
Polymorphisms in Genes Involved in Inflammation and Periodontitis: A Narrative Review


Abstract
:1. Introduction
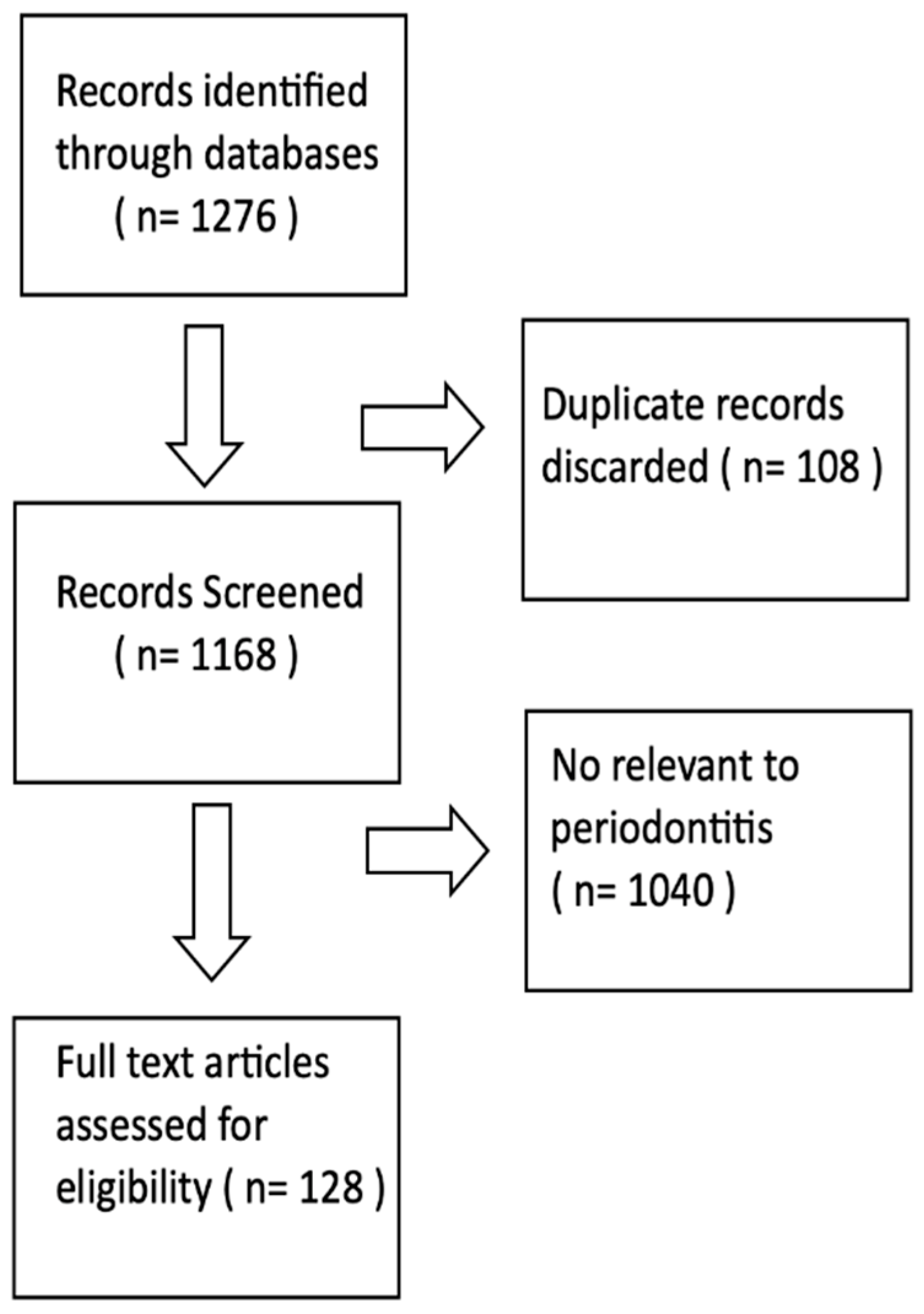
2. Methodology
3. Results
3.1. Human Leukocyte Antigens (HLA)
3.2. Antibodies (Ig)
3.3. Receptor for Antibody Fc Fragment (FcR)
3.4. CD14 Molecules and Toll-like Receptors
3.5. Vitamin D Receptor (VDR)
3.6. Other Cell Receptors
3.7. Matrix Metalloproteinases (MMPs)
3.8. Other Enzymes
3.9. Cytokines
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shaddox, L.M.; Morford, L.A.; Nibali, L. Periodontal health and disease: The contribution of genetics. Periodontology 2000 2021, 85, 161–181. [Google Scholar] [CrossRef] [PubMed]
- Yoshie, H.; Kobayashi, T.; Tai, H.; Galicia, J.C. The role of genetic polymorphisms in periodontitis. Periodontology 2000 2007, 43, 102–132. [Google Scholar] [CrossRef] [PubMed]
- Armitage, G.C. Development of a classification system for periodontal diseases and conditions. Ann. Periodontol. 1999, 22, 1–6. [Google Scholar] [CrossRef]
- Fine, D.H.; Patil, A.G.; Loos, B.G. Classification and diagnosis of aggressive periodontitis. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S95–S111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonetti, M.S.; Greenwell, H.; Kornman, K.S. Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S149–S161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterer, G.W.; Wunderink, R.G. Science review: Genetic variability in the systemic inflammatory response. Crit. Care 2003, 7, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Hollegaard, M.V.; Bidwell, J.L. Cytokine gene polymorphism in human disease: On-line databases. Genes Immun. 2006, 7 (Suppl. 3), 269–276. [Google Scholar] [CrossRef]
- Lafuente-Ibáñez de Mendoza, I.; Setien-Olarra, A.; García-De la Fuente, A.M.; Aguirre-Urizar, J.M.; Marichalar-Mendia, X. Role of proinflammatory mutations in peri-implantitis: Systematic review and meta-analysis. Int. J. Implant Dent. 2022, 8, 2. [Google Scholar] [CrossRef]
- Mattuella, L.G.; Bernardi, L.; Zambra, F.M.B.; Campagnaro, M.B.; Oppermann, R.V.; Xavier, L.L.; Chies, J.A.B.; Miranda, L.A. Human leukocyte antigen-G polymorphisms in periodontitis. Acta Odontol. Scand. 2020, 78, 141–145. [Google Scholar] [CrossRef]
- Takashiba, S.; Ohyama, H.; Oyaizu, K.; Kogoe-Kato, N.; Murayama, Y. HLA genetics fofr diagnosis of susceptiobility to early-onset periodontitis. J. Periodont. Res. 1999, 34, 374–378. [Google Scholar] [CrossRef]
- Kulshrestha, R.; Biswas, J. Role of Immunoglobulin G anadA in Periodontitis: A Review. J. Pure Appl. Microbiol. 2013, 70, 673–676. [Google Scholar]
- Hans, V.M.; Mehta, D.S.; Hans, M. Association of Fc gamma-receptors IIa, IIIa and IIIb genetic polymorphism with susceptibility to chronic periodontitis in South Indian population. Contep. Clin. Dent. 2015, 6 (Suppl. 1), 141–146. [Google Scholar]
- Dimou, N.L.; Nikolopoulos, G.; Hamodrakas, S.; Bagos, P. Fc gamma receptor polymorphism and their association with periodontal disease: A meta-analysis. J. Clin. Periodontol. 2010, 37, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Sakunrangsit, N.; Pholtaisong, J.; Sucharitakul, J.; Wanna-Udom, S.; Prombutara, P.; Pisitkun, P.; Leelahavanichkul, A.; Aporntewan, C.; Greenblatt, M.B.; Lotinun, S. Identification of candidate regulators of mandibular bone loss in FcγRIIB-/- Mice. Sci. Rep. 2021, 21, 18726. [Google Scholar] [CrossRef] [PubMed]
- Donati, M.; Berglundh, T.; Hytonen, A.M.; Hahn-Zoric, M.; Hanson, L.A.; Padyukov, L. Association of the −159 CD14 gene polymorphism and lack of association of the −308 TNFA and Q551R IL-4RA polymorphisms with severe chronic periodontitis in Swedish Caucasians. J. Clin. Periodontol. 2005, 32, 474–479. [Google Scholar] [CrossRef]
- Zacarias, J.; Bazzo de Alencar, J.; Yumeko Tsuneto, P.; Hugo de Souza, V. The influence of TLR4, CD14, OPG and RANKL Polymorphism in Periodontitis: A case control study. Mediat. Inflamm. 2019, 2019, 4029217. [Google Scholar] [CrossRef]
- Song, B.; Zhang, Y.L.; Chen, L.J.; Zhou, T.; Huang, W.K.; Zhou, X.; Shao, L.Q. The role of Toll-like receptors in periodontitis. Oral Dis. 2017, 23, 168–180. [Google Scholar] [CrossRef]
- Murthykumar, K.; Arjunkumar, R.; Jayaseelan, V.P. Association of vitamin D receptor gene polymorphism (rs 10735810) and chronic periodontitis. J. Investig. Clin. Dent. 2019, 10, e12440. [Google Scholar] [CrossRef]
- Skvortsov, S.S.; Gabdoulkhakova, A.G. Formyl peptide receptor polymorphism: 27 most possible ways for phagocyte dysfunction. Biochemistry 2017, 82, 426–437. [Google Scholar] [CrossRef]
- Holla, L.I.; Kankova, K.; Fassmann, A.; Buckova, D.; Halabala, T.; Znojil, V.; Vanek, J. Distribution of the receptor for advanced glycation and products gene polymorphisms in patients with chronic periodontitis: A preliminary study. J. Periodontol. 2001, 72, 1742–1746. [Google Scholar] [CrossRef]
- Slotwinska, S.M. The Interleukin-1 receptor antagonist (IL-1-Ra) and soluble tumor necrosis factor receptor I (sTNF RI) in periodontal disease. Open J. Immunol. 2013, 3, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhu, Y.; Singh, P.; Ajmera, D.H.; Song, J.; Ji, P. Association of Common Variants in MMPs with Periodontitis Risk. Dis. Markers 2016, 2016, 1545974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holla, L.I.; Fassmann, A.; Vasku, A.; Znojil, V.; Vanek, J.; Vacha, J. Interactions of lymphotoxin alpha (TNF-beta), angiotensin-converting enzyme (ACE), and endothelin-1 (ET-1) gene polymorphisms in adult periodontitis. J. Periodontol. 2001, 72, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Reichert, S.; Altermann, W.; Stein, J.M.; Schaller, H.G.; Machulla, H.K.G.; Schulz, S. Individual Composition of HLA and Periodontopathogens in the Background of Periodontitis. J. Periodontol. 2012, 84, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Bonfil, J.J.; Dillier, F.L.; Mercier, P.; Reviron, D.; Foti, B.; Sambuc, R.; Brodeur, J.M.; Sedarat, C. A “case control” study on the role of HLA DR4 in severe periodontitis and rapidly progressive periodontoitis. Identification of types and subtypes using molecular biology (PCR.SSO). J. Clin. Periodontol. 1999, 26, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, H.; Takashiba, S.; Oyaizu, K.; Nagai, A.; Narusz, T.; Ikono, H.; Kurihara, H.; Murayama, Y. HLA class II genotype associated with early-onset periodontitis: DQB1 molecule primarily confers susceptibility to the disease. J. Periodontol. 1996, 67, 888–894. [Google Scholar] [CrossRef]
- Stein, J.M.; Machulla, H.K.G.; Smeets, R.; Lampert, F.; Reichert, S. Human leukocyte antigen polymorphism in chronic and aggressive periodontitis among Caucasians: A meta-analysis. J. Clin. Periodontol. 2008, 35, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Megha, K.B.; Mohanan, P.V. Role of immunoglobulin and antibodies in disease management. Int. J. Biol. Macromol. 2021, 169, 28–38. [Google Scholar] [CrossRef]
- Page, R.C.; Offenbacher, S.; Schroeder, H.E.; Seymour, G.J.; Kornman, K.S. Advances in the pathogenesis of periodontitis: Summary of developments, clinical implications and future directions. Periodontology 2000 1997, 14, 216–248. [Google Scholar] [CrossRef]
- Saraiva, L.; Rebeis, E.S.; Martins Ede, S.; Sekiguchi, R.T.; Ando-Suguimoto, E.S.; Mafra, C.E.S.; Holzhausen, M.; Romito, G.A.; Mayer, M.P.A. IgG Sera Levels Against a Subset of Periodontopathogens and Severity of Disease in Aggressive Periodontitis Patients: A Cross-Sectional Study of Selected Pocket Sites. J. Clin. Periodontol. 2014, 41, 943–951. [Google Scholar] [CrossRef]
- Hwang, M.A.; Stoupel, J.; Celenti, R.; Demmer, R.T.; Papapanou, P.N. Serum Antibody Responses to Periodontal Microbiota in Chronic and Aggessive Periodontitis. A Postulate Revised. J. Periodontol. 2014, 85, 592–600. [Google Scholar] [CrossRef]
- Chai, L.; Song, Y.Q.; Leung, W.K. Genetic polymorphism studies in periodontitis and Fcy receptors. J. Periodontal. Res. 2012, 47, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, B.G.; Leppers-Van de Styraat, F.G.J.; Van de Winkel, J.G.J.; Van der Velden, U. Fcgamma Receptor Polymorphisms in Relation to Periodontitis. J. Clin. Periodontol. 2003, 30, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Ito, S.; Yasuda, K.; Kuroda, T.; Yamamoto, K.; Sugita, N.; Tai, H.; Narita, I.; Gejyo, F.; Yoshie, H. The combined genotypes of stimulatory and inhibitory Fc gamma receptors associated with systemic lupus erythematosus and periodontitis in Japanese adults. J. Periodontol. 2007, 78, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Yamamoto, K.; Sugita, N.; van der Pol, W.L.; Yasuda, K.; Kaneko, S.; van de Winkel, J.G.J.; Yoshie, H. The Fcy receptor genotype as a severity factor for chronic periodontitis in Japanese patients. J. Periodontol. 2001, 72, 1324–1331. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sugita, N.; van der Pol, W.L.; Nunokawa, Y.; Westerdaal, N.A.C.; Yamamoto, K.; van de Winkel, J.G.J.; Yoshie, H. The Fcy receptor genotype as a risk factor for generalized early-onset periodontitis in Japanese patients. J. Periodontol. 2000, 71, 1425–1432. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kobayashi, T.; Grossi, S.; Ho, A.W.; Genco, R.J.; Yoshie, H.; De Nardin, E. Association of Fcy receptor IIa genotype with chronic periodontitis in Caucasians. J. Periodontol. 2004, 75, 517–552. [Google Scholar] [CrossRef]
- Saremi, L.; Esmaeilzadeh, E.; Ghorashi, T.; Sohrtabi, M.; Kermani, M.E.; Kadkhofazadeh, M. Association of Fc gamma-receptor genes polymorphisms with chronic periodontitis and Peri-Implantitis. J. Cell Biochem. 2019, 120, 12010–12017. [Google Scholar] [CrossRef]
- Kobayashi, T.; van der Pol, W.L.; van de Winkel, J.G.J.; Hara, K.; Sugita, N.; Westerdaal, N.A.C.; Yoshie, H.; Horigome, T. Relevance of IgG receptor III (CD16) polymorphism to handling of Porphyromonas gingivalis: Implications for the pathogenesis of adult periodontitis. J. Periodont. Res. 2000, 35, 65–73. [Google Scholar] [CrossRef]
- Yoshihara, A.; Sugita, N.; Yamamoto, K.; Kobayashi, T.; Hirotomi, T.; Ogawa, H.; Miyazaki, H.; Yoshi, H. FcyRIIIb genotypes and smoking in periodontal disease progression among community-dwelling older adults in Japan. J. Periodontol. 2005, 76, 250–255. [Google Scholar] [CrossRef]
- Kaneko, S.; Kobayashi, T.; Yamamoto, K.; Jansen, M.D.; van de Winkel, J.G.J.; Yoshie, H. A novel polymorphism of Fc&RI (CD89) associated with aggressive periodontitis. Tissue Antigens 2004, 63, 572–577. [Google Scholar]
- Behm, C.; Blufstein, A.; Gahn, J.; Noroozkhan, N.; Moritz, A.; Raush-Fan, X.; Andrukhov, O. Soluble CD14 Enhances the Response of Periodontal Loigament Stem Cells to Toll-Like Receptor 2 Agonists. Mediat. Inflamm. 2019, 2019, 8127301. [Google Scholar] [CrossRef]
- Holla, L.I.; Buckova, D.; Fassmann, A.; Halabala, T.; Vasku, A.; Vacha, J. Promoter polymorphisms in the CD14 receptor gene and their potential association with the severity of chronic periodontitis. J. Med. Genet. 2002, 39, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Aisaiti, A.; Wu, Z.P.; Wang, T.T.; Zhao, J. Association of TLR-2 Gene Polymorphisms with the Risk of Periodontitis: A Meta-Analysis. Dis. Markers 2020, 2020, 9353958. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.-H.; Guan, Z.-Y.; Liang, W.-H.; Bai, G.-H.; Liu, J.-G. TLR4 polymorphism and periodontitis susceptibility: A meta-analysis. Medicine 2016, 95, e4845. [Google Scholar] [CrossRef] [PubMed]
- Khammissa, R.A.G.; Ballyram, R.; Jadwat, Y.; Fourie, J.; Lemmer, J.; Feller, L. Vitamin D Deficiency as It Relates to Oral Immunity and chronic Periodontitis. Int. J. Dent. 2018, 2018, 7315797. [Google Scholar] [CrossRef] [Green Version]
- Musacchio, E.; Binotto, P.; Silva-Netto, F.; Perissinotto, E.; Sartori, L. Bone-related polymorphisms and dental status in older men and women. Results of the longitudinal Pro.V.A. study. J. Dent. Sci. 2022, 17, 528–534. [Google Scholar] [CrossRef]
- Holla, L.I.; Jurajda, M.; Fassmann, A.; Dvorakova, N.; Znojil, V.; Vacha, J. Genetic variations in the matrix metalloproteinase-1 promoter and risk of susceptibility and/or severity of chronic periodontitis in the Czech population. J. Clin. Periodontol. 2004, 31, 685–690. [Google Scholar] [CrossRef]
- Hsiao, Y.F.; Yang, L.C.; Chou, Y.S.; Ho, Y.P.; Lin, Y.C.; Ho, K.Y.; Wu, Y.M.; Tsai, C.C. Matrix metalloproteinase-2, -9, and tissue inhibitor of MMP-2 gene polymorphisms in Taiwanese periodontitis patients. J. Dent. Sci. 2016, 11, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Mashhadiabbas, F.; Neamatzadeh, H.; Foroughi, E.; Dastgheib, S.A.; Farahnak, S.; Nasiri, R.; Ahmadi, S. Association of MMP-2-753C>T and MMP-9-1562C>T Polymorphisms with Chronic/Aggressive Periodontitis Risk: A Systematic Review and Meta-Analysis. J. Public Health 2019, 48, 1227–1238. [Google Scholar] [CrossRef]
- Salminen, A.; Pietiäinen, M.; Paju, S.; Sorsa, T.; Mäntylä, P.; Buhlin, K.; Sinisalo, J.; Pussinen, P.J. Common complement factor H polymorphisms are linked with periodontitis in elderly patients. J. Periodontol. 2022, 22. Epub ahead of print. [Google Scholar] [CrossRef]
- Kocher, T.; Sawaf, H.; Fanghanel, J.; Timm, R.; Meisel, P. Association between bone loss in periodontal disease and polymorphism of N-acetylotransferaze (NAT2). J. Clin. Periodontol. 2002, 29, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Cochran, D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, H. A systematic review and meta-anlysis on multiple cytokine gene polymorphisms in the pathogenesis of periodontits. Front. Immunol. 2022, 12, 713198. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.R.P.; Pessoa, L.D.S.; Shin, J., II; Alves, E.H.P.; Koga, R.S.; Smith, C.V.; Vasconcelos, D.F.P.V.; De Cuhna Pereira, A.C.T. Polymorphisms in the interleukin genes and chronic periodontitis: A field synopsis and revaluation by Bayesian approaches. Cytokines 2021, 138, 155361. [Google Scholar] [CrossRef] [PubMed]
- de Alencar, J.B.; Zacarias, J.M.V.; Tsuneto, P.Y.; Souza, V.H.; Silva, C.O.E.; Visentainer, J.E.L.; Sell, A.M. Influence of inflammasome NLRP3, and IL1B and IL2 gene polymorphisms in periodontitis susceptibility. PLoS ONE 2020, 15, e0227905. [Google Scholar] [CrossRef]
- Da Silva, F.R.P.; Galeno, J.G.; Bentes Leal, A.L.A.; Koga, R.S.; Batista, N.Y.; da Conceição Furtado, S.; Vasconcelos, D.F.P.; Carvalho, M.D.; Barcellos, J.F.M. Non-significant association between—330 T/G polymorphism in interleukin-2 gene and chronic periodontitis: Findings from a meta-analysis. BMC Oral Health 2020, 20, 58. [Google Scholar] [CrossRef] [Green Version]
- Reichert, S.; Stein, J.M.; Klapproth, J.; Zimmermann, U.; Reichert, Y.; Glaser, C.; Schaller, H.G.; Schulz, S. The genetic impact of the Q551R interleukin-4 receptor alpha polymorphism for aggressive or chronic periodontitis and occurrence of periodonrhopatic bacteria. Arch. Oral Biol. 2011, 6, 1485–1493. [Google Scholar] [CrossRef]
- Gonzales, J.R.; Kobayashi, T.; Michel, J.; Mann, J.; Yoshi, H.; Meyle, J. Interleukin-4 gene polymorphisms in Japanese and Caucasian patients with aggressive periodontitis. J. Clin. Periodontol. 2004, 31, 384–389. [Google Scholar] [CrossRef]
- Scarel-Caminga, R.M.; Trevilatto, P.C.; Souza, A.P.; Brito Jr, R.B.; Line, S.R.P. Investigation of IL-4 gene polymorphism in individuals with different levels of chronic periodontitis in a Brazilian population. J. Clin. Periodontol. 2003, 30, 341–345. [Google Scholar] [CrossRef]
- Jia, X.-W.; Yuan, Y.-D.; Yao, Z.-X.; Wu, C.-J.; Chen, X.; Chen, X.-H.; Lin, Y.-M.; Meng, X.-Y.; Zeng, X.-T.; Shao, J. Association between IL-4 and IL-4R Polymorphisms and Periodontitis: A Meta-Analysis. Dis. Markers 2017, 2017, 8021279. [Google Scholar] [CrossRef]
- Holla, L.I.; Fassmann, A.; Stejskalova, A.; Znojil, V.; Vanek, J.; Vacha, J. Analysis of the Interleukin-6 gene promoter polymorphisms in Czech patients with chronic periodontitis. J. Periodontol. 2004, 75, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Trevilatto, P.C.; Scarel-Caminaga, R.M.; de Brito, R.B., Jr.; de Souza, A.P.; Line, S.R.P. Polymorphism at position-174 of IL-6 gene is associated with susceptibility to chronic periodontitis in a Caucasian Brazilian population. J. Clin. Periodontol. 2003, 30, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Mehar, R.; Swarnakar, S.; Lakkakula, S.; Verma, H.K.; Bhaskar, L.V.K.S. Interleukin-6 gene-174G>C promoter polymorphism reduces the risk of periodontitis in Brazilian populations: A meta-analysis. J. Oral Biosci. 2021, 63, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Li, X.; Li, R. Genetic Relationship Between IL-6 rs1800796 Polymorphism and Susceptibility to Periodontitis. Immunol. Investig. 2019, 48, 268–282. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Zhou, Y.; Qiao, Y. Association between the IL-10 rs1800872 polymorphisms and periodontitis susceptibility. A meta-analysis. Medicine 2019, 98, e17113. [Google Scholar] [CrossRef]
- Sumer, A.P.; Nurten, K.; Gonca, C.K.; Sezgin, G.; Hulya, K.; Hasan, B. Association of interleukin-10 gene polymorphisms with severe generalized chronic periodontitis. J. Periodontol. 2007, 78, 493–497. [Google Scholar] [CrossRef]
- Bita, M.; Zahra, H.; Hamidreza, M.S.; Mehrnoosh, M. Analysis of interleukin-10 gene polymorphisms in patients with chronic periodontitis and healthy controls. Dent. Res. J. 2018, 15, 71–79. [Google Scholar]
- Ying, G.; Lu, L.; Xiaogian, W.; Fanzhen, H.; Yi, Z.; Mifang, Y.; Yan, X. Interleukin-10 polymorphisms affect the key periodontal pathogens in Chinese periodontitis patients. Sci. Rep. 2018, 8, 9068. [Google Scholar]
- Yamazaki, K.; Tabeta, K.; Nakajima, T.; Ohsawa, Y.; Ueki, K.; Itoh, H.; Yoshie, H. Interleukin-10 gene promotor polymorphism in Japanese patients with adult and early-onset periodontitis. J. Clin. Periodontol. 2001, 28, 828–832. [Google Scholar] [CrossRef]
- Dashash, M.; Blinkhorn, A.S.; Huthinson, I.V.; Pravica, V.; Drucker, D.B. The relationship between inyterleukin-10 gene polymorphism at position -1082 and susceptibility to gingivitis in children. J. Periodontol. 2005, 76, 1455–1462. [Google Scholar] [CrossRef]
- Mashhadiabbas, F.; Dastgheib, S.A.; Hashemzehi, A.; Bahrololoomi, Z.; Asadian, F.; Neamatzadeh, H.; Zare-Shehneh, M.; Daliri, K. Association of IL-10 -1082A>G, -819C>T, and -592C>A polymorphisms with susceptibility to chronic and aggressive periodontitis: A systematic review and meta-analysis. Inflamm. Res. 2021, 70, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Stolf, C.S.; Sacramento, C.M.; Paz, H.E.S.; Machado, R.A.; Ramos, L.P.; de Oliveira, L.D.; Cogo-Müller, K.; Santamaria, M.P.; Ruiz, K.G.S.; Casarin, R.C.V. IL10 promoter rs6667202 polymorphism is functional in health but not in grade c periodontitis patients: A pilot study. J. Periodontal. Res. 2022, 57, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Dhayanand, J.V.; Sangeetha, S.; Prakash, P.S.G.; Bob, J.J. Tumor necrosis factor-alpha-308 gene polymorphism in the association between gestational diabetes mellitus and chronic periodontitis in South Indian population. J. Pharmacol. Pharmacother. 2018, 9, 109–112. [Google Scholar]
- Jakovljevic, A.; Nikolic, N.; Jacimovic, J.; Miletic, M.; Andric, M.; Milasin, J.; Aminoshariae, A.; Azarpazhooh, A. Tumor Necrosis Factor Alpha-308 G/A Single-Nucleoide Polymorphism and Apical Periodontitis: An Updated Systematic Review and Meta-analysis. J. Endod. 2021, 47, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, X.; Sun, W. Association Between Tumor Necrosis Factor-α (G-308A) Polymorphism and Chronic Periodontitis, Aggressive Periodontitis, and Peri-implantitis: A Meta-analysis. J. Evid. Based Dent. Pract. 2021, 21, 101528. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, C.; Zheng, Y.; Huang, Y.; Zhong, Y.; Zhao, Z.; Ma, N.; Zhang, Z.; Zhang, L. Association of TNF-α -308G/A, -238G/A, -863C/A, -1031T/C, -857C/T polymorphisms with periodontitis susceptibility: Evidence from a meta-analysis of 52 studies. Medicine 2020, 99, e21851. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Sun, Y.-P.; Li, D.-G.; Wang, S.-H.; Shao, D. Transforming growth factor-β1 rs1800469 polymorphism and periodontitis risk: A meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 15569–15574. [Google Scholar] [PubMed]
- Shimada, Y.; Tai, H.; Endo, M.; Kobayashi, T.; Akazawa, K.; Yamazaki, K. Association of tumor necrosis factor receptor type 2 +587 gene polymorphism with severe chronic periodontitis. J. Clin. Periodontol. 2004, 31, 436–463. [Google Scholar] [CrossRef] [PubMed]
- Izakovicova Holla, L.; Fassmann, A.; Benes, P.; Halabala, T.; Znojil, V. 5 polymorphisms in the transforming growth factor-B 1 gene (TGF-B1) in adult periodontitis. J. Clin. Periodontol. 2002, 29, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ding, C.; Chen, X.; He, R.; Chen, N. Association of TGF-β1-509C/T, +869T/C, and +915G/C polymorphisms with periodontitis susceptibility. Oral Dis. 2015, 21, 443–450. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Position | Periodontitis | Race | Type of Study | References |
|---|---|---|---|---|---|
| HLA | DRB1*1501–DQB1*0602 | Positively associated with AgP | - | review | [10] |
| HLA | B*57, DQB1*08, DRB1*04, DRB4*, DQB1*0302 | Negatively associated with AgP and CP | German with Caucasian descent | a case control study | [24] |
| HLA | DR4 (subtypes 0401, 0404, 0405, 0408) | Positively associated with AgP | The ethno-geographic origin of the subjects was neutralized by stratified analysis | a case control study | [25] |
| HLA | DRB1*1401, DRB1*1501, DQB1*0503, DQBA*0602 | Positively associated with AgP | Japanese | a case control study | [26] |
| HLA | A9, B15 A2, B5 | Susceptibility factors for AgP Protective factors against AgP | Caucasian | meta-analysis | [27] |
| Gene | Position | Periodontitis | Race | Type of Study | References |
|---|---|---|---|---|---|
| MMP-1 | −1607 1G/2G, −519 A/G, −422 A/T | No associated with the susceptibility to periodontitis | Caucasian, Asian, or mixed (excluding the detailed ethnic results of mixed population in the original study) | meta-analysis | [22] |
| MMP-2 | −1575 G/A, −1306 C/T, −790 T/G, −735 C/T | ||||
| MMP-3 | −1171 5A/6A | ||||
| MMP-8 | −381 A/G, +17 C/G | ||||
| MMP-9 | −1562 C/T, +279 R/Q | ||||
| MMP-12 | −357 Asn/Ser | ||||
| MMP-13 | −77 A/G, 11A/12A | ||||
| MMP-1 | −519 A/G, −422 A/T | No associated with CP | Czech | a case control study | [48] |
| −1607 1G | Increased frequency of CP | ||||
| MMP-2 | −790 T, −790 TT, −790 T/G | Less susceptible to CP | Taiwanese | a case control study | [49] |
| −1562 T | Less susceptible to AgP | ||||
| MMP-2 | −753 C > T | No associated with periodontitis in overall population | Caucasians, Asians, Latinos | meta-analysis | [50] |
| MMP-9 | −1562 C > T | ||||
| MMP-2 | −753 C > T | Associated with periodontitis in Asians | |||
| MMP-9 | −1562 C > T | Associated with periodontitis in Caucasians |
| Gene | Chromosome | Position | Periodontitis | Race | Type of Study | References |
|---|---|---|---|---|---|---|
| Il-1α | 2q13-2q21 | —889 C/T T | Prevent periodontitis risk | - | meta-analysis | [54] |
| —889 C/T C | Strong association with periodontitis development | |||||
| rs1800587 | Positively associated with CP | - | meta-analysis | [55] | ||
| Il-1β | 2q13-2q21 | −511 C > T, −3954 C > T | Very trong association with periodontitis development | - | meta-analysis | [54] |
| rs1143634 | Positively associated with CP | - | meta-analysis | [55] | ||
| IL-2 | 4q26-2 | −330 T, −330 G | Positively associated with periodontitis | - | meta-analysis | [54] |
| −330T/G | No associated with CP | Caucasian, Asian, Mixed | meta-analysis | [57] | ||
| −166, −330 | Positively associated with CP | Mixed | acase-control study | [56] | ||
| Il-4 | 5q31.1 | Q551R | Positively associated with CP | German | a case control study | [58] |
| promoter, intron, allele 1, allele 2 | No associated with AgP | Caucasian, Japanese | a case-control study | [59] | ||
| −590 (C—>T) | No associated with CP | Brazilian | a case-control study | [60] | ||
| Q551R−33 C/T | Positively associated with CP Negatively associated with AgP | Caucasian, Asian, Mixed, Dravidian | meta-analysis | [61] | ||
| Il-6 | 7p21 | −572 G/C | Lower susceptibility to CP | Czech | a case-control study | [62] |
| −174 G/G | Higher susceptibility to CP | Brazilian | a case-control study | [63] | ||
| −174 G > C | Lower susceptibility to CP | Brazilian | meta-analysis | [64] | ||
| Il-10 | 1q31-32 | −824 AA/CC + CA | Positively associated with CP | Turkish | a case control study | [67] |
| −1082 G | Positively associated with CP | Iranian | ||||
| −592 AA, −819 TT, −819 ATA/ATA | Positively associated with CP | Chinese | a case-control study | [68] | ||
| −592 C > A | Increased risk of periodontitis | Overall population | ||||
| −1082 A > G | Increased risk of periodontitis | Caucasians | a case-control study | [69] | ||
| −819 C > T | Increased risk of periodontitis | Mixed | ||||
| −592 C > A | Increased risk of periodontitis | Asians, Mixed | meta-analysis | [71] | ||
| −592 C > A | Increased risk of CA | Overall population |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brodzikowska, A.; Górski, B. Polymorphisms in Genes Involved in Inflammation and Periodontitis: A Narrative Review. Biomolecules 2022, 12, 552. https://doi.org/10.3390/biom12040552
Brodzikowska A, Górski B. Polymorphisms in Genes Involved in Inflammation and Periodontitis: A Narrative Review. Biomolecules. 2022; 12(4):552. https://doi.org/10.3390/biom12040552
Chicago/Turabian StyleBrodzikowska, Aniela, and Bartłomiej Górski. 2022. "Polymorphisms in Genes Involved in Inflammation and Periodontitis: A Narrative Review" Biomolecules 12, no. 4: 552. https://doi.org/10.3390/biom12040552