
MTHFR SNPs (Methyl Tetrahydrofolate Reductase, Single Nucleotide Polymorphisms) C677T and A1298C Prevalence and Serum Homocysteine Levels in >2100 Hypofertile Caucasian Male Patients

 , ,
, ,
Abstract
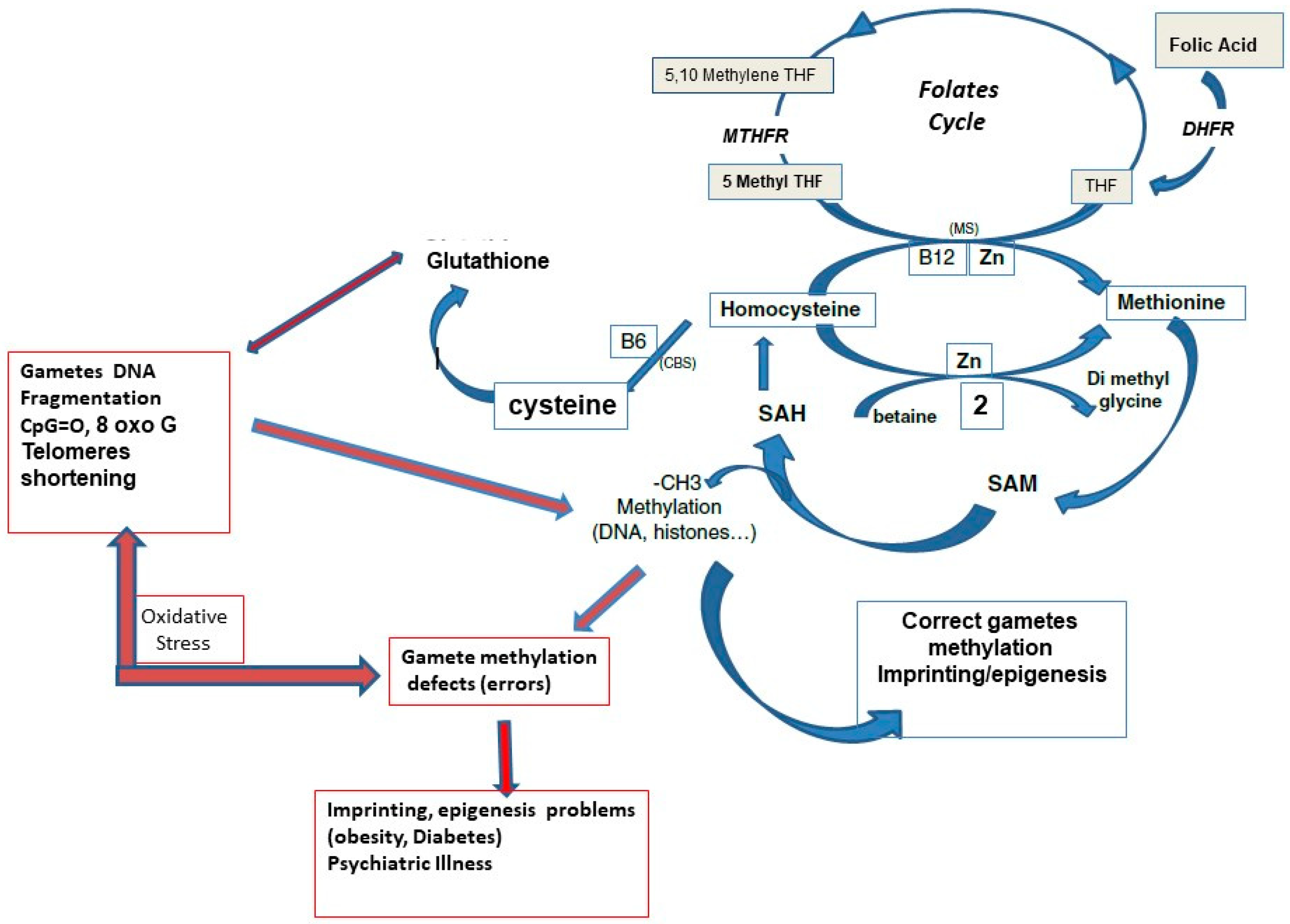
:1. Introduction
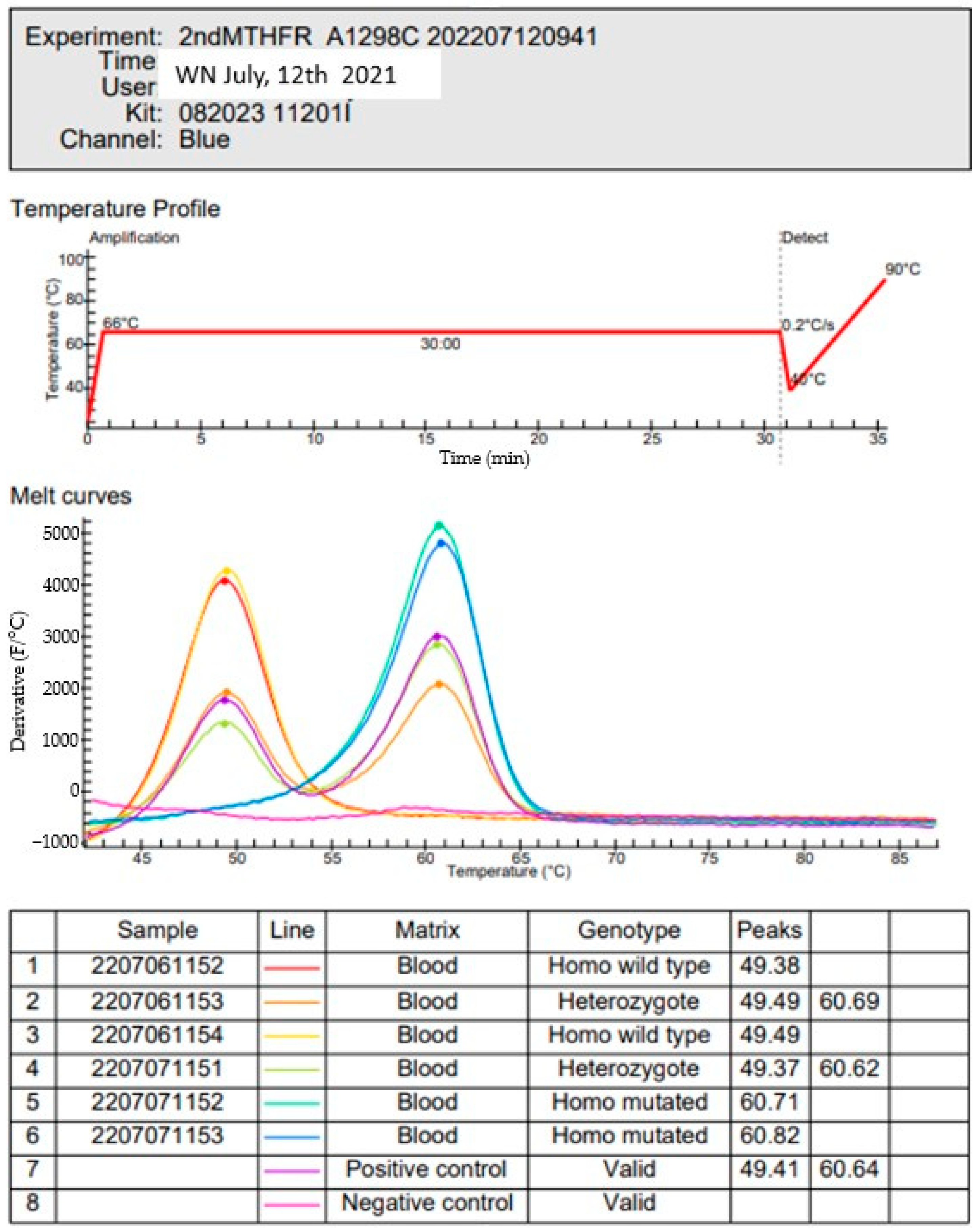
2. Materials and Methods
Population and Ethical Considerations
3. Results
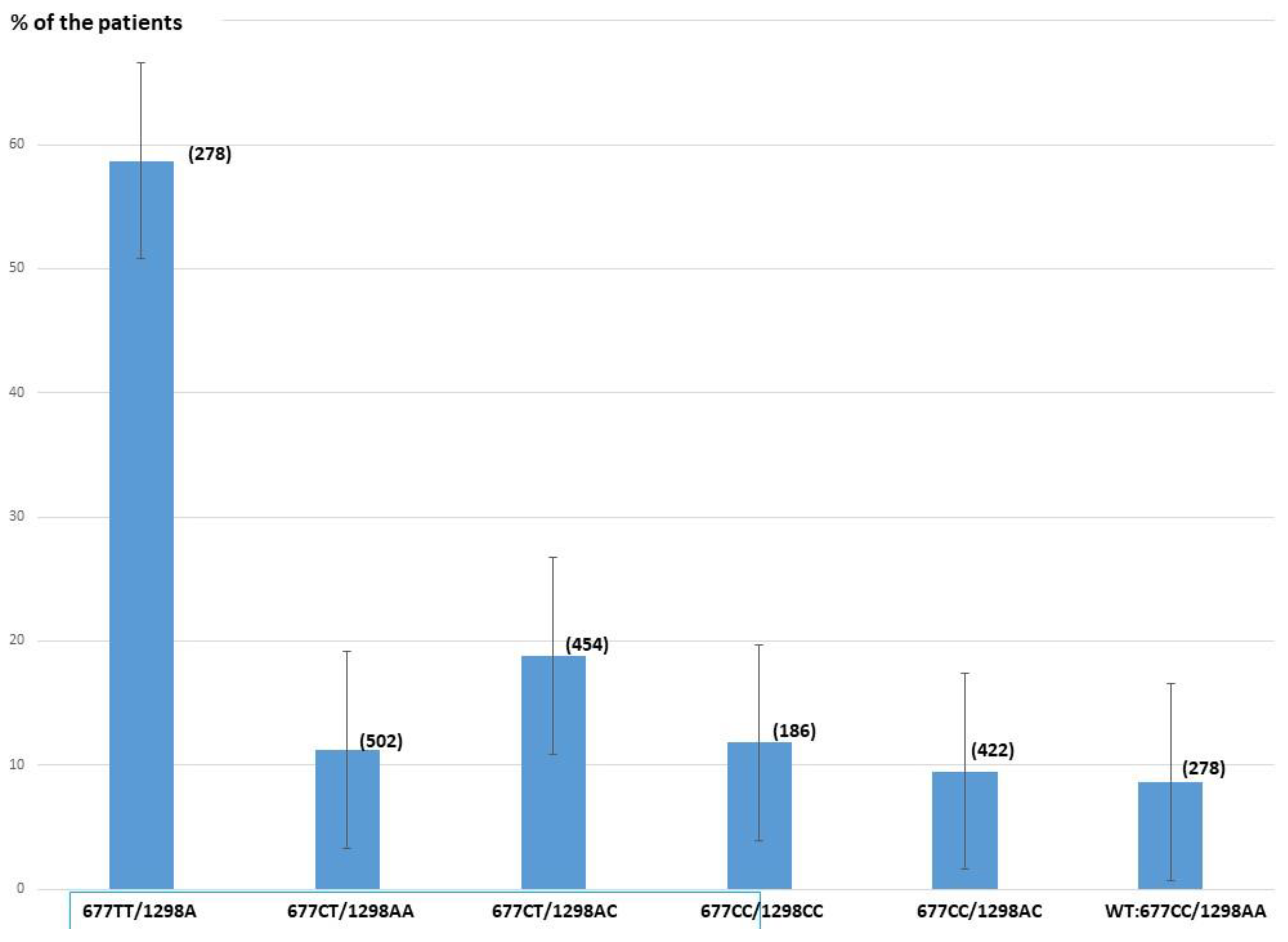
3.1. Distribution of the Two SNP Combinations
3.2. Impact of the SNP Combinations on Hcy Levels >15 µMolar
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Monk, M. Genomic imprinting. Genes Dev. 1988, 2, 921–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, M. Memories of mother and father. Nature 1987, 328, 203–204. [Google Scholar] [CrossRef] [PubMed]
- Menezo, Y.; Clement, P.; Clement, A.; Elder, K. Methylation: An Ineluctable Biochemical and Physiological Process Essential to the Transmission of Life. Int. J. Mol. Sci. 2020, 21, 9311. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, P.; Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Hine, R.J.; James, S.J. Increase in plasma homocysteine associated with parallel increases in plasma S-adenosylhomocysteine and lymphocyte DNA hypomethylation. J. Biol. Chem. 2000, 275, 29318–29323. [Google Scholar] [CrossRef] [Green Version]
- van der Put, N.M.; Gabreëls, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Blom, H.J. A second common mutation in the methyl-enetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Zappacosta, B.; Graziano, M.; Persichilli, S.; Di Castelnuovo, A.; Mastroiacovo, P.; Iacoviello, L. 5,10-Methylenetetrahydrofolate reductase (MTHFR) C677T and A1298C polymorphisms: Genotype frequency and association with homocysteine and folate levels in middle-southern Italian adults. Cell Biochem. Funct. 2014, 32, 1–4. [Google Scholar] [CrossRef]
- Hoffman, M. Hypothesis: Hyperhomocysteinemia is an indicator of oxidant stress. Med. Hypothes. 2011, 77, 1088–1093. [Google Scholar] [CrossRef]
- Tunc, O.; Tremellen, K. Oxidative DNA damage impairs global sperm DNA methylation in infertile men. J. Assist. Reprod. Genet. 2009, 26, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Menezo, Y.J.; Silvestris, E.; Dale, B.; Elder, K. Oxidative stress and alterations in DNA methylation: Two sides of the same coin in reproduction. Reprod. BioMed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef] [Green Version]
- Duthie, S.J.; Narayanan, S.; Brand, G.M.; Pirie, L.; Grant, G. Impact of Folate Deficiency on DNA Stability. J. Nutr. 2002, 132, 2444S–2449S. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Flanagan, H.M.; Connaughton, H.; Whiting, S.; Hedges, A.; Baker, M.A. Involvement of homocysteine, homocysteine thiolactone, and paraoxonase type 1 (PON-1) in the etiology of defective human sperm function. Andrology 2016, 4, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarabi, M.; San Gabriel, M.C.; Chan, D.; Behan, N.A.; Caron, M.; Pastinen, T.; Bourque, G.; MacFarlane, A.J.; Zini, A.; Trasler, J. High-dose folic acid supplementation alters the human sperm methylome and is influenced by the MTHFR C677T polymorphism. J. Hum. Mol. Genet. 2015, 24, 6301–6313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarabi, M.; Christensen, K.E.; Chan, D.; Leclerc, D.; Landry, M.; Ly, L.; Rozen, R.; Trasler, J. Testicular MTHFR deficiency may ex-plain sperm DNA hypomethylation associated with high dose folic acid supplementation. J. Hum. Mol. Genet. 2018, 27, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Luo, Y.; Wu, S.; Tang, Y.; Rao, X.; Xiong, L.; Tan, M.; Deng, M.; Liu, H. Association between C677T and A1298C polymorphisms of the MTHFR gene and risk of male infertility: A meta-analysis. Genet. Mol. Res. 2016, 15, grm.15027631. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luo, Y.; Yuan, J.; Tang, Y.; Xiong, L.; Xu, M.; Rao, X.; Liu, H. Association between maternal, fetal and paternal MTHFR gene C677T and A1298C polymorphisms and risk of recurrent pregnancy loss: A comprehensive evaluation. Arch. Gynecol. Obstet. 2016, 293, 1197–1211. [Google Scholar] [CrossRef]
- Gong, M.; Dong, W.; He, T.; Shi, Z.; Huang, G. MTHFR 677C>T polymorphism increases the male infertility risk: A meta-analysis involving 26 studies. PLoS ONE 2015, 10, e0121147. [Google Scholar] [CrossRef] [PubMed]
- Jacquesson-Fournols, L.; Alvarez, S.; Cohen, M.; Clement, P.; Menezo, Y. A paternal effect of MTHFR SNPs on gametes and embryos should not be overlooked: Case reports. J. Assist. Reprod. Genet. 2019, 36, 1351–1353. [Google Scholar] [CrossRef] [Green Version]
- Han, L.J.; He, X.F.; Ye, X.H. Methylenetetrahydrofolate reductase C677T and A1298C polymorphisms and male infertility risk: An updated meta-analysis. Medicine 2020, 99, e23662. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, C.; Shi, Q.; Zhu, Y.; Liu, Y. Hyperhomocysteinemia in men with a reproductive history of fetal neural tube defects Three case reports and literature review. Medicine 2019, 98, e13998. [Google Scholar] [CrossRef]
- Enciso, M.; Sarasa, J.; Xanthopoulou, L.; Bristow, S.; Bowles, M.; Fragouli, E.; Delhanty, J.; Wells, D. Polymorphisms in the MTHFR gene influence embryo viability and the incidence of aneuploidy. Hum. Genet. 2016, 135, 555–568. [Google Scholar] [CrossRef] [PubMed]
- D’Elia, P.Q.; dos Santos, A.A.; Bianco, B.; Barbosa, C.; Christofolini, D.M.; Aoki, T. MTHFR polymorphisms C677T and A1298C and associations with IVF outcomes in Brazilian women. Reprod. BioMed. Online 2014, 28, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Szymański, W.; Kazdepka-Ziemińska, A. Effect of homocysteine concentration in follicular fluid on a degree of oocyte maturity. Ginekol. Pol. 2003, 74, 1392–1396. [Google Scholar] [PubMed]
- Berker, B.; Kaya, C.; Aytac, R.; Satıroglu, H. Homocysteine concentrations in follicular fluid are associated with poor oocyte and embryo qualities in polycystic ovary syndrome patients undergoing assisted reproduction. Hum. Reprod. 2009, 24, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocal, P.; Ersoylu, B.; Cepni, I.; Guralp, O.; Atakul, N.; Irez, T.; Idil, M. The association between homocysteine in the follicular fluid with embryo quality and pregnancy rate in assisted reproductive techniques. J. Assist. Reprod. Genet. 2012, 29, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, C.A.; Sherman, S.L.; Yi, P.; Hopkins, S.E.; Torfs, C.P.; Hine, R.J.; Pogribna, M.; Rozen, R.; James, S.J. Polymorphisms in genes involved in folate metabolism as maternal risk factors for Down syndrome. Am. J. Hum. Genet. 2000, 67, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Fei, Y.; Li, J.; Shi, Y.; Yang, X. Novel Review of Homocysteine and Pregnancy Complications. BioMed Res. Int. 2021, 2021, 6652231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fu, H.; Wei, T. MTHFR gene polymorphism and homocysteine levels in spontaneous abortion of pregnant women. Am. J. Transl. Res. 2021, 13, 7083–7088. [Google Scholar]
- Kutchy, N.A.; Menezes, E.S.; Ugur, M.R.; Husna, A.U.; ElDebaky, H.; Evans, H.C.; Beaty, E.; Santos, F.C.; Tan, W.; Wills, R.W.; et al. Sperm cellular and nuclear dynamics associated with bull fertility. Anim. Reprod. Sci. 2019, 211, 106203. [Google Scholar] [CrossRef]
- Karahan, G.; Chan, D.; Shirane, K.; McClatchie, T.; Janssen, S.; Baltz, J.M.; Lorincz, M.; Trasler, J. Paternal MTHFR deficiency leads to hypomethylation of young retrotransposons and reproductive decline across two successive generations. Development 2021, 148, dev199492. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Lanzillotti, C.; Mazziotta, C.; Tognon, M.; Martini, F. Epigenetics of Male Infertility: The Role of DNA Methylation. Front. Cell Dev. Biol. 2021, 9, 689624. [Google Scholar] [CrossRef] [PubMed]
- Clément, A.; Menezo, Y.; Cohen, M.; Cornet, D.; Clément, P. 5-Methyltetrahydrofolate reduces blood homocysteine level significantly in C677T methyltetrahydrofolate reductase single-nucleotide polymorphism carriers consulting for infertility. J. Gynecol. Obstet. Hum. Reprod. 2020, 49, 101622. [Google Scholar] [CrossRef] [PubMed]
- Clément, A.; Chouteau, J.; Clément, P.; Ménézo, Y. Importance of the determination of MTHFR SNPs (Methylene Tetrahydrofolate Reductase Single Nucleotide Polymorphisms) in couple infertility. Gynecol. Obstet. Fertil. Senol. 2020, 48, 422–427. [Google Scholar] [PubMed]
- Leclerc, D.; Rozen, R. Molecular genetics of MTHFR: Polymorphisms are not all benign. Med. Sci. 2007, 23, 297–302. [Google Scholar]
- Škovierová, H.; Vidomanová, E.; Mahmood, S.; Sopková, J.; Drgová, A.; Červeňová, T.; Halašová, E.; Lehotský, J. The Molecular and Cellular Effect of Homocysteine Metabolism Imbalance on Human Health. Int. J. Mol. Sci. 2016, 17, 1733. [Google Scholar] [CrossRef] [Green Version]
- Jamil, K. Clinical Implications of MTHFR Gene Polymorphism in Various Diseases. Biol. Med. 2014, 6, e107. [Google Scholar]
- Servy, E.J.; Jacquesson-Fournols, L.; Cohen, M.; Menezo, Y. MTHFR isoform carriers. 5-MTHF (5-methyl tetrahydro-folate) vs. folic acid: A key to pregnancy outcome: A case series. J. Assist. Reprod. Genet. 2018, 35, 1431–1435. [Google Scholar] [CrossRef]
- Tara, S.-S.; Ghaemimanesh, F.; Zarei, S.; Reihani-Sabet, F.; Pahlevanzadeh, Z.; Modarresi, M.H.; Jeddi-Tehrani, M. Methylenetetrahydrofolate Reductase C677T and A1298C Polymorphisms in Male Partners of Recurrent Miscarriage Couples. J. Reprod. Infertil. 2015, 16, 193–198. [Google Scholar]
- Mtiraoui, N.; Zammiti, W.; Ghazouani, L.; Braham, N.J.; Saidi, S.; Finan, R.R.; Almawi, W.Y.; Mahjoub, T. Methylenetetrahydrofolate reductase C677T and A1298C polymorphism and changes in homocysteine concentrations in women with idiopathic recurrent pregnancy losses. Reproduction 2006, 131, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Hong, X.; Wang, J.; Jiang, Y.; Zhang, Y.; Chen, J.; Niu, X. Estradiol-17β inhibits homocysteine mediated damage by promoting H2S production via upregulating CBS and CSE expression in human umbilical vein endothelial cells. J. Cell. Biochem. 2019, 122, 915–925. [Google Scholar] [CrossRef]
- Rubini, E.; Snoek, K.M.; Schoenmakers, S.; Willemsen, S.P.; Sinclair, K.D.; Rousian, M.; Steegers-Theunissen, R.P. First Trimester Maternal Homocysteine and Embryonic and Fetal Growth: The Rotterdam Periconception Cohort. Nutrients 2022, 14, 1129. [Google Scholar] [CrossRef] [PubMed]
- Menezo, Y.; Elder, K.; Clement, A.; Clement, P. Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules 2022, 12, 197. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.W.; Ayling, J.E. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc. Natl. Acad. Sci. USA 2009, 106, 15424–15429. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| SNP Combination | Number (% of the Population) | Number, (%) of the Patients w Hcy >15 µM) |
|---|---|---|
| C677C/A1298A (WT) | 278(13.1%) | 24 (8.6%) |
| T677T/A1298A | 278(13.1%) | 159 (57.8%) |
| C677T/A1298A | 502(23.5%) | 56 (11.2%) |
| C677T/A1298C | 454 (21.4%) | 86 (18.8%) |
| C677C/C1298C | 186(8.7%) | 22 (11.8%) |
| C677C/A1298C | 422(19.7%) | 40 (9.5%) |
| C677T/C1298C | 4(0.2%) | - |
| T667T/A1298C | 3 (0.2%) | 1 |
| Total | 2127 (100%) | 388 (18.2%) |
| Odds Ratio | Lower 95% | Upper 95% | |
|---|---|---|---|
| C677T/A1298C | 2.47 | 1.5 | 4 |
| T677T/A1298A | 14.14 | 8.7 | 22.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clément, A.; Amar, E.; Brami, C.; Clément, P.; Alvarez, S.; Jacquesson-Fournols, L.; Davy, C.; Lalau-Keraly, M.; Menezo, Y. MTHFR SNPs (Methyl Tetrahydrofolate Reductase, Single Nucleotide Polymorphisms) C677T and A1298C Prevalence and Serum Homocysteine Levels in >2100 Hypofertile Caucasian Male Patients. Biomolecules 2022, 12, 1086. https://doi.org/10.3390/biom12081086
Clément A, Amar E, Brami C, Clément P, Alvarez S, Jacquesson-Fournols L, Davy C, Lalau-Keraly M, Menezo Y. MTHFR SNPs (Methyl Tetrahydrofolate Reductase, Single Nucleotide Polymorphisms) C677T and A1298C Prevalence and Serum Homocysteine Levels in >2100 Hypofertile Caucasian Male Patients. Biomolecules. 2022; 12(8):1086. https://doi.org/10.3390/biom12081086
Chicago/Turabian StyleClément, Arthur, Edouard Amar, Charles Brami, Patrice Clément, Silvia Alvarez, Laetitia Jacquesson-Fournols, Céline Davy, Marc Lalau-Keraly, and Yves Menezo. 2022. "MTHFR SNPs (Methyl Tetrahydrofolate Reductase, Single Nucleotide Polymorphisms) C677T and A1298C Prevalence and Serum Homocysteine Levels in >2100 Hypofertile Caucasian Male Patients" Biomolecules 12, no. 8: 1086. https://doi.org/10.3390/biom12081086