
Application of Single-Cell RNA Sequencing in Ovarian Development

Abstract
:1. Introduction
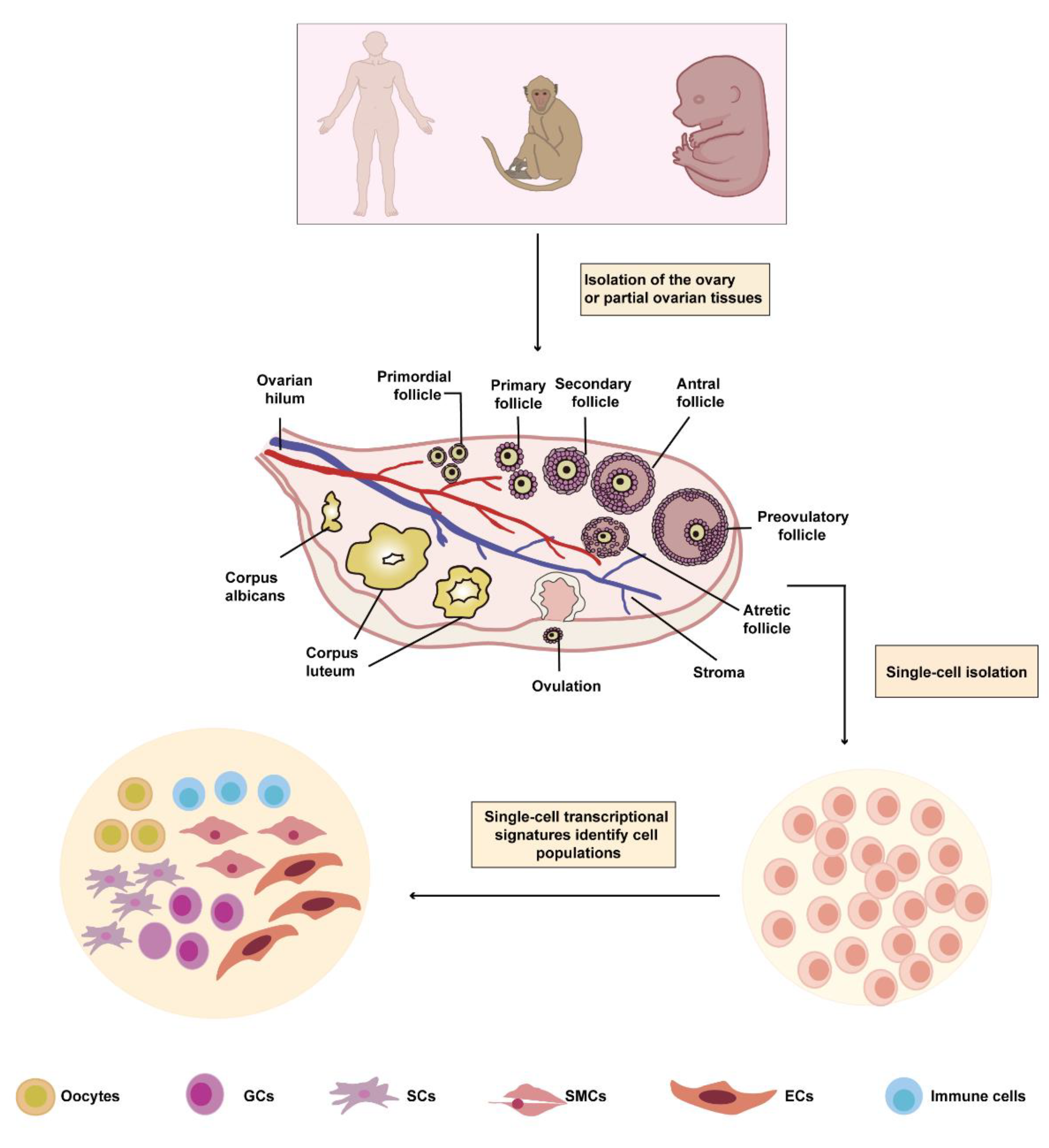
2. Ovarian Cell Heterogeneity at A Single-Cell Resolution
2.1. The Single-Cell Atlas of the Ovary
2.2. The Cell-Type-Specific Genes of Ovarian Cells
3. A Single-Cell RNA Perspective of Ovarian Germ Cells
3.1. A Single-Cell Perspective of Fetal Germ Cells
3.1.1. The Cell Subpopulations and Characteristics of Fetal Germ Cells
3.1.2. Molecular Mechanisms of the Mitotic-to-Meiotic Transition and the Initiation of Meiosis in Fetal Germ Cells
3.2. A Single-Cell Perspective of Oocytes
3.2.1. Oocyte Characteristics from Different Follicular Stages
3.2.2. The Characteristics of In Vitro Matured Oocytes
3.2.3. The Characteristics of Oocytes from Aged Females
3.2.4. The Characteristics of Oocytes from Polycystic Ovarian Syndrome Patients
3.3. A Single-Cell Perspective of Oogonial Stem Cells
4. A Single-Cell RNA Perspective of Ovarian Somatic Cells
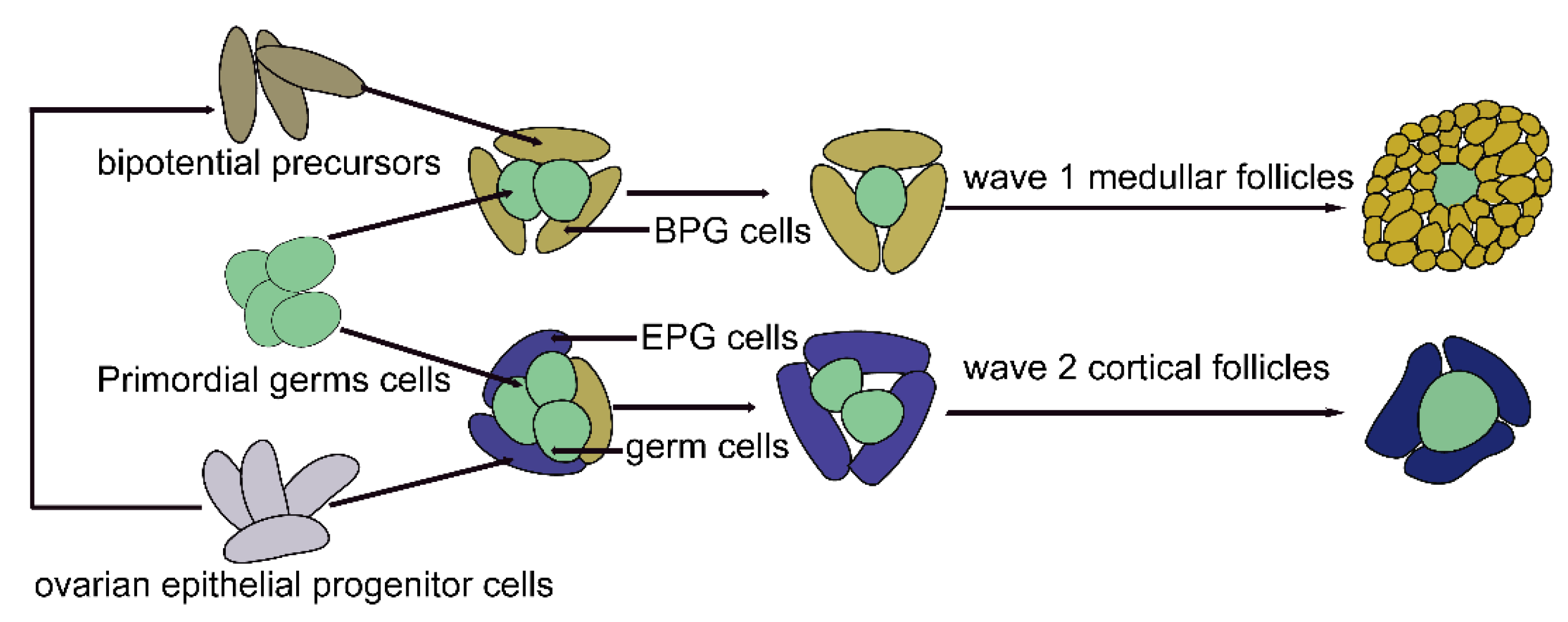
4.1. A Single-Cell Perspective of Pregranulosa Cells
4.2. A Single-Cell Perspective of Granulosa Cells
4.2.1. The Characteristics of Granulosa Cells from Different Stages of Follicles
4.2.2. The Characteristics of Granulosa Cells during Ovulation
4.2.3. The Characteristics of Granulosa Cells in Atretic Follicles
4.2.4. The Characteristics of Granulosa Cells from Aged Females
4.3. A Single-Cell Perspective of Ovarian Stromal Cells
4.4. A Single-Cell Perspective of Ovarian Smooth Muscle Cells
4.5. A Single-Cell Perspective of Ovarian Endothelial Cells
4.6. A Single-Cell Perspective of Ovarian Immune Cells
5. Applications of Single-Cell Transcriptome Sequencing in Ovarian Intercellular Interactions
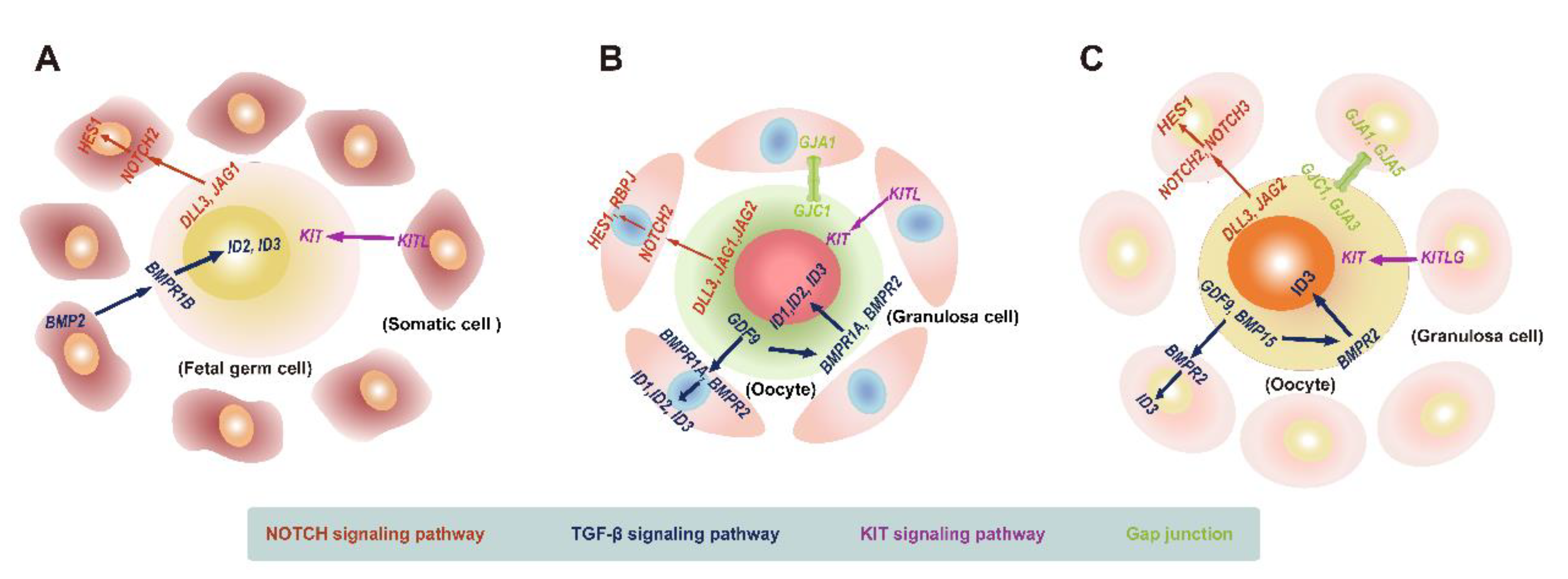
5.1. Landscape of Interactions between Female Human Embryonic Germ Cells and Gonadal Somatic Cells
5.2. Landscape of Crosstalk between Oocytes and Granulosa Cells during Primordial Follicular Assembly
5.3. Landscape of Interactions between Oocytes and Granulosa Cells during Folliculogenesis
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oktem, O.; Oktay, K. The ovary: Anatomy and function throughout human life. Ann. N. Y. Acad. Sci. 2008, 1127, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Muraoka, M.; Kato, Y.; Saga, Y. Decoding the transcriptome of pre-granulosa cells during the formation of primordial follicles in the mouse†. Biol. Reprod. 2021, 105, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Cowan, R.G.; Quirk, S.M. Cells responding to hedgehog signaling contribute to the theca of ovarian follicles. Reproduction 2021, 161, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Li, S.Y.; DeFalco, T. Immune and vascular contributions to organogenesis of the testis and ovary. FEBS J. 2022, 289, 2386–2408. [Google Scholar] [CrossRef]
- He, J.; Yao, G.; He, Q.; Zhang, T.; Fan, H.; Bai, Y.; Zhang, J.; Yang, G.; Xu, Z.; Hu, J.; et al. Theaflavin 3, 3′-Digallate Delays Ovarian Aging by Improving Oocyte Quality and Regulating Granulosa Cell Function. Oxidative Med. Cell. Longev. 2021, 2021, 7064179. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, B.; Mosser, D.M.; Zhang, X. TLRs, macrophages, and NK cells: Our understandings of their functions in uterus and ovary. Int. Immunopharmacol. 2011, 11, 1442–1450. [Google Scholar] [CrossRef]
- Fabbri, R.; Vicenti, R.; Macciocca, M.; Pasquinelli, G.; Paradisi, R.; Battaglia, C.; Martino, N.A.; Venturoli, S. Good preservation of stromal cells and no apoptosis in human ovarian tissue after vitrification. BioMed Res. Int. 2014, 2014, 673537. [Google Scholar] [CrossRef]
- Choi, D.H.; Kim, E.K.; Kim, K.H.; Lee, K.A.; Kang, D.W.; Kim, H.Y.; Bridges, P.; Ko, C. Expression pattern of endothelin system components and localization of smooth muscle cells in the human pre-ovulatory follicle. Hum. Reprod. 2011, 26, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Kedem, A.; Aelion-Brauer, A.; Guo, P.; Wen, D.; Ding, B.S.; Lis, R.; Cheng, D.; Sandler, V.M.; Rafii, S.; Rosenwaks, Z. Activated ovarian endothelial cells promote early follicular development and survival. J. Ovarian Res. 2017, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, L.A.; Egbert, J.R. Regulation of Mammalian Oocyte Meiosis by Intercellular Communication Within the Ovarian Follicle. Ann. Rev. Physiol. 2017, 79, 237–260. [Google Scholar] [CrossRef]
- Herrera, L.; Ottolenghi, C.; Garcia-Ortiz, J.E.; Pellegrini, M.; Manini, F.; Ko, M.S.; Nagaraja, R.; Forabosco, A.; Schlessinger, D. Mouse ovary developmental RNA and protein markers from gene expression profiling. Dev. Biol. 2005, 279, 271–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artini, P.G.; Tatone, C.; Sperduti, S.; D’Aurora, M.; Franchi, S.; Di Emidio, G.; Ciriminna, R.; Vento, M.; Di Pietro, C.; Stuppia, L.; et al. Cumulus cells surrounding oocytes with high developmental competence exhibit down-regulation of phosphoinositol 1,3 kinase/protein kinase B (PI3K/AKT) signalling genes involved in proliferation and survival. Hum. Reprod. 2017, 32, 2474–2484. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Gong, W.; Zhou, Y.; Li, X.; Yu, J.; Hu, S. A comprehensive transcriptomic analysis of infant and adult mouse ovary. Genom. Proteom. Bioinform. 2014, 12, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, G.; Zhang, N.; Hua, K. A time-resolved transcriptome landscape of the developing mouse ovary. Biochem. Biophys. Res. Commun. 2021, 572, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, S.E.; Collado-Torres, L.; Jaffe, A.; Leek, J.T. Improving the value of public RNA-seq expression data by phenotype prediction. Nucleic Acids Res. 2018, 46, e542018. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Liu, Y.; Mao, X.; Jia, C.; Ferguson, J.F.; Xue, C.; Reilly, M.P.; Li, H.; Li, M. PennSeq: Accurate isoform-specific gene expression quantification in RNA-Seq by modeling non-uniform read distribution. Nucleic Acids Res. 2014, 42, e202014. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, C.Y. From bulk, single-cell to spatial RNA sequencing. Int. J. Oral Sci. 2021, 13, 36. [Google Scholar] [CrossRef]
- Olsen, T.K.; Baryawno, N. Introduction to Single-Cell RNA Sequencing. Curr. Protoc. Mol. Biol. 2018, 122, e572018. [Google Scholar] [CrossRef]
- Noé, A.; Cargill, T.N.; Nielsen, C.M.; Russell, A.J.C.; Barnes, E. The Application of Single-Cell RNA Sequencing in Vaccinology. J. Immunol. Res. 2020, 2020, 8624963. [Google Scholar] [CrossRef]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A.; et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Anderson, A.G.; Merullo, D.P.; Konopka, G. Beyond bulk: A review of single cell transcriptomics methodologies and applications. Curr. Opin. Biotechnol. 2019, 58, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.S. Single-cell RNA sequencing for the study of development, physiology and disease. Nat. Rev. Nephrol. 2018, 14, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ye, F.; Guo, G. Revolutionizing immunology with single-cell RNA sequencing. Cell Mol. Immunol. 2019, 16, 242–249. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.; Lee, J.H.; Bang, D. Single-cell RNA sequencing technologies and bioinformatics pipelines. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [Green Version]
- MacParland, S.A.; Liu, J.C.; Ma, X.Z.; Innes, B.T.; Bartczak, A.M.; Gage, B.K.; Manuel, J.; Khuu, N.; Echeverri, J.; Linares, I.; et al. Single cell RNA sequencing of human liver reveals distinct intrahepatic macrophage populations. Nat. Commun. 2018, 9, 4383. [Google Scholar] [CrossRef] [Green Version]
- Travaglini, K.J.; Nabhan, A.N.; Penland, L.; Sinha, R.; Gillich, A.; Sit, R.V.; Chang, S.; Conley, S.D.; Mori, Y.; Seita, J.; et al. A molecular cell atlas of the human lung from single-cell RNA sequencing. Nature 2020, 587, 619–625. [Google Scholar] [CrossRef]
- Astapova, O.; Minor, B.M.N.; Hammes, S.R. Physiological and Pathological Androgen Actions in the Ovary. Endocrinology 2019, 160, 1166–1174. [Google Scholar] [CrossRef]
- Richani, D.; Dunning, K.R.; Thompson, J.G.; Gilchrist, R.B. Metabolic co-dependence of the oocyte and cumulus cells: Essential role in determining oocyte developmental competence. Hum. Reprod. Update 2021, 27, 27–47. [Google Scholar] [CrossRef]
- Sun, N.; Yu, X.; Li, F.; Liu, D.; Suo, S.; Chen, W.; Chen, S.; Song, L.; Green, C.D.; McDermott, J.; et al. Inference of differentiation time for single cell transcriptomes using cell population reference data. Nat. Commun. 2017, 8, 1856. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, J.; Wu, Z.; Sun, X. Inferring microenvironmental regulation of gene expression from single-cell RNA sequencing data using scMLnet with an application to COVID-19. Brief. Bioinform. 2021, 22, 988–1005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X. Interaction and signal transduction between oocyte and samatic cells in the ovary. Front. Biosci. A J. Virtual Libr. 2007, 12, 2782–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Gardeux, V.; Meireles-Filho, A.; Deplancke, B. Profiling of Single-Cell Transcriptomes. Curr. Protoc. Mouse Biol. 2017, 7, 145–175. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Bialecka, M.; Moustakas, I.; Lam, E.; Torrens-Juaneda, V.; Borggreven, N.V.; Trouw, L.; Louwe, L.A.; Pilgram, G.S.K.; Mei, H.; et al. Single-cell reconstruction of follicular remodeling in the human adult ovary. Nat. Commun. 2019, 10, 3164. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Ge, W.; Zhai, Q.Y.; Liu, J.C.; Sun, X.W.; Liu, W.X.; Li, L.; Lei, C.Z.; Dyce, P.W.; De Felici, M.; et al. Single-cell transcriptome landscape of ovarian cells during primordial follicle assembly in mice. PLoS Biol. 2020, 18, e30010252020. [Google Scholar] [CrossRef]
- Wang, J.J.; Tian, Y.; Li, M.H.; Feng, Y.Q.; Kong, L.; Zhang, F.L.; Shen, W. Single-cell transcriptome dissection of the toxic impact of Di (2-ethylhexyl) phthalate on primordial follicle assembly. Theranostics 2021, 11, 4992–5009. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, M.Y.; Zhao, A.H.; Kong, L.; Wang, J.J.; Shen, W.; Li, L. Single-cell transcriptomic profiling provides insights into the toxic effects of Zearalenone exposure on primordial follicle assembly. Theranostics 2021, 11, 5197–5213. [Google Scholar] [CrossRef]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef] [Green Version]
- Paranko, J.; Pelliniemi, L.J. Differentiation of smooth muscle cells in the fetal rat testis and ovary: Localization of alkaline phosphatase, smooth muscle myosin, F-actin, and desmin. Cell Tissue Res. 1992, 268, 521–530. [Google Scholar] [CrossRef]
- Han, Y.J.; Hu, W.Y.; Chernaya, O.; Antic, N.; Gu, L.; Gupta, M.; Piano, M.; de Lanerolle, P. Increased myosin light chain kinase expression in hypertension: Regulation by serum response factor via an insertion mutation in the promoter. Mol. Biol. Cell 2006, 17, 4039–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haaksma, C.J.; Schwartz, R.J.; Tomasek, J.J. Myoepithelial cell contraction and milk ejection are impaired in mammary glands of mice lacking smooth muscle alpha-actin. Biol. Reprod. 2011, 85, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migone, F.F.; Cowan, R.G.; Williams, R.M.; Gorse, K.J.; Zipfel, W.R.; Quirk, S.M. In vivo imaging reveals an essential role of vasoconstriction in rupture of the ovarian follicle at ovulation. Proc. Natl. Acad. Sci. USA 2016, 113, 2294–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doss, D.N.; Boulos, I.K.; Fredens, K. The presence of smooth muscle cells in the rat ovary. Histochemistry 1984, 81, 205–206. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600.e19. [Google Scholar] [CrossRef]
- Ziegenhain, C.; Vieth, B.; Parekh, S.; Reinius, B.; Guillaumet-Adkins, A.; Smets, M.; Leonhardt, H.; Heyn, H.; Hellmann, I.; Enard, W. Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol. Cell 2017, 65, 631–643.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.H.; Ma, J.Y.; Meng, T.G.; Wang, Z.B.; Yue, W.; Zhou, Q.; Li, S.; Feng, X.; Hou, Y.; Schatten, H.; et al. Single-cell RNA sequencing reveals the landscape of early female germ cell development. FASEB J. 2020, 34, 12634–12645. [Google Scholar] [CrossRef]
- Niu, W.; Spradling, A.C. Two distinct pathways of pregranulosa cell differentiation support follicle formation in the mouse ovary. Proc. Natl. Acad. Sci. USA 2020, 117, 20015–20026. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Moustakas, I.; Bialecka, M.; Del Valle, J.S.; Overeem, A.W.; Louwe, L.A.; Pilgram, G.S.K.; van der Westerlaken, L.A.J.; Mei, H.; Chuva de Sousa Lopes, S.M. Single-Cell Transcriptomics Analysis of Human Small Antral Follicles. Int. J. Mol. Sci. 2021, 22, 11955. [Google Scholar] [CrossRef]
- Li, S.; Chen, L.N.; Zhu, H.J.; Feng, X.; Xie, F.Y.; Luo, S.M.; Ou, X.H.; Ma, J.Y. Single-cell RNA sequencing analysis of mouse follicular somatic cells†. Biol. Reprod. 2021, 105, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liu, C.; Liu, Y. Advances in research on the development of female germ cells. Zhonghua Yi Xue Yi Chuan Xue Za Zhi = Zhonghua Yixue Yichuanxue Zazhi = Chin. J. Med. Genet. 2021, 38, 286–289. [Google Scholar]
- Guigon, C.J.; Magre, S. Contribution of germ cells to the differentiation and maturation of the ovary: Insights from models of germ cell depletion. Biol. Reprod. 2006, 74, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, A. Development of the mammalian gonad: The fate of the supporting cell lineage. BioEssays News Rev. Mol. Cell. Dev. Biol. 1991, 13, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Monget, P.; McNatty, K.; Monniaux, D. The Crazy Ovary. Genes 2021, 12, 928. [Google Scholar] [CrossRef] [PubMed]
- Broekmans, F.J.; Soules, M.R.; Fauser, B.C. Ovarian aging: Mechanisms and clinical consequences. Endocr. Rev. 2009, 30, 465–493. [Google Scholar] [CrossRef] [Green Version]
- Ben-Meir, A.; Burstein, E.; Borrego-Alvarez, A.; Chong, J.; Wong, E.; Yavorska, T.; Naranian, T.; Chi, M.; Wang, Y.; Bentov, Y.; et al. Coenzyme Q10 restores oocyte mitochondrial function and fertility during reproductive aging. Aging Cell 2015, 14, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Cafforio, P.; Felici, C.; Cormio, G.; D’Oronzo, S. Ddx4(+) Oogonial Stem Cells in Postmenopausal Women’s Ovaries: A Controversial, Undefined Role. Cells 2019, 8, 650. [Google Scholar] [CrossRef] [Green Version]
- Sequeira, R.C.; Sittadjody, S.; Criswell, T.; Atala, A.; Jackson, J.D.; Yoo, J.J. Enhanced method to select human oogonial stem cells for fertility research. Cell Tissue Res. 2021, 386, 145–156. [Google Scholar] [CrossRef]
- Gondos, B.; Bhiraleus, P.; Hobel, C.J. Ultrastructural observations on germ cells in human fetal ovaries. Am. J. Obstet. Gynecol. 1971, 110, 644–652. [Google Scholar] [CrossRef]
- Mamsen, L.S.; Brøchner, C.B.; Byskov, A.G.; Møllgard, K. The migration and loss of human primordial germ stem cells from the hind gut epithelium towards the gonadal ridge. Int. J. Dev. Biol. 2012, 56, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Wear, H.M.; McPike, M.J.; Watanabe, K.H. From primordial germ cells to primordial follicles: A review and visual representation of early ovarian development in mice. J. Ovarian Res. 2016, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereda, J.; Zorn, T.; Soto-Suazo, M. Migration of human and mouse primordial germ cells and colonization of the developing ovary: An ultrastructural and cytochemical study. Microsc. Res. Tech. 2006, 69, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Spiller, C.M.; Bowles, J.; Koopman, P. Regulation of germ cell meiosis in the fetal ovary. Int. J. Dev. Biol. 2012, 56, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, C.M.; Bowles, J. Sex determination in mammalian germ cells. Asian J. Androl. 2015, 17, 427–432. [Google Scholar]
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452. [Google Scholar] [CrossRef] [Green Version]
- Morita-Fujimura, Y.; Tokitake, Y.; Matsui, Y. Heterogeneity of mouse primordial germ cells reflecting the distinct status of their differentiation, proliferation and apoptosis can be classified by the expression of cell surface proteins integrin α6 and c-Kit. Dev. Growth Differ. 2009, 51, 567–583. [Google Scholar] [CrossRef]
- Zhong, C.; Liu, M.; Tao, Y.; Wu, X.; Yang, Y.; Wang, T.; Meng, Z.; Xu, H.; Liu, X. Pou5f1 and Nanog Are Reliable Germ Cell-Specific Genes in Gonad of a Protogynous Hermaphroditic Fish, Orange-Spotted Grouper (Epinephelus coioides). Genes 2021, 13, 79. [Google Scholar] [CrossRef]
- de Jong, J.; Stoop, H.; Gillis, A.J.; van Gurp, R.J.; van de Geijn, G.J.; Boer, M.; Hersmus, R.; Saunders, P.T.; Anderson, R.A.; Oosterhuis, J.W.; et al. Differential expression of SOX17 and SOX2 in germ cells and stem cells has biological and clinical implications. J. Pathol. 2008, 215, 21–30. [Google Scholar] [CrossRef]
- Lei, L.; Spradling, A.C. Mouse primordial germ cells produce cysts that partially fragment prior to meiosis. Development 2013, 140, 2075–2081. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.X.; Zhang, Y.; Li, Y.Y.; Liu, X.M.; Wang, X.X.; Zhang, C.L.; Hao, C.F.; Deng, S.L. Regulation of follicular development and differentiation by intra-ovarian factors and endocrine hormones. Front. Biosci. 2019, 24, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Chen, C.; De Felici, M.; Shen, W. In vitro differentiation of germ cells from stem cells: A comparison between primordial germ cells and in vitro derived primordial germ cell-like cells. Cell Death Dis. 2015, 6, e19062015. [Google Scholar] [CrossRef] [PubMed]
- Grive, K.J.; Freiman, R.N. The developmental origins of the mammalian ovarian reserve. Development 2015, 142, 2554–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, W.; Wang, J.J.; Zhang, R.Q.; Tan, S.J.; Zhang, F.L.; Liu, W.X.; Li, L.; Sun, X.F.; Cheng, S.F.; Dyce, P.W.; et al. Dissecting the initiation of female meiosis in the mouse at single-cell resolution. Cell. Mol. Life Sci. CMLS 2021, 78, 695–713. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Osada, A.; Ohta, M.; Yokota, K.; Nishiyama, A.; Niikura, Y.; Tamura, T.; Sekita, Y.; Kimura, T. SWI/SNF chromatin remodeling complex is required for initiation of sex-dependent differentiation in mouse germline. Sci. Rep. 2021, 11, 24074. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, Y.; Qin, Y.; Xu, B.; Guo, T.; Ke, H.; Chen, M.; Zhang, L.; Han, F.; Li, Y.; et al. Wdr62 is involved in female meiotic initiation via activating JNK signaling and associated with POI in humans. PLoS Genet. 2018, 14, e1008504. [Google Scholar] [CrossRef] [Green Version]
- Eppig, J.J. Growth and development of mammalian oocytes in vitro. Arch. Pathol. Lab. Med. 1992, 116, 379–382. [Google Scholar] [CrossRef]
- Pelland, A.M.; Corbett, H.E.; Baltz, J.M. Amino Acid transport mechanisms in mouse oocytes during growth and meiotic maturation. Biol. Reprod. 2009, 81, 1041–1054. [Google Scholar] [CrossRef] [Green Version]
- Ferrero, H.; Corachán, A.; Aguilar, A.; Quiñonero, A.; Carbajo-García, M.C.; Alamá, P.; Tejera, A.; Taboas, E.; Muñoz, E.; Pellicer, A.; et al. Single-cell RNA sequencing of oocytes from ovarian endometriosis patients reveals a differential transcriptomic profile associated with lower quality. Hum. Reprod. 2019, 34, 1302–1312. [Google Scholar] [CrossRef] [Green Version]
- Sieber, M.H.; Thomsen, M.B.; Spradling, A.C. Electron Transport Chain Remodeling by GSK3 during Oogenesis Connects Nutrient State to Reproduction. Cell 2016, 164, 420–432. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J. Cell. Physiol. 2021, 236, 7966–7983. [Google Scholar] [CrossRef] [PubMed]
- Kindsfather, A.J.; Czekalski, M.A.; Pressimone, C.A.; Erisman, M.P.; Mann, M.R.W. Perturbations in imprinted methylation from assisted reproductive technologies but not advanced maternal age in mouse preimplantation embryos. Clin. Epigenetics 2019, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, J.; Zhang, Y.; Zhang, Y.; Jia, L.; Zhang, D.; Zhang, J.; Han, Y.; Luo, S. The Efficacy of Complementary and Alternative Medicine in the Treatment of Female Infertility. Evid.-Based Complement. Altern. Med. Ecam 2021, 2021, 6634309. [Google Scholar] [CrossRef] [PubMed]
- Carson, S.A.; Kallen, A.N. Diagnosis and Management of Infertility: A Review. JAMA 2021, 326, 65–76. [Google Scholar] [CrossRef]
- Luke, B. Pregnancy and birth outcomes in couples with infertility with and without assisted reproductive technology: With an emphasis on US population-based studies. Am. J. Obstet. Gynecol. 2017, 217, 270–281. [Google Scholar] [CrossRef]
- Hendriks, W.K.; Colleoni, S.; Galli, C.; Paris, D.B.; Colenbrander, B.; Roelen, B.A.; Stout, T.A. Maternal age and in vitro culture affect mitochondrial number and function in equine oocytes and embryos. Reprod. Fertil. Dev. 2015, 27, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.W.T.; Ng, J.K.W.; Liao, J.; Luk, A.C.; Suen, A.H.C.; Chan, T.T.H.; Cheung, M.Y.; Chu, H.T.; Tang, N.L.S.; Zhao, M.P.; et al. Single-cell RNA sequencing identifies molecular targets associated with poor in vitro maturation performance of oocytes collected from ovarian stimulation. Hum. Reprod. 2021, 36, 1907–1921. [Google Scholar] [CrossRef]
- Rodriguez-Wallberg, K.A.; Oktay, K. Recent advances in oocyte and ovarian tissue cryopreservation and transplantation. Best Pract. Res. Clin. Obstet. Gynaecol. 2012, 26, 391–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataruka, S.; Modrak, M.; Kinterova, V.; Malik, R.; Zeitler, D.M.; Horvat, F.; Kanka, J.; Meister, G.; Svoboda, P. MicroRNA dilution during oocyte growth disables the microRNA pathway in mammalian oocytes. Nucleic Acids Res. 2020, 48, 8050–8062. [Google Scholar] [CrossRef]
- Keefe, D.; Kumar, M.; Kalmbach, K. Oocyte competency is the key to embryo potential. Fertil. Steril. 2015, 103, 317–322. [Google Scholar] [CrossRef]
- Dong, J.P.; Dai, Z.H.; Jiang, Z.X.; He, Y.; Wang, L.; Liao, Q.Y.; Sun, N.X.; Wang, Y.N.; Sun, S.H.; Lin, W.; et al. CD24: A marker of granulosa cell subpopulation and a mediator of ovulation. Cell Death Dis. 2019, 10, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hayek, S.; Clarke, H.J. Control of Oocyte Growth and Development by Intercellular Communication Within the Follicular Niche. Results Probl. Cell Differ. 2016, 58, 191–224. [Google Scholar] [PubMed]
- Pan, B.; Li, J. The art of oocyte meiotic arrest regulation. Reprod. Biol. Endocrinol. RBE 2019, 17, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, R.M.; Zhang, Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature 2013, 502, 472–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Aguilera, O.; Depreux, P.; Halby, L.; Arimondo, P.B.; Goossens, L. DNA Methylation Targeting: The DNMT/HMT Crosstalk Challenge. Biomolecules 2017, 7, 3. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Nomura, S. Review of Single-Cell RNA Sequencing in the Heart. Int. J. Mol. Sci. 2020, 21, 8345. [Google Scholar] [CrossRef]
- Westergaard, C.G.; Byskov, A.G.; Andersen, C.Y. Morphometric characteristics of the primordial to primary follicle transition in the human ovary in relation to age. Hum. Reprod. 2007, 22, 2225–2231. [Google Scholar] [CrossRef] [Green Version]
- Pongsuthirak, P.; Vutyavanich, T. Developmental competence of human embryos derived from in vitro maturation of immature oocytes retrieved during cesarean section. J. Obstet. Gynaecol. Res. 2014, 40, 459–464. [Google Scholar] [CrossRef]
- Reyes, J.M.; Ross, P.J. Cytoplasmic polyadenylation in mammalian oocyte maturation. Wiley Interdiscip Rev RNA 2016, 7, 71–89. [Google Scholar] [CrossRef]
- Warzych, E.; Lipinska, P. Energy metabolism of follicular environment during oocyte growth and maturation. J. Reprod. Dev. 2020, 66, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer-Vaquer, A.; Barragán, M.; Rodríguez, A.; Vassena, R. Altered cytoplasmic maturation in rescued in vitro matured oocytes. Hum. Reprod. 2019, 34, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, T.; Zhao, Y.; Tan, T.; Liu, C.; Liu, Y.; Chang, L.; Huang, N.; Li, C.; Fan, Y.; et al. Single-Cell Transcriptomics of Human Oocytes: Environment-Driven Metabolic Competition and Compensatory Mechanisms During Oocyte Maturation. Antioxid. Redox Signal. 2019, 30, 542–559. [Google Scholar] [CrossRef] [PubMed]
- Collado-Fernandez, E.; Picton, H.M.; Dumollard, R. Metabolism throughout follicle and oocyte development in mammals. Int. J. Dev. Biol. 2012, 56, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.J.; Liu, X.; Chen, L.; Zhang, S.; Zhang, X.; Hao, C.; Miao, Y.L. Advanced maternal age alters expression of maternal effect genes that are essential for human oocyte quality. Aging 2020, 12, 3950–3961. [Google Scholar] [CrossRef]
- Zhang, T.; Xi, Q.; Wang, D.; Li, J.; Wang, M.; Li, D.; Zhu, L.; Jin, L. Mitochondrial dysfunction and endoplasmic reticulum stress involved in oocyte aging: An analysis using single-cell RNA-sequencing of mouse oocytes. J. Ovarian Res. 2019, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.E.; Mihalas, B.P.; Bromfield, E.G.; Roman, S.D.; Nixon, B.; Sutherland, J.M. Autophagy in Female Fertility: A Role in Oxidative Stress and Aging. Antioxid. Redox Signal. 2020, 32, 550–568. [Google Scholar] [CrossRef]
- Buyalos, R.P.; Lee, C.T. Polycystic ovary syndrome: Pathophysiology and outcome with in vitro fertilization. Fertil. Steril. 1996, 65, 1–10. [Google Scholar]
- Qi, L.; Liu, B.; Chen, X.; Liu, Q.; Li, W.; Lv, B.; Xu, X.; Wang, L.; Zeng, Q.; Xue, J.; et al. Single-Cell Transcriptomic Analysis Reveals Mitochondrial Dynamics in Oocytes of Patients With Polycystic Ovary Syndrome. Front. Genet. 2020, 11, 396. [Google Scholar] [CrossRef]
- Faraci, C.; Annis, S.; Jin, J.; Li, H.; Khrapko, K.; Woods, D.C. Impact of exercise on oocyte quality in the POLG mitochondrial DNA mutator mouse. Reproduction 2018, 156, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Grive, K.J. Pathways coordinating oocyte attrition and abundance during mammalian ovarian reserve establishment. Mol. Reprod. Dev. 2020, 87, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Testing and interpreting measures of ovarian reserve: A committee opinion. Fertil. Steril. 2020, 114, 1151–1157. [CrossRef] [PubMed]
- Stimpfel, M.; Skutella, T.; Cvjeticanin, B.; Meznaric, M.; Dovc, P.; Novakovic, S.; Cerkovnik, P.; Vrtacnik-Bokal, E.; Virant-Klun, I. Isolation, characterization and differentiation of cells expressing pluripotent/multipotent markers from adult human ovaries. Cell Tissue Res. 2013, 354, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.J.; Woods, D.C.; Tilly, J.L. Implications and Current Limitations of Oogenesis from Female Germline or Oogonial Stem Cells in Adult Mammalian Ovaries. Cells 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef]
- White, Y.A.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Xu, B.; Li, X.; Ma, W.; Zhang, P.; Chen, X.; Wu, J. Tracing and Characterizing the Development of Transplanted Female Germline Stem Cells In Vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1408–1419. [Google Scholar] [CrossRef] [Green Version]
- Silvestris, E.; Cafforio, P.; D’Oronzo, S.; Felici, C.; Silvestris, F.; Loverro, G. In vitro differentiation of human oocyte-like cells from oogonial stem cells: Single-cell isolation and molecular characterization. Hum. Reprod. 2018, 33, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, Y.L.; Weatherall, E.; Waterfall, M.; McLaughlin, M.; Lu, H.; Skehel, P.A.; Anderson, R.A.; Telfer, E.E. Extracellular Localisation of the C-Terminus of DDX4 Confirmed by Immunocytochemistry and Fluorescence-Activated Cell Sorting. Cells 2019, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Maidarti, M.; Anderson, R.A.; Telfer, E.E. Crosstalk between PTEN/PI3K/Akt Signalling and DNA Damage in the Oocyte: Implications for Primordial Follicle Activation, Oocyte Quality and Ageing. Cells 2020, 9, 200. [Google Scholar] [CrossRef] [Green Version]
- Devos, M.; Grosbois, J.; Demeestere, I. Interaction between PI3K/AKT and Hippo pathways during in vitro follicular activation and response to fragmentation and chemotherapy exposure using a mouse immature ovary model. Biol. Reprod. 2020, 102, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Ilicic, T.; Kim, J.K.; Kolodziejczyk, A.A.; Bagger, F.O.; McCarthy, D.J.; Marioni, J.C.; Teichmann, S.A. Classification of low quality cells from single-cell RNA-seq data. Genome Biol. 2016, 17, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhartiya, D.; Sharma, D. Ovary does harbor stem cells—Size of the cells matter! J. Ovarian Res. 2020, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- Nicol, B.; Grimm, S.A.; Chalmel, F.; Lecluze, E.; Pannetier, M.; Pailhoux, E.; Dupin-De-Beyssat, E.; Guiguen, Y.; Capel, B.; Yao, H.H. RUNX1 maintains the identity of the fetal ovary through an interplay with FOXL2. Nat. Commun. 2019, 10, 5116. [Google Scholar] [CrossRef] [Green Version]
- De Cian, M.C.; Pauper, E.; Bandiera, R.; Vidal, V.P.; Sacco, S.; Gregoire, E.P.; Chassot, A.A.; Panzolini, C.; Wilhelm, D.; Pailhoux, E.; et al. Amplification of R-spondin1 signaling induces granulosa cell fate defects and cancers in mouse adult ovary. Oncogene 2017, 36, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Liu, B.; Wang, H.; Sun, G.; Feng, L.; Chen, Z.; Zhou, J.; Zhang, J.; Zhang, T.; He, M.; et al. SP1 governs primordial folliculogenesis by regulating pregranulosa cell development in mice. J. Mol. Cell Biol. 2020, 12, 230–244. [Google Scholar] [CrossRef] [Green Version]
- Habara, O.; Logan, C.Y.; Kanai-Azuma, M.; Nusse, R.; Takase, H.M. WNT signaling in pre-granulosa cells is required for ovarian folliculogenesis and female fertility. Development 2021, 148, dev198846. [Google Scholar] [CrossRef]
- Stévant, I.; Kühne, F.; Greenfield, A.; Chaboissier, M.C.; Dermitzakis, E.T.; Nef, S. Dissecting Cell Lineage Specification and Sex Fate Determination in Gonadal Somatic Cells Using Single-Cell Transcriptomics. Cell Rep. 2019, 26, 3272–3283.e3. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Mark-Kappeler, C.J.; Hoyer, P.B.; Pepling, M.E. The steroid hormone environment during primordial follicle formation in perinatal mouse ovaries. Biol. Reprod. 2014, 91, 68. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, B.; Xia, G. Mechanisms controlling germline cyst breakdown and primordial follicle formation. Cell. Mol. Life Sci. CMLS 2017, 74, 2547–2566. [Google Scholar] [CrossRef]
- Mork, L.; Maatouk, D.M.; McMahon, J.A.; Guo, J.J.; Zhang, P.; McMahon, A.P.; Capel, B. Temporal differences in granulosa cell specification in the ovary reflect distinct follicle fates in mice. Biol. Reprod. 2012, 86, 37. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhu, J.; Zhou, X. Statistical analysis of spatial expression patterns for spatially resolved transcriptomic studies. Nat. Methods 2020, 17, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, O.A.; Bartels, T.; Holmqvist, S.; Kleshchevnikov, V.; Martirosyan, A.; Polioudakis, D.; Ben Haim, L.; Young, A.M.H.; Batiuk, M.Y.; Prakash, K.; et al. Astrocyte layers in the mammalian cerebral cortex revealed by a single-cell in situ transcriptomic map. Nat. Neurosci. 2020, 23, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Camaioni, A.; Ucci, M.A.; Campagnolo, L.; De Felici, M.; Klinger, F.G. The process of ovarian aging: It is not just about oocytes and granulosa cells. J. Assist. Reprod. Genet. 2022, 39, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Kranc, W.; Budna, J.; Kahan, R.; Chachuła, A.; Bryja, A.; Ciesiółka, S.; Borys, S.; Antosik, M.P.; Bukowska, D.; Brussow, K.P.; et al. Molecular basis of growth, proliferation, and differentiation of mammalian follicular granulosa cells. J. Biol. Regul. Homeost. Agents 2017, 31, 1–8. [Google Scholar]
- Turathum, B.; Gao, E.M.; Chian, R.C. The Function of Cumulus Cells in Oocyte Growth and Maturation and in Subsequent Ovulation and Fertilization. Cells 2021, 10, 2292. [Google Scholar] [CrossRef]
- Khristi, V.; Chakravarthi, V.P.; Singh, P.; Ghosh, S.; Pramanik, A.; Ratri, A.; Borosha, S.; Roby, K.F.; Wolfe, M.W.; Rumi, M.A.K. ESR2 regulates granulosa cell genes essential for follicle maturation and ovulation. Mol. Cell. Endocrinol. 2018, 474, 214–226. [Google Scholar] [CrossRef]
- Glamoclija, V.; Vilović, K.; Saraga-Babić, M.; Baranović, A.; Sapunar, D. Apoptosis and active caspase-3 expression in human granulosa cells. Fertil. Steril. 2005, 83, 426–431. [Google Scholar] [CrossRef]
- Tatone, C.; Amicarelli, F. The aging ovary--the poor granulosa cells. Fertil. Steril. 2013, 99, 12–17. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Swelum, A.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Abdo, M. Cellular and functional adaptation to thermal stress in ovarian granulosa cells in mammals. J. Biol. 2020, 92, 102688. [Google Scholar] [CrossRef]
- Alam, M.H.; Miyano, T. Interaction between growing oocytes and granulosa cells in vitro. Reprod. Med. Biol. 2020, 19, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellix, M.T.; Menaker, M. Circadian clocks in the ovary. Trends Endocrinol. Metab. TEM 2010, 21, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Diaz, F.J. Ovulatory signals alter granulosa cell behavior through YAP1 signaling. Reprod. Biol. Endocrinol. RBE 2019, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Shirafuta, Y.; Tamura, I.; Ohkawa, Y.; Maekawa, R.; Doi-Tanaka, Y.; Takagi, H.; Mihara, Y.; Shinagawa, M.; Taketani, T.; Sato, S.; et al. Integrated Analysis of Transcriptome and Histone Modifications in Granulosa Cells During Ovulation in Female Mice. Endocrinology 2021, 162, bqab128. [Google Scholar] [CrossRef]
- Choi, Y.; Rosewell, K.L.; Brännström, M.; Akin, J.W.; Curry, T.E., Jr.; Jo, M. FOS, a Critical Downstream Mediator of PGR and EGF Signaling Necessary for Ovulatory Prostaglandins in the Human Ovary. J. Clin. Endocrinol. Metab. 2018, 103, 4241–4252. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.H.K.; Murphy, B.D. Nuclear receptors: Key regulators of somatic cell functions in the ovulatory process. Mol. Asp. Med. 2021, 78, 100937. [Google Scholar] [CrossRef]
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E. Ovulation: Parallels With Inflammatory Processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef] [Green Version]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Coordination of Ovulation and Oocyte Maturation: A Good Egg at the Right Time. Endocrinology 2018, 159, 3209–3218. [Google Scholar] [CrossRef]
- Park, C.J.; Lin, P.C.; Zhou, S.; Barakat, R.; Bashir, S.T.; Choi, J.M.; Cacioppo, J.A.; Oakley, O.R.; Duffy, D.M.; Lydon, J.P.; et al. Progesterone Receptor Serves the Ovary as a Trigger of Ovulation and a Terminator of Inflammation. Cell Rep. 2020, 31, 107496. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, X.; Mei, S. Autophagy in Ovarian Follicular Development and Atresia. Int. J. Biol. Sci. 2019, 15, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Wang, H.; Zhang, Y.; Zhang, J.; Liu, J.; Pan, Z. circRNA-Mediated Inhibin-Activin Balance Regulation in Ovarian Granulosa Cell Apoptosis and Follicular Atresia. Int. J. Mol. Sci. 2021, 22, 9113. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Van der Hoek, K.H.; Ryan, N.K.; Norman, R.J.; Robker, R.L. Macrophage contributions to ovarian function. Hum. Reprod. Update 2004, 10, 119–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townson, D.H.; Liptak, A.R. Chemokines in the corpus luteum: Implications of leukocyte chemotaxis. Reprod. Biol. Endocrinol. RBE 2003, 1, 94. [Google Scholar] [CrossRef] [Green Version]
- Petrovská, M.; Dimitrov, D.G.; Michael, S.D. Quantitative changes in macrophage distribution in normal mouse ovary over the course of the estrous cycle examined with an image analysis system. Am. J. Reprod. Immunol. 1996, 36, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Zhang, X.; Li, N.; Zhang, D.; Zhao, S.; Dang, Y.; Zanvit, P.; Jin, W.; Chen, Z.J.; Chen, W.; et al. T(reg) deficiency-mediated T(H) 1 response causes human premature ovarian insufficiency through apoptosis and steroidogenesis dysfunction of granulosa cells. Clin. Transl. Med. 2021, 11, e448. [Google Scholar] [CrossRef]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. N-Cadherin Is Critical for the Survival of Germ Cells, the Formation of Steroidogenic Cells, and the Architecture of Developing Mouse Gonads. Cells 2019, 8, 1610. [Google Scholar] [CrossRef] [Green Version]
- Cajas, Y.N.; Cañón-Beltrán, K.; Ladrón de Guevara, M.; Millán de la Blanca, M.G.; Ramos-Ibeas, P.; Gutiérrez-Adán, A.; Rizos, D.; González, E.M. Antioxidant Nobiletin Enhances Oocyte Maturation and Subsequent Embryo Development and Quality. Int. J. Mol. Sci. 2020, 21, 5340. [Google Scholar] [CrossRef]
- Li, S.H.; Lin, M.H.; Hwu, Y.M.; Lu, C.H.; Yeh, L.Y.; Chen, Y.J.; Lee, R.K. Correlation of cumulus gene expression of GJA1, PRSS35, PTX3, and SERPINE2 with oocyte maturation, fertilization, and embryo development. Reprod. Biol. Endocrinol. RBE 2015, 13, 93. [Google Scholar] [CrossRef] [Green Version]
- Iwata, H. Age-associated changes in granulosa cells and follicular fluid in cows. J. Reprod. Dev. 2017, 63, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Eslami, M.; Esfandyari, S.; Aghahosseini, M.; Rashidi, Z.; Hosseinishental, S.H.; Brenjian, S.; Sobhani, A.; Amidi, F. Astaxanthin Protects Human Granulosa Cells against Oxidative Stress through Activation of NRF2/ARE Pathway and Its Downstream Phase II Enzymes. Cell J. 2021, 23, 319–328. [Google Scholar]
- Nie, Z.; Hua, R.; Zhang, Y.; Zhang, N.; Zhang, Y.; Li, Q.; Wu, H. Resveratrol protects human luteinised granulosa cells against hydrogen peroxide-induced oxidative injury through the Sirt1. Reprod. Fertil. Dev. 2021, 33, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Reeves, G. Specific stroma in the cortex and medulla of the ovary. Cell types and vascular supply in relation to follicular apparatus and ovulation. Obstet. Gynecol. 1971, 37, 832–844. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, H.M.; Tomaszewski, C.E.; Chang, F.L.; Moravek, M.B.; Xu, M.; Padmanabhan, V.; Shikanov, A. The ovarian stroma as a new frontier. Reproduction 2020, 160, R25–R39. [Google Scholar] [CrossRef] [PubMed]
- Grasa, P.; Sheikh, S.; Krzys, N.; Millar, K.; Janjua, S.; Nawaggi, P.; Williams, S.A. Dysregulation of follicle development in a mouse model of premature ovarian insufficiency. Reproduction 2016, 152, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharma, S.J.; Kelkar, R.L.; Nandedkar, T.D. Fas and Fas ligand protein and mRNA in normal and atretic mouse ovarian follicles. Reproduction 2003, 126, 783–789. [Google Scholar] [CrossRef]
- Lin, L.; Rodrigues, F.; Kary, C.; Contet, A.; Logan, M.; Baxter, R.H.G.; Wood, W.; Baehrecke, E.H. Complement-Related Regulates Autophagy in Neighboring Cells. Cell 2017, 170, 158–171.e8. [Google Scholar] [CrossRef] [Green Version]
- Winterhoff, B.J.; Maile, M.; Mitra, A.K.; Sebe, A.; Bazzaro, M.; Geller, M.A.; Abrahante, J.E.; Klein, M.; Hellweg, R.; Mullany, S.A.; et al. Single cell sequencing reveals heterogeneity within ovarian cancer epithelium and cancer associated stromal cells. Gynecol. Oncol. 2017, 144, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Yu, T.; Han, Y.; Jiang, H.; Wang, C.; You, T.; Zhao, X.; Shan, H.; Yang, R.; Yang, L.; et al. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding miR-101-3p to regulate ZEB1 expression. Mol. Cancer 2018, 17, 119. [Google Scholar] [CrossRef]
- Talbot, P.; Chacon, R.S. In vitro ovulation of hamster oocytes depends on contraction of follicular smooth muscle cells. J. Exp. Zool. 1982, 224, 409–415. [Google Scholar] [CrossRef]
- Miyamoto, A.; Shirasuna, K.; Shimizu, T.; Bollwein, H.; Schams, D. Regulation of corpus luteum development and maintenance: Specific roles of angiogenesis and action of prostaglandin F2alpha. Soc. Reprod. Fertil. Suppl. 2010, 67, 289–304. [Google Scholar]
- Martin, G.G.; Van Steenwyk, G.; Miller-Walker, C. The fate of thecal smooth muscle cells in postovulatory hamster follicles. Anat. Rec. 1983, 207, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Kizuka-Shibuya, F.; Tokuda, N.; Takagi, K.; Adachi, Y.; Lee, L.; Tamura, I.; Maekawa, R.; Tamura, H.; Suzuki, T.; Owada, Y.; et al. Locally existing endothelial cells and pericytes in ovarian stroma, but not bone marrow-derived vascular progenitor cells, play a central role in neovascularization during follicular development in mice. J. Ovarian Res. 2014, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.S.; Rueda, B.R.; Spanel-Borowski, K. Microvascular endothelial cells of the corpus luteum. Reprod. Biol. Endocrinol. RBE 2003, 1, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Fujii, S.; Ryan, N.K.; Van der Hoek, K.H.; Jasper, M.J.; Sini, I.; Robertson, S.A.; Robker, R.L.; Norman, R.J. Ovarian leukocyte distribution and cytokine/chemokine mRNA expression in follicular fluid cells in women with polycystic ovary syndrome. Hum. Reprod. 2007, 22, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walusimbi, S.S.; Pate, J.L. Physiology and Endocrinology Symposium: Role of immune cells in the corpus luteum. J. Anim. Sci. 2013, 91, 1650–1659. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Burns, K.H.; Viveiros, M.M.; Eppig, J.J. Intercellular communication in the mammalian ovary: Oocytes carry the conversation. Science 2002, 296, 2178–2180. [Google Scholar] [CrossRef]
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef] [Green Version]
- Porras-Gómez, T.J.; Moreno-Mendoza, N. Interaction between oocytes, cortical germ cells and granulosa cells of the mouse and bat, following the dissociation-re-aggregation of adult ovaries. Zygote 2020, 28, 223–232. [Google Scholar] [CrossRef]
- Komatsu, K.; Masubuchi, S. Mouse oocytes connect with granulosa cells by fusing with cell membranes and form a large complex during follicle development. Biol. Reprod. 2018, 99, 527–535. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, L.K.; Limback, S.D.; Albertini, D.F. Signaling modalities during oogenesis in mammals. Curr. Top. Dev. Biol. 2013, 102, 227–242. [Google Scholar]
- Buccione, R.; Schroeder, A.C.; Eppig, J.J. Interactions between somatic cells and germ cells throughout mammalian oogenesis. Biol. Reprod. 1990, 43, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, F.; Smitz, J. Molecular control of oogenesis. Biochim. Biophys. Acta 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, H.J. Regulation of germ cell development by intercellular signaling in the mammalian ovarian follicle. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e294. [Google Scholar] [CrossRef]
- Chang, H.M.; Qiao, J.; Leung, P.C. Oocyte-somatic cell interactions in the human ovary-novel role of bone morphogenetic proteins and growth differentiation factors. Hum. Reprod. Update 2016, 23, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Overeem, A.W.; Chang, Y.W.; Spruit, J.; Roelse, C.M.; Chuva De Sousa Lopes, S.M. Ligand-Receptor Interactions Elucidate Sex-Specific Pathways in the Trajectory From Primordial Germ Cells to Gonia During Human Development. Front. Cell Dev. Biol. 2021, 9, 661243. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Giladi, A.; Gorki, A.D.; Solodkin, D.G.; Zada, M.; Hladik, A.; Miklosi, A.; Salame, T.M.; Halpern, K.B.; David, E.; et al. Lung Single-Cell Signaling Interaction Map Reveals Basophil Role in Macrophage Imprinting. Cell 2018, 175, 1031–1044.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Kuang, H.; Ansari, S.; Liu, T.; Gong, J.; Wang, S.; Zhao, X.Y.; Ji, Y.; Li, C.; Guo, L.; et al. Landscape of Intercellular Crosstalk in Healthy and NASH Liver Revealed by Single-Cell Secretome Gene Analysis. Mol. Cell 2019, 75, 644–660.e5. [Google Scholar] [CrossRef] [PubMed]
- Ximerakis, M.; Lipnick, S.L.; Innes, B.T.; Simmons, S.K.; Adiconis, X.; Dionne, D.; Mayweather, B.A.; Nguyen, L.; Niziolek, Z.; Ozek, C.; et al. Single-cell transcriptomic profiling of the aging mouse brain. Nat. Neurosci. 2019, 22, 1696–1708. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, H.; Wan, L.; Wang, Z.; Wang, H.; Ge, C.; Liu, Y.; Hao, Y.; Zhang, D.; Shi, G.; et al. Single-cell transcriptomic architecture and intercellular crosstalk of human intrahepatic cholangiocarcinoma. J. Hepatol. 2020, 73, 1118–1130. [Google Scholar] [CrossRef]
- Kim, B.; Kim, Y.; Cooke, P.S.; Rüther, U.; Jorgensen, J.S. The fused toes locus is essential for somatic-germ cell interactions that foster germ cell maturation in developing gonads in mice. Biol. Reprod. 2011, 84, 1024–1032. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Chen, Y.; Shu, Y.; Cheng, Y.; Qiao, J.; Behr, B.; Pera, R.A.; Hsueh, A.J. Promotion of human early embryonic development and blastocyst outgrowth in vitro using autocrine/paracrine growth factors. PLoS One 2012, 7, e493282012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K. Regulation of primordial follicle assembly and development. Hum. Reprod. Update 2005, 11, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, D.; Busayavalasa, K.; Zhang, J.; Hu, M.; Risal, S.; Bayazit, M.B.; Singh, M.; Diril, M.K.; Kaldis, P.; Liu, K. Inhibitory phosphorylation of Cdk1 mediates prolonged prophase I arrest in female germ cells and is essential for female reproductive lifespan. Cell Res. 2016, 26, 1212–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, A.; Nilsson, E.; Skinner, M.K. Cytokine (IL16) and tyrphostin actions on ovarian primordial follicle development. Reproduction 2014, 148, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kezele, P.; Nilsson, E.; Skinner, M.K. Cell-cell interactions in primordial follicle assembly and development. Front. Biosci. A J. Virtual Libr. 2002, 7, d1990-62002. [Google Scholar] [CrossRef] [Green Version]
- Cecconi, S.; Ciccarelli, C.; Barberi, M.; Macchiarelli, G.; Canipari, R. Granulosa cell-oocyte interactions. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115 (Suppl. 1), S19–S22. [Google Scholar] [CrossRef]
- Wu, Z.; Guan, K.L. Hippo Signaling in Embryogenesis and Development. Trends Biochem. Sci. 2021, 46, 51–63. [Google Scholar] [CrossRef]
- Lew, R. Natural history of ovarian function including assessment of ovarian reserve and premature ovarian failure. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 55, 2–13. [Google Scholar] [CrossRef]
- Rimon-Dahari, N.; Yerushalmi-Heinemann, L.; Alyagor, L.; Dekel, N. Ovarian Folliculogenesis. Results Probl. Cell Differ. 2016, 58, 167–190. [Google Scholar]
- Ford, E.A.; Beckett, E.L.; Roman, S.D.; McLaughlin, E.A.; Sutherland, J.M. Advances in human primordial follicle activation and premature ovarian insufficiency. Reprod. 2020, 159, R15–R29. [Google Scholar] [CrossRef] [Green Version]
- Nagamatsu, G. Regulation of primordial follicle formation, dormancy, and activation in mice. J. Reprod. Dev. 2021, 67, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, A.J.; Kawamura, K.; Cheng, Y.; Fauser, B.C. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, R.; Campbell, B.K. Development of the dominant follicle: Mechanisms of selection and maintenance of oocyte quality. Soc. Reprod. Fertil. Suppl. 2007, 64, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Knight, P.G.; Glister, C. TGF-beta superfamily members and ovarian follicle development. Reproduction 2006, 132, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; He, M.; Zhao, L.; Qin, S.; Zhu, Z.; Du, X.; Zhou, B.; Yang, Y.; Liu, X.; Xia, G.; et al. HDAC6 regulates primordial follicle activation through mTOR signaling pathway. Cell Death Dis. 2021, 12, 559. [Google Scholar] [CrossRef]
- Dissen, G.A.; Romero, C.; Hirshfield, A.N.; Ojeda, S.R. Nerve growth factor is required for early follicular development in the mammalian ovary. Endocrinology 2001, 142, 2078–2086. [Google Scholar] [CrossRef]
- Harel, S.; Jin, S.; Fisch, B.; Feldberg, D.; Krissi, H.; Felz, C.; Freimann, S.; Tan, S.L.; Ao, A.; Abir, R. Tyrosine kinase B receptor and its activated neurotrophins in ovaries from human fetuses and adults. Mol. Hum. Reprod. 2006, 12, 357–365. [Google Scholar] [CrossRef]
- Macklon, N.S.; Fauser, B.C. Aspects of ovarian follicle development throughout life. Horm. Res. 1999, 52, 161–170. [Google Scholar] [CrossRef]
- Gougeon, A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Ann. D’endocrinol. 2010, 71, 132–143. [Google Scholar] [CrossRef]
- Bachelot, A.; Gilleron, J.; Meduri, G.; Guberto, M.; Dulon, J.; Boucherie, S.; Touraine, P.; Misrahi, M. A common African variant of human connexin 37 is associated with Caucasian primary ovarian insufficiency and has a deleterious effect in vitro. Int. J. Mol. Med. 2018, 41, 640–648. [Google Scholar]
- Chambers, D.C.; Carew, A.M.; Lukowski, S.W.; Powell, J.E. Transcriptomics and single-cell RNA-sequencing. Respirology 2019, 24, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, E.; Deng, Q. Single-cell RNA sequencing: Technical advancements and biological applications. Mol. Asp. Med. 2018, 59, 36–46. [Google Scholar] [CrossRef] [PubMed]
- García-Castro, H.; Solana, J. Single-cell transcriptomics in planaria: New tools allow new insights into cellular and evolutionary features. Biochem. Soc. Trans. 2022, 50, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Pensold, D.; Zimmer-Bensch, G. Methods for Single-Cell Isolation and Preparation. Adv. Exp. Med. Biol. 2020, 1255, 7–27. [Google Scholar]
- Zhai, X.; Wang, W.; Dou, D.; Ma, Y.; Gang, D.; Jiang, Z.; Shi, B.; Jin, B. A novel technique to prepare a single cell suspension of isolated quiescent human hepatic stellate cells. Sci. Rep. 2019, 9, 12757. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, J.B.; George, J.; Christenson, L.K. Non-coding RNA in Ovarian Development and Disease. Adv. Exp. Med. Biol. 2016, 886, 79–93. [Google Scholar]
- Li, D.; Xu, D.; Zou, Y.; Xu, Y.; Fu, L.; Xu, X.; Liu, Y.; Zhang, X.; Zhang, J.; Ming, H.; et al. Non-coding RNAs and ovarian diseases (Review). Mol. Med. Rep. 2017, 15, 1435–1440. [Google Scholar] [CrossRef] [Green Version]
- Longo, S.K.; Guo, M.G.; Ji, A.L.; Khavari, P.A. Integrating single-cell and spatial transcriptomics to elucidate intercellular tissue dynamics. Nat. Rev. Genet. 2021, 22, 627–644. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cell Type | Marker Genes | Species | Technology | Reference | |
|---|---|---|---|---|---|
| Fetal germ cells (FGCs) | FGCs | DDX4, DAZL, POU5F1, PRDM1, TFAP2C, KIT, NANOG, SALL4, LIN28A, LEFTY1, LEFTY2 | Mouse | 10XGenomics | [47] |
| DDX4, DAZL | Mouse | 10XGenomics | [48] | ||
| Mitotic FGCs | POU5F1, NANOG | Human | Smart-seq2 | [39] | |
| Retinoid-acid-signaling-responsive FGCs | STRA8, ZGLP1, ANHX, ASB9, THRA/BTR | Human | Smart-seq2 | [39] | |
| Meiotic prophase FGCs | Il13RA2 | Human | Smart-seq2 | [39] | |
| Oogenesis stage FGCs | PECAM1, ZP3, OOSP2 | Human | Smart-seq2 | [39] | |
| Oocyte | GDF9, ZP3, FIGLA, OOSP2 | Human | 10XGenomics | [34] | |
| DDX4, ZP2, SYCP3, SOX30, ZAR1, DAZL | Human | Tang method | [49] | ||
| DDX4, ZP2, FIGLA, SOX30 | Human | Smart-seq2 | [50] | ||
| GDF9, DDX4, SYCP3, ZP3, LMOD3, RBM46 NET01 | Monkey | STRT-seq | [45] | ||
| DDX4, DAZL | Mouse | 10XGenomics | [36,38] | ||
| DAZL | Mouse | 10XGenomics | [37] | ||
| Pregranulosa cells | FOXL2, WNT4, WNT6, KITL | Mouse | 10XGenomics | [48] | |
| Granulosa cells (GCs) | FOXL2, WT1 | Human | Smart-seq2 | [39] | |
| FOXL2, AMH | Human | 10XGenomics | [34] | ||
| AMH, CYP11A1, STAR, INHBA | Human | 10XGenomics | [49] | ||
| FOXL2, CDH2, GJA1, TNNI3 | Human | Smart-seq2 | [50] | ||
| AMH, HSD17B1, SERPINE2, GSTA1 | Human | 10XGenomics | [35] | ||
| AMH, WT1, INH4 | Monkey | STRT-seq | [45] | ||
| AMHR2, KITL | Mouse | 10XGenomics | [36,38] | ||
| AMHR2 | Mouse | 10XGenomics | [37] | ||
| AMH, AMHR2, KITL, CYP19A1, FSHR | Mouse | 10XGenomics | [51] | ||
| Cumulus GCs | IGFBP2, INHBB, IHH | Human | Smart-seq2 | [50] | |
| IGFBP2, INHBB, IHH, VCAN, FST, HTRA1 | Human | 10XGenomics | [35] | ||
| HAS2, NPR2 | Mouse | 10XGenomics | [51] | ||
| Mural GCs | KRT18, AKIRIN1, CYP19A1, | Human | Smart-seq2 | [50] | |
| KRT18, AKIRIN1, LIHP, CITED2 | Human | 10XGenomics | [35] | ||
| FSHR, BMPR2, NPPC | Mouse | 10XGenomics | [51] | ||
| Stromal cells | DCN, COLLA1, COL6A1, PDGFRA | Human | 10XGenomics | [34] | |
| DCN, COLLA1, COL3A1, APOE | Human | Smart-seq2 | [50] | ||
| DCN, LUM | Human | 10XGenomics | [35] | ||
| TCF21, COLLA2 | Monkey | STRT-seq | [45] | ||
| MFAP4, NR2F2 | Mouse | 10XGenomics | [38] | ||
| TCF21, NR2F2 | Mouse | 10XGenomics | [36] | ||
| MFAP4 | Mouse | 10XGenomics | [37] | ||
| Smooth muscle cells | TAGLN, RGS5, MYH11, MCAM, RERGl | Human | 10XGenomics | [34] | |
| TAGLN, RGS5 | Human | 10XGenomics, Smart-seq2 | [35,50] | ||
| DES, ACTA2 | Monkey | STRT-seq | [45] | ||
| Endothelial cells | CDH5 (CD144), PECAM1 (CD31) | Human | Smart-seq2 | [39] | |
| vWF, CDH5 | Human | 10XGenomics | [34] | ||
| vWF, CD34 | Human | 10XGenomics | [50] | ||
| vWF, CLDN5 | Human | 10XGenomics | [35] | ||
| vWF, CDH5 | Monkey | STRT-seq | [45] | ||
| APLNR, EGFL7 | Mouse | 10XGenomics | [38] | ||
| APLNR | Mouse | 10XGenomics | [36,37] | ||
| Immune cells | Immune cells | CD69, ITGB2 | Human | 10XGenomics | [34] |
| CD53, CXCR4 | Human | 10XGenomics | [35] | ||
| ELNE, MPO | Mouse | 10XGenomics | [38] | ||
| TYROBP | Mouse | 10XGenomics | [36,37] | ||
| Innate immune cells | CD68, IFI30 | Human | 10XGenomics | [35] | |
| NK cells | CD3d, KLRB1 | Monkey | STRT-seq | [45] | |
| Macrophages | CD68, CD14 | Monkey | STRT-seq | [45] | |
| Antigen-presenting cells | CD14, HLA-DRA, B2M, HLA-DQB1 | Human | 10XGenomics | [34] | |
| T lymphocytes | CD2, CD3G, CD8A | Human | 10XGenomics | [34] | |
| T lymphocytes | AW112010, CD3G | Mouse | 10XGenomics | [51] | |
| B lymphocytes | IGHM, CD37 | Mouse | 10XGenomics | [51] | |
| Monocytes or monocyte-derived cells | CD14 | Mouse | 10XGenomics | [51] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Zhang, Y.; Ai, J.; Li, K. Application of Single-Cell RNA Sequencing in Ovarian Development. Biomolecules 2023, 13, 47. https://doi.org/10.3390/biom13010047
Gong X, Zhang Y, Ai J, Li K. Application of Single-Cell RNA Sequencing in Ovarian Development. Biomolecules. 2023; 13(1):47. https://doi.org/10.3390/biom13010047
Chicago/Turabian StyleGong, Xiaoqin, Yan Zhang, Jihui Ai, and Kezhen Li. 2023. "Application of Single-Cell RNA Sequencing in Ovarian Development" Biomolecules 13, no. 1: 47. https://doi.org/10.3390/biom13010047