits cell proliferation and induces apoptosis in human osteosarcoma cells via modulation of the JAK2/STAT3 and MAPK pathways. Exp. Ther. Med. 2017, 14, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Kaewmeesri, P.; Kukongviriyapan, V.; Prawan, A.; Kongpetch, S.; Senggunprai, L. Cucurbitacin B Diminishes Metastatic Behavior of Cholangiocarcinoma Cells by Suppressing Focal Adhesion Kinase. Asian Pac. J. Cancer Prev. 2021, 22, 219–225. [Google Scholar] [CrossRef]
- Tao, B.; Wang, D.; Yang, S.; Liu, Y.; Wu, H.; Li, Z.; Chang, L.; Yang, Z.; Liu, W. Cucurbitacin B Inhibits Cell Proliferation by Regulating X-Inactive Specific Transcript Expression in Tongue Cancer. Front. Oncol. 2021, 11, 651648. [Google Scholar] [CrossRef]
- Liu, T.; Peng, H.; Zhang, M.; Deng, Y.; Wu, Z. Cucurbitacin B, a small molecule inhibitor of the Stat3 signaling pathway, enhances the chemosensitivity of laryngeal squamous cell carcinoma cells to cisplatin. Eur. J. Pharmacol. 2010, 641, 15–22. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, M.; Zhang, H.; Sun, C.; Yang, X.; Deng, Y.; Ji, W. Combined antitumor activity of cucurbitacin B and docetaxel in laryngeal cancer. Eur. J. Pharmacol. 2008, 587, 78–84. [Google Scholar] [CrossRef]
- LUO, W.W.; ZHAO, W.W.; LU, J.J.; WANG, Y.T.; CHEN, X.P. Cucurbitacin B suppresses metastasis mediated by reactive oxygen species (ROS) via focal adhesion kinase (FAK) in breast cancer MDA-MB-231 cells. Chin. J. Nat. Med. 2018, 16, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Sha, T.; Guo, J.; Li, W.; Lu, J.; Chen, X. Cucurbitacin B induces DNA damage and autophagy mediated by reactive oxygen species (ROS) in MCF-7 breast cancer cells. J. Nat. Med. 2015, 69, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, G.; Bao, J.; Hao, W.; Lu, J.; Chen, X. Cucurbitacin B induced ATM-mediated DNA damage causes G2/M cell cycle arrest in a ROS-dependent manner. PLoS ONE 2014, 9, e88140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikander, M.; Malik, S.; Khan, S.; Kumari, S.; Chauhan, N.; Khan, P.; Halaweish, F.T.; Chauhan, B.; Yallapu, M.M.; Jaggi, M.; et al. Novel Mechanistic Insight into the Anticancer Activity of Cucurbitacin D against Pancreatic Cancer (Cuc D Attenuates Pancreatic Cancer). Cells 2019, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Fan, Q.; Liang, X.; Han, S.; He, J.; Wang, Q.Q.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits TGF-β1-induced epithelial-mesenchymal transition (EMT) in NSCLC through regulating ROS and PI3K/Akt/mTOR pathways. Chin. Med. 2022, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Baodan, Y.U.; Zheng, L.; Tang, H.; Wang, W.; Yongping, L.I.N. Cucurbitacin B enhances apoptosis in gefitinib resistant non-small cell lung cancer by modulating the miR-17-5p/STAT3 axis. Mol. Med. Rep. 2021, 240, 12349. [Google Scholar] [CrossRef]
- Liu, P.; Xiang, Y.; Liu, X.; Zhang, T.; Yang, R.; Chen, S.; Xu, L.; Yu, Q.; Zhao, H.; Zhang, L.; et al. Cucurbitacin B Induces the Lysosomal Degradation of EGFR and Suppresses the CIP2A/PP2A/Akt Signaling Axis in Gefitinib-Resistant Non-Small Cell Lung Cancer. Molecules 2019, 24, 647. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Huang, H.; Zhang, J.; Liu, H.; Ao, R.; Xiao, M.; Wu, Y. The anticancer effects of Cucurbitacin I inhibited cell growth of human non-small cell lung cancer through PI3K/AKT/p70S6K pathway. Mol. Med. Rep. 2018, 17, 2750–2756. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Wu, S.; Wang, X.; Zhu, G.; Chen, X.; Ding, Y.; Jiang, W. Cucurbitacin I induces pro-death autophagy in A549 cells via the ERK-mTOR-STAT3 signaling pathway. J. Cell. Biochem. 2018, 119, 6104–6112. [Google Scholar] [CrossRef]
- Wang, W.D.; Liu, Y.; Su, Y.; Xiong, X.Z.; Shang, D.; Xu, J.J.; Liu, H.J. Antitumor And Apoptotic Effects Of Cucurbitacin A In A-549 Lung Carcinoma Cells Is Mediated Via G2/M Cell Cycle Arrest And M-Tor/Pi3k/Akt Signalling Pathway. Afr. J. Tradit. Complement. Altern. Med. AJTCAM 2017, 14, 75–82. [Google Scholar] [CrossRef]
- Zhang, M.; Bian, Z.G.; Zhang, Y.; Wang, J.H.; Kan, L.; Wang, X.; Niu, H.Y.; He, P. Cucurbitacin B inhibits proliferation and induces apoptosis via STAT3 pathway inhibition in A549 lung cancer cells. Mol. Med. Rep. 2014, 10, 2905–2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.S.; Huang, P.I.; Chang, Y.L.; Tzao, C.; Chen, Y.W.; Shih, H.C.; Hung, S.C.; Chen, Y.C.; Tseng, L.M.; Chiou, S.H. Cucurbitacin I inhibits tumorigenic ability and enhances radiochemosensitivity in nonsmall cell lung cancer-derived CD133-positive cells. Cancer 2011, 117, 2970–2985. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Duan, C.; Ji, J.; Zhang, T.; Yuan, X.; Zhang, Y.; Ma, W.; Yang, J.; Yang, L.; Jiang, Z.; et al. Cucurbitacin B induces autophagy and apoptosis by suppressing CIP2A/PP2A/mTORC1 signaling axis in human cisplatin resistant gastric cancer cells. Oncol. Rep. 2017, 38, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.L.; Tao, W.H.; Yang, T.X.; Qiao, J.G. Anticancer effect of cucurbitacin B on MKN-45 cells via inhibition of the JAK2/STAT3 signaling pathway. Exp. Ther. Med. 2016, 12, 2709–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.; Xiang, K.; Wu, Y.; Zhang, T.; Liu, Y.; Liu, X.; Zhen, W.; Si, Y. Cucurbitacin B Inhibits the Hippo-YAP Signaling Pathway and Exerts Anticancer Activity in Colorectal Cancer Cells. Med. Sci. Monit. 2018, 24, 9251–9258. [Google Scholar] [CrossRef] [PubMed]
- Kurman, Y.; Kiliccioglu, I.; Dikmen, A.U.; Esendagli, G.; Bilen, C.Y.; Sozen, S.; Konac, E. Cucurbitacin B and cisplatin induce the cell death pathways in MB49 mouse bladder cancer model. Exp. Biol. Med. 2020, 245, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Ma, W.; Kou, W.; Li, C.; Zhao, J. Anticancer activity of cucurbitacin-A in ovarian cancer cell line SKOV3 involves cell cycle arrest, apoptosis and inhibition of mTOR/PI3K/Akt signaling pathway-PubMed. J. Buon. 2018, 23, 124–128. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Tumor | Cell Lines | Effects | Mechanisms | Concentration | References |
|---|---|---|---|---|---|
| Neuroblastoma | SH-SY5Y | Regulation of cell cycle and induces apoptosis | ↑ cell-cycle arrest at the G2/M phase, ↓ p-JAK2, ↓ p-STAT3, ↓ phospho-extracellular signal-regulated kinases, ↓ c-Jun N-terminal kinase, ↓ p38, ↓ MAPK, ↓ Cyclin B1 ↓ Bcl2- x, ↑ p53 and p21, ↓ Bcl-2 ↑ Bax | 0–128 µM | [95] |
| Osteosarcoma | U-2 OS | Cell-cycle apprehension, apoptosis and inhibition of angiogenesis | ↓ cell viability, proliferation, migration ability, ↓ MMP-2 and 9, ↑ apoptotic pathway, ↓ MAPK signaling andJAK2/STAT3 cascade, ↓ VEG F, ↑ caspase-3, -8 and -9, ↑ Bad and Bax, ↓ Bcl-2 and Bcl-xL, ↓ p38, ERK1/2, JNK and p-JNK | 20, 40, 80 and 100 µM | [96] |
| Cholangiocarcinoma | KKU-452 | Inhibits metastatic behavior | ↓ FAK activation, ↓ phospho-FAK protein, ↓ migration, invasion and adhesion abilities, ↓ MMP-9, ICAM-1 and VEGF, | 0, 5, 10, 25, 50, 100 nM | [97] |
| Tongue squamous | CAL27 and SCC9 | Induced apoptosis and microRNA mediated | ↓ proliferation, migration and invasion, ↓ Xinactive specific transcript (XIST), ↑ miR-29b | 0.001, 0.01, 0.1, 1, 10, 100 µM | [98] |
| Laryngeal squamous | Hep-2 | Regulation of cell cycle and apoptosis | compared with single treatment, combination treatment ↓ cell proliferation and viability, G2/M enrichment was accompanied by a reduction in G0/G1 phase cells, ↑ condensation of chromatin, ↑ nuclear fragmentations and apoptotic bodies, ↓ p-STAT3, ↓ Bcl-2, ↓ cyclin B1 | 1 µM cucurbitacin B + 2, 5, 10, 20, and 30 µM cisplatin | [99] |
| Hep-2 | Regulation of cell cycle, apoptosis, enhance docetaxel chemosensitivity | compared with single treatment, combination treatment ↓ cell proliferation and viability, G2/M enrichment was accompanied by a reduction in G0/G1 phase cells, ↑ condensation of chromatin, ↑ nuclear fragmentations and apoptotic bodies, significant activation (phosphorylation) of ERK1/2, ↓ p-STAT3, ↓ Bcl-2, ↓ cyclin B1 | 1 µM cucurbitacin B + 25 nM docetaxel | [100] | |
| Colon | HCT116; | Apoptosis induction | ↓ proliferation and migration ability, G2/M arrest, ↓ cyclin A, ↓ cyclin D1, ↑ p21, ↑ early apoptosis, ↓ p- Akt (Ser473) | 0.001 μM–10 μM | [14] |
| Breast | MDA-MB-231 | Suppresses metastasis | ↓ migration, invasion and adhesion ability, ↓ p-FAK (focal adhesion kinase), ↓ p- paxillin, ↑ intracellular ROS generation, | 0–100 nmol·L−1 | [101] |
| MCF-7 | Induces autophagy | ↓ cell viability, ↑ γH2AX, comet tails were significantly longer, ↑ phosphorylation of ATM (Ser-1981) and ATR (Ser428, ↑ LC3 II, ↓ p-mTOR, ↓ p-Akt (Ser308 and Ser473), ↓ p62, ↑ Beclin-1 and p-ULK1 (Ser 317), ↑ intracellular ROS | 0–200 nM | [102] | |
| MCF-7 | Regulation of cell cycle and induces apoptosis | ↑ γH2AX, comet tails were significantly longer, ↑ phosphorylation of ATM (Ser-1981) ↑ p- p53(Ser-15) | 0–800 nM | [103] | |
| Pancreatic | ASPC-1,BXPC-3, CFPAC-1, SW 1990 | Induced cell-cycle Arrest and apoptosis | ↓ proliferation, viability, ↑ percentage of cells in G2/M phase ↓ decrease in S and G0/G phase cells, ↓ cyclin B1, ↓ cyclin D1 and cyclin A2, ↓ Caspase3 and PARP1, ↓ p-JAK2 andp-STAT3 | 0, 0.25, 0.5 and 1.0 μM | [19] |
| AsPC-1, BxPC-3, CaPan-1, and HPAF-II | Cell-cycle arrest | ↓ viability of PanCa cells, ↓ colony formation capacity, ↑ G2/M Phase, ↓ invasion and migration ability, ↓ MUC13, restores miR-145 expression, ↓ proliferation of gemcitabine resistant PanCa cells, ↓ RRM1/2 expression, | 0.1, 0.25, and 0.5 µM | [104] | |
| Hepatoblastoma | HepG2 | Induced cell-cycle arrest and apoptosis | ↓ proliferation and migration ability, G2/M arrest, ↓ cyclin A, ↓ cyclin D1, ↑ p21, ↑ early apoptosis, ↓ p- Akt (Ser473), ↑ caspase-8 and PARP, | 0.001 μM–10 μM | [14] |
| Lung | A549 cells | Inhibits Metastatic Behavior, Cell-cycle arrest and apoptosis | ↓ proliferation and cell viabilitygradually, ↑ condensation, ↓ p-STAT3 levels, ↑ percentage of cells in G2/M phase, ↑ levels of ROS, ↑ membrane pore formation, ↑ pyroptosis, ↑ caspase-3, 9 activity | Cucumber-derived nanovesicles (CDNVs)- CDNVs containing 10 nM CuB | [15] |
| A549 | Apoptosis and microRNA mediated | ↓ proliferation of lung cancer cells, ↑ cell apoptosis frequency, ↑ Bax and cleaved caspase3, ↓ cyclin B1 and Bcl-2, ↓ XIST and IL-6, ↑ miR-let-7c expression, ↓ IL-6/STAT3 pathway | 0.1, 0.3, 0.6, and 0.9 μM | [52] | |
| A549, A549-GR | Anti-metastasis | ↓ EMT, ↓ p-PI3K, ↓ p-Akt, ↓ p-mTOR | 5, 10, 15, 20 nM | [105] | |
| PC9 (gefitinib resistance) | Apoptosis and microRNA mediated | ↑ miR-17-5p in PC9/GR cells, caspases, STAT3, ↓ p- STAT3 | -- | [106] | |
| A549 | Induced cell-cycle arrest and apoptosis | ↓ proliferation and migration ability, G2/M arrest, ↓ cyclin A,↓ cyclin D1, ↑ p21, ↑ early apoptosis, ↓ p- Akt (Ser473) | 0.001 μM–10 μM | [14] | |
| A549, NCI-H1299 (H1299), NCI-H1975 (H1975), and NCI-H820 (H820) (gefitinib resistance) | Induces apoptosis | ↓ anchorage-dependent growth and clonogenic ability, ↓ proliferation, invasion and migration ability, ↑ caspase-8 and 3, caspase-3, c-PARP, ↑ Lysosomal Degradation of EGFR and thus Inhibits ERK Signaling, ↓ CIP2A expression, ↑ PP2A, ↓ pAkt | IC50 (µM) H1299-0.77 A549-0.76 H197-0.63 H820-0.19 | [107] | |
| A549 | Induces apoptosis | ↓ cell proliferation, ↑ capase-3/9, ↓ PI3K, p-AKT and p-p70S6K | 0, 50, 100 and 200 nM | [108] | |
| A549 | Induces autophagy | ↓ cell viability, colony formation ability, ↑apoptosis frequency, ↑ apoptotic and necrotic, ↑ autophagosomes, ↑ LC3-II/LC3-I, ↓ p-mTOR (S2448), ↓ p- ERK and p- STAT3 | 0, 100, 200, 300, 400 and 500 nM | [109] | |
| A-549 | Induces cell-cycle arrest and apoptosis | ↑ G2/M phase cell-cycle collapse, ↓ m-TOR/PI3K/Akt proteins | 0, 10, 20, 40, 100, 150 and 200 μM | [110] | |
| A549 | Regulation of cell cycle and induces apoptosis | ↓ proliferation and colony forming ability, ↑ γH2AX, comet tails were significantly longer, ↑ G2/M phase, ↑ phosphorylation of ATM (Ser-1981), ↑ p- p53(Ser-15) | 0–800 nM | [103] | |
| A549 | Regulation of cell cycle and induces apoptosis | ↓ cell proliferation, ↑ G2/M phasecells, % early and lateapoptotic cells, ↑ cell shrinking, ↑ intracytoplasmic vacuoles, ↑chromatin condensation, ↑mitochondrial swelling, ↑caspase-3 and caspase-9, ↑ disruption of the ΔΨm, ↓ (p)-STAT3, ↓ cyclinB1 ↓ Bcl-2 | 0.02,0.1, 0.5, 2.5, 12.5 and 62.5 µmol/L | [111] | |
| CD133-positive and CD133-negative | Inhibits tumorigenic ability and enhances radiochemo-sensitivity | ↓ STAT3, ↓ tumorigenic capacity,↓ sphere formation ability, ↓ radioresistance and chemoresistance in CD133-positive, ↓ stemness gene signature of CD133-positive, ↓ Bcl-2, ↓ Bcl-xL ↓survivin, ↑ Bax | radiation doses (0 Gy, 2Gy, 4 Gy, 6 Gy, 8 Gy, and 10 Gy) + 0, 50, 100 and 150 nM | [112] | |
| Gastric | Human DDP-resistant gastric cancer cell lineSGC7901/DDP and human GC cell line SGC7901 | Induces autophagy and apoptosis | ↓ viability, ↓ clonogenic ability, ↑ cytoplasmic shrinkage, ↓ pro-caspases-3 and -9 and cleaved PARP, ↑ LC3 II and Beclin1, ↓ P-gp, and HIF-1α, ↓ phosphorylation of mTORC1 effectors (mTOR, p70S6K and 4E-BP1), ↓ pAkt, ↑PP2A, ↓ CIP2A | IC50 (nM) SGC7901- 216.70 SGC7901/DDP- 170.25 | [113] |
| MKN-45 | Cell-cycle arrest, apoptosis | ↓ proliferation of cancer cells, ↑ progression of the cell cycle from G0/G1 to S phase, ↓ cyclin D1, ↓ cyclin E, ↓ CDK4 and CDK2 (cyclin-dependent kinase), ↑ p27, ↑ cell apoptosis frequency, ↑Bax, ↓ Bcl-2, ↓ JAK2/STAT3 signaling pathway | 0.1, 1 or 10 µM | [114] | |
| Colorectal | SW620 and HT29 | Regulation of cell cycle and apoptosis | ↑ proliferation and invasion, ↑ chromatin condensation and fragmentation, ↓ pro-cas-3, ↓cleaved PARP, ↓ YAP ↓Cyr 61 and c-Myc, ↑ LATS1 | 0, 0.1, 0.2, 0.4, 0.6, 0.8 and 1.0 μM | [115] |
| Prostate | LNCaP, DU145, and PC-3; | Induced cell-cycle arrest and apoptosis | ↓ proliferation and migration ability, G2/M arrest, ↓ cyclin A,↓ cyclin D1, ↑ p21, ↑ early apoptosis, ↓ p- Akt (Ser473), ↑ caspase-8 and PARP | 0.001 μM–10 μM | [15] |
| Bladder | MB49 | Induce the cell death pathways, apoptosis and autophagy | ↓ viability of cancer cells, ↓Bcl-2, ↑ LC3II, ↓ phosphorylation of p27, PRAS40 and Raf-1 proteins, ↑ p- AKT, ↓ p-ERK1/ ERK2, ↓ p-mTOR, ↑ BAD, ↑AMPKα | CuB- 0.01–50 μM and Cisplatin- 0.5–50 μM | [116] |
| T24 | Induced cell-cycle arrest and apoptosis | ↓ proliferation and migration ability, G2/M arrest, ↓ cyclin A,↓ cyclin D1, ↑ p21, ↑ early apoptosis, ↓ p- Akt (Ser473), ↑ caspase-8 and PARP | 0.001 μM–10 μM | [14] | |
| Ovarian | SKOV3 | Cell-cycle arrest, apoptosis | ↑ chromatin condensation, ↑ apoptotic body formation, ↑ deformed cell morphology, ↑ intracellular ROS levels, ↓ MMP, ↓ mTOR/PI3K/Akt signaling pathway, ↓ m-TOR, ↓ phospho m-TOR proteins. ↓ PI3K/Akt protein expressions, ↑ DNA damage, ↑ cell-cycle arrest at G2/M checkpoint | 0, 10, 20, 40, 80 and 160 μM | [117] |
| Type of Tumor | Animal Models | Effects | Mechanisms | Dosage | Duration | References |
|---|---|---|---|---|---|---|
| Laryngeal | Nude mice injected with 5 × 106 Hep-2 cells | Inhibits tumor growth | ↓ tumor weight and volume, | 55 μg/kg/day ofcucurbitacin B, or 7.5 mg/kg/week of docetaxel | 14 days | [100] |
| Athymic nude mice 5 × 106 Hep-2 cells | Inhibits tumor growth | ↓ tumor growth inhibition, no significant adverse effects were observed | 55 µg/kg daily for 14 days, orcisplatin (intraperitoneal injection) 10 mg/kg daily | 14 days | [99] | |
| Tongue squamous | Nude mice injected with SCC9 cell lines (8 × 106) | Inhibits tumor growth | ↓ XIST, XIST expression was lost in XIST KO (knockout) | 0.5 mg/kg | 14 days | [98] |
| Lung | C57BL/6 J mice injected with 5 × 105 B16-F10-Luc cells | Anti-metastasis | ↓ lung index, ↓ lung metastasis | CuB (0.25 mg/kg, and 0.5 mg/kg) groups and Geftinib (40 mg/kg) | 14 days | [105] |
| BALB/c nude mice injected with A549 cells (2×106 cells) | Inhibits tumor growth | ↓tumor weight, ↑necrotic and apoptotic cells were observed in tumor sections, ↓ p-STAT3, ↓ CD31, | CsDNVscontaining 472 nM CuB | 14 days | [15] | |
| nu/nu mice injected with GR NSCLC H1975 cells (2.5 × 106) | Inhibits tumor growth | ↓ tumor weight, ↓ CIP2Aand EGFR | gefitinib (30 mg/kg) or CucB 0.5 mg/kg | 24 days | [107] | |
| BALB/c mice injected with 1 × 105 CD133-positive and CD133-negative cells | Inhibited tumor growth | ↓ lung metastasis, ↓ tumor size, ↓ tumorigenic and metastatic capabilities, ↑ survival rate | 1 mg/kg | 4 weeks | [112] | |
| Pancreatic | BALB/c-nu injected with BXPC-3cells (2 × 106 cells) | Inhibits tumor growth | ↓ p-STAT3 in the tumors of mice, ↑ PCNA in control | 1 mg/kg and 2 mg/kg | 30 days | [19] |
| NOD-SCID gamma mice injected with HPAF-II cells (4 × 106 cells) | Inhibits tumor growth | ↓ MUC13, ↓ PCNA (nuclear proliferating cell antigen), ↑ miRNA-145 | 1 mg/kg | 40 days | [104] | |
| Hepatocellular | SCID mice injected with HepG2 (3 × 106) | Inhibits tumor growth | No significant differences of body weight, ↑ DNA cleavage, ↑ CDKN1A (p21), ↑ CDKN1B (p27), ↑ FOXO, ↑ p- Akt | 0.1 mg/kg | 3 weeks | [14] |
| Prostrate | SCID mice injected with HepG2 (3 × 106) cells | Inhibits tumor growth | No significant differences of body weight, ↑ DNA cleavage, ↑ CDKN1A (p21), ↑ CDKN1B (p27), ↑ FOXO, ↓ p- Akt | 0.1 mg/kg | 8 weeks | [14] |
| Bladder | C57BL/6 injected with 1 × 106 MB49 cells | Reduced the tumor growth | No histopathological changes, ↓ Bcl-2, ↑ LC3II, ↓ phosphorylation of p27, PRAS40 and Raf-1 proteins, ↑ p- AKT, ↓ p-ERK1/ ERK2, ↓ p-mTOR, | CuB (0.5–1 mg/kg) and Cis (1–3 mg/kg) | 19 days | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuli, H.S.; Rath, P.; Chauhan, A.; Ranjan, A.; Ramniwas, S.; Sak, K.; Aggarwal, D.; Kumar, M.; Dhama, K.; Lee, E.H.C.; et al. Cucurbitacins as Potent Chemo-Preventive Agents: Mechanistic Insight and Recent Trends. Biomolecules 2023, 13, 57. https://doi.org/10.3390/biom13010057
Tuli HS, Rath P, Chauhan A, Ranjan A, Ramniwas S, Sak K, Aggarwal D, Kumar M, Dhama K, Lee EHC, et al. Cucurbitacins as Potent Chemo-Preventive Agents: Mechanistic Insight and Recent Trends. Biomolecules. 2023; 13(1):57. https://doi.org/10.3390/biom13010057
Chicago/Turabian StyleTuli, Hardeep Singh, Prangya Rath, Abhishek Chauhan, Anuj Ranjan, Seema Ramniwas, Katrin Sak, Diwakar Aggarwal, Manoj Kumar, Kuldeep Dhama, E Hui Clarissa Lee, and et al. 2023. "Cucurbitacins as Potent Chemo-Preventive Agents: Mechanistic Insight and Recent Trends" Biomolecules 13, no. 1: 57. https://doi.org/10.3390/biom13010057