
Ghrelin and Cancer: Examining the Roles of the Ghrelin Axis in Tumor Growth and Progression



Abstract
:1. Introduction
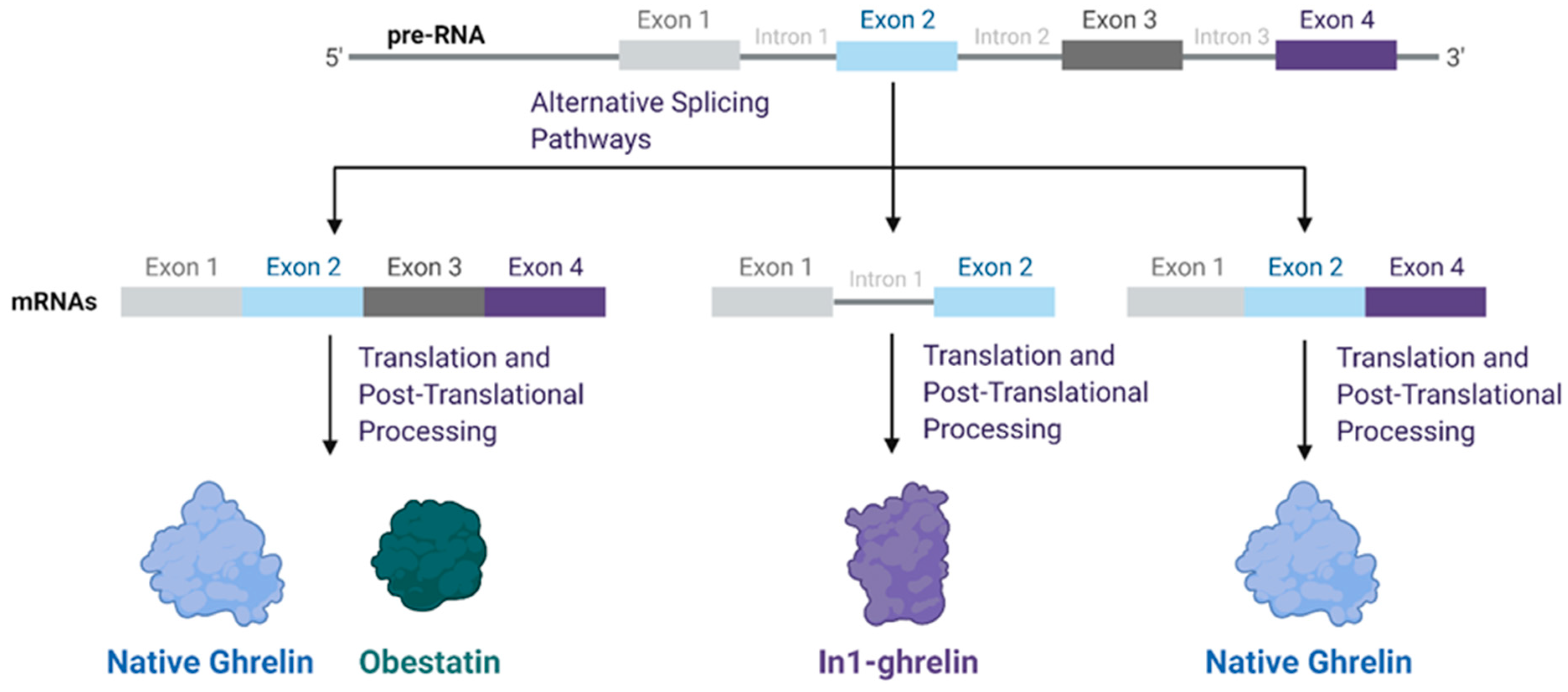
2. Ghrelin Structure and Function
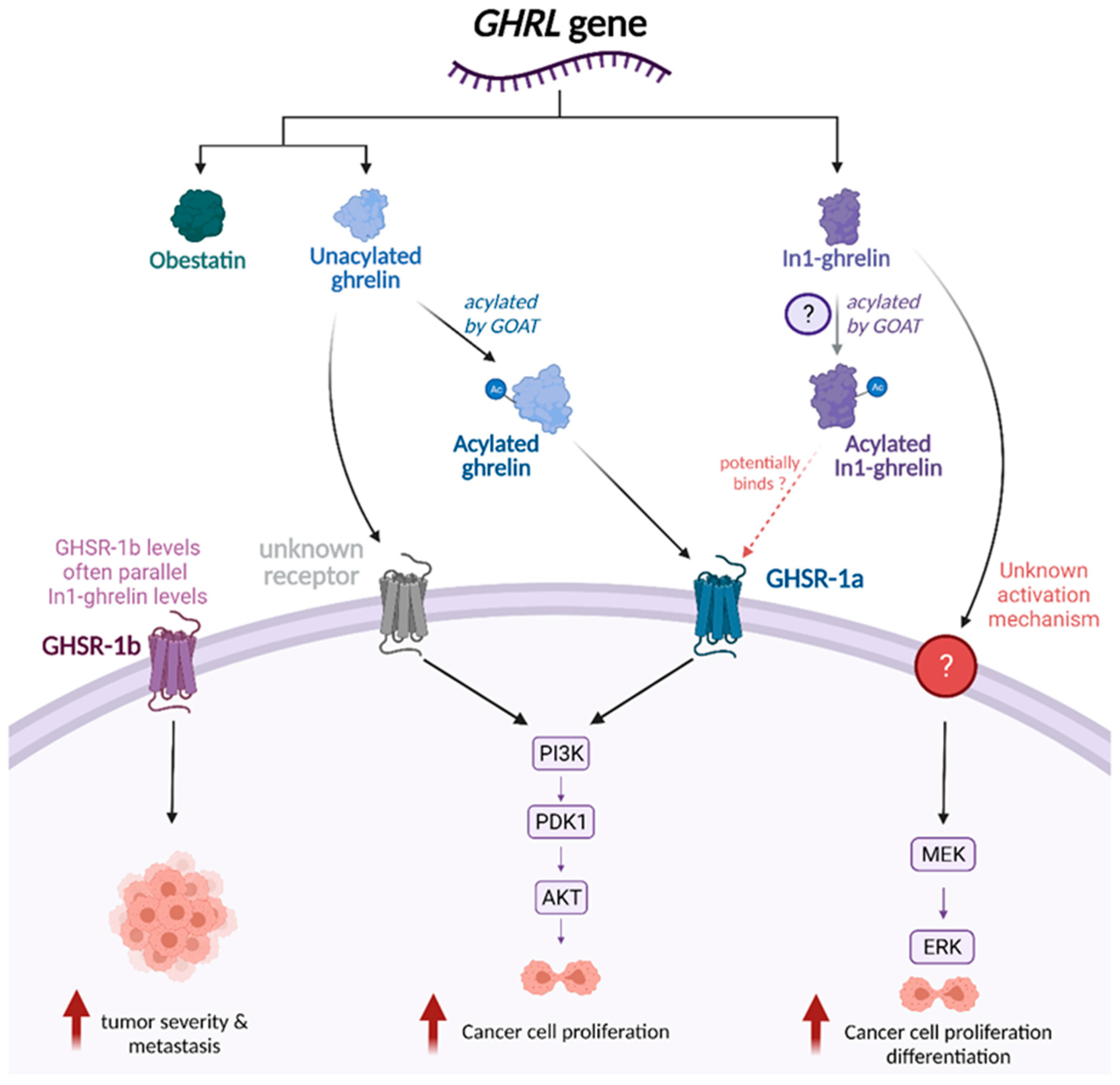
3. Functional Role of Ghrelin Splice Variants across Tumor Types
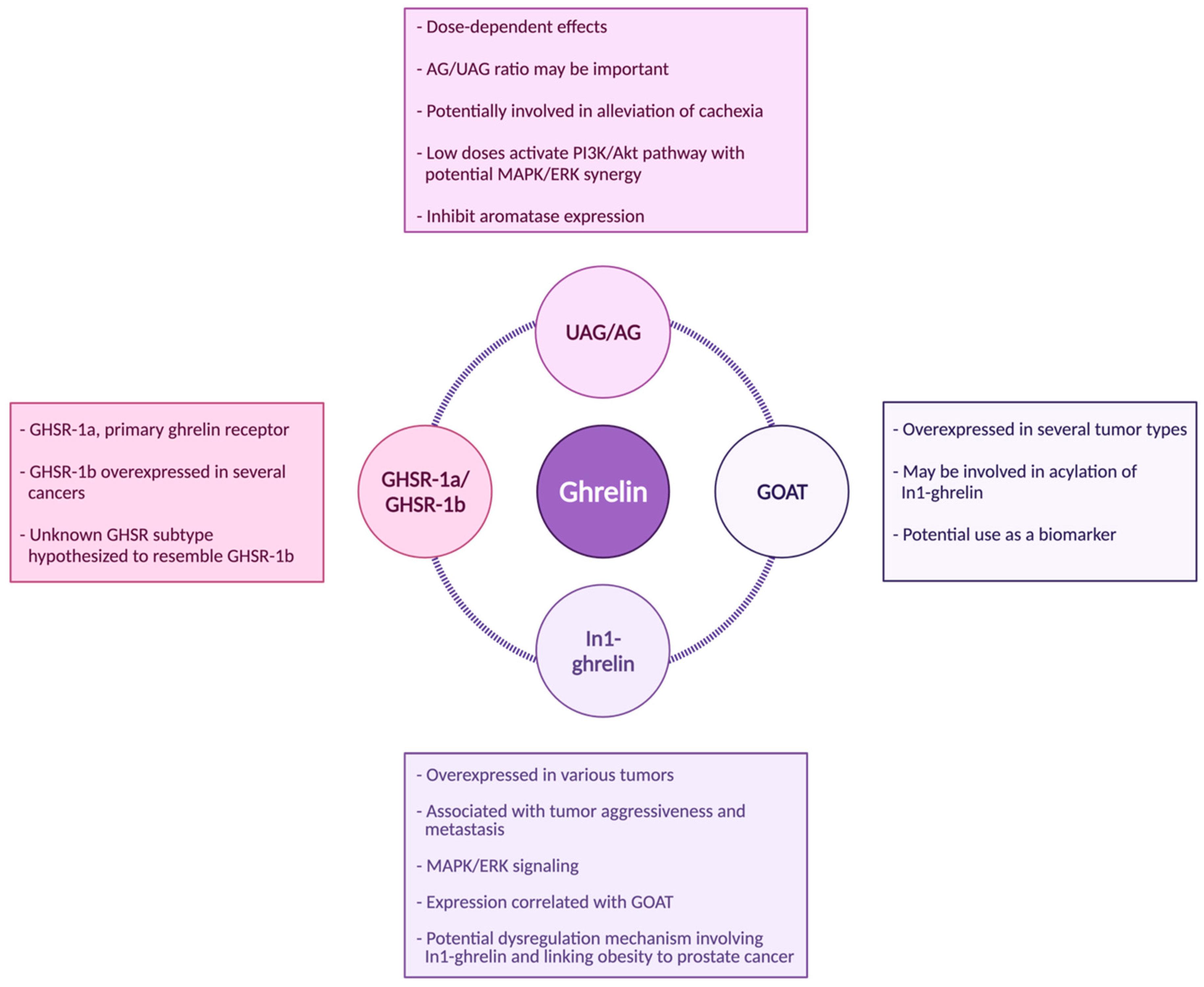
4. Acylated Ghrelin (AG) and Unacylated Ghrelin (UAG)
5. GHSR-1a/1b
6. GOAT
7. Ghrelin-Related Signaling Pathways
8. Ghrelin in the Prevention of Cancer Cachexia
9. Ghrelin, Obesity, and Cancer Feedback Loop
10. Interactions between Ghrelin and Other Hormones: Implications for Cancer
11. Strengths and Limitations
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gahete, M.D.; Cordoba-Chacon, J.; Salvatori, R.; Castano, J.P.; Kineman, R.D.; Luque, R.M. Metabolic regulation of ghrelin O-acyl transferase (GOAT) expression in the mouse hypothalamus, pituitary, and stomach. Mol. Cell Endocrinol. 2010, 317, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.M.; Wang, G.; Englander, E.W.; Kojima, M.; Greeley, G.H., Jr. Ghrelin, a new gastrointestinal endocrine peptide that stimulates insulin secretion: Enteric distribution, ontogeny, influence of endocrine, and dietary manipulations. Endocrinology 2002, 143, 185–190. [Google Scholar] [CrossRef]
- Wu, C.S.; Wei, Q.; Wang, H.; Kim, D.M.; Balderas, M.; Wu, G.; Lawler, J.; Safe, S.; Guo, S.; Devaraj, S.; et al. Protective Effects of Ghrelin on Fasting-Induced Muscle Atrophy in Aging Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Seim, I.; Collet, C.; Herington, A.C.; Chopin, L.K. Revised genomic structure of the human ghrelin gene and identification of novel exons, alternative splice variants and natural antisense transcripts. BMC Genomics 2007, 8, 298. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Lee, J.H.; Buras, E.D.; Yu, K.; Wang, R.; Smith, C.W.; Wu, H.; Sheikh-Hamad, D.; Sun, Y. Ghrelin receptor regulates adipose tissue inflammation in aging. Aging (Albany NY) 2016, 8, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.H.; Rees, D.J.; Andrews, Z.B.; Davies, J.S. Ghrelin mediated neuroprotection - A possible therapy for Parkinson’s disease? Neuropharmacology 2018, 136, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Buntwal, L.; Sassi, M.; Morgan, A.H.; Andrews, Z.B.; Davies, J.S. Ghrelin-Mediated Hippocampal Neurogenesis: Implications for Health and Disease. Trends Endocrinol. Metab. 2019, 30, 844–859. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Xu, H.; Guo, S.; Mertens-Talcott, S.U.; Sun, Y. Ghrelin Signaling in Immunometabolism and Inflamm-Aging. Adv. Exp. Med. Biol. 2018, 1090, 165–182. [Google Scholar] [CrossRef]
- Pradhan, G.; Samson, S.L.; Sun, Y. Ghrelin: Much more than a hunger hormone. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Asnicar, M.; Smith, R.G. Central and peripheral roles of ghrelin on glucose homeostasis. Neuroendocrinology 2007, 86, 215–228. [Google Scholar] [CrossRef]
- Smith, R.G.; Jiang, H.; Sun, Y. Developments in ghrelin biology and potential clinical relevance. Trends Endocrinol. Metab. 2005, 16, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Delporte, C. Structure and physiological actions of ghrelin. Scientifica (Cairo) 2013, 2013, 518909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nass, R.; Gaylinn, B.D.; Thorner, M.O. The ghrelin axis in disease: Potential therapeutic indications. Mol. Cell Endocrinol. 2011, 340, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stempniewicz, A.; Ceranowicz, P.; Warzecha, Z. Potential Therapeutic Effects of Gut Hormones, Ghrelin and Obestatin in Oral Mucositis. Int. J. Mol. Sci. 2019, 20, 1534. [Google Scholar] [CrossRef] [Green Version]
- El-Kott, A.F.; Shati, A.A.; Al-Kahtani, M.A.; Alqahtani, S. Acylated Ghrelin Renders Chemosensitive Ovarian Cancer Cells Resistant to Cisplatin Chemotherapy via Activation of the PI3K/Akt/mTOR Survival Pathway. Anal. Cell Pathol. (Amst.) 2019, 2019, 9627810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfili, L.; Cuccioloni, M.; Cecarini, V.; Mozzicafreddo, M.; Palermo, F.A.; Cocci, P.; Angeletti, M.; Eleuteri, A.M. Ghrelin induces apoptosis in colon adenocarcinoma cells via proteasome inhibition and autophagy induction. Apoptosis 2013, 18, 1188–1200. [Google Scholar] [CrossRef] [PubMed]
- Fung, J.N.; Seim, I.; Wang, D.; Obermair, A.; Chopin, L.K.; Chen, C. Expression and in vitro functions of the ghrelin axis in endometrial cancer. Horm. Cancer 2010, 1, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Soleyman-Jahi, S.; Sadeghi, F.; Pastaki Khoshbin, A.; Khani, L.; Roosta, V.; Zendehdel, K. Attribution of Ghrelin to Cancer; Attempts to Unravel an Apparent Controversy. Front. Oncol. 2019, 9, 1014. [Google Scholar] [CrossRef]
- Au, C.C.; Furness, J.B.; Brown, K.A. Ghrelin and Breast Cancer: Emerging Roles in Obesity, Estrogen Regulation, and Cancer. Front. Oncol. 2016, 6, 265. [Google Scholar] [CrossRef] [Green Version]
- Lanfranco, F.; Baldi, M.; Cassoni, P.; Bosco, M.; Ghe, C.; Muccioli, G. Ghrelin and prostate cancer. Vitam. Horm. 2008, 77, 301–324. [Google Scholar] [CrossRef]
- Ginter, G.; Ceranowicz, P.; Warzecha, Z. Protective and Healing Effects of Ghrelin and Risk of Cancer in the Digestive System. Int. J. Mol. Sci. 2021, 22, 10571. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, S.; Sato, T.; Kangawa, K.; Nakazato, M. The Homeostatic Force of Ghrelin. Cell Metab. 2018, 27, 786–804. [Google Scholar] [CrossRef] [Green Version]
- Ibanez-Costa, A.; Gahete, M.D.; Rivero-Cortes, E.; Rincon-Fernandez, D.; Nelson, R.; Beltran, M.; de la Riva, A.; Japon, M.A.; Venegas-Moreno, E.; Galvez, M.A.; et al. In1-ghrelin splicing variant is overexpressed in pituitary adenomas and increases their aggressive features. Sci. Rep. 2015, 5, 8714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahete, M.D.; Cordoba-Chacon, J.; Hergueta-Redondo, M.; Martinez-Fuentes, A.J.; Kineman, R.D.; Moreno-Bueno, G.; Luque, R.M.; Castano, J.P. A novel human ghrelin variant (In1-ghrelin) and ghrelin-O-acyltransferase are overexpressed in breast cancer: Potential pathophysiological relevance. PLoS ONE 2011, 6, e23302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, M.; Murphy, K.G.; le Roux, C.W.; Ghatei, M.A.; Bloom, S.R. Characterization of ghrelin-like immunoreactivity in human plasma. J. Clin. Endocrinol. Metab. 2005, 90, 2205–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatnik, M.; Soderstrom, C.I.; Dysinger, M.; Fraser, S.A. Prandial ghrelin attenuation provides evidence that des-acyl ghrelin may be an artifact of sample handling in human plasma. Bioanalysis 2012, 4, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Hormaechea-Agulla, D.; Gahete, M.D.; Jimenez-Vacas, J.M.; Gomez-Gomez, E.; Ibanez-Costa, A.; F, L.L.; Rivero-Cortes, E.; Sarmento-Cabral, A.; Valero-Rosa, J.; Carrasco-Valiente, J.; et al. The oncogenic role of the In1-ghrelin splicing variant in prostate cancer aggressiveness. Mol. Cancer 2017, 16, 146. [Google Scholar] [CrossRef]
- Luque, R.M.; Sampedro-Nunez, M.; Gahete, M.D.; Ramos-Levi, A.; Ibanez-Costa, A.; Rivero-Cortes, E.; Serrano-Somavilla, A.; Adrados, M.; Culler, M.D.; Castano, J.P.; et al. In1-ghrelin, a splice variant of ghrelin gene, is associated with the evolution and aggressiveness of human neuroendocrine tumors: Evidence from clinical, cellular and molecular parameters. Oncotarget 2015, 6, 19619–19633. [Google Scholar] [CrossRef] [Green Version]
- Rincon-Fernandez, D.; Culler, M.D.; Tsomaia, N.; Moreno-Bueno, G.; Luque, R.M.; Gahete, M.D.; Castano, J.P. In1-ghrelin splicing variant is associated with reduced disease-free survival of breast cancer patients and increases malignancy of breast cancer cells lines. Carcinogenesis 2018, 39, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Vacas, J.M.; Montero-Hidalgo, A.J.; Gomez-Gomez, E.; Fuentes-Fayos, A.C.; Ruiz-Pino, F.; Guler, I.; Camargo, A.; Anglada, F.J.; Carrasco-Valiente, J.; Tena-Sempere, M.; et al. In1-Ghrelin Splicing Variant as a Key Element in the Pathophysiological Association Between Obesity and Prostate Cancer. J. Clin. Endocrinol. Metab 2021, 106, e4956–e4968. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, P.L.; Murray, R.E.; Yeh, A.H.; McNamara, J.F.; Duncan, R.P.; Francis, G.D.; Herington, A.C.; Chopin, L.K. Expression and function of the ghrelin axis, including a novel preproghrelin isoform, in human breast cancer tissues and cell lines. Endocr. Relat. Cancer 2005, 12, 839–850. [Google Scholar] [CrossRef]
- Yeh, A.H.; Jeffery, P.L.; Duncan, R.P.; Herington, A.C.; Chopin, L.K. Ghrelin and a novel preproghrelin isoform are highly expressed in prostate cancer and ghrelin activates mitogen-activated protein kinase in prostate cancer. Clin. Cancer Res. 2005, 11, 8295–8303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Heshmati, M.; Fattahi, S.; Bagheri, N.; Alibeigi, F.M.; Taheri, F.; Anjomshoa, M.; Jami, M.S.; Ghatreh Samani, M. The relation between the ghrelin receptor and FOXP3 in bladder cancer. Biotech. Histochem 2021, 96, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.X.; Wang, W.P.; Zhao, P.W.; Li, C.B. Ghrelin attenuates the growth of HO-8910 ovarian cancer cells through the ERK pathway. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef] [Green Version]
- Fooladi, S.; Akbari, H.; Abolhassani, M.; Sadeghi, E.; Fallah, H. Can Estradiol and Ghrelin Play a Protective Role in Epithelial Ovarian Cancer Incidence in Postmenopausal Women? Arch. Med. Res. 2021, 52, 324–331. [Google Scholar] [CrossRef]
- Pritchett, N.R.; Maziarz, M.; Shu, X.O.; Kamangar, F.; Dawsey, S.M.; Fan, J.H.; Ji, B.T.; Gao, Y.T.; Xiang, Y.B.; Qiao, Y.L.; et al. Serum ghrelin and esophageal and gastric cancer in two cohorts in China. Int. J. Cancer 2020, 146, 2728–2735. [Google Scholar] [CrossRef]
- Hu, X.L.; Zhu, Y.J.; Hu, C.H.; You, L.; Wu, J.; He, X.Y.; Huang, W.J.; Wu, Z.H. Ghrelin Affects Gastric Cancer Progression by Activating AMPK Signaling Pathway. Biochem. Genet. 2021, 59, 652–667. [Google Scholar] [CrossRef]
- Tian, C.; Zhang, L.; Hu, D.; Ji, J. Ghrelin induces gastric cancer cell proliferation, migration, and invasion through GHS-R/NF-kappaB signaling pathway. Mol. Cell Biochem. 2013, 382, 163–172. [Google Scholar] [CrossRef]
- Au, C.C.; Furness, J.B.; Britt, K.; Oshchepkova, S.; Ladumor, H.; Soo, K.Y.; Callaghan, B.; Gerard, C.; Inghirami, G.; Mittal, V.; et al. Three-dimensional growth of breast cancer cells potentiates the anti-tumor effects of unacylated ghrelin and AZP-531. Elife 2020, 9, e56913. [Google Scholar] [CrossRef]
- Konturek, P.C.; Burnat, G.; Rau, T.; Hahn, E.G.; Konturek, S. Effect of adiponectin and ghrelin on apoptosis of Barrett adenocarcinoma cell line. Dig. Dis. Sci. 2008, 53, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, P.L.; Herington, A.C.; Chopin, L.K. Expression and action of the growth hormone releasing peptide ghrelin and its receptor in prostate cancer cell lines. J. Endocrinol. 2002, 172, R7–R11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawnicka, H.; Melen-Mucha, G.; Motylewska, E.; Mucha, S.; Stepien, H. Modulation of ghrelin axis influences the growth of colonic and prostatic cancer cells in vitro. Pharmacol. Rep. 2012, 64, 951–959. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Feng, L. Ghrelin Regulates Cyclooxygenase-2 Expression and Promotes Gastric Cancer Cell Progression. Comput. Math. Methods Med. 2021, 2021, 5576808. [Google Scholar] [CrossRef]
- Lien, G.S.; Lin, C.H.; Yang, Y.L.; Wu, M.S.; Chen, B.C. Ghrelin induces colon cancer cell proliferation through the GHS-R, Ras, PI3K, Akt, and mTOR signaling pathways. Eur. J. Pharmacol. 2016, 776, 124–131. [Google Scholar] [CrossRef]
- Waseem, T.; Duxbury, M.; Ashley, S.W.; Robinson, M.K. Ghrelin promotes intestinal epithelial cell proliferation through PI3K/Akt pathway and EGFR trans-activation both converging to ERK 1/2 phosphorylation. Peptides 2014, 52, 113–121. [Google Scholar] [CrossRef]
- Waseem, T.; Javaid Ur, R.; Ahmad, F.; Azam, M.; Qureshi, M.A. Role of ghrelin axis in colorectal cancer: A novel association. Peptides 2008, 29, 1369–1376. [Google Scholar] [CrossRef]
- Barzon, L.; Pacenti, M.; Masi, G.; Stefani, A.L.; Fincati, K.; Palu, G. Loss of growth hormone secretagogue receptor 1a and overexpression of type 1b receptor transcripts in human adrenocortical tumors. Oncology 2005, 68, 414–421. [Google Scholar] [CrossRef]
- Yang, J.; Brown, M.S.; Liang, G.; Grishin, N.V.; Goldstein, J.L. Identification of the acyltransferase that octanoylates ghrelin, an appetite-stimulating peptide hormone. Cell 2008, 132, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Martinez, A.D.; Gahete, M.D.; Sanchez-Sanchez, R.; Alors-Perez, E.; Pedraza-Arevalo, S.; Serrano-Blanch, R.; Martinez-Fuentes, A.J.; Galvez-Moreno, M.A.; Castano, J.P.; Luque, R.M. Ghrelin-O-Acyltransferase (GOAT) Enzyme as a Novel Potential Biomarker in Gastroenteropancreatic Neuroendocrine Tumors. Clin. Transl. Gastroenterol. 2018, 9, 196. [Google Scholar] [CrossRef]
- Gomez-Gomez, E.; Jimenez-Vacas, J.M.; Carrasco-Valiente, J.; Herrero-Aguayo, V.; Blanca-Pedregosa, A.M.; Leon-Gonzalez, A.J.; Valero-Rosa, J.; Fernandez-Rueda, J.L.; Gonzalez-Serrano, T.; Lopez-Miranda, J.; et al. Plasma ghrelin O-acyltransferase (GOAT) enzyme levels: A novel non-invasive diagnosis tool for patients with significant prostate cancer. J. Cell Mol. Med. 2018, 22, 5688–5697. [Google Scholar] [CrossRef] [PubMed]
- Hormaechea-Agulla, D.; Gomez-Gomez, E.; Ibanez-Costa, A.; Carrasco-Valiente, J.; Rivero-Cortes, E.; F, L.L.; Pedraza-Arevalo, S.; Valero-Rosa, J.; Sanchez-Sanchez, R.; Ortega-Salas, R.; et al. Ghrelin O-acyltransferase (GOAT) enzyme is overexpressed in prostate cancer, and its levels are associated with patient’s metabolic status: Potential value as a non-invasive biomarker. Cancer Lett. 2016, 383, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Vacas, J.M.; Gomez-Gomez, E.; Montero-Hidalgo, A.J.; Herrero-Aguayo, V.; F, L.L.; Sanchez-Sanchez, R.; Guler, I.; Blanca, A.; Mendez-Vidal, M.J.; Carrasco, J.; et al. Clinical Utility of Ghrelin-O-Acyltransferase (GOAT) Enzyme as a Diagnostic Tool and Potential Therapeutic Target in Prostate Cancer. J. Clin. Med. 2019, 8, 2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, P.Y.; Fan, X.M. The proliferative effects of ghrelin on human gastric cancer AGS cells. J. Dig. Dis. 2012, 13, 453–458. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, T. Ghrelin inhibits cisplatin-induced MDA-MB-231 breast cancer cell apoptosis via PI3K/Akt/mTOR signaling. Exp. Ther. Med. 2020, 19, 1633–1640. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Yao, J.; Huang, R.; Wang, Y.; Jia, M.; Huang, Y. Ghrelin promotes human non-small cell lung cancer A549 cell proliferation through PI3K/Akt/mTOR/P70S6K and ERK signaling pathways. Biochem. Biophys. Res. Commun. 2018, 498, 616–620. [Google Scholar] [CrossRef]
- Duxbury, M.S.; Waseem, T.; Ito, H.; Robinson, M.K.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. Ghrelin promotes pancreatic adenocarcinoma cellular proliferation and invasiveness. Biochem. Biophys. Res. Commun. 2003, 309, 464–468. [Google Scholar] [CrossRef]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef] [Green Version]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef]
- Granata, R.; Settanni, F.; Biancone, L.; Trovato, L.; Nano, R.; Bertuzzi, F.; Destefanis, S.; Annunziata, M.; Martinetti, M.; Catapano, F.; et al. Acylated and unacylated ghrelin promote proliferation and inhibit apoptosis of pancreatic beta-cells and human islets: Involvement of 3’,5’-cyclic adenosine monophosphate/protein kinase A, extracellular signal-regulated kinase 1/2, and phosphatidyl inositol 3-Kinase/Akt signaling. Endocrinology 2007, 148, 512–529. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Ahmed, M.; Hussain, A.; Assad, L.; Al-Dayel, F.; Bavi, P.; Al-Kuraya, K.S.; Munkarah, A. Cyclooxygenase-2 inhibition inhibits PI3K/AKT kinase activity in epithelial ovarian cancer. Int. J. Cancer 2010, 126, 382–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Sharma, K. COX-2 signaling and cancer: New players in old arena. Curr. Drug Targets 2014, 15, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Martinson, H.A.; Lyons, T.R.; Giles, E.D.; Borges, V.F.; Schedin, P. Developmental windows of breast cancer risk provide opportunities for targeted chemoprevention. Exp. Cell Res. 2013, 319, 1671–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, T.R.; Borges, V.F.; Betts, C.B.; Guo, Q.; Kapoor, P.; Martinson, H.A.; Jindal, S.; Schedin, P. Cyclooxygenase-2-dependent lymphangiogenesis promotes nodal metastasis of postpartum breast cancer. J. Clin. Investig. 2014, 124, 3901–3912. [Google Scholar] [CrossRef] [PubMed]
- Lyons, T.R.; O’Brien, J.; Borges, V.F.; Conklin, M.W.; Keely, P.J.; Eliceiri, K.W.; Marusyk, A.; Tan, A.C.; Schedin, P. Postpartum mammary gland involution drives progression of ductal carcinoma in situ through collagen and COX-2. Nat. Med. 2011, 17, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, G.L.; Crump, L.S.; Young, C.M.; Wessells, V.M.; McQueen, C.M.; Wall, S.W.; Gustafson, T.L.; Fan, Y.Y.; Chapkin, R.S.; Porter, W.W.; et al. Cross-talk between SIM2s and NFkappaB regulates cyclooxygenase 2 expression in breast cancer. Breast Cancer Res. 2019, 21, 131. [Google Scholar] [CrossRef]
- Slomiany, B.L.; Slomiany, A. Mechanism of Cytosolic Phospholipase A(2) Activation in Ghrelin Protection of Salivary Gland Acinar Cells against Ethanol Cytotoxicity. Adv. Pharmacol. Sci. 2010, 2010, 269274. [Google Scholar] [CrossRef] [Green Version]
- Slomiany, B.L.; Slomiany, A. Involvement of constitutive nitric oxide synthase in ghrelin-induced cytosolic phospholipase A(2) activation in gastric mucosal cell protection against ethanol cytotoxicity. Inflammopharmacology 2009, 17, 245–253. [Google Scholar] [CrossRef]
- Slomiany, B.L.; Slomiany, A. Ghrelin Protection against Cytotoxic Effect of Ethanol on Rat Salivary Mucin Synthesis involves Cytosolic Phospholipase A2 Activation through S-Nitrosylation. Int. J. Biomed. Sci. 2010, 6, 37–44. [Google Scholar] [PubMed]
- Zeng, X.; Chen, P.; Zhao, L.; Chen, S. Acylated and unacylated ghrelin relieve cancer cachexia in mice through multiple mechanisms. Chin. J. Physiol. 2020, 63, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Uysal, P.; Afsar, C.U.; Sozer, V.; Inanc, B.; Agaoglu, F.; Gural, Z.; Fazlioglu, N.Y.; Cuhadaroglu, C.; Uzun, H. Evaluation of the relationship between serum ghrelin levels and cancer cachexia in patients with locally advanced nonsmall-cell lung cancer treated with chemoradiotherapy. J. Cancer Res. Ther. 2020, 16, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Blum, D.; de Wolf-Linder, S.; Oberholzer, R.; Brandle, M.; Hundsberger, T.; Strasser, F. Natural ghrelin in advanced cancer patients with cachexia, a case series. J. Cachexia Sarcopenia Muscle 2021. [Google Scholar] [CrossRef]
- DeBoer, M.D. Emergence of ghrelin as a treatment for cachexia syndromes. Nutrition 2008, 24, 806–814. [Google Scholar] [CrossRef]
- Currow, D.C.; Maddocks, M.; Cella, D.; Muscaritoli, M. Efficacy of Anamorelin, a Novel Non-Peptide Ghrelin Analogue, in Patients with Advanced Non-Small Cell Lung Cancer (NSCLC) and Cachexia-Review and Expert Opinion. Int. J. Mol. Sci. 2018, 19, 3471. [Google Scholar] [CrossRef] [Green Version]
- Temel, J.S.; Abernethy, A.P.; Currow, D.C.; Friend, J.; Duus, E.M.; Yan, Y.; Fearon, K.C. Anamorelin in patients with non-small-cell lung cancer and cachexia (ROMANA 1 and ROMANA 2): Results from two randomised, double-blind, phase 3 trials. Lancet Oncol. 2016, 17, 519–531. [Google Scholar] [CrossRef]
- Katakami, N.; Uchino, J.; Yokoyama, T.; Naito, T.; Kondo, M.; Yamada, K.; Kitajima, H.; Yoshimori, K.; Sato, K.; Saito, H.; et al. Anamorelin (ONO-7643) for the treatment of patients with non-small cell lung cancer and cachexia: Results from a randomized, double-blind, placebo-controlled, multicenter study of Japanese patients (ONO-7643-04). Cancer 2018, 124, 606–616. [Google Scholar] [CrossRef]
- Chen, M.; Tsai, C.W.; Chang, W.S.; Xiong, G.Y.; Xu, Y.; Bau, D.T.; Gu, J. High circulating insulin-like growth factor-1 reduces the risk of renal cell carcinoma: A Mendelian randomization study. Carcinogenesis 2021, 42, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Knuppel, A.; Papadimitriou, N.; Martin, R.M.; Tsilidis, K.K.; Smith-Byrne, K.; Fensom, G.; Perez-Cornago, A.; Travis, R.C.; Key, T.J.; et al. Insulin-like growth factor-1, insulin-like growth factor-binding protein-3, and breast cancer risk: Observational and Mendelian randomization analyses with approximately 430,000 women. Ann. Oncol. 2020, 31, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Northrup, R.; Kuroda, K.; Duus, E.M.; Barnes, S.R.; Cheatham, L.; Wiley, T.; Pietra, C. Effect of ghrelin and anamorelin (ONO-7643), a selective ghrelin receptor agonist, on tumor growth in a lung cancer mouse xenograft model. Support. Care Cancer 2013, 21, 2409–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Roux, C.W.; Patterson, M.; Vincent, R.P.; Hunt, C.; Ghatei, M.A.; Bloom, S.R. Postprandial plasma ghrelin is suppressed proportional to meal calorie content in normal-weight but not obese subjects. J. Clin. Endocrinol. Metab. 2005, 90, 1068–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docanto, M.M.; Yang, F.; Callaghan, B.; Au, C.C.; Ragavan, R.; Wang, X.; Furness, J.B.; Andrews, Z.B.; Brown, K.A. Ghrelin and des-acyl ghrelin inhibit aromatase expression and activity in human adipose stromal cells: Suppression of cAMP as a possible mechanism. Breast Cancer Res. Treat. 2014, 147, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lin, L.; Yue, J.; Wu, C.S.; Guo, C.A.; Wang, R.; Yu, K.J.; Devaraj, S.; Murano, P.; Chen, Z.; et al. Suppression of Ghrelin Exacerbates HFCS-Induced Adiposity and Insulin Resistance. Int. J. Mol. Sci. 2017, 18, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Asnicar, M.; Saha, P.K.; Chan, L.; Smith, R.G. Ablation of ghrelin improves the diabetic but not obese phenotype of ob/ob mice. Cell Metab. 2006, 3, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Au, C.C.; Docanto, M.M.; Zahid, H.; Raffaelli, F.M.; Ferrero, R.L.; Furness, J.B.; Brown, K.A. Des-acyl ghrelin inhibits the capacity of macrophages to stimulate the expression of aromatase in breast adipose stromal cells. J. Steroid Biochem. Mol. Biol. 2017, 170, 49–53. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef]
- Shan, X.; Yeo, G.S. Central leptin and ghrelin signalling: Comparing and contrasting their mechanisms of action in the brain. Rev. Endocr. Metab. Disord. 2011, 12, 197–209. [Google Scholar] [CrossRef]
- Monti, V.; Carlson, J.J.; Hunt, S.C.; Adams, T.D. Relationship of ghrelin and leptin hormones with body mass index and waist circumference in a random sample of adults. J. Am. Diet. Assoc. 2006, 106, 822–828; quiz 829–830. [Google Scholar] [CrossRef]
- Lin, T.C.; Hsiao, M. Leptin and Cancer: Updated Functional Roles in Carcinogenesis, Therapeutic Niches, and Developments. Int. J. Mol. Sci. 2021, 22, 2870. [Google Scholar] [CrossRef]
- Fabian, C.J.; Klemp, J.R.; Marchello, N.J.; Vidoni, E.D.; Sullivan, D.K.; Nydegger, J.L.; Phillips, T.A.; Kreutzjans, A.L.; Hendry, B.; Befort, C.A.; et al. Rapid Escalation of High-Volume Exercise during Caloric Restriction; Change in Visceral Adipose Tissue and Adipocytokines in Obese Sedentary Breast Cancer Survivors. Cancers 2021, 13, 4871. [Google Scholar] [CrossRef]
- Yoon, Y.S.; Kwon, A.R.; Lee, Y.K.; Oh, S.W. Circulating adipokines and risk of obesity related cancers: A systematic review and meta-analysis. Obes. Res. Clin. Pract. 2019, 13, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Bowers, L.W.; Rossi, E.L.; McDonell, S.B.; Doerstling, S.S.; Khatib, S.A.; Lineberger, C.G.; Albright, J.E.; Tang, X.; deGraffenried, L.A.; Hursting, S.D. Leptin Signaling Mediates Obesity-Associated CSC Enrichment and EMT in Preclinical TNBC Models. Mol. Cancer Res. 2018, 16, 869–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, N.K.; Sharma, D.; Ding, X.; Lin, S.; Marra, F.; Merlin, D.; Anania, F.A. Concomitant activation of the JAK/STAT, PI3K/AKT, and ERK signaling is involved in leptin-mediated promotion of invasion and migration of hepatocellular carcinoma cells. Cancer Res. 2007, 67, 2497–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunwobi, O.; Mutungi, G.; Beales, I.L. Leptin stimulates proliferation and inhibits apoptosis in Barrett’s esophageal adenocarcinoma cells by cyclooxygenase-2-dependent, prostaglandin-E2-mediated transactivation of the epidermal growth factor receptor and c-Jun NH2-terminal kinase activation. Endocrinology 2006, 147, 4505–4516. [Google Scholar] [CrossRef]
- Beales, I.L.; Ogunwobi, O.O. Leptin synergistically enhances the anti-apoptotic and growth-promoting effects of acid in OE33 oesophageal adenocarcinoma cells in culture. Mol. Cell. Endocrinol. 2007, 274, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Vargas, A.K.; Garcia-Rodriguez, E.; Olea-Flores, M.; Mendoza-Catalan, M.A.; Flores-Alfaro, E.; Navarro-Tito, N. Pro-angiogenic activity and vasculogenic mimicry in the tumor microenvironment by leptin in cancer. Cytokine Growth Factor Rev. 2021, 62, 23–41. [Google Scholar] [CrossRef]
- Tong, J.; Prigeon, R.L.; Davis, H.W.; Bidlingmaier, M.; Kahn, S.E.; Cummings, D.E.; Tschop, M.H.; D’Alessio, D. Ghrelin suppresses glucose-stimulated insulin secretion and deteriorates glucose tolerance in healthy humans. Diabetes 2010, 59, 2145–2151. [Google Scholar] [CrossRef] [Green Version]
- Gauna, C.; Meyler, F.M.; Janssen, J.A.; Delhanty, P.J.; Abribat, T.; van Koetsveld, P.; Hofland, L.J.; Broglio, F.; Ghigo, E.; van der Lely, A.J. Administration of acylated ghrelin reduces insulin sensitivity, whereas the combination of acylated plus unacylated ghrelin strongly improves insulin sensitivity. J. Clin. Endocrinol. Metab. 2004, 89, 5035–5042. [Google Scholar] [CrossRef] [Green Version]
- Benso, A.; St-Pierre, D.H.; Prodam, F.; Gramaglia, E.; Granata, R.; van der Lely, A.J.; Ghigo, E.; Broglio, F. Metabolic effects of overnight continuous infusion of unacylated ghrelin in humans. Eur. J. Endocrinol. 2012, 166, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Zhang, Q.; Dong, H.; Xiang, X.; Zhang, W.; Zhang, Y.; Li, Y. Effects of Des-acyl Ghrelin on Insulin Sensitivity and Macrophage Polarization in Adipose Tissue. J. Transl. Int. Med. 2021, 9, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Chiefari, E.; Mirabelli, M.; La Vignera, S.; Tanyolac, S.; Foti, D.P.; Aversa, A.; Brunetti, A. Insulin Resistance and Cancer: In Search for a Causal Link. Int. J. Mol. Sci. 2021, 22, 11137. [Google Scholar] [CrossRef] [PubMed]
- Bowers, L.W.; Rossi, E.L.; O’Flanagan, C.H.; deGraffenried, L.A.; Hursting, S.D. The Role of the Insulin/IGF System in Cancer: Lessons Learned from Clinical Trials and the Energy Balance-Cancer Link. Front. Endocrinol. (Lausanne) 2015, 6, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giles, E.D.; Wellberg, E.A.; Astling, D.P.; Anderson, S.M.; Thor, A.D.; Jindal, S.; Tan, A.C.; Schedin, P.S.; Maclean, P.S. Obesity and overfeeding affecting both tumor and systemic metabolism activates the progesterone receptor to contribute to postmenopausal breast cancer. Cancer Res. 2012, 72, 6490–6501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.M.Y.; Wellberg, E.A.; Kopp, J.L.; Johnson, J.D. Hyperinsulinemia in Obesity, Inflammation, and Cancer. Diabetes Metab J. 2021, 45, 285–311. [Google Scholar] [CrossRef]
- Giles, E.D.; Jindal, S.; Wellberg, E.A.; Schedin, T.; Anderson, S.M.; Thor, A.D.; Edwards, D.P.; MacLean, P.S.; Schedin, P. Metformin inhibits stromal aromatase expression and tumor progression in a rodent model of postmenopausal breast cancer. Breast Cancer Res. 2018, 20, 50. [Google Scholar] [CrossRef]
- Al-Massadi, O.; Muller, T.; Tschop, M.; Dieguez, C.; Nogueiras, R. Ghrelin and LEAP-2: Rivals in Energy Metabolism. Trends Pharmacol. Sci. 2018, 39, 685–694. [Google Scholar] [CrossRef]
- Aslanipour, B.; Alan, M.; Demir, I. Decreased levels of liver-expressed antimicrobial peptide-2 and ghrelin are related to insulin resistance in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2020, 36, 222–225. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, R.; Tena-Sempere, M.; Aguilar, E.; Pinilla, L. Ghrelin effects on gonadotropin secretion in male and female rats. Neurosci. Lett. 2004, 362, 103–107. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, R.; Tena-Sempere, M.; Navarro, V.M.; Barreiro, M.L.; Castellano, J.M.; Aguilar, E.; Pinilla, L. Effects of ghrelin upon gonadotropin-releasing hormone and gonadotropin secretion in adult female rats: In vivo and in vitro studies. Neuroendocrinology 2005, 82, 245–255. [Google Scholar] [CrossRef]
- Grundker, C.; Emons, G. The Role of Gonadotropin-Releasing Hormone in Cancer Cell Proliferation and Metastasis. Front. Endocrinol. (Lausanne) 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Cell Line | Ghrelin Acylation Status | Concentration | Effect | Reference |
|---|---|---|---|---|---|
| Breast | MCF7 (ER+/PR+/HER2-) | Acylated | 0–1000 nM | No effect | [32] |
| SKBR3 (HER2+) | Unacylated | 0.1 nM | Inhibited growth | [40] | |
| MDA-MB-435 (TNBC) | Acylated | 0.1, 1, 10, 100 nM | ↑ proliferation | [32] | |
| MDA-MB-231 (TNBC) | Acylated | 10 and 100 nM | ↑ proliferation | [32] | |
| Ovarian | A2780 | Acylated | 1 nM | ↑ proliferation | [15] |
| Endometrial | HEC1B | Acylated | 10 and 100 nM | ↑ proliferation | [17] |
| KLE | Acylated | 1, 10, 100 nM | ↑ proliferation | [17] | |
| Esophageal | OE-19 | Not Specified | 20–450 nM | No effect on apoptosis | [41] |
| Prostate | PC3 | Acylated | 5 and 10 nM | ↑ proliferation | [42] |
| DU145 | Unacylated | 100 and 10,000 nM | Inhibited growth | [43] | |
| Acylated | 10–1000 nM | Inhibited growth | [43] | ||
| Gastric | AGS | Not Specified | Overexpression | ↑ apoptosis | [38] |
| Not Specified | 10 and 100 nM | ↑ proliferation | [39] | ||
| Not Specified | 10 nM | ↑ cell migration and invasion ↓ apoptosis | [44] | ||
| SGC7901 | Not Specified | 1, 10, 100 nM | ↑ proliferation | [39] | |
| Colon | HT-29 | Acylated | 0.1 and 1 nM | ↑ proliferation | [45] |
| Caco-2 | Acylated | 1 and 10 nM | ↑ proliferation | [46] | |
| Unacylated | 1 and 10 nM | ↑ proliferation | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotta, A.S.; Kelling, A.S.; Corleto, K.A.; Sun, Y.; Giles, E.D. Ghrelin and Cancer: Examining the Roles of the Ghrelin Axis in Tumor Growth and Progression. Biomolecules 2022, 12, 483. https://doi.org/10.3390/biom12040483
Kotta AS, Kelling AS, Corleto KA, Sun Y, Giles ED. Ghrelin and Cancer: Examining the Roles of the Ghrelin Axis in Tumor Growth and Progression. Biomolecules. 2022; 12(4):483. https://doi.org/10.3390/biom12040483
Chicago/Turabian StyleKotta, Anuhya S., Abigail S. Kelling, Karen A. Corleto, Yuxiang Sun, and Erin D. Giles. 2022. "Ghrelin and Cancer: Examining the Roles of the Ghrelin Axis in Tumor Growth and Progression" Biomolecules 12, no. 4: 483. https://doi.org/10.3390/biom12040483