

Message in a Scaffold: Natural Biomaterials for Three-Dimensional (3D) Bioprinting of Human Brain Organoids

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Brain Organoids: Potential and Limits
3. Three-Dimensional Bioprinting: Benefits and Challenges
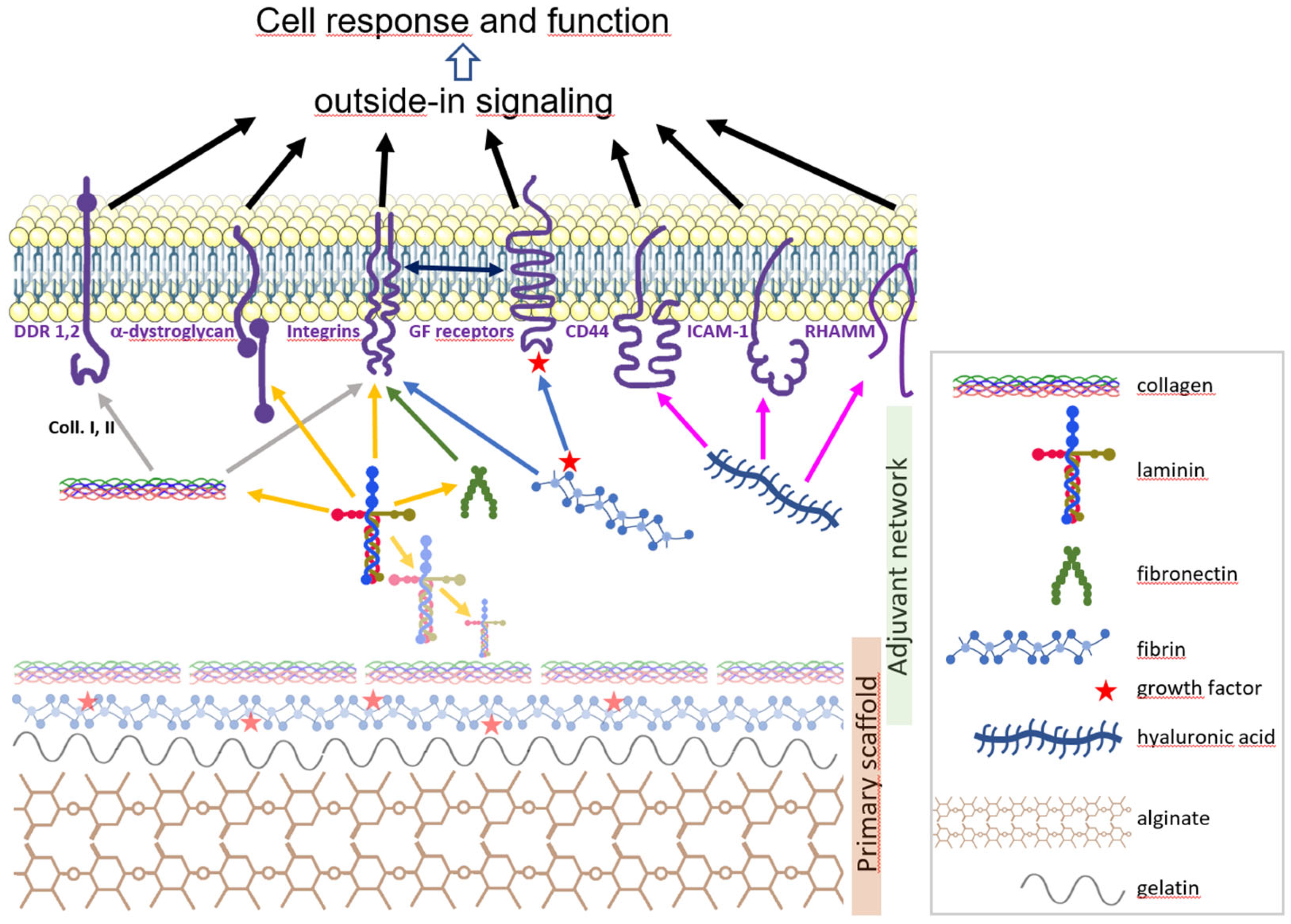
4. The Brain Extracellular Matrix
5. Scaffold Hydrogel Biomaterials
5.1. Scaffolds from Extracellular Matrix Extracts
5.2. Scaffolds from Single Polymers: Agarose and Alginate
6. Natural Adjuvants
6.1. Collagen
6.2. Fibrin
6.3. Gelatine
6.4. Hyaluronic Acid
6.5. Laminin
6.6. Other Natural Adjuvants: Fibronectin, Chitosan and Tenascin
7. Simple Adjuvant Molecules
8. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Feigin, V.L.; Vos, T.; Nichols, E.; Owolabi, M.O.; Carroll, W.M.; Dichgans, M.; Deuschl, G.; Parmar, P.; Brainin, M.; Murray, C. The Global Burden of Neurological Disorders: Translating Evidence into Policy. Lancet Neurol. 2020, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- McGraw, C.M.; Ward, C.S.; Samaco, R.C. Genetic Rodent Models of Brain Disorders: Perspectives on Experimental Approaches and Therapeutic Strategies. Am. J. Med. Genet. Part C Semin. Med. Genet. 2017, 175, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, J.J.; Mason, J.O. Mouse vs Man: Organoid Models of Brain Development & Disease. Brain Res. 2019, 1724, 146427. [Google Scholar] [CrossRef]
- Rolland, M.; Martin, H.; Bergamelli, M.; Sellier, Y.; Bessières, B.; Aziza, J.; Benchoua, A.; Leruez-Ville, M.; Gonzalez-Dunia, D.; Chavanas, S. Human Cytomegalovirus Infection Is Associated with Increased Expression of the Lissencephaly Gene PAFAH1B1 Encoding LIS1 in Neural Stem Cells and Congenitally Infected Brains. J. Pathol. 2021, 254, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Shahsavani, M.; Pronk, R.J.; Falk, R.; Lam, M.; Moslem, M.; Linker, S.B.; Salma, J.; Day, K.; Schuster, J.; Anderlid, B.M.; et al. An in Vitro Model of Lissencephaly: Expanding the Role of DCX during Neurogenesis. Mol. Psychiatry 2018, 23, 1674–1684. [Google Scholar] [CrossRef] [Green Version]
- Kajtez, J.; Nilsson, F.; Fiorenzano, A.; Parmar, M.; Emnéus, J. 3D Biomaterial Models of Human Brain Disease. Neurochem. Int. 2021, 147, 105043. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Kelley, K.W.; Pașca, S.P. Human Brain Organogenesis: Toward a Cellular Understanding of Development and Disease. Cell 2022, 185, 42–61. [Google Scholar] [CrossRef]
- Mansour, A.A.F.; Schafer, S.T.; Gage, F.H. Cellular Complexity in Brain Organoids: Current Progress and Unsolved Issues. Semin. Cell Dev. Biol. 2021, 111, 32–39. [Google Scholar] [CrossRef]
- Procès, A.; Luciano, M.; Kalukula, Y.; Ris, L.; Gabriele, S. Multiscale Mechanobiology in Brain Physiology and Diseases. Front. Cell Dev. Biol. 2022, 10, 823857. [Google Scholar] [CrossRef]
- Amirifar, L.; Shamloo, A.; Nasiri, R.; de Barros, N.R.; Wang, Z.Z.; Unluturk, B.D.; Libanori, A.; Ievglevskyi, O.; Diltemiz, S.E.; Sances, S.; et al. Brain-on-a-Chip: Recent Advances in Design and Techniques for Microfluidic Models of the Brain in Health and Disease. Biomaterials 2022, 285, 121531. [Google Scholar] [CrossRef]
- Liu, X.; Inda, M.E.; Lai, Y.; Lu, T.K.; Zhao, X.; Liu, X.; Zhao, X.; Inda, M.E.; Lai, Y.; Lu, T.K. Engineered Living Hydrogels. Adv. Mater. 2022, 34, 2201326. [Google Scholar] [CrossRef]
- Samanipour, R.; Tahmooressi, H.; Rezaei Nejad, H.; Hirano, M.; Shin, S.R.; Hoorfar, M. A Review on 3D Printing Functional Brain Model. Biomicrofluidics 2022, 16, 011501. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Martino, F.; Perestrelo, A.R.; Vinarský, V.; Pagliari, S.; Forte, G. Cellular Mechanotransduction: From Tension to Function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef] [Green Version]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in Vitro Model of Human Development and Disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Trujillo, C.A.; Gao, R.; Negraes, P.D.; Gu, J.; Buchanan, J.; Preissl, S.; Wang, A.; Wu, W.; Haddad, G.G.; Chaim, I.A.; et al. Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development. Cell Stem Cell 2019, 25, 558. [Google Scholar] [CrossRef]
- Samarasinghe, R.A.; Miranda, O.A.; Buth, J.E.; Mitchell, S.; Ferando, I.; Watanabe, M.; Allison, T.F.; Kurdian, A.; Fotion, N.N.; Gandal, M.J.; et al. Identification of Neural Oscillations and Epileptiform Changes in Human Brain Organoids. Nat. Neurosci. 2021, 24, 1488–1500. [Google Scholar] [CrossRef]
- Qian, X.; Song, H.; Ming, G.L. Brain Organoids: Advances, Applications and Challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef] [Green Version]
- Todd, G.K.; Boosalis, C.A.; Burzycki, A.A.; Steinman, M.Q.; Hester, L.D.; Shuster, P.W.; Patterson, R.L. Towards Neuronal Organoids: A Method for Long-Term Culturing of High-Density Hippocampal Neurons. PLoS ONE 2013, 8, e58996. [Google Scholar] [CrossRef]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional Cortical Neurons and Astrocytes from Human Pluripotent Stem Cells in 3D Culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieng, V.; Stoppini, L.; Villy, S.; Fathi, M.; Dubois-Dauphin, M.; Krause, K.-H. Engineering of Midbrain Organoids Containing Long-Lived Dopaminergic Neurons. Stem Cells Dev. 2014, 23, 1535–1547. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Fang, K.-H.; Cao, S.-Y.; Qu, Z.-Y.; Li, Q.; Krencik, R.; Xu, M.; Bhattacharyya, A.; Su, Y.-W.; Zhu, D.-Y.; et al. Efficient Generation of Region-Specific Forebrain Neurons from Human Pluripotent Stem Cells under Highly Defined Condition. Sci. Rep. 2016, 5, 18550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, K.P.; Hewitt, A.W.; Davidson, K.C.; Pébay, A.; Wong, R.C.B. Methods of Retinal Ganglion Cell Differentiation From Pluripotent Stem Cells. Transl. Vis. Sci. Technol. 2014, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Dezonne, R.S.; Sartore, R.C.; Nascimento, J.M.; Saia-Cereda, V.M.; Romão, L.F.; Alves-Leon, S.V.; de Souza, J.M.; Martins-de-Souza, D.; Rehen, S.K.; Gomes, F.C.A. Derivation of Functional Human Astrocytes from Cerebral Organoids. Sci. Rep. 2017, 7, 45091. [Google Scholar] [CrossRef] [Green Version]
- Pandya, H.; Shen, M.J.; Ichikawa, D.M.; Sedlock, A.B.; Choi, Y.; Johnson, K.R.; Kim, G.; Brown, M.A.; Elkahloun, A.G.; Maric, D.; et al. Differentiation of Human and Murine Induced Pluripotent Stem Cells to Microglia-like Cells. Nat. Neurosci. 2017, 20, 753. [Google Scholar] [CrossRef]
- Muffat, J.; Li, Y.; Omer, A.; Durbin, A.; Bosch, I.; Bakiasi, G.; Richards, E.; Meyer, A.; Gehrke, L.; Jaenisch, R. Human Induced Pluripotent Stem Cell-Derived Glial Cells and Neural Progenitors Display Divergent Responses to Zika and Dengue Infections. Proc. Natl. Acad. Sci. USA 2018, 115, 7117–7122. [Google Scholar] [CrossRef] [Green Version]
- Marton, R.M.; Miura, Y.; Sloan, S.A.; Li, Q.; Revah, O.; Levy, R.J.; Huguenard, J.R.; Pașca, S.P. Differentiation and Maturation of Oligodendrocytes in Human Three-Dimensional Neural Cultures. Nat. Neurosci. 2019, 22, 484–491. [Google Scholar] [CrossRef]
- Gabriel, E.; Albanna, W.; Pasquini, G.; Ramani, A.; Josipovic, N.; Mariappan, A.; Schinzel, F.; Karch, C.M.; Bao, G.; Gottardo, M.; et al. Human Brain Organoids Assemble Functionally Integrated Bilateral Optic Vesicles. Cell Stem Cell 2021, 28, 1740–1757.e8. [Google Scholar] [CrossRef]
- Lancaster, M.A. Brain Organoids Get Vascularized. Nat. Biotechnol. 2018, 36, 407–408. [Google Scholar] [CrossRef]
- Mansour, A.A.; Gonçalves, J.T.; Bloyd, C.W.; Li, H.; Fernandes, S.; Quang, D.; Johnston, S.; Parylak, S.L.; Jin, X.; Gage, F.H. An in Vivo Model of Functional and Vascularized Human Brain Organoids. Nat. Biotechnol. 2018, 36, 432–441. [Google Scholar] [CrossRef]
- Costamagna, G.; Comi, G.P.; Corti, S. Advancing Drug Discovery for Neurological Disorders Using IPSC-Derived Neural Organoids. Int. J. Mol. Sci. 2021, 22, 2659. [Google Scholar] [CrossRef]
- Javier-Torrent, M.; Zimmer-Bensch, G.; Nguyen, L. Mechanical Forces Orchestrate Brain Development. Trends Neurosci. 2021, 44, 110–121. [Google Scholar] [CrossRef]
- Wang, H. Modeling Neurological Diseases With Human Brain Organoids. Front. Synaptic Neurosci. 2018, 10, 15. [Google Scholar] [CrossRef]
- Sloan, S.A.; Andersen, J.; Pașca, A.M.; Birey, F.; Pașca, S.P. Generation and Assembly of Human Brain Region–Specific Three-Dimensional Cultures. Nat. Protoc. 2018, 13, 2062–2085. [Google Scholar] [CrossRef]
- Shen, Q.; Goderie, S.K.; Jin, L.; Karanth, N.; Sun, Y.; Abramova, N.; Vincent, P.; Pumiglia, K.; Temple, S. Endothelial Cells Stimulate Self-Renewal and Expand Neurogenesis of Neural Stem Cells. Science 2004, 304, 1338–1340. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.K.; Tsuru, Y.; Hasegawa, K.; Kuwako, K. ichiro Vascularization of Human Brain Organoids. Stem Cells 2021, 39, 1017–1024. [Google Scholar] [CrossRef]
- Ozbolat, I.T.; Peng, W.; Ozbolat, V. Application Areas of 3D Bioprinting. Drug Discov. Today 2016, 21, 1257–1271. [Google Scholar] [CrossRef] [Green Version]
- Ayan, B.; Heo, D.N.; Zhang, Z.; Dey, M.; Povilianskas, A.; Drapaca, C.; Ozbolat, I.T. Aspiration-Assisted Bioprinting for Precise Positioning of Biologics. Sci. Adv. 2020, 6, eaaw5111. [Google Scholar] [CrossRef] [Green Version]
- Kolesky, D.B.; Homan, K.A.; Skylar-Scott, M.A.; Lewis, J.A. Three-Dimensional Bioprinting of Thick Vascularized Tissues. Proc. Natl. Acad. Sci. USA 2016, 113, 3179–3184. [Google Scholar] [CrossRef]
- Hospodiuk, M.; Dey, M.; Sosnoski, D.; Ozbolat, I.T. The Bioink: A Comprehensive Review on Bioprintable Materials. Biotechnol. Adv. 2017, 35, 217–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovett, M.L.; Nieland, T.J.F.; Dingle, Y.T.L.; Kaplan, D.L. Innovations in 3-Dimensional Tissue Models of Human Brain Physiology and Diseases. Adv. Funct. Mater. 2020, 30, 1909146. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Molotnikov, A.; Parkington, H.C.; Meagher, L.; Forsythe, J.S. Extrusion 3D Bioprinting of Functional Self-Supporting Neural Constructs Using a Photoclickable Gelatin Bioink. Biofabrication 2022, 14, 035014. [Google Scholar] [CrossRef] [PubMed]
- Hölzl, K.; Lin, S.; Tytgat, L.; Vlierberghe, S.V.; Gu, L.; Ovsianikov, A. Bioink Properties before, during and after 3D Bioprinting. Biofabrication 2016, 8, 032002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular Matrix Assembly: A Multiscale Deconstruction. Nat. Reviews. Mol. Cell Biol. 2014, 15, 771. [Google Scholar] [CrossRef] [Green Version]
- Walma, D.A.C.; Yamada, K.M. The Extracellular Matrix in Development. Development 2020, 147, dev175596. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.H.; et al. A Guide to the Composition and Functions of the Extracellular Matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Milošević, N.J.; Judaš, M.; Aronica, E.; Kostovic, I. Neural ECM in Laminar Organization and Connectivity Development in Healthy and Diseased Human Brain. Prog. Brain Res. 2014, 214, 159–178. [Google Scholar] [CrossRef]
- Freitas, A.; Aroso, M.; Rocha, S.; Ferreira, R.; Vitorino, R.; Gomez-Lazaro, M. Bioinformatic Analysis of the Human Brain Extracellular Matrix Proteome in Neurodegenerative Disorders. Eur. J. Neurosci. 2021, 53, 4016–4033. [Google Scholar] [CrossRef]
- Ruoslahti, E. Brain Extracellular Matrix. Glycobiology 1996, 6, 489–492. [Google Scholar] [CrossRef]
- Kerever, A.; Arikawa-Hirasawa, E. Optimal Extracellular Matrix Niches for Neurogenesis: Identifying Glycosaminoglycan Chain Composition in the Subventricular Neurogenic Zone. Front. Neuroanat. 2021, 15, 764458. [Google Scholar] [CrossRef]
- Dankovich, T.M.; Rizzoli, S.O. The Synaptic Extracellular Matrix: Long-Lived, Stable, and Still Remarkably Dynamic. Front. Synaptic Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Chaudhuri, O.; Cooper-White, J.; Janmey, P.A.; Mooney, D.J.; Shenoy, V.B. The Impact of Extracellular Matrix Viscoelasticity on Cellular Behavior. Nature 2020, 584, 535. [Google Scholar] [CrossRef]
- Bejoy, J.; Wang, Z.; Bijonowski, B.; Yang, M.; Ma, T.; Sang, Q.X.; Li, Y. Differential Effects of Heparin and Hyaluronic Acid on Neural Patterning of Human Induced Pluripotent Stem Cells. ACS Biomater. Sci. Eng. 2018, 4, 4354. [Google Scholar] [CrossRef]
- Stiles, J.; Jernigan, T.L. The Basics of Brain Development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [Green Version]
- Teffer, K.; Semendeferi, K. Chapter 9—Human Prefrontal Cortex: Evolution, Development, and Pathology. In Progress in Brain Research; Evolution of the Primate Brain; Hofman, M.A., Falk, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 195, pp. 191–218. [Google Scholar]
- Kondo, T.; Asai, M.; Tsukita, K.; Kutoku, Y.; Ohsawa, Y.; Sunada, Y.; Imamura, K.; Egawa, N.; Yahata, N.; Okita, K.; et al. Modeling Alzheimer’s Disease with IPSCs Reveals Stress Phenotypes Associated with Intracellular Aβ and Differential Drug Responsiveness. Cell Stem Cell 2013, 12, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Mattis, V.B.; Svendsen, S.P.; Ebert, A.; King, A.R.; Casale, M.; Winokur, S.T.; Batugedara, G.; Vawter, M.; Donovan, P.J.; Lock, L.F.; et al. Induced Pluripotent Stem Cells from Patients with Huntington’s Disease Show CAG-Repeat-Expansion-Associated Phenotypes. Cell Stem Cell 2012, 11, 264–278. [Google Scholar] [CrossRef] [Green Version]
- Gilmozzi, V.; Gentile, G.; Riekschnitz, D.A.; Von Troyer, M.; Lavdas, A.A.; Kerschbamer, E.; Weichenberger, C.X.; Rosato-Siri, M.D.; Casarosa, S.; Conti, L.; et al. Generation of HiPSC-Derived Functional Dopaminergic Neurons in Alginate-Based 3D Culture. Front. Cell Dev. Biol. 2021, 9, 708389. [Google Scholar] [CrossRef]
- Garcia-Campmany, L.; Stam, F.J.; Goulding, M. From Circuits to Behaviour: Motor Networks in Vertebrates. Curr. Opin. Neurobiol. 2010, 20, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Valiulahi, P.; Vidyawan, V.; Puspita, L.; Oh, Y.; Juwono, V.B.; Sittipo, P.; Friedlander, G.; Yahalomi, D.; Sohn, J.-W.; Lee, Y.K.; et al. Generation of Caudal-Type Serotonin Neurons and Hindbrain-Fate Organoids from HPSCs. Stem Cell Rep. 2021, 16, 1938–1952. [Google Scholar] [CrossRef]
- Reinhard, J.; Brösicke, N.; Theocharidis, U.; Faissner, A. The Extracellular Matrix Niche Microenvironment of Neural and Cancer Stem Cells in the Brain. Int. J. Biochem. Cell Biol. 2016, 81, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement Membrane Matrix with Biological Activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A Complex Protein Mixture Required for Optimal Growth of Cell Culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of Cerebral Organoids from Human Pluripotent Stem Cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, M.T.; Crook, C.J.; Ku, H.T. Towards Organoid Culture without Matrigel. Commun. Biol. 2021, 4, 1387. [Google Scholar] [CrossRef]
- Vukicevic, S.; Kleinman, H.K.; Luyten, F.P.; Roberts, A.B.; Roche, N.S.; Reddi, A.H. Identification of Multiple Active Growth Factors in Basement Membrane Matrigel Suggests Caution in Interpretation of Cellular Activity Related to Extracellular Matrix Components. Exp. Cell Res. 1992, 202, 1–8. [Google Scholar] [CrossRef]
- Morales, X.; Cortés-Domínguez, I.; Ortiz-de-Solorzano, C. Modeling the Mechanobiology of Cancer Cell Migration Using 3D Biomimetic Hydrogels. Gels 2021, 7, 17. [Google Scholar] [CrossRef]
- Fan, R.; Piou, M.; Darling, E.; Cormier, D.; Sun, J.; Wan, J. Bio-Printing Cell-Laden Matrigel–Agarose Constructs. J. Biomater. Appl. 2016, 31, 684–692. [Google Scholar] [CrossRef]
- De Stefano, P.; Briatico-Vangosa, F.; Bianchi, E.; Pellegata, A.F.; Hartung de Hartungen, A.; Corti, P.; Dubini, G. Bioprinting of Matrigel Scaffolds for Cancer Research. Polymers 2021, 13, 2026. [Google Scholar] [CrossRef]
- Dequach, J.A.; Yuan, S.H.; Goldstein, L.S.B.; Christman, K.L. Decellularized Porcine Brain Matrix for Cell Culture and Tissue Engineering Scaffolds. Tissue Eng. Part A 2011, 17, 2583–2592. [Google Scholar] [CrossRef]
- Medberry, C.J.; Crapo, P.M.; Siu, B.F.; Carruthers, C.A.; Wolf, M.T.; Nagarkar, S.P.; Agrawal, V.; Jones, K.E.; Kelly, J.; Johnson, S.A.; et al. Hydrogels Derived from Central Nervous System Extracellular Matrix. Biomaterials 2013, 34, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Sood, D.; Chwalek, K.; Stuntz, E.; Pouli, D.; Du, C.; Tang-Schomer, M.; Georgakoudi, I.; Black, L.D.; Kaplan, D.L. Fetal Brain Extracellular Matrix Boosts Neuronal Network Formation in 3D Bioengineered Model of Cortical Brain Tissue. ACS Biomater. Sci. Eng. 2016, 2, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Crapo, P.M.; Medberry, C.J.; Reing, J.E.; Tottey, S.; van der Merwe, Y.; Jones, K.E.; Badylak, S.F. Biologic Scaffolds Composed of Central Nervous System Extracellular Matrix. Biomaterials 2012, 33, 3539–3547. [Google Scholar] [CrossRef] [Green Version]
- Dauth, S.; Grevesse, T.; Pantazopoulos, H.; Campbell, P.H.; Maoz, B.M.; Berretta, S.; Parker, K.K. Extracellular Matrix Protein Expression Is Brain Region Dependent. J. Comp. Neurol. 2016, 524, 1309–1336. [Google Scholar] [CrossRef]
- Balgude, A.P.; Yu, X.; Szymanski, A.; Bellamkonda, R.V. Agarose Gel Stiffness Determines Rate of DRG Neurite Extension in 3D Cultures. Biomaterials 2001, 22, 1077–1084. [Google Scholar] [CrossRef]
- Lin, P.W.; Wu, C.C.; Chen, C.H.; Ho, H.O.; Chen, Y.C.; Sheu, M.T. Characterization of Cortical Neuron Outgrowth in Two- and Three-Dimensional Culture Systems. J. Biomed. Mater. Res. Part B Appl. Biomater. 2005, 75B, 146–157. [Google Scholar] [CrossRef]
- Cao, Z.; Gilbert, R.J.; He, W. Simple Agarose−Chitosan Gel Composite System for Enhanced Neuronal Growth in Three Dimensions. Biomacromolecules 2009, 10, 2954–2959. [Google Scholar] [CrossRef]
- Gu, Q.; Tomaskovic-Crook, E.; Lozano, R.; Chen, Y.; Kapsa, R.M.; Zhou, Q.; Wallace, G.G.; Crook, J.M. Functional 3D Neural Mini-Tissues from Printed Gel-Based Bioink and Human Neural Stem Cells. Adv. Healthc. Mater. 2016, 5, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Tabriz, A.G.; Hermida, M.A.; Leslie, N.R.; Shu, W. Three-Dimensional Bioprinting of Complex Cell Laden Alginate Hydrogel Structures. Biofabrication 2015, 7, 045012. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and Biomedical Applications. Prog. Polym. Sci. 2012, 37, 106. [Google Scholar] [CrossRef]
- Łabowska, M.B.; Cierluk, K.; Jankowska, A.M.; Kulbacka, J.; Detyna, J.; Michalak, I. A Review on the Adaption of Alginate-Gelatin Hydrogels for 3D Cultures and Bioprinting. Materials 2021, 14, 858. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.H.Y.; Naficy, S.; Yue, Z.; Kapsa, R.; Quigley, A.E.; Moulton, S.; Wallace, G. Bio-Ink Properties and Printability for Extrusion Printing Living Cells. Biomater. Sci. 2013, 1, 763–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozza, A.; Coates, E.E.; Incitti, T.; Ferlin, K.M.; Messina, A.; Menna, E.; Bozzi, Y.; Fisher, J.P.; Casarosa, S. Neural Differentiation of Pluripotent Cells in 3D Alginate-Based Cultures. Biomaterials 2014, 35, 4636–4645. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.K.; Goda, Y. Integrins in Synapse Regulation. Nat. Rev. Neurosci. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Pinkstaff, J.K.; Detterich, J.; Lynch, G.; Gall, C. Integrin Subunit Gene Expression Is Regionally Differentiated in Adult Brain. J. Neurosci. 1999, 19, 1541. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, L.A.; Rebaza, L.M.; Derzic, S.; Schwartz, P.H.; Monuki, E.S. Regulation of Human Neural Precursor Cells by Laminin and Integrins. J. Neurosci. Res. 2006, 83, 845. [Google Scholar] [CrossRef] [Green Version]
- Vilella, E.; Gas, C.; Garcia-Ruiz, B.; Rivera, F.J. Expression of DDR1 in the CNS and in Myelinating Oligodendrocytes. Biochim. Et Biophys. Acta (BBA)—Mol. Cell Res. 2019, 1866, 118483. [Google Scholar] [CrossRef]
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Tunable Collagen I Hydrogels for Engineered Physiological Tissue Micro-Environments. PLoS ONE 2015, 10, e0122500. [Google Scholar] [CrossRef] [Green Version]
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Review of Collagen I Hydrogels for Bioengineered Tissue Microenvironments: Characterization of Mechanics, Structure, and Transport. Tissue Eng. Part B Rev. 2014, 20, 683–696. [Google Scholar] [CrossRef]
- Ma, W.; Fitzgerald, W.; Liu, Q.Y.; O’Shaughnessy, T.J.; Maric, D.; Lin, H.J.; Alkon, D.L.; Barker, J.L. CNS Stem and Progenitor Cell Differentiation into Functional Neuronal Circuits in Three-Dimensional Collagen Gels. Exp. Neurol. 2004, 190, 276–288. [Google Scholar] [CrossRef]
- Tang-Schomer, M.D.; White, J.D.; Tien, L.W.; Schmitt, L.I.; Valentin, T.M.; Graziano, D.J.; Hopkins, A.M.; Omenetto, F.G.; Haydon, P.G.; Kaplan, D.L. Bioengineered Functional Brain-like Cortical Tissue. Proc. Natl. Acad. Sci. USA 2014, 111, 13811–13816. [Google Scholar] [CrossRef] [Green Version]
- Chwalek, K.; Tang-Schomer, M.D.; Omenetto, F.G.; Kaplan, D.L. In Vitro Bioengineered Model of Cortical Brain Tissue. Nat. Protoc. 2015, 10, 1362–1373. [Google Scholar] [CrossRef]
- Chwalek, K.; Sood, D.; Cantley, W.L.; White, J.D.; Tang-Schomer, M.; Kaplan, D.L. Engineered 3D Silk-Collagen-Based Model of Polarized Neural Tissue. J. Vis. Exp. 2015, 52970. [Google Scholar] [CrossRef] [Green Version]
- Tang-Schomer, M.D.; Wu, W.B.; Kaplan, D.L.; Bookland, M.J. In Vitro 3D Regeneration-like Growth of Human Patient Brain Tissue. J. Tissue Eng. Regen. Med. 2018, 12, 1247–1260. [Google Scholar] [CrossRef]
- Cantley, W.; Du, C.; Lomoio, S.; DePalma, T.; Peirent, E.; Kleinknecht, D.; Hunter, M.; Tang-Schomer, M.; Tesco, G.; Kaplan, D.L. Functional and Sustainable 3D Human Neural Network Models from Pluripotent Stem Cells. ACS Biomater. Sci. Eng. 2018, 4, 4278–4288. [Google Scholar] [CrossRef]
- Moxon, S.R.; Corbett, N.J.; Fisher, K.; Potjewyd, G.; Domingos, M.; Hooper, N.M. Blended Alginate/Collagen Hydrogels Promote Neurogenesis and Neuronal Maturation. Mater. Sci. Eng. C 2019, 104, 109904. [Google Scholar] [CrossRef]
- Benwood, C.; Chrenek, J.; Kirsch, R.L.; Masri, N.Z.; Richards, H.; Teetzen, K.; Willerth, S.M. Natural Biomaterials and Their Use as Bioinks for Printing Tissues. Bioengineering 2021, 8, 27. [Google Scholar] [CrossRef]
- Janmey, P.A.; Winer, J.P.; Weisel, J.W. Fibrin Gels and Their Clinical and Bioengineering Applications. J. R. Soc. Interface 2009, 6, 1–10. [Google Scholar] [CrossRef]
- Willerth, S.M.; Arendas, K.J.; Gottlieb, D.I.; Sakiyama-Elbert, S.E. Optimization of Fibrin Scaffolds for Differentiation of Murine Embryonic Stem Cells into Neural Lineage Cells. Biomaterials 2006, 27, 5990–6003. [Google Scholar] [CrossRef]
- Martino, M.M.; Briquez, P.S.; Ranga, A.; Lutolf, M.P.; Hubbell, J.A. Heparin-Binding Domain of Fibrin(Ogen) Binds Growth Factors and Promotes Tissue Repair When Incorporated within a Synthetic Matrix. Proc. Natl. Acad. Sci. USA 2013, 110, 4563–4568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, H.E.; Asker, M.E.; Kotb, N.S.; El Habab, A.M. Human Platelet Lysate Efficiency, Stability, and Optimal Heparin Concentration Required in Culture of Mammalian Cells. Blood Res. 2020, 55, 35. [Google Scholar] [CrossRef] [PubMed]
- Bento, A.R.; Quelhas, P.; Oliveira, M.J.; Pêgo, A.P.; Amaral, I.F. Three-Dimensional Culture of Single Embryonic Stem-Derived Neural/Stem Progenitor Cells in Fibrin Hydrogels: Neuronal Network Formation and Matrix Remodelling. J. Tissue Eng. Regen. Med. 2017, 11, 3494–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.; Douglas, S.; Willerth, S.M. Mechanically Stable Fibrin Scaffolds Promote Viability and Induce Neurite Outgrowth in Neural Aggregates Derived from Human Induced Pluripotent Stem Cells. Sci. Rep. 2017, 7, 6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Gregory, C.A.; Molnar, P.; Cui, X.; Jalota, S.; Bhaduri, S.B.; Boland, T. Viability and Electrophysiology of Neural Cell Structures Generated by the Inkjet Printing Method. Biomaterials 2006, 27, 3580–3588. [Google Scholar] [CrossRef]
- Schachtrup, C.; Lu, P.; Jones, L.L.; Lee, J.K.; Lu, J.; Sachs, B.D.; Zheng, B.; Akassoglou, K. Fibrinogen Inhibits Neurite Outgrowth via Β3 Integrin-Mediated Phosphorylation of the EGF Receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 11814. [Google Scholar] [CrossRef] [Green Version]
- Uibo, R.; Laidmäe, I.; Sawyer, E.S.; Flanagan, L.A.; Georges, P.C.; Winer, J.P.; Janmey, P.A. Soft Materials to Treat Central Nervous System Injuries: Evaluation of the Suitability of Non-Mammalian Fibrin Gels. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2009, 1793, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Copland, I.B.; Garcia, M.A.; Waller, E.K.; Roback, J.D.; Galipeau, J. The Effect of Platelet Lysate Fibrinogen on the Functionality of MSCs in Immunotherapy. Biomaterials 2013, 34, 7840–7850. [Google Scholar] [CrossRef]
- Pous, L.; Deshpande, S.S.; Nath, S.; Mezey, S.; Malik, S.C.; Schildge, S.; Bohrer, C.; Topp, K.; Pfeifer, D.; Fernández-Klett, F.; et al. Fibrinogen Induces Neural Stem Cell Differentiation into Astrocytes in the Subventricular Zone via BMP Signaling. Nat. Commun. 2020, 11, 630. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tao, Y.; Bradley, R.; Du, Z.; Tao, Y.; Kong, L.; Dong, Y.; Jones, J.; Yan, Y.; Harder, C.R.K.; et al. Fast Generation of Functional Subtype Astrocytes from Human Pluripotent Stem Cells. Stem Cell Rep. 2018, 11, 998–1008. [Google Scholar] [CrossRef]
- Stern, R.; Asari, A.A.; Sugahara, K.N. Hyaluronan Fragments: An Information-Rich System. Eur. J. Cell Biol. 2006, 85, 699–715. [Google Scholar] [CrossRef]
- Snetkov, P.; Zakharova, K.; Morozkina, S.; Olekhnovich, R.; Uspenskaya, M. Hyaluronic Acid: The Influence of Molecular Weight on Structural, Physical, Physico-Chemical, and Degradable Properties of Biopolymer. Polymers 2020, 12, 1800. [Google Scholar] [CrossRef]
- Fundarò, S.P.; Salti, G.; Malgapo, D.M.H.; Innocenti, S. The Rheology and Physicochemical Characteristics of Hyaluronic Acid Fillers: Their Clinical Implications. Int. J. Mol. Sci. 2022, 23, 10518. [Google Scholar] [CrossRef]
- Casini, P.; Nardi, I.; Ori, M. Hyaluronan Is Required for Cranial Neural Crest Cells Migration and Craniofacial Development. Dev. Dyn. 2012, 241, 294–302. [Google Scholar] [CrossRef]
- Margolis, R.U.; Margolis, R.K.; Chang, L.B.; Preti, C. Glycosaminoglycans of Brain during Development. Biochemistry 1975, 14, 85–88. [Google Scholar] [CrossRef]
- Preston, M.; Sherman, L.S. Neural Stem Cell Niches: Roles for the Hyaluronan-Based Extracellular Matrix. Front. Biosci. -Sch. 2011, 3, 1165–1179. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Foster, S.C.; Xing, R.; Feistel, K.; Olsen, R.H.J.; Acevedo, S.F.; Raber, J.; Sherman, L.S. CD44 Transmembrane Receptor and Hyaluronan Regulate Adult Hippocampal Neural Stem Cell Quiescence and Differentiation. J. Biol. Chem. 2017, 292, 4434–4445. [Google Scholar] [CrossRef] [Green Version]
- Vasvani, S.; Kulkarni, P.; Rawtani, D. Hyaluronic Acid: A Review on Its Biology, Aspects of Drug Delivery, Route of Administrations and a Special Emphasis on Its Approved Marketed Products and Recent Clinical Studies. Int. J. Biol. Macromol. 2020, 151, 1012–1029. [Google Scholar] [CrossRef]
- Seidlits, S.K.; Khaing, Z.Z.; Petersen, R.R.; Nickels, J.D.; Vanscoy, J.E.; Shear, J.B.; Schmidt, C.E. The Effects of Hyaluronic Acid Hydrogels with Tunable Mechanical Properties on Neural Progenitor Cell Differentiation. Biomaterials 2010, 31, 3930–3940. [Google Scholar] [CrossRef]
- Gerecht, S.; Burdick, J.A.; Ferreira, L.S.; Townsend, S.A.; Langer, R.; Vunjak-Novakovic, G. Hyaluronic Acid Hydrogel for Controlled Self-Renewal and Differentiation of Human Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2007, 104, 11298. [Google Scholar] [CrossRef]
- Wilson, E.; Knudson, W.; Newell-Litwa, K. Hyaluronan Regulates Synapse Formation and Function in Developing Neural Networks. Sci. Rep. 2020, 10, 16459. [Google Scholar] [CrossRef] [PubMed]
- Ylä-Outinen, L.; Harju, V.; Joki, T.; Koivisto, J.T.; Karvinen, J.; Kellomäki, M.; Narkilahti Ylä-Outinen, S.L.; Harju, V.; Joki, T.; Narkilahti, S.; et al. Screening of Hydrogels for Human Pluripotent Stem Cell–Derived Neural Cells: Hyaluronan-Polyvinyl Alcohol-Collagen-Based Interpenetrating Polymer Network Provides an Improved Hydrogel Scaffold. Macromol. Biosci. 2019, 19, 1900096. [Google Scholar] [CrossRef] [PubMed]
- Van Drunen, R.; Jimenez-Vergara, A.C.; Tsai, E.H.; Tchen, R.; Cagle, T.; Agee, A.B.; Roberts, J.; Steele, J.M.; Munoz-Pinto, D.J. Collagen Based Multicomponent Interpenetrating Networks as Promising Scaffolds for 3D Culture of Human Neural Stem Cells, Human Astrocytes, and Human Microglia. ACS Appl. Bio Mater. 2019, 2, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.C.; George, J.H.; Waller, S.; Besnard, C.; Nagel, D.A.; Hill, E.J.; Coleman, M.D.; Korsunsky, A.M.; Cui, Z.; Ye, H. Increased Connectivity of HiPSC-Derived Neural Networks in Multiphase Granular Hydrogel Scaffolds. Bioact. Mater. 2022, 9, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Nikolopoulou, E.; Galea, G.L.; Rolo, A.; Greene, N.D.E.; Copp, A.J. Neural Tube Closure: Cellular, Molecular and Biomechanical Mechanisms. Development 2017, 144, 552. [Google Scholar] [CrossRef] [Green Version]
- Miner, J.H.; Li, C.; Mudd, J.L.; Go, G.; Sutherland, A.E. Compositional and Structural Requirements for Laminin and Basement Membranes during Mouse Embryo Implantation and Gastrulation. Development 2004, 131, 2247–2256. [Google Scholar] [CrossRef] [Green Version]
- Colognato, H.; Ffrench-Constant, C.; Feltri, M.L. Human Diseases Reveal Novel Roles for Neural Laminins. Trends Neurosci. 2005, 28, 480–486. [Google Scholar] [CrossRef]
- Cloutier, G.; Sallenbach-Morrissette, A.; Beaulieu, J.F. Non-Integrin Laminin Receptors in Epithelia. Tissue Cell 2019, 56, 71–78. [Google Scholar] [CrossRef]
- Lindenmaier, L.B.; Parmentier, N.; Guo, C.; Tissir, F.; Wright, K.M. Dystroglycan Is a Scaffold for Extracellular Axon Guidance Decisions. eLife 2019, 8, e42143. [Google Scholar] [CrossRef]
- Nickolls, A.R.; Bönnemann, C.G. The Roles of Dystroglycan in the Nervous System: Insights from Animal Models of Muscular Dystrophy. DMM Dis. Model. Mech. 2018, 11, dmm035931. [Google Scholar] [CrossRef]
- Hall, P.E.; Lathia, J.D.; Caldwell, M.A.; Ffrench-Constant, C. Laminin Enhances the Growth of Human Neural Stem Cells in Defined Culture Media. BMC Neurosci. 2008, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Pavathuparambil Abdul Manaph, N.; Al-Hawwas, M.; Bobrovskaya, L.; Xiong, L.L.; Zhou, X.F. Coating Materials for Neural Stem/Progenitor Cell Culture and Differentiation. Stem Cells Dev. 2020, 29, 463–474. [Google Scholar] [CrossRef]
- Arulmoli, J.; Wright, H.J.; Phan, D.T.T.; Sheth, U.; Que, R.A.; Botten, G.A.; Keating, M.; Botvinick, E.L.; Pathak, M.M.; Zarembinski, T.I.; et al. Combination Scaffolds of Salmon Fibrin, Hyaluronic Acid, and Laminin for Human Neural Stem Cell and Vascular Tissue Engineering. Acta Biomater. 2016, 43, 122. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.D.; Pandanaboina, S.C.; Srivatsan, M.; Xie, J.Y. Predominant Differentiation of Rat Fetal Neural Stem Cells into Functional Oligodendrocytes in Vitro. Neurosci. Lett. 2020, 736, 135264. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Josey, B.; Chou, J.; Tan, Y.; Zhang, N.; Wen, X.; Chou, C.J.; Tan, Y.U. Short Laminin Peptide for Improved Neural Stem Cell Growth. Stem Cells Transl. Med. 2014, 3, 662–670. [Google Scholar] [CrossRef]
- Perris, R.; Perissinotto, D. Role of the Extracellular Matrix during Neural Crest Cell Migration. Mech. Dev. 2000, 95, 3–21. [Google Scholar] [CrossRef]
- Gnavi, S.; Barwig, C.; Freier, T.; Haastert-Talini, K.; Grothe, C.; Geuna, S. The Use of Chitosan-Based Scaffolds to Enhance Regeneration in the Nervous System. Int. Rev. Neurobiol. 2013, 109, 1–62. [Google Scholar] [CrossRef]
- Li, Y.; Ju, D. Chapter 12—The Application, Neurotoxicity, and Related Mechanism of Cationic Polymers. In Neurotoxicity of Nanomaterials and Nanomedicine; Academic press: Cambridge, MA, USA, 2017; pp. 285–329. ISBN 9780128045985. [Google Scholar]
- Ojeda-Hernández, D.D.; Canales-Aguirre, A.A.; Matias-Guiu, J.; Gomez-Pinedo, U.; Mateos-Díaz, J.C. Potential of Chitosan and Its Derivatives for Biomedical Applications in the Central Nervous System. Front. Bioeng. Biotechnol. 2020, 8, 389. [Google Scholar] [CrossRef]
- Wu, Q.; Therriault, D.; Heuzey, M.-C. Processing and Properties of Chitosan Inks for 3D Printing of Hydrogel Microstructures. ACS Biomater. Sci. Eng. 2018, 4, 2643–2652. [Google Scholar] [CrossRef]
- Faissner, A.; Roll, L.; Theocharidis, U. Tenascin-C in the Matrisome of Neural Stem and Progenitor Cells. Mol. Cell. Neurosci. 2017, 81, 22–31. [Google Scholar] [CrossRef]
- Shahi, M.; Mohammadnejad, D.; Karimipour, M.; Rahbarghazi, R.; Abedelahi, A. Evaluation of the Effect of Hyaluronic Acid–Based Biomaterial Enriched With Tenascin-C on the Behavior of the Neural Stem Cells. Int. J. Toxicol. 2021, 40, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Naghdi, P.; Tiraihi, T.; Ganji, F.; Darabi, S.; Taheri, T.; Kazemi, H. Survival, Proliferation and Differentiation Enhancement of Neural Stem Cells Cultured in Three-Dimensional Polyethylene Glycol–RGD Hydrogel with Tenascin. J. Tissue Eng. Regen. Med. 2016, 10, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Yu, A.; Chen, J.; Yuan, J.; Yin, Y.; Duanmu, W.; Tan, L.; Yang, Y.; Lan, C.; Chen, W.; et al. Poly-L-Ornithine Enhances Migration of Neural Stem/Progenitor Cells via Promoting α-Actinin 4 Binding to Actin Filaments. Sci. Rep. 2016, 6, 37681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiya, Y.; Yamaki, T.; Uchida, M.; Hatanaka, T.; Kimura, M.; Ogihara, M.; Morimoto, Y.; Natsume, H. Preparation and Evaluation of PEGylated Poly-L-Ornithine Complex as a Novel Absorption Enhancer. Biol. Pharm. Bull. 2017, 40, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Guy, B.; Zhang, J.S.; Duncan, L.H.; Johnston, R.J. Human Neural Organoids: Models for Developmental Neurobiology and Disease. Dev. Biol. 2021, 478, 102–121. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Layrolle, P.; Payoux, P.; Chavanas, S. Message in a Scaffold: Natural Biomaterials for Three-Dimensional (3D) Bioprinting of Human Brain Organoids. Biomolecules 2023, 13, 25. https://doi.org/10.3390/biom13010025
Layrolle P, Payoux P, Chavanas S. Message in a Scaffold: Natural Biomaterials for Three-Dimensional (3D) Bioprinting of Human Brain Organoids. Biomolecules. 2023; 13(1):25. https://doi.org/10.3390/biom13010025
Chicago/Turabian StyleLayrolle, Pierre, Pierre Payoux, and Stéphane Chavanas. 2023. "Message in a Scaffold: Natural Biomaterials for Three-Dimensional (3D) Bioprinting of Human Brain Organoids" Biomolecules 13, no. 1: 25. https://doi.org/10.3390/biom13010025